
Effects of CaCl2 Treatment Alleviates Chilling Injury of Loquat Fruit (Eribotrya japonica) by Modulating ROS Homeostasis


College of Food Science and Technology, Nanjing Agricultural University, Nanjing 210095, China
*
Author to whom correspondence should be addressed.
Foods 2021, 10(7), 1662; https://0-doi-org.brum.beds.ac.uk/10.3390/foods10071662
Submission received: 11 June 2021
/
Revised: 14 July 2021
/
Accepted: 15 July 2021
/
Published: 19 July 2021
(This article belongs to the Special Issue Latest Advances in Preservation Technology for Fresh Fruit and Vegetables)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:The effects of calcium chloride (CaCl2) treatment on chilling injury (CI), reactive oxygen species (ROS) metabolism, and ascorbate-glutathione (AsA-GSH) cycle in loquat fruit at 1 °C storage for 35 d were investigated. The results indicated that CaCl2 treatment remarkably suppressed the increase in browning index and firmness as well as the decrease in extractable juice rate. CaCl2 treatment also decreased the production of superoxide radical (), hydrogen peroxide (H2O2) content, but increased the 1,1-diphenyl-2-picrylhydrazyl (DPPH), hydroxyl radical () scavenging ability, the activities of superoxide dismutase (SOD), catalase (CAT), and their gene expressions. Moreover, compared to the control loquat fruit, CaCl2-treated fruit maintained higher contents of AsA, GSH, higher levels of activities of ascorbate peroxidase (APX), glutathione reductase (GR), dehydroascorbate reductase (DHAR), and monodehydroascorbate reductase (MDHAR) and expressions of EjAPX, EjGR, EjMDHAR, and EjDHAR, but exhibited lower glutathione disulfide (GSSG) content. These results suggested that CaCl2 treatment alleviated CI in loquat fruit through enhancing antioxidant enzymes activities and AsA-GSH cycle system to quench ROS.
1. Introduction
Loquat (Eriobotrya japonica Lindl.), as a type of non-climacteric fruit, presents a short postharvest life at ambient temperature because of its physiological deterioration and microbial decay. Refrigeration is widely adopted to retain quality and prolong the shelf life of loquat fruit. Nevertheless, loquat fruit is vulnerable to chilling injury (CI), displaying symptoms including enhanced fruit firmness, internal browning, and reduced extractable juice rate after long-term low temperature storage [1,2], which severely influences the texture and commercial value of the fruit, and eventually reduces consumer acceptance. Therefore, the exploration into the mechanism of CI in loquat fruit is of great significance.
It is well-established that the occurrence of chilling injury in higher plants is closely associated with oxidative stress resulted from excessive reactive oxygen species (ROS) such as superoxide anion (), hydrogen peroxide (H2O2), and hydroxyl radical () [3]. This large quantity of ROS accumulation alter the membrane organization and cause or exacerbate lipid peroxidation, leading to damage to the cell system [4]. To protect against oxidative damage and maintain homeostasis of ROS, plants have evolved a complicated antioxidant system including enzymatic and non-enzymatic antioxidant components. Superoxide dismutase (SOD), catalase (CAT) and peroxidase (POD) are primary antioxidant enzymes, while ascorbate (AsA) and glutathione (GSH) are non-enzymatic antioxidants [5]. The ascorbate–glutathione (AsA-GSH) cycle, which mainly consists of ascorbate peroxidase (APX), glutathione reductase (GR), dehydroascorbate reductase (DHAR), and monodehydroascorbate reductase (MDHAR), plays a critical role in ROS elimination and preventing oxidative damage [6]. It has been reported that increasing the activities of antioxidant enzymes (SOD, CAT, APX, GR, MDHAR, and DHAR) to eliminate excessive ROS is beneficial for alleviating the occurrence of CI in many fruit, such as peach [7], litchi [8], and bell pepper [9]. Furthermore, previous studies have confirmed that the increased antioxidant enzymes activities (SOD, CAT, and POD) and AsA-GSH cycle and mitigated peroxidation of membrane lipids might contribute to the enhancement of chilling tolerance in loquat fruit [10,11]; nevertheless, their regulation mechanism at a molecular level remains unknown.
Calcium ion (Ca2+), as an essential nutrient for fruit, plays a crucial role in building the structure of cell wall and cellular membranes [12]. Moreover, Ca2+ is also a crucial second messenger in plant signal transduction involving in physiological processes and the responses to various stresses [13]. Under cold stress, induced transient elevations of cytosolic Ca2+ are sensed by different calcium binding proteins, consequently initiating various physiological responses in the cell [14]. Dong et al. [15] reported that overexpression of the MdCPK1a gene increased tobacco chilling tolerance by inducing the expression of SOD, CAT, APX and scavenging ROS accumulation. Li et al. [16] suggested that postharvest Ca2+ application significantly increased Ca2+ and calmodulin content, and concomitantly, endogenous GABA content in ‘Nanguo’ pear fruit, which in turn delayed fruit browning after low-temperature storage. In addition, recent studies have reported that calcium chloride (CaCl2) application can trigger antioxidant system activity and maintain ROS homeostasis to increase cold tolerance of postharvest vegetables and fruits. For instance, CaCl2 application effectively alleviated peel browning caused by chilling injury of pear fruit on account of the inhibition of membrane lipid peroxidation and a higher activity and expression of SOD [17]. Wei and Zhao [18] also found that CaCl2 treatment could alleviate chilling injury symptoms in winter jujube fruit by promoting the SOD, CAT, and POD activities to scavenge ROS. In green peppers, CaCl2 treatment significantly suppressed ROS levels by regulating activities of SOD, POD, CAT as well as promoting the AsA-GSH cycle and, in turn, enhanced chilling tolerance [19]. For loquat fruit, a previous study demonstrated that CaCl2 treatment could significantly enhance the cold tolerance of loquat fruit through regulating energy metabolism and accumulating osmotic substances [20]. However, whether the inhibitory impact of calcium application is associated with the modulation of the ROS-scavenging system for the chilling injury of loquat fruit remains unclear.
Therefore, the impact of exogenous CaCl2 treatment on the ROS metabolism and AsA–GSH cycle system in loquat fruit were examined, which aimed to elucidate the antioxidant system triggered by Ca2+ treatment in alleviating oxidative damage during CI. The results will extend the mechanistic understanding of CaCl2-inhibited chilling injury in postharvest loquat fruit, and supply a scientific basis for the application of CaCl2 treatment as a useful technology to prolong the storage life of vegetables and fruits.
2. Materials and Methods
2.1. Materials and Treatments
Loquat fruit (Eriobotrya japonica L. cv. ‘Changhong’) of uniform maturity, size, and color were harvested from a commercial orchard in Fujian, China. Fruit with no mechanical damage or disease were chosen and then randomly divided into two groups (375 fruits in each group). One percent of CaCl2 solution was employed to soak loquat fruit for 10 min as CaCl2 treatment group, with the concentration chosen according to Li et al. [20]. Distilled water was used to immerse fruits for 10 min as control group. After treatment, all fruit were stored at 1 ± 1 °C with 90−95% relative humidity for 35 days. A total of 75 fruits from each group comprising three replicates were randomly sampled at 7 days intervals during storage. Fruit firmness was evaluated using 10 fruits, and 35 fruits were cut into small pieces, frozen using liquid nitrogen and stored at −80 °C for subsequent analysis. At each sampling day, another 30 fruits were transferred from 1 °C and kept at 20 °C for 3 days to stimulate shelf condition, and then internal browning index, firmness and extractable juice were measured. Three independent replicates were evaluated.
2.2. Measurement of Browning Index, Firmness, and Extractable Juice
The browning index was visually assayed using the reported method [20]. Browning index was scored based on a 5-grade scale (0, no visible symptoms; 1, <5% of browning; 2, 5–25% of browning; 3, 25–50% of browning; 4, >50% of browning). The result was acquired by the equation given below:
Fruit firmness was determined following the method of Zhang et al. [21]. Firmness from 10 loquat fruits selected randomly were assayed by a TA-XT2i texture analyzer (Stable Micro System Ltd., Surrey, UK), with a 5 mm diameter probe (test speed 1 mm s−1). Firmness measurement was operated at two opposite sides of fruit, and the result was expressed as N.
Extractable juice was evaluated based on the description of Cao et al. [10]. The unit of % was used to express extractable juice rate.
2.3. Measurement of , Generation Rate and H2O2 Content
The generation rate of and H2O2 content were assayed based on the description of Cao et al. [10]. The generation rate was acquired based on standard curve obtained from sodium nitrite and denoted as the unit of nmol g−1 min−1 on the basis of fresh weight (FW). The H2O2 content was acquired based on a standard curve and represented as μmol g−1 FW.
2.4. Measurement of 1,1-Diphenyl-2-Picrylhydrazyl (DPPH) and Radical Scavenging Capacity
Frozen tissue (2 g) was employed to determine DPPH and radical scavenging capacity according to Wang et al. [22].
For DPPH radical scavenging rate, the reaction mixture contained 0.1 mL of crude enzyme and 1.9 mL of 120 μmol L−1 DPPH. The result was acquired using the equation given below:
A0 and A1 indicate the absorbance of control and sample, respectively. Percentage was used for expressing DPPH radical scavenging rate.
For scavenging rate, the reaction mixture included 0.5 mL of crude enzyme, 1.5 mL of salicylic acid, 2 mL of water and 0.1 mL of 0.3% H2O2. Results were calculated by the following equation:
A0 and A1 refer to absorbance of the control and samples, respectively. The unit of % was regarded as radical scavenging ability.
2.5. Determination of SOD and CAT Activities
Determination of SOD and CAT enzymes activities were carried out based on the description of Zhang et al. [21]. One unit of SOD activity was defined as the amount of enzyme causing 50% inhibition of nitroblue tetrazolium (NBT) reduction. One unit of CAT was defined as the amount of enzyme that decomposed 1 μmol of H2O2 min−1. The unit of U g−1 was used to express SOD and CAT activities on the basis of fresh weight.
2.6. Measurement of Parameters Related to the AsA-GSH Cycle
Determination of AsA content was performed based on the description of Wang et al. [22], and the unit of AsA content was represented as mg g−1 FW based on the standard curve.
GSH and glutathione disulfide (GSSG) contents were assayed using assay kits (Solarbio, Beijing, China), according to manufacturer’s instructions. GSH and GSSG contents were represented as μg g−1 on the basis of fresh weight.
APX activity was performed following the procedures reported by Liu et al. [8]. The APX activity was acquired from the alteration in absorbance (290 nm) for 5 min promoted by the AsA after adding H2O2. One unit of APX activity was defined as the amount of enzyme that led to a 0.01 variation in absorbance every minute at 290 nm, and the APX activity was represented as U g−1 FW.
The activity of GR was determined through monitoring the variation in absorbance at 340 nm for 2 min due to oxidation of NADPH with GSSG [8], and the results were represented as U g−1 FW.
Frozen tissue (2 g) was used to measure DHAR and MDHAR activities according to the description of Cao et al. [10]. DHAR activity was acquired from the variation in absorbance at 265 nm and the result was represented as U g−1 FW. MDHAR activity was acquired through evaluating the reduction in NADH absorbance at 340 nm and represented as U g−1 FW.
2.7. Real-Time Quantitative PCR (RT-qPCR) Analysis
Total RNA was extracted from loquat fruit according to an improved CTAB method [23]. First-strand cDNA was synthesized using Hifair Ⅲ First Strand cDNA Synthesis Super Mix for qPCR (gDNA digester plus) (11141ES60, YEASEN, Shanghai, China). RT-qPCR analysis was conducted with a 7500 Fast Real-Time PCR System (Thermo Fisher Scientific, Waltham, MA, USA) by mixing the primers, Hieff qPCR SYBR Green Master Mix (11202ES08, YEASEN), cDNA template and RNase-free water in a total volume of 20 μL. EjACT was selected as the internal control to normalize the relative gene expression in each sample [24]. The relative expressions were normalized against the threshold cycle (Ct) value of the target gene and calculated with the 2–ΔΔCt method. All analyses were conducted in triplicate, and all primer sequences were shown in Supplementary Table S1.
2.8. Statistical Analysis
Experiments were conducted at least three biological replicates and all data were expressed as mean ± standard error (SE). The statistical analyses were performed using IBM SPSS Statistics 22 (SPSS Inc., Chicago, IL, USA). The independent samples T-test was used to compare the data of control and CaCl2 treatment groups. The value of p < 0.05 indicated significant difference.
3. Results
3.1. Effects of CaCl2 Treatment on Internal Browning Index, Fruit Firmness, Extractable Juice of Loquat Fruit
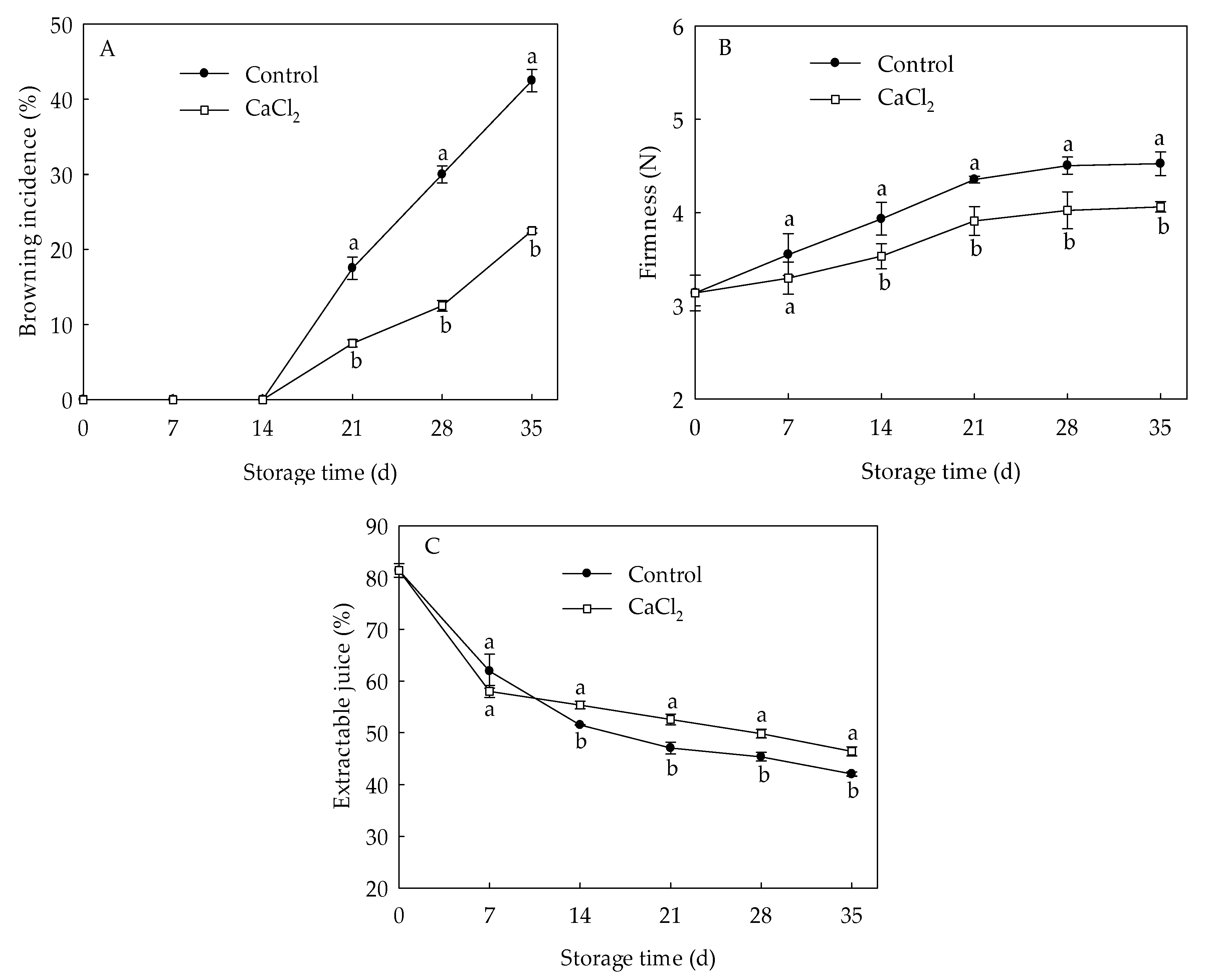
Internal browning in loquat fruit occurred on the 21st day of storage and increased sharply thereafter (Figure 1A). CaCl2 treatment significantly reduced the browning index under refrigeration. On day 35 of storage, browning index in CaCl2-treated fruit was 47.06% lower than that in control. The firmness in loquat fruit displayed an increase trend during the entire storage period (Figure 1B). CaCl2 treatment remarkably suppressed the upward trend of firmness compared to the control. Firmness in CaCl2-treated fruit was 10.18% lower compared with control on the 35th day of storage. Extractable juice of the control fruit declined gradually during cold storage (Figure 1C), whereas CaCl2-treated fruit displayed markedly higher extractable juice than that of the control from day 14 to day 35 of storage. These results indicated that CaCl2 treatment efficiently suppressed the increase in browning index, firmness, as well as retained higher extractable juice rate, consequently inhibiting CI symptoms of loquat fruit during cold storage.
3.2. Effects of CaCl2 Treatment on Generation Rate and H2O2 Content of Loquat Fruit
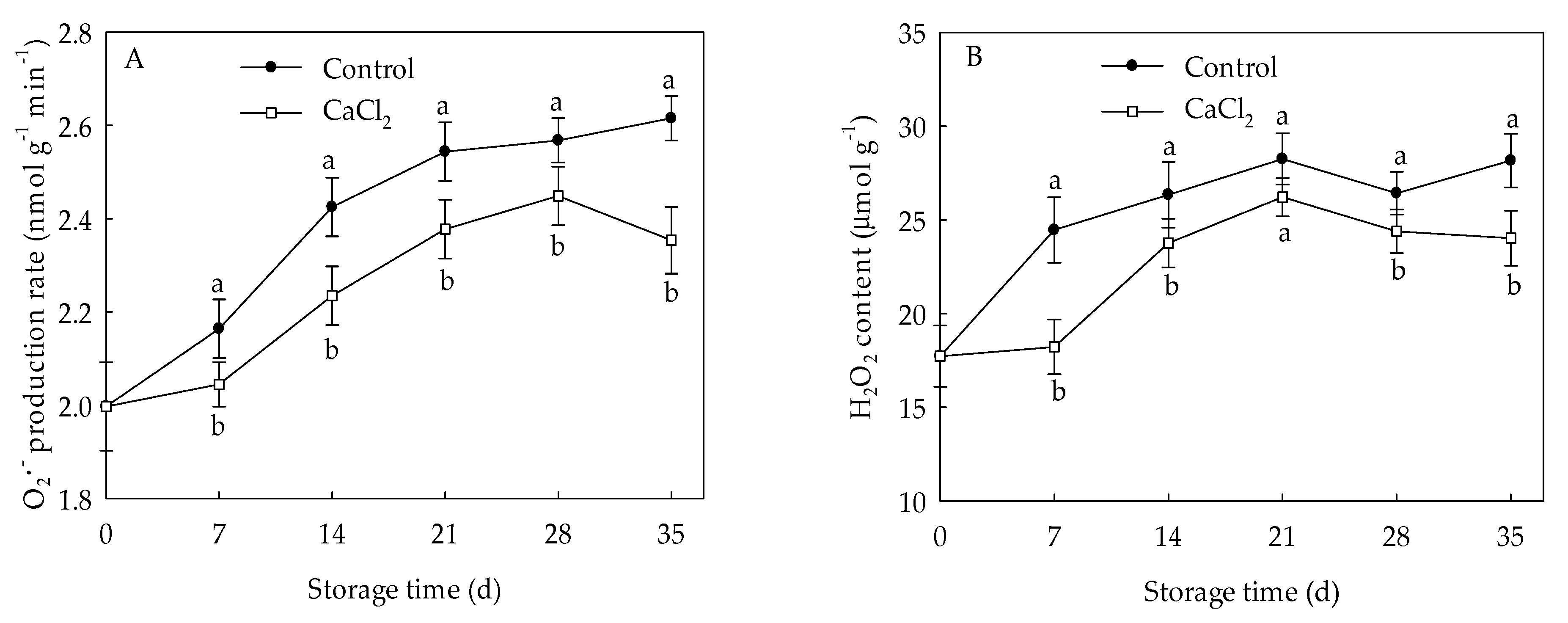
generation rate and H2O2 level in control loquat fruit displayed overall increments during cold storage (Figure 2). However, compared to the control, the application of CaCl2 remarkably reduced the accumulation of and H2O2 throughout the whole storage time. At the end of storage, the generation rate and H2O2 content of CaCl2-treated fruit were 10.00% and 14.71% lower (p < 0.05) than those in control fruit, respectively.
3.3. Effects of CaCl2 Treatment on DPPH and Radical Scavenging Capacity of Loquat Fruit
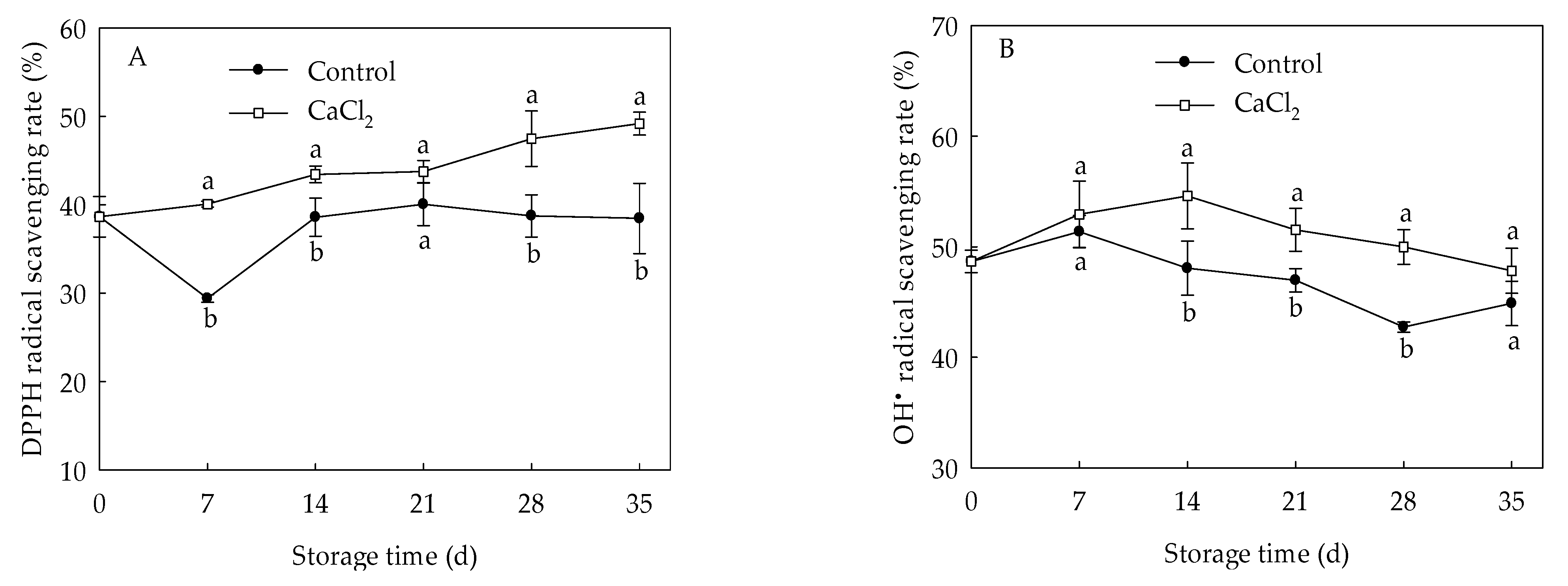
DPPH radical scavenging rate of control loquat fruit declined at the first 7 days of storage, and increased gradually from day 7 to day 21, then maintained relatively stable, while the CaCl2-treated fruit exhibited a gradually upward trend (Figure 3A). CaCl2 treatment maintained higher DPPH radical scavenging capacity within the entire storage period, and the DPPH radical scavenging rate was 27.85% higher in CaCl2-treated fruit than that of the control on the 35th day. The radical scavenging rate in control loquat fruit increased before day 7 of storage, but declined during day 7 to day 28, then displayed a slight increase (Figure 3B). However, the radical scavenging rate in the fruit treated with CaCl2 was higher compared to control fruit within the whole storage time, and presented a markedly (p < 0.05) higher radical scavenging rate during 14 d to 28 d of storage.
3.4. Effects of CaCl2 Treatment on CAT, SOD Activities and EjCAT, EjSOD Expressions of Loquat Fruit
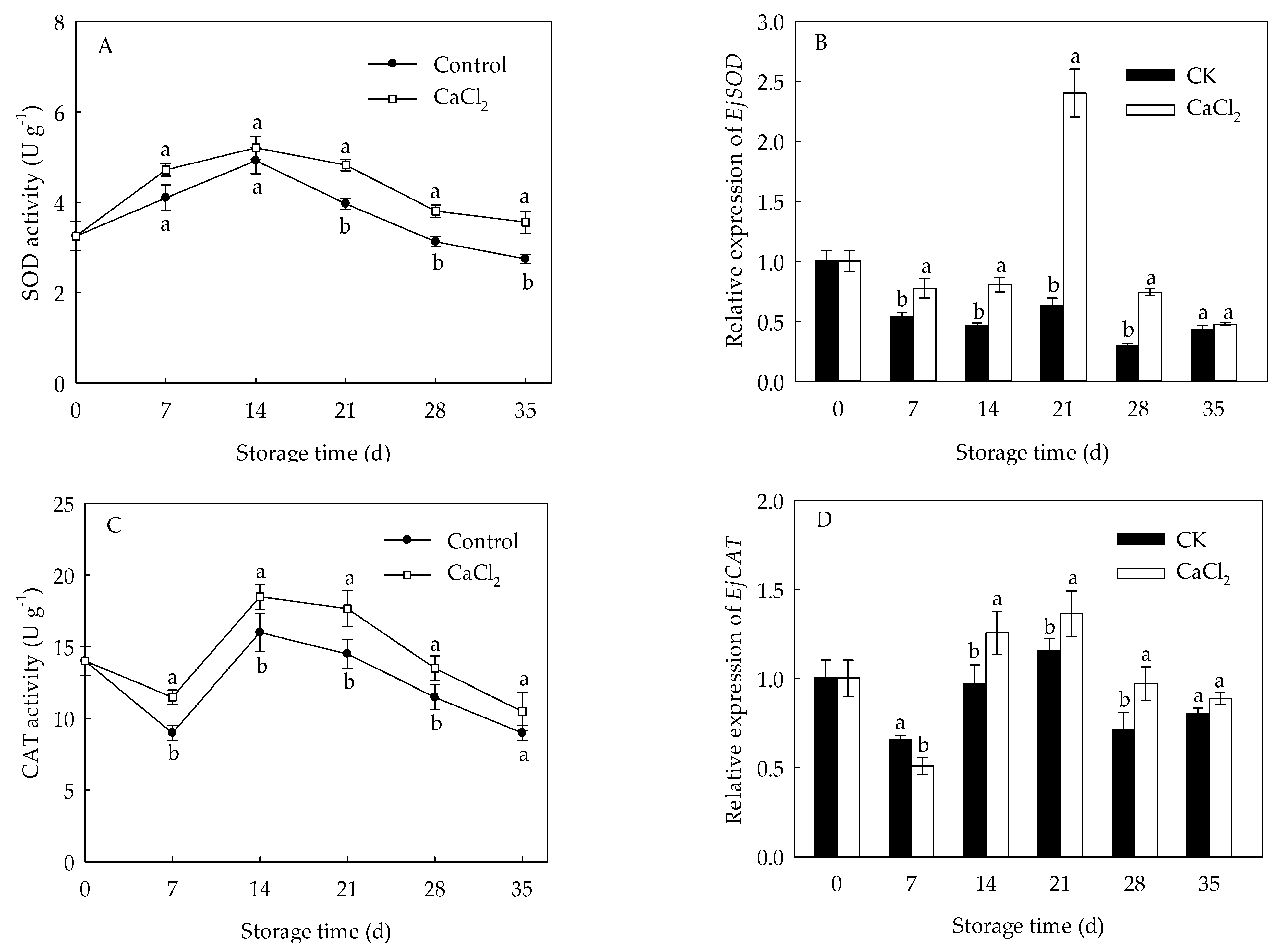
The SOD activity of loquat fruit rose quickly from 0 to 14 d, but gradually decreased thereafter (Figure 4A). The CAT activity in loquat fruit dropped slowly from day 0 to day 7 of storage, but quickly rose from 7 d to 14 d, followed by a gradual declination (Figure 4C). CaCl2 treatment enhanced SOD and CAT activities throughout the whole storage period, with 21.61% and 21.84% higher than those in control on day 21, respectively. Expression levels of EjSOD and EjCAT presented a similar trend with that of SOD and CAT enzymes activities (Figure 4B,D). CaCl2 treatment up regulated the expression of EjSOD and EjCAT, which were approximate 2.80-fold and 17.83% higher on the 21st day of storage compared to control, respectively.
3.5. Effects of CaCl2 Treatment on Contents of AsA, GSH, GSSG and GSH/GSSG Ratio of Loquat Fruit
The content of AsA in loquat fruit continuously declined with the extension of storage time (Figure 5A), whereas GSH content rose quickly from day 0 to day 21 of storage, then dropped slowly during 21–28 d, followed by a sharp increase (Figure 5B). Change in content of GSSG in loquat fruit was similar to that of AsA content (Figure 5C). On the contrary, GSH/GSSG in loquat fruit displayed a steady increase trend during cold storage (Figure 5D). CaCl2 treatment maintained higher AsA content and GSH content compared to the control, whereas GSSG content was lower in CaCl2-treated fruit than that in the control group. CaCl2 treatment remarkably increased GSH/GSSG within the entire storage time, and the GSH/GSSG in CaCl2-treated fruit was 34.92% higher on the 35th day of storage compared with control.
3.6. Effects of CaCl2 Treatment on Enzyme Activity and Gene Expression of APX, GR, DHAR, and MDHAR of Loquat Fruit
The APX activity in loquat fruit reached the peak on the 21st day of storage, and dropped quickly afterwards (Figure 6A). CaCl2-treated fruit exhibited higher APX activity compared to control fruit. Change of GR activity in loquat fruit displayed a similar trend with APX activity, but the maximum value reached on the 28th day (Figure 6C). Compared to control group, CaCl2 treatment maintained higher GR activity during cold storage, with a remarkable discrepancy from day 14 to day 35, except day 21 of storage.
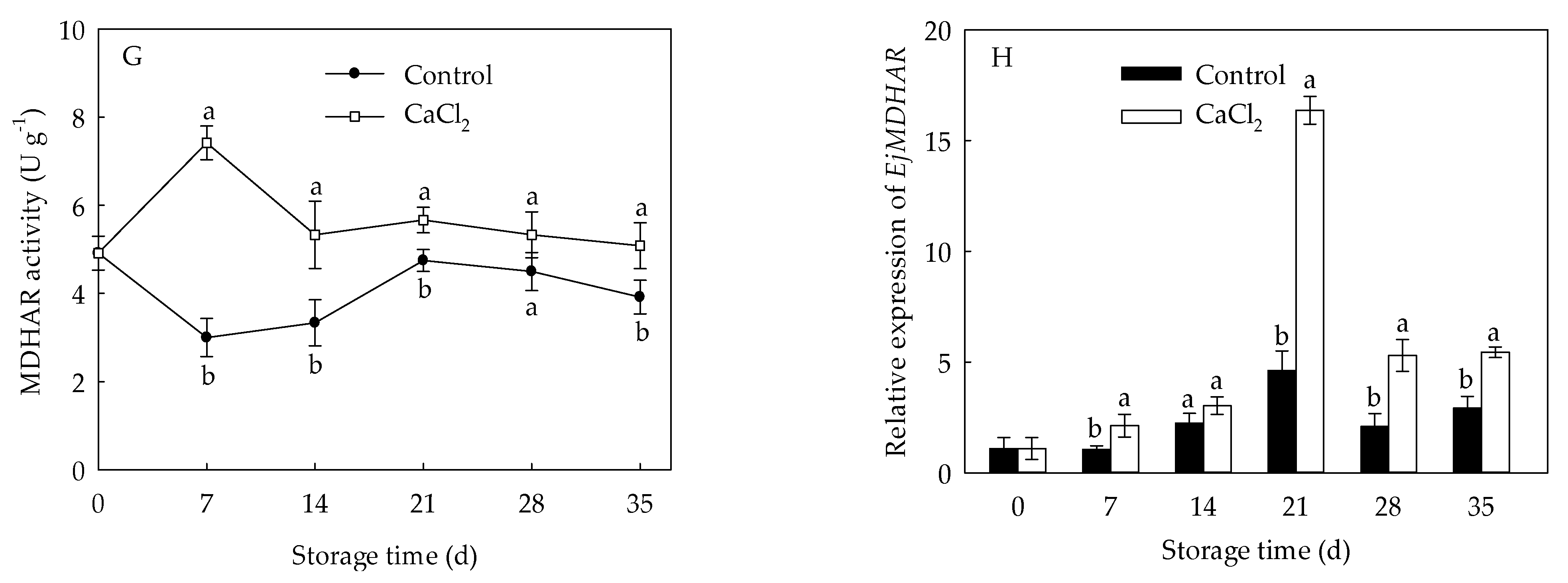
The DHAR activity in control loquat fruit rose at first 14 d of storage, then declined during 14–28 d, followed by a slight increase (Figure 6E). However, CaCl2-treated fruit exhibited remarkably higher DHAR activity from day 14 to day 35 compared to the control fruit. MDHAR activity of control fruit decreased sharply initially, and rose during day 7 to day 21, then a slight drop during day 21 to day 35 (Figure 6G). CaCl2 treatment displayed remarkably higher MDHAR activity than that in control group during day 7 to day 35, except day 28.
The expression of EjAPX and EjDHAR in control loquat fruit generally showed a downward trend, but the CaCl2 treatment remained higher expression of EjAPX and EjDHAR throughout the entire storage period (Figure 6B,F). On day 21 of storage, EjDHAR expression in CaCl2-treated loquat fruit was about 6.82-fold of control. Expression of EjGR and EjMDHAR in loquat fruit increased and peaked at 21 d of storage, followed by an overall decline (Figure 6D,H). Compared with control fruit, CaCl2 treatment up regulated the expression of EjGR and EjMDHAR within the whole storage time. Expression of EjGR and EjMDHAR in the fruit treated with CaCl2 were 3.08-fold and 3.55-fold of those in control on the 21st day of storage, respectively.
4. Discussion
Chilling injury, manifested as internal browning, an unusual increase in firmness, and juiceless pulp, is a primary problem that significantly impacts the quality of loquat fruit during cold storage [10]. It is reported that Ca2+ acts as an important second messenger fulfilling a crucial role in protection against chilling injury in plants [13]. Previous studies have confirmed that exogenous CaCl2 treatment improved chilling tolerance in loquat fruit under low temperature storage [20]. In the present study, CaCl2 treatment efficiently suppressed the increase in browning index and firmness, and maintained higher extractable juice in loquat fruit (Figure 1), demonstrating CaCl2 exerts a positive influence in enhancing chilling tolerance in harvested loquat fruit. Similar result was also obtained in pear fruit [17] and pineapple fruit [25].
Destabilization of the cell membrane has been well-known as the primary reason of chilling injury in plants. The overproduction of ROS, including , H2O2 and hydroxyl radical, may result in the occurrence of CI [26,27]. SOD, CAT, and APX are considered to be vital ROS-eliminating enzymes which involve an alleviation of chilling injury [9]. SOD can convert the overproduced into H2O2, while CAT and APX catalyze the H2O2 into H2O and O2 [28]. Antioxidant enzymes with higher activity and their integrated action have been demonstrated to be a part of the mechanism involved in the inhibition of oxidative damage and enhancement of cold tolerance in pear [29], blood orange [26], and banana [30]. In the current study, the accumulation of and H2O2 in loquat fruit (Figure 2) demonstrated the membrane damage from accumulation of ROS under chilling stress, and CaCl2 treatment efficiently reduced ROS accumulation, leading to a lower membrane peroxidation in loquat fruit [20]. Compared with the control fruit, CaCl2-treated loquat fruit maintained higher DPPH and radical scavenging ability (Figure 3), which are two crucial parameters on the antioxidant capacity of plants [31]. Meanwhile, the SOD and CAT activities in loquat fruit rose during the early stage of storage, and CaCl2 treatment enhanced SOD and CAT activities within the whole storage period (Figure 4A,C). Moreover, the expression of EjSOD and EjCAT was also up regulated by CaCl2 treatment (Figure 4B,D). The increased activities and gene expressions of SOD and CAT might explain the lower levels of and H2O2 in CaCl2-treated fruit. Zhang et al. [17] also reported that postharvest CaCl2 treatment promoted the activity and expression of SOD to against oxidative damage, inhibiting the browning in pear fruit after cold storage. Shi et al. [32] reported that CaCl2 treatment increased SOD and CAT activities to modulate redox homeostasis, contributing to improvement of chilling tolerance in bermudagrass. Similar results have also been confirmed in loquat fruit treated by methyl jasmonate [27]. These results indicated that the effect of CaCl2 on modulating the ROS homeostasis of loquat fruit was correlated to enhanced enzyme activities as well as gene expressions of SOD and CAT which, in turn, alleviated the occurrence of chilling injury.
The ASA–GSH cycle acts as a crucial antioxidant system to substantially scavenges H2O against oxidative damage, and APX, GR, DHAR, and MDHAR are key enzymes in this cycle [9]. AsA and GSH are products of the ASA-GSH cycle and important non-enzymatic antioxidants that are capable of directly or indirectly quenching ROS [7]. AsA is oxidized to the monodehydroascorbate (MDHA) radical, while APX converts H2O2 into the H2O with the help of AsA as an electron donor. MDHA regenerates AsA by MDHAR and is spontaneously converted to dehydroascorbate (DHA). DHA is reduced to AsA again through GSH, which leads to its oxidation to produce GSSG by DHAR. GSH regenerate from GSSG with catalyzation of GR [33]. Previous studies have demonstrated that enhanced AsA, GSH contents, and GSH/GSSG ratio are beneficial for the resistance to oxidative damage and improvement of chilling tolerance in plants [34,35]. Zhang et al. [19] confirmed that Ca2+ treatment could increase the activities of APX and GR to activate AsA-GSH cycle, and to effectively inhibit the increase in H2O2, thus, maintaining redox balance to better alleviate the occurrence of oxidative damage to green peppers under cold stress. In the current study, CaCl2-treated loquat fruit displayed higher levels of AsA and GSH contents (Figure 5A,B), lower GSSG content (Figure 5C), and retained higher GSH/GSSG ratio (Figure 5D) compared with the control fruit. Furthermore, CaCl2 enhanced the APX, GR, DHAR, and MDHAR activities in loquat fruit under cold storage (Figure 6A,C,E,G), which was conducive to maintaining a higher content of AsA, GSH and GSH/GSSG ratio. These findings were consistent with the result that ROS levels in CaCl2-treated fruit was lower than that in control (Figure 2). Similar results that AsA-GSH cycle played a crucial role in maintaining redox balance by loquat fruit under chilling injury were also reported by Cao et al. [10].
Additionally, the numerous reports showed that the increased expression of APX, GR, DHAR, MDHAR could alleviate oxidative injury due to their role in modulation of AsA–GSH cycle system in plants [33]. Song et al. [7] demonstrated that the mitigation of CI in peach fruit treated by hypobaric treatment was associated with its influence on activating the AsA-GSH cycle system through inducing GR, MDHAR1, and APX gene expressions to avoid oxidative damage. In the current study, CaCl2 treatment up regulated the EjAPX, EjGR, EjDHAR, and EjMDHR expressions in loquat fruit (Figure 6B,D,F,H), which exerted positive influence in the maintenance of high activities of antioxidant enzymes involved in AsA–GSH cycle, resulting in the inhibition of ROS accumulation and chilling injury symptoms. It was in agreement with previous work which stated that the up-regulated CaAPX1, CaMDHAR1, and CaDHAR1 expressions in bell peppers by GSH treatment promoted activities of AsA-GSH cycle enzymes and regenerated AsA and GSH, consequently bringing about lower oxidative damage and alleviation of chilling injury [9]. Thus, we conclude that CaCl2 treatment could mitigate oxidative damage by activating AsA-GSH cycle system in loquat fruit during chilling injury.
5. Conclusions
In conclusion, CaCl2 treatment effectively enhanced cold tolerance and alleviated the occurrence of CI in loquat fruit. The inhibition in occurrence of CI by CaCl2 treatment might be ascribed to its capacity to reduce ROS accumulation by triggering of the antioxidant system including enzymatic and non-enzymatic antioxidant activities, thereby reducing cell membrane oxidative damage. These findings provide insight into the improvement of cold tolerance in loquat fruit through modulating ROS homeostasis by CaCl2 treatment. However, further molecular evidence is needed to support an explanation of how Ca2+ triggers low-temperature tolerance.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/foods10071662/s1, Table S1. Primer sequences for Real-time PCR analysis.
Author Contributions
Conceptualization, Y.H. and P.J.; data curation, Y.H. and Z.L.; funding acquisition, P.J.; investigation, Y.H. and Z.L.; methodology, Y.H. and Z.L.; supervision, P.J.; validation, P.J. and Y.Z.; writing—original draft, Y.H. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the National Natural Science Foundation of China (No.31972125, 31671901).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors gratefully acknowledge laboratory colleagues for their help.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Zheng, Y.H.; Li, S.Y.; Xi, Y.F. Changes of cell wall substances in relation to flesh woodiness in cold-stored loquat fruits. Acta Phytophysiol. Sin. 2000, 26, 306–310. [Google Scholar] [CrossRef]
- Cai, C.; Xu, C.J.; Li, X.; Ferguson, I.B.; Chen, K.S. Accumulation of lignin in relation to change in activities of lignification enzymes in loquat fruit flesh after harvest. Postharvest Boil. Technol. 2006, 40, 163–169. [Google Scholar] [CrossRef]
- Dreyer, A.; Dietz, K.J. Reactive Oxygen species and the redox-regulatory network in cold stress acclimation. Antioxidants 2018, 7, 169. [Google Scholar] [CrossRef] [Green Version]
- You, J.; Chan, Z.L. ROS regulation during abiotic stress responses in crop plants. Front. Plant Sci. 2015, 6, 1092. [Google Scholar] [CrossRef] [Green Version]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Ye, L.; Wang, Y.; Zhou, X.; Yang, J.; Wang, J.; Cao, K.; Zou, Z. Melatonin increases the chilling tolerance of chloroplast in cucumber seedlings by regulating photosynthetic electron flux and the ascorbate-glutathione cycle. Front. Plant Sci. 2016, 7, 1814. [Google Scholar] [CrossRef] [Green Version]
- Song, L.L.; Wang, J.H.; Shafi, M.; Liu, Y.; Wang, J.; Wu, J.S.; Wu, A.M. Hypobaric treatment effects on chilling injury, mitochondrial dysfunction, and the ascorbate–glutathione (AsA-GSH) cycle in postharvest peach fruit. J. Agric. Food Chem. 2016, 64, 4665–4674. [Google Scholar] [CrossRef]
- Liu, J.L.; Sun, J.H.; Pan, Y.G.; Yun, Z.; Zhang, Z.K.; Guoxiang Jiang, G.X.; Jiang, Y.M. Endogenous melatonin generation plays a positive role in chilling tolerance in relation to redox homeostasis in litchi fruit during refrigeration. Postharvest Boil. Technol. 2021, 178, 111554. [Google Scholar] [CrossRef]
- Yao, M.; Ge, W.; Zhou, Q.; Zhou, X.; Luo, M.; Zhao, Y.; Wei, B.; Ji, S. Exogenous glutathione alleviates chilling injury in postharvest bell pepper by modulating the ascorbate-glutathione (AsA-GSH) cycle. Food Chem. 2021, 352, 129458. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Yang, Z.; Cai, Y.; Zheng, Y. Fatty acid composition and antioxidant system in relation to susceptibility of loquat fruit to chilling injury. Food Chem. 2011, 127, 1777–1783. [Google Scholar] [CrossRef]
- Cai, Y.; Cao, S.; Yang, Z.; Zheng, Y. MeJA regulates enzymes involved in ascorbic acid and glutathione metabolism and improves chilling tolerance in loquat fruit. Postharvest Boil. Technol. 2011, 59, 324–326. [Google Scholar] [CrossRef]
- Hocking, B.; Tyerman, S.D.; Burton, R.A.; Gilliham, M. Fruit calcium: Transport and physiology. Front. Plant Sci. 2016, 7, 569. [Google Scholar] [CrossRef] [Green Version]
- Ranty, B.; Aldon, D.; Cotelle, V.; Galaud, J.P.; Thuleau, P.; Mazars, C. Calcium sensors as key hubs in plant responses to biotic and abiotic stresses. Front. Plant Sci. 2016, 7, 327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, P.; Yang, T.; Poovaiah, B.W. Calcium signaling-mediated plant response to cold stress. Int. J. Mol. Sci. 2018, 19, 3896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, H.; Wu, C.; Luo, C.; Wei, M.; Qu, S.; Wang, S. Overexpression of MdCPK1a gene, a calcium dependent protein kinase in apple, increase tobacco cold tolerance via scavenging ROS accumulation. PLoS ONE 2020, 15, e0242139. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhou, Q.; Zhou, X.; Wei, B.; Ji, S. Calcium treatment alleviates pericarp browning of ‘nanguo’ pears by regulating the GABA shunt after cold storage. Front. Plant Sci. 2020, 11, 580986. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wang, J.; Zhou, B.; Li, G.; Liu, Y.; Xia, X.L.; Xiao, Z.G.; Fei, L.; Ji, S.J. Calcium inhibited peel browning by regulating enzymes in membrane metabolism of ‘Nanguo’ pears during post-ripeness after refrigerated storage. Sci. Hortic. 2019, 244, 15–21. [Google Scholar] [CrossRef]
- Wei, D.; Zhao, X. Calcium maintained higher quality and enhanced resistance against chilling stress by regulating enzymes in reactive oxygen and biofilm metabolism of chinese winter jujube fruit. J. Food Biochem. 2020, 44, e13161. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.Y.; Ma, M.J.; Ye, B.; Liu, L.; Ji, S.J. Calcium ion improves cold resistance of green peppers (Capsicum annuum L.) by regulating the activity of protective enzymes and membrane lipid composition. Sci. Hortic. 2021, 277, 109789. [Google Scholar] [CrossRef]
- Li, Z.Y.; Wang, L.; Xie, B.; Hu, S.Q.; Zheng, Y.H.; Jin, P. Effects of exogenous calcium and calcium chelant on cold tolerance of postharvest loquat fruit. Sci. Hortic. 2020, 269, 109391. [Google Scholar] [CrossRef]
- Zhang, Y.; Jin, P.; Huang, Y.P.; Shan, T.M.; Wang, L.; Li, Y.Y.; Zheng, Y.H. Effect of hot water combined with glycine betaine alleviates chilling injury in cold-stored loquat fruit. Postharvest Boil. Technol. 2016, 118, 141–147. [Google Scholar] [CrossRef]
- Wang, L.; Shao, S.; Madebo, M.P.; Hou, Y.; Zheng, Y.; Jin, P. Effect of nano-SiO2 packing on postharvest quality and antioxidant capacity of loquat fruit under ambient temperature storage. Food Chem. 2020, 315, 126295. [Google Scholar] [CrossRef] [PubMed]
- Asif, M.H.; Dhawan, P.; Math, P. A simple procedure for the isolation of high quality RNA from ripening banana fruit. Plant Mol. Biol. Rep. 2000, 18, 109–115. [Google Scholar] [CrossRef]
- Zeng, J.K.; Li, X.; Zhang, J.; Ge, H.; Yin, X.R.; Chen, K.S. Regulation of loquat fruit low temperature response and lignification involves interaction of heat shock factors and genes associated with lignin biosynthesis. Plant Cell Environ. 2016, 39, 1780–1789. [Google Scholar] [CrossRef]
- Youryon, P.; Supapvanich, S.; Kongtrakool, P.; Wongs-Aree, C. Calcium chloride and calcium gluconate peduncle infiltrations alleviate the internal browning of queen pineapple in refrigerated storage. Hortic. Environ. Biotechnol. 2018, 59, 205–213. [Google Scholar] [CrossRef]
- Habibi, F.; Ramezanian, A.; Guillén, F.; Martínez-Romero, D.; Serrano, M.; Valero, D. Susceptibility of blood orange cultivars to chilling injury based on antioxidant system, physiological and biochemical responses at different storage temperatures. Foods 2020, 9, 1609. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Zheng, Y.; Wang, K.; Peng, J.; Rui, H. Methyl jasmonate reduces chilling injury and enhances antioxidant enzyme activity in postharvest loquat fruit. Food Chem. 2009, 115, 1458–1463. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Sun, H.; Luo, M.L.; Zhou, X.; Zhou, Q.; Ji, S.J. Exogenous glycine betaine treatment alleviates low temperature-induced pericarp browning of ‘nanguo’ pears by regulating antioxidant enzymes and proline metabolism. Food Chem. 2019, 306, 125626. [Google Scholar] [CrossRef]
- Hao, J.; Li, X.; Xu, G.; Huo, Y.; Yang, H. Exogenous progesterone treatment alleviates chilling injury in postharvest banana fruit associated with induction of alternative oxidase and antioxidant defense. Food Chem. 2019, 286, 329–337. [Google Scholar] [CrossRef]
- Tan, X.L.; Zhao, Y.T.; Shan, W.; Kuang, J.F.; Chen, J.Y. Melatonin delays leaf senescence of postharvest chinese flowering cabbage through ros homeostasis. Food Res. Int. 2020, 138, 109790. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Ye, T.; Ba, O.Z.; Liu, X.; Chan, Z. Comparative proteomic and metabolomic analyses reveal mechanisms of improved cold stress tolerance in bermudagrass (cynodon dactylon (L.) Pers.) by exogenous calcium. J. Integr. Plant Biol. 2014, 56, 1064–1079. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Anee, T.I.; Parvin, K.; Nahar, K.; Mahmud, J.A.; Fujita, M. Regulation of ascorbate-glutathione pathway in mitigating oxidative damage in plants under abiotic stress. Antioxidants 2019, 8, 384. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.Y.; Wang, Y.T.; Pan, X.B.; Xi, Z.M. Amelioration of cold-induced oxidative stress by exogenous 24-epibrassinolide treatment in grapevine seedlings: Toward regulating the ascorbate–glutathione cycle. Sci. Hortic. 2019, 244, 379–387. [Google Scholar] [CrossRef]
- Huan, C.; Han, S.A.; Jiang, L.; An, X.J.; Yu, M.L.; Xu, Y.; Ma, R.J.; Yu, Z.F. Postharvest hot air and hot water treatments affect the antioxidant system in peach fruit during refrigerated storage. Postharvest Boil. Technol. 2017, 126, 1–14. [Google Scholar] [CrossRef]
Figure 1.
Browning index (A), firmness (B), and extractable juice (C) in loquat fruit stored at 1 °C after treatment with 1% CaCl2 or water (control). Vertical bars indicate the SE of mean (n = 3). The symbols a and b represent a significant difference in the two treatments (p < 0.05).
Figure 1.
Browning index (A), firmness (B), and extractable juice (C) in loquat fruit stored at 1 °C after treatment with 1% CaCl2 or water (control). Vertical bars indicate the SE of mean (n = 3). The symbols a and b represent a significant difference in the two treatments (p < 0.05).

Figure 2.
generation rate (A) and H2O2 content (B) in loquat fruit stored at 1 °C after treatment with 1% CaCl2 or water (control). Vertical bars indicate the SE of mean (n = 3). The symbols a and b represent a significant difference in the two treatments (p < 0.05).
Figure 2.
generation rate (A) and H2O2 content (B) in loquat fruit stored at 1 °C after treatment with 1% CaCl2 or water (control). Vertical bars indicate the SE of mean (n = 3). The symbols a and b represent a significant difference in the two treatments (p < 0.05).

Figure 3.
DPPH (A) and (B) radical scavenging capacity in loquat fruit stored at 1 °C after treatment with 1% CaCl2 or water (control). Vertical bars indicate the SE of mean (n = 3). The symbols a and b represent a significant difference in the two treatments (p < 0.05).
Figure 3.
DPPH (A) and (B) radical scavenging capacity in loquat fruit stored at 1 °C after treatment with 1% CaCl2 or water (control). Vertical bars indicate the SE of mean (n = 3). The symbols a and b represent a significant difference in the two treatments (p < 0.05).

Figure 4.
SOD activity (A), EjSOD expression (B), CAT activity (C) and EjCAT expression (D) in loquat fruit stored at 1 °C after treatment with 1% CaCl2 or water (control). Vertical bars indicate the SE of mean (n = 3). The symbols a and b represent a significant difference in the two treatments (p < 0.05).
Figure 4.
SOD activity (A), EjSOD expression (B), CAT activity (C) and EjCAT expression (D) in loquat fruit stored at 1 °C after treatment with 1% CaCl2 or water (control). Vertical bars indicate the SE of mean (n = 3). The symbols a and b represent a significant difference in the two treatments (p < 0.05).

Figure 5.
AsA (A), GSH (B), GSSG (C) contents and GSH/GSSG (D) in loquat fruit stored at 1 °C after treatment with 1% CaCl2 or water (control). Vertical bars indicate the SE of mean (n = 3). The symbols a and b represent a significant difference in the two treatments (p < 0.05).
Figure 5.
AsA (A), GSH (B), GSSG (C) contents and GSH/GSSG (D) in loquat fruit stored at 1 °C after treatment with 1% CaCl2 or water (control). Vertical bars indicate the SE of mean (n = 3). The symbols a and b represent a significant difference in the two treatments (p < 0.05).

Figure 6.
APX activity (A), EjAPX expression (B), GR activity (C), EjGR expression (D), DHAR activity (E), EjDHAR expression (F), MDHAR activity (G), and EjMDHAR expression (H) in loquat fruit stored at 1 °C after treatment with 1% CaCl2 or water (control). Vertical bars indicate the SE of mean (n = 3). The symbols a and b represent a significant difference in the two treatments (p < 0.05).
Figure 6.
APX activity (A), EjAPX expression (B), GR activity (C), EjGR expression (D), DHAR activity (E), EjDHAR expression (F), MDHAR activity (G), and EjMDHAR expression (H) in loquat fruit stored at 1 °C after treatment with 1% CaCl2 or water (control). Vertical bars indicate the SE of mean (n = 3). The symbols a and b represent a significant difference in the two treatments (p < 0.05).


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hou, Y.; Li, Z.; Zheng, Y.; Jin, P. Effects of CaCl2 Treatment Alleviates Chilling Injury of Loquat Fruit (Eribotrya japonica) by Modulating ROS Homeostasis. Foods 2021, 10, 1662. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10071662
AMA Style
Hou Y, Li Z, Zheng Y, Jin P. Effects of CaCl2 Treatment Alleviates Chilling Injury of Loquat Fruit (Eribotrya japonica) by Modulating ROS Homeostasis. Foods. 2021; 10(7):1662. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10071662
Chicago/Turabian StyleHou, Yuanyuan, Ziying Li, Yonghua Zheng, and Peng Jin. 2021. "Effects of CaCl2 Treatment Alleviates Chilling Injury of Loquat Fruit (Eribotrya japonica) by Modulating ROS Homeostasis" Foods 10, no. 7: 1662. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10071662
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.