
Seasonal Changes in the Nutritional Composition of Agarophyton vermiculophyllum (Rhodophyta, Gracilariales) from the Center of Portugal

 ,
, 
 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling Site, Collection, and Storage
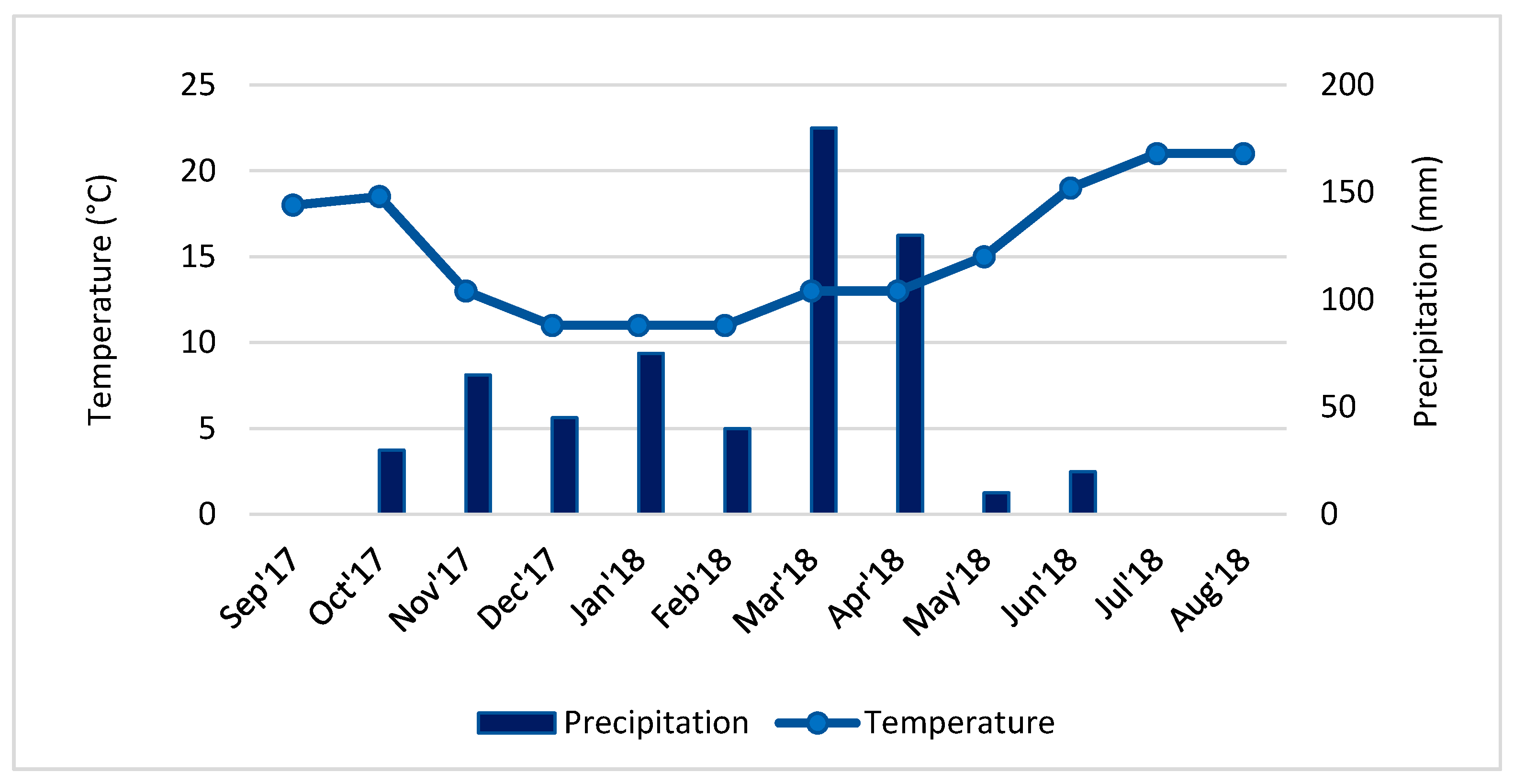
2.2. Seasonal Climatic Conditions
2.3. Estimation of Proximate Composition
2.3.1. Determination of Moisture and Ash Content
2.3.2. Total Dietary Fiber Determination
2.3.3. Total Protein Content
2.3.4. Total Lipid Content
2.3.5. Total Carbohydrates
2.3.6. Agar Extraction and Quantification
2.3.7. Determination of Ascorbic Acid Content
2.3.8. Amino Acid Profile
2.3.9. Fatty Acid Analysis
2.3.10. Statistical Analysis
3. Results
3.1. Seasonal Climatic Conditions
3.2. Proximate Composition of Agarophyton Vermiculophyllum
3.3. Fatty Acids Analysis
3.4. Amino Acid Profile
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bocanegra, A.; Bastida, S.; Benedí, J.; Ródenas, S.; Sánchez-Muniz, F.J. Characteristics and nutritional and cardiovascular-health properties of seaweeds. J. Med. Food 2009, 12, 236–258. [Google Scholar] [CrossRef] [PubMed]
- Holdt, S.L.; Kraan, S. Bioactive compounds in seaweed: Functional food applications and legislation. J. Appl. Phycol. 2011, 23, 543–597. [Google Scholar] [CrossRef]
- Angell, A.R.; Angell, S.F.; de Nys, R.; Paul, N.A. Seaweed as a protein source for mono-gastric livestock. Trends Food Sci. Technol. 2016, 54, 74–84. [Google Scholar] [CrossRef]
- Wells, M.L.; Potin, P.; Craigie, J.S.; Raven, J.A.; Merchant, S.S.; Helliwell, K.E.; Smith, A.G.; Camire, M.E.; Brawley, S.H. Algae as nutritional and functional food sources: Revisiting our understanding. J. Appl. Phycol. 2017, 29, 949–982. [Google Scholar] [CrossRef]
- Kazir, M.; Abuhassira, Y.; Robin, A.; Nahor, O.; Luo, J.; Israel, A.; Golberg, A.; Livney, Y.D. Extraction of proteins from two marine macroalgae, Ulva sp. and Gracilaria sp., for food application, and evaluating digestibility, amino acid composition and antioxidant properties of the protein concentrates. Food Hydrocoll. 2018, 87. [Google Scholar] [CrossRef]
- MaCartain, P.; Gill, C.I.R.; Brooks, M.; Campbell, R.; Rowland, I.R. Nutritional Value of Edible Seaweeds. Nutr. Rev. 2007, 65, 535–543. [Google Scholar] [CrossRef]
- Vieira, E.F.; Soares, C.; Machado, S.; Correia, M.; Ramalhosa, M.J.; Oliva-teles, M.T.; Paula Carvalho, A.; Domingues, V.F.; Antunes, F.; Oliveira, T.A.C.; et al. Seaweeds from the Portuguese coast as a source of proteinaceous material: Total and free amino acid composition profile. Food Chem. 2018, 269, 264–275. [Google Scholar] [CrossRef] [Green Version]
- Paiva, L.; Lima, E.; Neto, A.I.; Marcone, M.; Baptista, J. Nutritional and Functional Bioactivity Value of Selected Azorean Macroalgae: Ulva compressa, Ulva rigida, Gelidium microdon, and Pterocladiella capillacea. J. Food Sci. 2017, 82, 1757–1764. [Google Scholar] [CrossRef]
- Gamero-Vega, G.; Palacios-Palacios, M.; Quitral, V. Nutritional Composition and Bioactive Compounds of Red Seaweed: A Mini-Review. J. Food Nutr. Res. 2020, 8, 431–440. [Google Scholar] [CrossRef]
- Peñalver, R.; Lorenzo, J.M.; Ros, G.; Amarowicz, R.; Pateiro, M.; Nieto, G. Seaweeds as a functional ingredient for a healthy diet. Mar. Drugs 2020, 18, 301. [Google Scholar] [CrossRef]
- Pereira, L. A review of the nutrient composition of selected edible seaweeds. In Seaweed: Ecology, Nutrient Composition and Medicinal Uses; Pomin, V.H., Ed.; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2011; pp. 15–47. ISBN 978-1-61470-878-0. [Google Scholar]
- Gomez-Zavaglia, A.; Prieto Lage, M.A.; Jimenez-Lopez, C.; Mejuto, J.C.; Simal-Gandara, J. The potential of seaweeds as a source of functional ingredients of prebiotic and antioxidant value. Antioxidants 2019, 8, 406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hermund, D.B. Antioxidant Properties of Seaweed-Derived Substances. In Bioactive Seaweeds for Food Applications; Qin, Y., Ed.; Academic Press: Cambridge, MA, USA, 2018; pp. 201–221. [Google Scholar] [CrossRef]
- Barbalace, M.C.; Malaguti, M.; Giusti, L.; Lucacchini, A.; Hrelia, S.; Angeloni, C. Anti-inflammatory activities of marine algae in neurodegenerative diseases. Int. J. Mol. Sci. 2019, 20, 3061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, E.-J.; Ham, Y.; Lee, W.; Lee, N.; Hyun, C.-G. Anti-inflammatory effects of apo-9′-fucoxanthinone from the brown alga, Sargassum muticum. DARU J. Pharm. Sci. 2013, 21, 62. [Google Scholar] [CrossRef] [Green Version]
- De Jesus Raposo, M.F.; De Morais, A.M.B.; De Morais, R.M.S.C. Marine polysaccharides from algae with potential biomedical applications. Mar. Drugs 2015, 13, 2967–3028. [Google Scholar] [CrossRef] [PubMed]
- Telles, C.B.S.; Mendes-Aguiar, C.; Fidelis, G.P.; Frasson, A.P.; Pereira, W.O.; Scortecci, K.C.; Camara, R.B.G.; Nobre, L.T.D.B.; Costa, L.S.; Tasca, T.; et al. Immunomodulatory effects and antimicrobial activity of heterofucans from Sargassum filipendula. J. Appl. Phycol. 2018, 30, 569–578. [Google Scholar] [CrossRef]
- Schleder, D.D.; Peruch, L.G.B.; Poli, M.A.; Ferreira, T.H.; Silva, C.P.; Andreatta, E.R.; Hayashi, L.; do Nascimento Vieira, F. Effect of brown seaweeds on Pacific white shrimp growth performance, gut morphology, digestive enzymes activity and resistance to white spot virus. Aquaculture 2018, 495, 359–365. [Google Scholar] [CrossRef]
- Sakthivel, M.; Deivasigamani, B.; Rajasekar, T.; Kumaran, S.; Alagappan, K. Immunostimulatory Effects of Polysaccharide Compound from Seaweed Kappaphycus alvarezii on Asian seabass (Lates calcarifer) and it’s Resistance against Vibrio parahaemolyticus. J. Mar. Biol. Oceanogr. 2015, 4. [Google Scholar] [CrossRef] [Green Version]
- Lopes, G.; Sousa, C.; Silva, L.R.; Pinto, E.; Andrade, P.B.; Bernardo, J.; Mouga, T.; Valentão, P. Can phlorotannins purified extracts constitute a novel pharmacological alternative for microbial infections with associated inflammatory conditions? PLoS ONE 2012, 7, e31145. [Google Scholar] [CrossRef]
- Govindasamy, C.; Narayani, S.; Arulpriya, M.; Ruban, P.; Anantharaj, K.; Srinivasan, R. In vitro antimicrobial activities of seaweed extracts against human pathogens. J. Pharm. Res. 2011, 4, 2076–2077. [Google Scholar]
- Capillo, G.; Savoca, S.; Costa, R.; Sanfilippo, M.; Rizzo, C.; Giudice, A.L.; Albergamo, A.; Rando, R.; Bartolomeo, G.; Spanò, N.; et al. New Insights into the Culture Method and Antibacterial Potential of Gracilaria gracilis. Mar. Drugs 2018, 16, 492. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Y.V.; Walsh, N.A. Antioxidant and antiproliferative activities of extracts from a variety of edible seaweeds. Food Chem. Toxicol. 2006, 44, 1144–1150. [Google Scholar] [CrossRef]
- Milledge, J.J.; Nielsen, B.V.; Bailey, D. High-value products from macroalgae: The potential uses of the invasive brown seaweed, Sargassum muticum. Rev. Environ. Sci. Biotechnol. 2016, 15, 67–88. [Google Scholar] [CrossRef]
- Rodrigues, D.; Alves, C.; Horta, A.; Pinteus, S.; Silva, J.; Culioli, G.; Thomas, O.P.; Pedrosa, R. Antitumor and antimicrobial potential of bromoditerpenes isolated from the Red Alga, Sphaerococcus coronopifolius. Mar. Drugs 2015, 13, 713–726. [Google Scholar] [CrossRef] [Green Version]
- Francavilla, M.; Franchi, M.; Monteleone, M.; Caroppo, C. The red seaweed Gracilaria gracilis as a multi products source. Mar. Drugs 2013, 11, 3754–3776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soares, C.; Machado, S.; Vieira, E.F.; Morais, S.; Teles, M.T.; Correia, M.; Carvalho, A.; Domingues, V.F.; Ramalhosa, M.J.; Delerue-Matos, C.; et al. Seaweeds from the Portuguese coast: A potential food resource? In IOP Conference Series: Materials Science and Engineering; IOP Publishing Ltd: Bristol, UK, 2017; Volume 231, p. 012126. [Google Scholar] [CrossRef] [Green Version]
- Leandro, A.; Pacheco, D.; Cotas, J.; Marques, J.C.; Pereira, L.; Gonçalves, A.M.M. Seaweed’s bioactive candidate compounds to food industry and global food security. Life 2020, 10, 140. [Google Scholar] [CrossRef] [PubMed]
- Pacheco, D.; Araújo, G.S.; Cotas, J.; Gaspar, R.; Neto, J.M.; Pereira, L. Invasive Seaweeds in the Iberian Peninsula: A Contribution for Food Supply. Mar. Drugs 2020, 18, 560. [Google Scholar] [CrossRef] [PubMed]
- Fleurence, J. Seaweed proteins: Biochemical, nutritional aspects and potential uses. Trends Food Sci. Technol. 1999, 10, 25–28. [Google Scholar] [CrossRef]
- Fleurence, J.; Ele Morançais, M.; Dumay, J.; Decottignies, P.; Turpin, V.; Munier, M.; Garcia-Bueno, N.; Jaouen, P. What are the prospects for using seaweed in human nutrition and for marine animals raised through aquaculture? Trends Food Sci. Technol. 2012, 27, 57–61. [Google Scholar] [CrossRef]
- Lourenço, S.O.; Barbarino, E.; De-Paula, J.C.; Otávio, L.; Pereira, S.; Marquez, U.M.L. Amino acid composition, protein content and calculation of nitrogen-to-protein conversion factors for 19 tropical seaweeds. Phycol. Res. 2002, 50, 233–241. [Google Scholar] [CrossRef]
- Gressler, V.; Sumie Yokoya, N.; Fujii, T.; Colepicolo, P.; Filho, J.M.; Torres, R.P.; Pinto, E. Analytical Methods Lipid, fatty acid, protein, amino acid and ash contents in four Brazilian red algae species. Food Chem. 2010, 120, 585–590. [Google Scholar] [CrossRef]
- Schmid, M.; Kraft, L.G.K.; Van Der Loos, L.M.; Kraft, G.T.; Virtue, P.; Nichols, P.D.; Hurd, C.L. Southern Australian seaweeds: A promising resource for omega-3 fatty acids. Food Chem. 2018, 265, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Di Pasquale, M.G. The Essentials of Essential Fatty Acids. J. Diet. Suppl. 2009, 6, 143–161. [Google Scholar] [CrossRef] [PubMed]
- Dawczynski, C.; Schubert, R.; Jahreis, G. Amino acids, fatty acids, and dietary fibre in edible seaweed products. Food Chem. 2007, 103, 891–899. [Google Scholar] [CrossRef]
- Neveux, N.; Bolton, J.J.; Bruhn, A.; Ras, M. The Bioremediation Potential of Seaweeds: Recycling Nitrogen, Phosphorus, and Other Waste Products. In Blue Biotechnology: Production and Use of Marine Molecules, 1st ed.; La Barre, S., Bates, S.S., Eds.; Wiley: Weinheim, Germany, 2018; pp. 217–239. [Google Scholar] [CrossRef]
- Lozano Muñoz, I.; Díaz, N.F. Minerals in edible seaweed: Health benefits and food safety issues. Crit. Rev. Food Sci. Nutr. 2020, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Rajapakse, N. Nutritional and Digestive Health Benefits of Seaweed. Adv. Food Nutr. Res. 2011, 64, 17–28. [Google Scholar] [CrossRef]
- Cardoso, S.M.; Carvalho, L.G.; Silva, P.J.; Rodrigues, M.S.; Pereira, O.; Pereira, L. Bioproducts From Seaweeds: A Review With Special Focus On The Iberian Peninsula. Curr. Org. Chem. 2014, 18, 896–917. [Google Scholar] [CrossRef]
- Sousa, A.M.M.; Sereno, A.M.; Hilliou, L.; Gonçalves, M.P. Biodegradable Agar extracted from Gracilaria vermiculophylla: Film Properties and Application to Edible Coating. Mater. Sci. Forum 2010, 636–637, 739–744. [Google Scholar] [CrossRef]
- Bermejo, R.; MacMonagail, M.; Heesch, S.; Mendes, A.; Edwards, M.; Fenton, O.; Knöller, K.; Daly, E.; Morrison, L. The arrival of a red invasive seaweed to a nutrient over-enriched estuary increases the spatial extent of macroalgal blooms. Mar. Environ. Res. 2020, 158. [Google Scholar] [CrossRef]
- Rueness, J. Life history and molecular sequences of Gracilaria vermiculophylla (Gracilariales, Rhodophyta), a new introduction to European waters. Phycologia 2005, 44, 120–128. [Google Scholar] [CrossRef]
- Nejrup, L.B.; Pedersen, M.F. The effect of temporal variability in salinity on the invasive red alga Gracilaria vermiculophylla. Eur. J. Phycol. 2012, 47, 254–263. [Google Scholar] [CrossRef]
- Shields, R.J.; Lupatsch, I. Algae for Aquaculture and Animal Feeds. TATuP Z. Tech. Theor. Prax. 2012, 21, 23–37. [Google Scholar]
- Climate Portal Data. 2020. Available online: http://portaldoclima.pt/pt/ (accessed on 4 February 2021).
- AOAC. Official Methods of Analysis of AOAC International, 20th ed.; AOAC Int.: Gaithersburg, MD, USA, 2016. [Google Scholar]
- Folch, J.; Lees, M.; Sloane Stanley, G. A Simple Method for the Isolation and Purification of Total Lipides from Animal Tissues. J. Biol. Chem. 1957, 55, 999–1033. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K. Colorimetric Method for Determination of Sugars and related Substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Lepage, G.; Roy, C.C. Direct transesterification of all classes of lipids in one-step reactions. J. Lipid Res. 1986, 27, 114–120. [Google Scholar] [CrossRef]
- Simopoulos, A.P. The importance of the ratio of omega-6/omega-3 essential fatty acids. Biomed. Pharmacother. 2002, 56, 365–379. [Google Scholar] [CrossRef]
- Mota, C.; Santos, M.; Mauro, R.; Samman, N.; Matos, A.S.; Torres, D.; Castanheira, I. Protein content and amino acids profile of pseudocereals. Food Chem. 2016, 193, 55–61. [Google Scholar] [CrossRef] [PubMed]
- U.S. Department of Agriculture, Agricultural Research Service. Food Data Central, Nutrient Data Laboratory, Beltsville Human Nutrition Research Center, ARS, USDA. 2019. Available online: https://fdc.nal.usda.gov/fdc-app.html#/food-details/169282/nutrients (accessed on 20 January 2021).
- WHO; FAO; UNU. Protein and Amino Acid Requirements in Human Nutrition: Report of a Joint WHO/FAO/UNU Expert Consultation; World Health Organization: Geneva, Switzerland, 2007; ISBN 9789241209359/9241209356. [Google Scholar]
- Cherry, P.; O’Hara, C.; Magee, P.J.; Mcsorley, E.M.; Allsopp, P.J. Risks and benefits of consuming edible seaweeds. Nutr. Rev. 2019, 77, 307–329. [Google Scholar] [CrossRef] [Green Version]
- Samanta, P.; Jang, S.; Shin, S.; Kim, J.K. Effects of pH on growth and biochemical responses in Agarophyton vermiculophyllum under different temperature conditions. J. Appl. Phycol. 2020, 32, 499–509. [Google Scholar] [CrossRef]
- Rasyid, A.; Ardiansyah, A.; Pangestuti, R. Nutrient Composition of Dried Seaweed Gracilaria gracilis. Indones. J. Mar. Sci. 2019, 24, 1. [Google Scholar] [CrossRef]
- Ben Said, R.; Mensi, F.; Majdoub, H.; Ben Said, A.; Ben Said, B.; Bouraoui, A. Effects of depth and initial fragment weights of Gracilaria gracilis on the growth, agar yield, quality, and biochemical composition. J. Appl. Phycol. 2018, 30, 2499–2512. [Google Scholar] [CrossRef]
- Madden, M.; Mitra, M.; Ruby, D. Seasonality of selected nutritional constituents of edible Delmarva Seaweeds. J. Phycol. 2012, 48, 1289–1298. [Google Scholar] [CrossRef]
- Tabarsa, M.; Rezaei, M.; Ramezanpour, Z.; Waaland, J.R. Chemical compositions of the marine algae Gracilaria salicornia (Rhodophyta) and Ulva lactuca (Chlorophyta) as a potential food source. J. Sci. Food Agric. 2012, 92, 2500–2506. [Google Scholar] [CrossRef] [PubMed]
- Morales, C.; Schwartz, M.; Sepúlveda, M.; Quitral, V. Composición química y propiedades tecnológicas de alga roja, Agarophyton chilensis (ex Gracilaria chilensis). Rev. Cienc. Tecnol. 2019, 31, 59–67. [Google Scholar]
- Nazni, P.; Deepa, S. Minerals and Heavy metal present in the selected red seaweeds of south coast region of Tamilnadu. Int. J. Res. Biol. Sci. 2015, 5, 45–49. [Google Scholar]
- Debbarma, J.; Rao, B.M.; Murthy, L.N.; Mathew, S.; Venkateshwarlu, G.; Ravishankar, C.N. Nutritional profiling of the edible seaweeds Gracilaria edulis, Ulva lactuca and Sargassum sp. Indian J. Fish. 2016, 63, 81–87. [Google Scholar] [CrossRef] [Green Version]
- National Institute of Health. Iron—Fact Sheet for Health Professionals. Available online: https://ods.od.nih.gov/factsheets/Iron-HealthProfessional/ (accessed on 20 January 2021).
- NHS. Vitamins and Minerals. Available online: https://www.nhs.uk/conditions/vitamins-and-minerals/ (accessed on 20 January 2021).
- Torres, P.; Santos, J.P.; Chow, F.; dos Santos, D.Y.A.C. A comprehensive review of traditional uses, bioactivity potential, and chemical diversity of the genus Gracilaria (Gracilariales, Rhodophyta). Algal Res. 2019, 37, 288–306. [Google Scholar] [CrossRef]
- Sakthivel, R.; Devi, K.P. Evaluation of physicochemical properties, proximate and nutritional composition of Gracilaria edulis collected from Palk Bay. Food Chem. 2015, 174, 68–74. [Google Scholar] [CrossRef]
- Raposo, M.F.D.J.; De Morais, A.M.M.B.; De Morais, R.M.S.C. Emergent sources of prebiotics: Seaweeds and microalgae. Mar. Drugs 2016, 14, 27. [Google Scholar] [CrossRef]
- Martín, L.A.; Rodríguez, M.C.; Matulewicz, M.C.; Fissore, E.N.; Gerschenson, L.N.; Leonardi, P.I. Seasonal variation in agar composition and properties from Gracilaria gracilis (Gracilariales, Rhodophyta) of the Patagonian coast of Argentina. Phycol. Res. 2013, 61, 163–171. [Google Scholar] [CrossRef]
- Denis, C.; Morançais, M.; Li, M.; Deniaud, E.; Gaudin, P.; Wielgosz-Collin, G.; Barnathan, G.; Jaouen, P.; Fleurence, J. Study of the chemical composition of edible red macroalgae Grateloupia turuturu from Brittany (France). Food Chem. 2010, 119, 913–917. [Google Scholar] [CrossRef]
- Marinho-Soriano, E.; Fonseca, P.C.; Carneiro, M.A.A.; Moreira, W.S.C. Seasonal variation in the chemical composition of two tropical seaweeds. Bioresour. Technol. 2006, 97, 2402–2406. [Google Scholar] [CrossRef] [PubMed]
- Rajauria, G.; Cornish, L.; Ometto, F.; Msuya, F.E.; Villa, R. Identification and selection of algae for food, feed, and fuel applications. Seaweed Sustain. Food Non Food Appl. 2015, 315–345. [Google Scholar] [CrossRef]
- Rodrigues, D.; Freitas, A.C.; Pereira, L.; Rocha-Santos, T.A.P.; Vasconcelos, M.W.; Roriz, M.; Rodríguez-Alcalá, L.M.; Gomes, A.M.P.; Duarte, A.C. Chemical composition of red, brown and green macroalgae from Buarcos bay in Central West Coast of Portugal. Food Chem. 2015, 183, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Gorissen, S.H.M.; Crombag, J.J.R.; Senden, J.M.G.; Waterval, W.A.H.; Bierau, J.; Verdijk, L.B.; van Loon, L.J.C. Protein content and amino acid composition of commercially available plant-based protein isolates. Amino Acids 2018, 50, 1685–1695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parjikolaei, B.R.; Bruhn, A.; Eybye, K.L.; Larsen, M.M.; Rasmussen, M.B.; Christensen, K.V.; Fretté, X.C. Valuable Biomolecules from Nine North Atlantic Red Macroalgae: Amino Acids, Fatty Acids, Carotenoids, Minerals and Metals. Nat. Resour. 2016, 7, 157–183. [Google Scholar] [CrossRef] [Green Version]
- Syad, A.N.; Shunmugiah, K.P.; Kasi, P.D. Seaweeds as nutritional supplements: Analysis of nutritional profile, physicochemical properties and proximate composition of G. acerosa and S. wightii. Biomed. Prev. Nutr. 2013, 3, 139–144. [Google Scholar] [CrossRef]
- Roleda, M.Y.; Hurd, C.L. Seaweed nutrient physiology: Application of concepts to aquaculture and bioremediation. Phycologia 2019, 58, 552–562. [Google Scholar] [CrossRef] [Green Version]
- Hurd, C.L. Shaken and stirred: The fundamental role of water motion in resource acquisition and seaweed productivity. Perspect. Phycol. 2017, 4, 73–81. [Google Scholar] [CrossRef]
- Misurcová, L. Chemical Composition of Seaweeds. In Handbook Marine Macroalgae: Biotechnology Applied Phycology; John Wiley & Sons, Ltd.: Chichester, UK, 2011. [Google Scholar] [CrossRef]
- Marrion, O.; Schwertz, A.; Fleurence, J.; Guéant, J.L.; Villaume, C. Improvement of the digestibility of the proteins of the red alga Palmaria palmata by physical processes and fermentation. Nahr. Food 2003, 47, 339–344. [Google Scholar] [CrossRef]
- Brown, M.R.; Jeffrey, S.W. Biochemical composition of microalgae from the green algal classes Chlorophyceae and Prasinophyceae. 1. Amino acids, sugars and pigments. J. Exp. Mar. Biol. Ecol. 1992, 161, 91–113. [Google Scholar] [CrossRef]
- Rosemary, T.; Arulkumar, A.; Paramasivam, S.; Mondragon-Portocarrero, A.; Miranda, J.; Rosemary, T.; Arulkumar, A.; Paramasivam, S.; Mondragon-Portocarrero, A.; Miranda, J.M. Biochemical, Micronutrient and Physicochemical Properties of the Dried Red Seaweeds Gracilaria edulis and Gracilaria corticata. Molecules 2019, 24, 2225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morais, T.; Inácio, A.; Coutinho, T.; Ministro, M.; Cotas, J.; Pereira, L.; Bahcevandziev, K. Seaweed potential in the animal feed: A review. J. Mar. Sci. Eng. 2020, 8, 559. [Google Scholar] [CrossRef]
- Candela, C.G.; López, L.B.; Kohen, V.L. Importance of a balanced omega 6/omega 3 ratio for the maintenance of health. Nutritional recommendations. Nutr. Hosp. 2011, 26, 323–329. [Google Scholar] [CrossRef]
- Khotimchenko, S.; Vaskovsky, V.; Przhemenetskaya, V. Distribution of eicosapentaenoic and arachidonic acids in different species of Gracilaria. Phytochemistry 1991, 30, 207–209. [Google Scholar] [CrossRef]
- Hafting, J.T.; Craigie, J.S.; Stengel, D.B.; Loureiro, R.R.; Buschmann, A.H.; Yarish, C.; Edwards, M.D.; Critchley, A.T. Prospects, and challenges for industrial production of seaweed bioactives. J. Phycol. 2015, 51, 821–837. [Google Scholar] [CrossRef]
- Paiva, L.; Lima, E.; Neto, A.I.; Baptista, J. Seasonal Variability of the Biochemical Composition and Antioxidant Properties of Fucus spiralis at Two Azorean Islands. Mar. Drugs 2018, 16, 248. [Google Scholar] [CrossRef] [Green Version]
- WHO. Interim Summary of Conclusions and Dietary Recommendations on Total Fat and Fatty Acids. In Proceedings of the Joint FAO/WHO Expert Consultation on Fats and Fatty Acids in Human Nutrition, Geneva, Switzerland, 10–14 November 2008. [Google Scholar]
- Norziah, M.H.; Ching, C.Y. Nutritional Composition of Edible Seaweed Gracilaria changgi. Food Chem. 2000, 68, 69–76. [Google Scholar] [CrossRef]
- Yildiz, G.; Dere, E.; Dere, Ş. Comparison of the antioxidative components of some marine macroalgae from Turkey. Pak. J. Bot. 2014, 46, 753–757. [Google Scholar]
- Ortiz, J.; Romero, N.; Robert, P.; Araya, J.; Lopez-Hernández, J.; Bozzo, C.; Navarrete, E.; Osorio, A.; Rios, A. Dietary fiber, amino acid, fatty acid and tocopherol contents of the edible seaweeds Ulva lactuca and Durvillaea antarctica. Food Chem. 2006, 99, 98–104. [Google Scholar] [CrossRef]
- Gioele, C.; Marilena, S.; Valbona, A.; Nunziacarla, S.; Andrea, S.; Antonio, M. Gracilaria gracilis, Source of Agar: A Short Review. Curr. Org. Chem. 2017, 21, 380–386. [Google Scholar] [CrossRef]
- Pangestuti, R.; Kim, S.K. Seaweed proteins, peptides, and amino acids. In Seaweed Sustainability: Food and Non-Food Applications; Academic Press: Waltham, MA, USA, 2015; ISBN 9780124199583. [Google Scholar] [CrossRef]
- Marinho-Soriano, E. Agar polysaccharides from Gracilaria species (Rhodophyta, Gracilariaceae). J. Biotechnol. 2001, 89, 81–84. [Google Scholar] [CrossRef]


{kind=link}
| 17 September | 17 October | 17 November | 17 December | 18 January | |
| Collecting site | FF | LO | FF | LO | FF |
| Moisture (% fw) | 74.1 ± 0.5 ab | 76.0 ± 0.2 b | 74.1 ± 1.4 ab | 71.3 ± 0.7 a | 74.7 ± 1.4 b |
| Ash (%) | 24.5 ± 1.3 | 19.6 ± 2.7 | 19.6 ± 2.1 | 19.9 ± 1.5 | 19.6 ± 16 |
| Calcium (mg 100 g−1 dw) | 138.5 | 190.7 | n.a. | 157.9 | 129.8 |
| Iron (mg 100 g−1 dw) | 148.6 | 80.8 | n.a. | 107.2 | 221.3 |
| Zinc (mg 100 g−1 dw) | 2.1 | 2.1 | n.a | 2.0 | 2.6 |
| Fiber (%) | 25.15 | 28.26 | n.a | 29.76 | 26.37 |
| Carbohydrates (%) | 46.7 ± 3.2 | 48.9 ± 2.6 | 45.9 ± 5.6 | 45.2 ± 8.1 | 40.2 ± 5.3 |
| Protein (%) | 21.6 ± 2.1 e | 15.5 ± 0.5 bcd | 19.0 ± 1.2 cde | 18.9 ± 1.4 de | 20.8 ± 0.6 de |
| Lipids (%) | 1.8 ± 0.1 bcde | 1.6 ± 0.1 bc | 1.9 ± 0.1 cde | 2.0 ± 0.1 de | 1.2 ± 0.1 bcde |
| Vitamin C (mg g−1 dw) | 0.15 ± 0.0 a | 0.15 ± 0.03 a | 0.17 ± 0.2 ab | 0.14 ± 0.1 a | 0.25 ± 0.2 bc |
| Agar (%) | 21.5 ± 2.1 c | 18.3 ± 1.5 bc | 18.3 ± 2.9 bc | 8.8 ± 0.9 a | 9.9 ± 1.0 a |
| 18 March | 18 April | 18 May | 18 July | 18 August | |
| Collecting site | FF | LO | FF | FF | LO |
| Moisture (% fw) | 75.2 ± 0.6 b | 74.2 ± 0.6 b | 76.9 ± 0.9 ab | 77.6 ± 0.5 b | 75.9 ± 0.4 b |
| Ash (%) | 22.5 ± 1.0 | 23.7 ± 1.1 | 23.9 ± 0.7 | 21.7 ± 0.5 | 21.6 ± 0.5 |
| Calcium (mg 100 g−1 dw) | 154.3 | 145.9 | n.a. | 188.92 | 138.1 |
| Iron (mg 100 g−1 dw) | 69.1 | 145.1 | n.a. | 94.7 | 63.9 |
| Zinc (mg 100 g−1 dw) | 1.9 | 2.4 | n.a. | 2.5 | 1.7 |
| Fiber (%) | 29.59 | 29.89 | n.a. | 26.35 | 32.75 |
| Carbohydrates (%) | 37.6 ± 2.1 | 47.0 ± 6.8 | 46.4 ± 2.7 | 45.1 ± 5.6 | 48.6 ± 0.4 |
| Protein (%) | 21.2 ± 0.2 e | 17.1 ± 2.0 cde | 15.2 ± 0.3 bc | 14.7 ± 0.1 b | 6.2 ± 0.3 a |
| Lipids (%) | 1.7 ± 0.0 bcd | 2.0 ± 0.1 e | 1.5 ± 0.1 ab | 1.8 ± 0.1 bcde | 1.2 ± 0.1 a |
| Vitamin C (mg g−1 dw) | 0.31 ± 0.04 c | 0.14 ± 0.01 a | 0.17 ± 0.01 ab | 0.18 ± 0.02 ab | 0.22 ± 0.04 abc |
| Agar (%) | 16.3 ± 1.4 abc | 17.7 ± 3.8 bc | 9.2 ± 1.7 a | 11.2 ± 0.2 ab | 13.6 ± 2.5 ab |
| Fatty Acids (%TFA) | 17 September | 17 October | 17 November | 17 December | 18 January | 18 March | 18 April | 18 May | 18 July | 18 August |
| 14:0 [myristic acid] | n.d. | 0.31 ± 0.02 | 0.39 ± 0.02 | 0.37 ± 0.25 | 0.34 ± 0.07 | 0.27 ± 0.01 | 0.22 ± 0.01 | 0.29 ± 0.04 | 0.42 ± 0.06 | 0.26 ± 0.00 |
| 15:0 [pentadecylic acid] | 0.47 ± 0.15 | 0.70 ± 0.04 | 0.76 ± 0.08 | 0.74 ± 0.15 | 0.53 ± 0.17 | 0.33 ± 0.13 | 0.35 ± 0.04 | 0.51 ± 0.17 | 0.83 ± 0.61 | 0.61 ± 0.00 |
| 16:0 [palmitic acid] | 57.88 ± 5.14 | 62.95 ± 5.69 | 53.62 ± 4.12 | 50.43 ± 4.19 | 44.72 ± 9.06 | 32.35 ± 2.21 | 51.18 ± 5.19 | 39.35 ± 8.83 | 58.70 ± 5.08 | 39.75 ± 10.8 |
| 17:0 [margaric acid] | 0.30 ± 0.01 | n.d. | 0.25 ± 0.10 | n.d. | 0.19 ± 0.13 | 0.22 ± 0.11 | n.d. | 0.14 ± 0.02 | ||
| 20:0 [arachidic acid] | 1.13 ± 0.00 | n.d. | n.d. | 0.06 ± 0.00 | n.d. | |||||
| 21:0 [heneicosylic acid] | n.d. | n.d. | n.d. | 0.10 ± 0.05 | 0.11 ± 0.00 | 0.07 ± 0.03 | ||||
| 23:0 [tricosylic acid] | n.d. | 0.06 ± 0.00 | n.d. | n.d. | ||||||
| Sum SFA (%) | 59.78 | 64.02 | 55.02 | 51.70 | 45.89 | 33.24 | 51.89 | 40.29 | 60.91 | 40.81 |
| 14:1 [myristoleic acid] | 5.76 ± 0.77 | 6.16 ± 0.43 | 1.97 ± 0.08 | 4.07 ± 0.94 | 3.81 ± 0.51 | 2.85 ± 0.95 | 4.87 ± 0.62 | 4.61 ± 0.63 | 5.38 ± 1.80 | 4.45 ± 1.60 |
| 15:1 [pentadecenoic acid] | n.d. | 0.27 ± 0.06 | 0.27 ± 0.04 | 0.60 ± 0.18 | 0.38 ± 0.14 | 0.50 ± 0.48 | 0.20 ± 0.12 | 0.37 ± 0.12 | 0.61 ± 0.64 | 0.43 ± 0.00 |
| 16:1 n-7 [palmitoleic acid] | 0.63 ± 0.15 | n.d. | n.d. | 0.21 ± 0.01 | 0.76 ± 0.25 | 0.43 ± 0.22 | 0.43 ± 0.25 | 0.44 ± 0.09 | 1.17 ± 0.27 | 0.47 ± 0.08 |
| 17:1 [heptadecenoic acid] | 0.46 ± 0.03 | 0.48 ± 0.09 | 0.53 ± 0.16 | 0.52 ± 0.11 | 0.43 ± 0.10 | 0.33 ± 0.12 | 0.41 ± 0.04 | 0.45 ± 0.16 | 0.39 ± 0.05 | 0.24 ± 0.03 |
| 18:1 n-9 [oleic acid) | 7.67 ± 2.84 | 6.81 ± 0.61 | 7.40 ± 2.19 | 4.42 ± 1.02 | 4.46 ± 1.08 | 3.78 ± 0.18 | 4.82 ± 1.16 | 3.22 ± 2.42 | 3.14 ± 0.82 | 1.33 ± 0.13 |
| 20:1 n-9 [eicosenoic acid] | n.d. | n.d. | 0.08 ± 0.00 | 0.1 ± 0.04 | 0.11 ± 0.00 | 0.11 ± 0.04 | 0.05 ± 0.00 | n.d. | 0.55 ± 0.00 | 0.15 ± 0.04 |
| 24:1 n-9 [nervonic acid] | n.d. | 0.4 ± 0.02 | n.d. | 1.21 ± 1.21 | 1.22 ± 1.08 | 0.40 ± 0.20 | 1.06 ± 0.10 | 0.69 ± 0.29 | 1.15 ± 0.54 | 0.46 ± 0.22 |
| Sum MUFAs (%) | 14.52 | 14.12 | 10.25 | 11.13 | 11.17 | 8.40 | 11.84 | 9.78 | 12.39 | 7.53 |
| 18:2 n-6 cis [linoleic acid, LA] | 4.53 ± 0.32 | 8.27 ± 2.51 | 1.85 ± 1.12 | 2.67 ± 1.21 | 1.32 ± 0.96 | 1.34 ± 0.37 | 2.87 ± 0.72 | 4.93 ± 2.12 | 5.95 ± 4.20 | 5.14 ± 4.75 |
| 18:2 n-6 trans [linolelaidic acid] | 0.36 ± 0.08 | 0.37 ± 0.04 | 0.30 ± 0.07 | 0.57 ± 0.09 | 0.63 ± 0.11 | 0.52 ± 0.00 | 1.02 ± 0.03 | 1.17 ± 0.74 | 1.78 ± 0.13 | 1.25 ± 0.47 |
| 18:3 n-6 [γ-linolenic acid] | n.d. | n.d. | n.d. | n.d. | n.d. | 0.60 ± 0.00 | 0.56 ± 0.10 | 0.39 ± 0.11 | 0.99 ± 0.11 | 0.95 ± 0.15 |
| 18:3 n-3 [α-linolenic acid] | n.d. | n.d. | 0.55 ± 0.36 | 0.47 ± 0.06 | 0.41 ± 0.01 | 0.33 ± 0.04 | 0.32 ± 0.00 | 0.80 ± 0.58 | 0.12 ± 0.00 | 0.46 ± 0.07 |
| 20:2 [eicosatrienoic acid] | 1.40 ± 0.41 | n.d. | n.d. | n.d. | n.d. | 0.12 ± 0.00 | 0.03 ± 0.00 | n.d. | n.d. | n.d. |
| 20:3 n-3 [eicosatrienoic acid] 20:3 n-6 [eicosatrienoic acid] 20:4 n-6 [arachidonic acid, AA] | n.d. | 1.56 ± 0.56 | 1.34 ± 0.58 | 1.65 ± 0.08 | 0.97 ± 0.00 | 0.60 ± 0.88 | 2.67 ± 0.22 | 1.90 ± 0.13 | 1.45 ± 0.07 | 2.72 ± 0.39 |
| n.d. | n.d. | n.d. | 0.13 ± 0.03 | 1.69 ± 0.08 | 0.95 ± 0.78 | n.d. | n.d. | 0.16 ± 0.00 | n.d. | |
| 21.83 ± 6.31 | 14.83 ± 5.55 | 27.16 ± 5.16 | 29.74 ± 4.81 | 37.9 ± 10.26 | 52.88 ± 3.83 | 27.88 ± 5.94 | 39.83 ± 8.99 | 15.79 ± 0.76 | 42.14 ± 15.51 | |
| 20:5 n-3 [eicosapentaenoic acid, EPA] 22:6 n-3 [Docosahexaenoic acid DHA] | n.d. | 0.28 ± 0.00 | n.d. | 0.13 ± 0.01 | 0.21 ± 0.00 | 0.18 ± 0.02 | 0.11 ± 0.00 | 0.26 ± 0.07 | n.d. | n.d. |
| n.d. | 0.51 ± 0.00 | n.d. | 4.41 ± 0.00 | n.d. | n.d. | n.d. | n.d. | n.d. | 0.36 ± 0.39 | |
| Sum PUFAs (%) | 28.12 | 25.82 | 31.20 | 39.77 | 43.13 | 57.52 | 35.46 | 49.28 | 26.24 | 53.02 |
| Nutritional Index | ||||||||||
| Σ n-3 | 2.35 | 1.89 | 6.66 | 1.59 | 1.11 | 3.10 | 2.96 | 1.57 | 3.54 | |
| Σ n-6 | 26.72 | 23.98 | 29.31 | 33.11 | 41.54 | 56.41 | 32.36 | 46.32 | 24.67 | 49.48 |
| ratio n-6/n-3 | 10.20 | 15.51 | 4.97 | 26.13 | 50.82 | 10.44 | 15.65 | 15.71 | 13.98 | |
| ratio h/H | 0.74 | 0.63 | 0.77 | 1.00 | 1.21 | 2.02 | 0.92 | 1.49 | 0.65 | 1.51 |
| A. vermiculophyllum (g 100 g−1 fw) | Quinoa [52] (g 100 g−1 fw) | Soybean [53] (g 100 g−1 fw) | FAO/WHO [54] (g/kg/day) | |
|---|---|---|---|---|
| Essential Amino Acids | ||||
| Thr | 0.178 ± 0.023 | 0.426 ± 0.034 | 0.516 | 0.015 |
| Met (1) | 0.085 ± 0.014 | 0.257 ± 0.014 | 0.157 | 0.015(1 + 3) |
| Phe (2) | 0.169 ± 0.022 | 0.615 ± 0.074 | 0.586 | 0.025(2 + 4) |
| His | 0.053 ± 0.006 | 0.475 ± 0.048 | 0.348 | 0.010 |
| Lys | 0.155 ± 0.023 | 0.481 ± 0.046 | 0.775 | 0.030 |
| Val | 0.154 ± 0.019 | 0.458 ± 0.046 | 0.576 | 0.026 |
| Ile | 0.126 ± 0.017 | 0.372 ± 0.031 | 0.570 | 0.020 |
| Leu | 0.271 ± 0.034 | 0.713 ± 0.039 | 0.926 | 0.039 |
| Σ EAA | 1.190 ± 0.159 | |||
| Non-essential Amino Acids | ||||
| Ser | 0.211 ± 0.026 | 0.594 ± 0.072 | 0.721 | - |
| Gly | 0.207 ± 0.031 | 0.648 ± 0.048 | 0.539 | - |
| Asx | 0.465 ± 0.060 | 0.924 ± 0.049 | 1.508 | - |
| Glx | 0.635 ± 0.122 | 1.888 ± 0.079 | 2.433 | - |
| Pro | 0.168 ± 0.023 | 0.433 ± 0.034 | 0.607 | - |
| Cys (3) | 0.007 ± 0.002 | 0.165 ± 0.052 | 0.118 | - |
| Ala | 0.278 ± 0.035 | 0.466 ± 0.035 | 0.582 | - |
| Tyr (4) | 0.134 ± 0.018 | 0.718 ± 0.058 | 0.464 | - |
| Arg | 0.227 ± 0.040 | 1.101 ± 0.095 | 1.042 | - |
| Σ NEAA | 2.332 ± 0.357 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Afonso, C.; Correia, A.P.; Freitas, M.V.; Baptista, T.; Neves, M.; Mouga, T. Seasonal Changes in the Nutritional Composition of Agarophyton vermiculophyllum (Rhodophyta, Gracilariales) from the Center of Portugal. Foods 2021, 10, 1145. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10051145
Afonso C, Correia AP, Freitas MV, Baptista T, Neves M, Mouga T. Seasonal Changes in the Nutritional Composition of Agarophyton vermiculophyllum (Rhodophyta, Gracilariales) from the Center of Portugal. Foods. 2021; 10(5):1145. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10051145
Chicago/Turabian StyleAfonso, Clélia, Ana Patrícia Correia, Marta V. Freitas, Teresa Baptista, Marta Neves, and Teresa Mouga. 2021. "Seasonal Changes in the Nutritional Composition of Agarophyton vermiculophyllum (Rhodophyta, Gracilariales) from the Center of Portugal" Foods 10, no. 5: 1145. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10051145