
Determination of Oxidized Lipids in Commonly Consumed Foods and a Preliminary Analysis of Their Binding Affinity to PPARγ

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Thermal Oxidation of Vegetable Oil
2.3. Room-Temperature Foods (RTF) and Fried Foods
2.4. Extraction of Lipids
2.5. Quantifying the Products of Oxidation
2.5.1. Primary Oxidation Products
2.5.2. Secondary Oxidation Products
2.6. Determining the Binding Affinity of Extracted Lipids for PPARγ
2.7. Alkaline Hydrolysis of Vegetable Oil
2.8. Data Analysis
3. Results
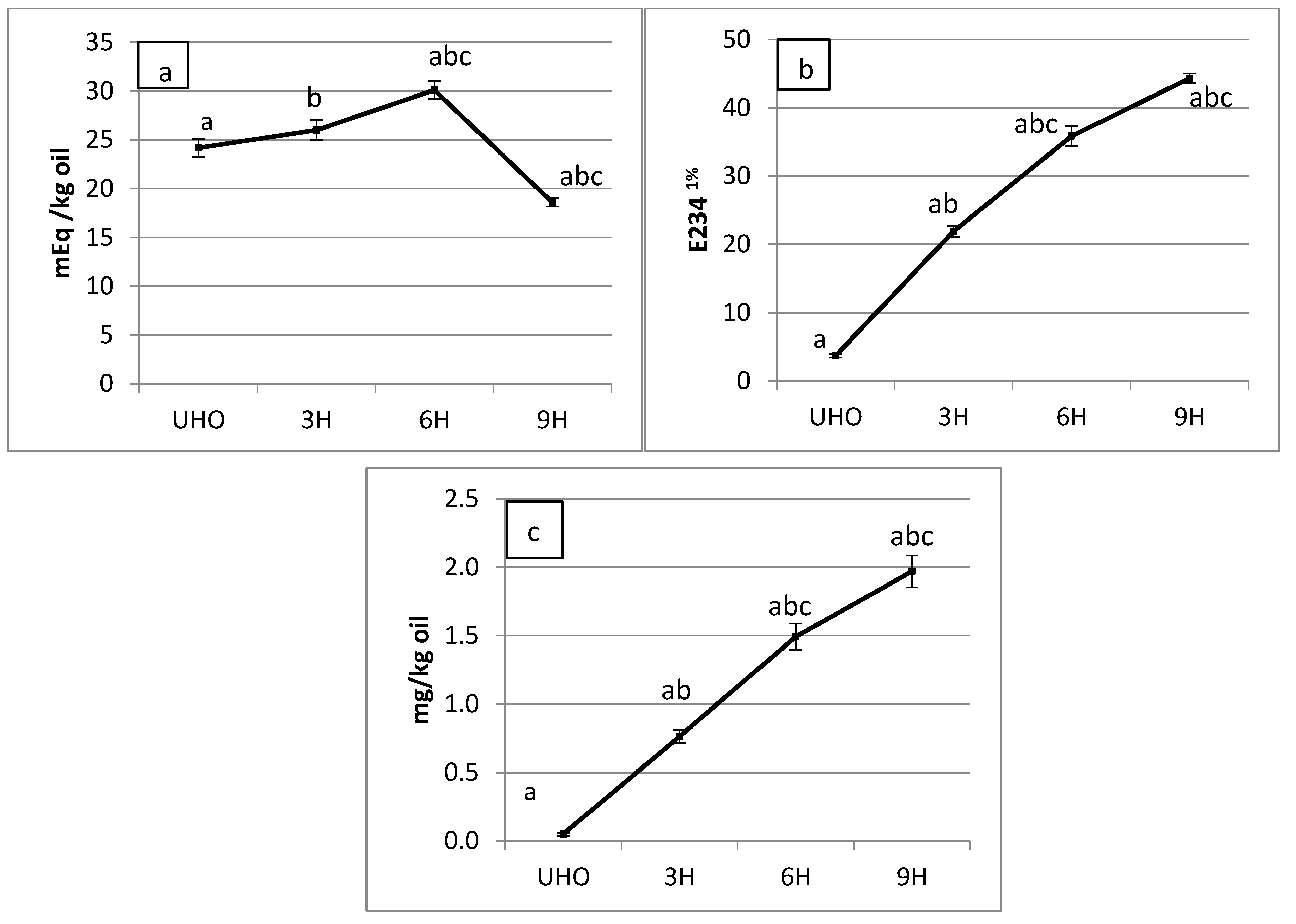
3.1. Lipid Oxidation Products in Soy Vegetable Oil
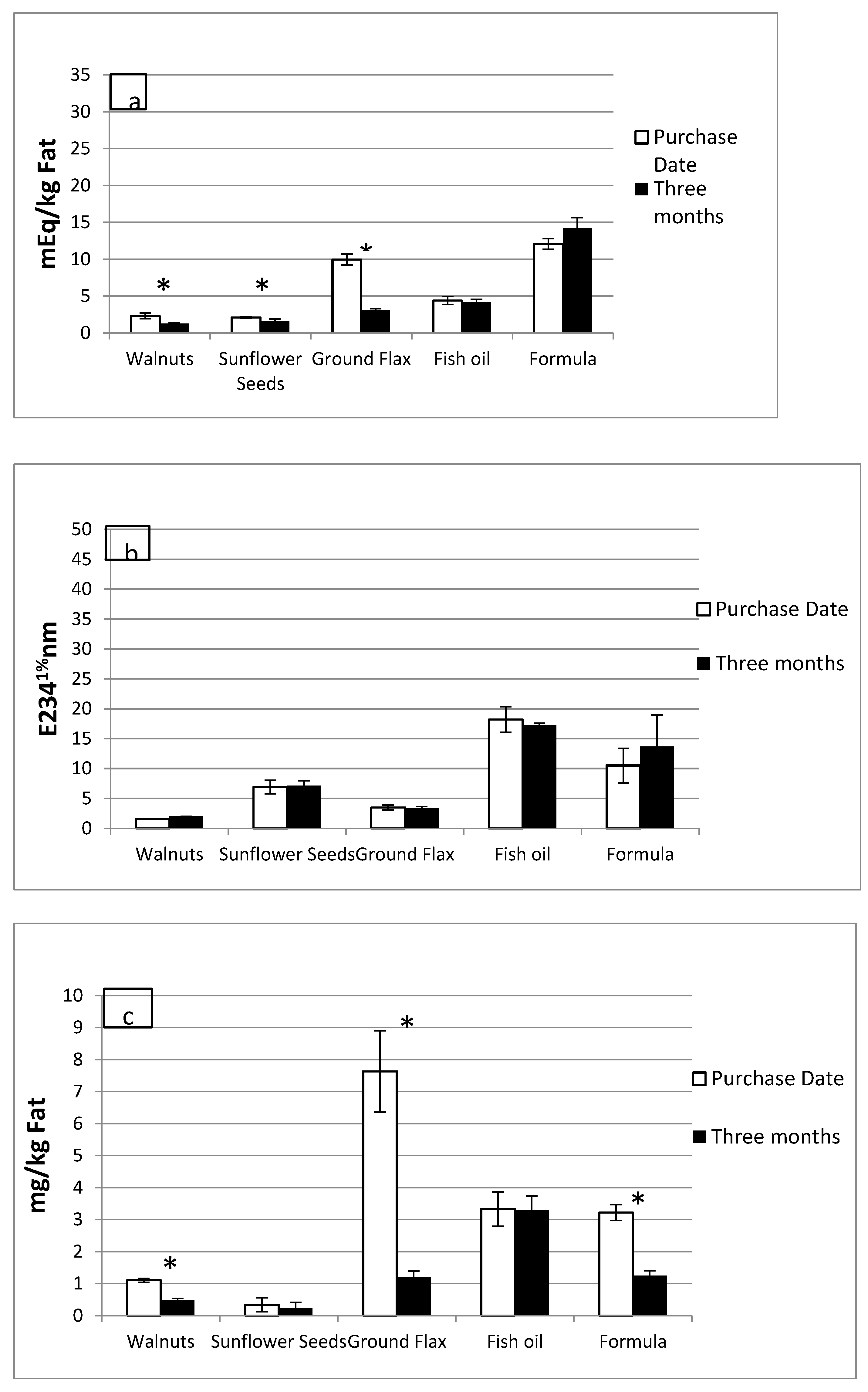
3.2. Lipid Oxidation Products in RTF and Fried Foods
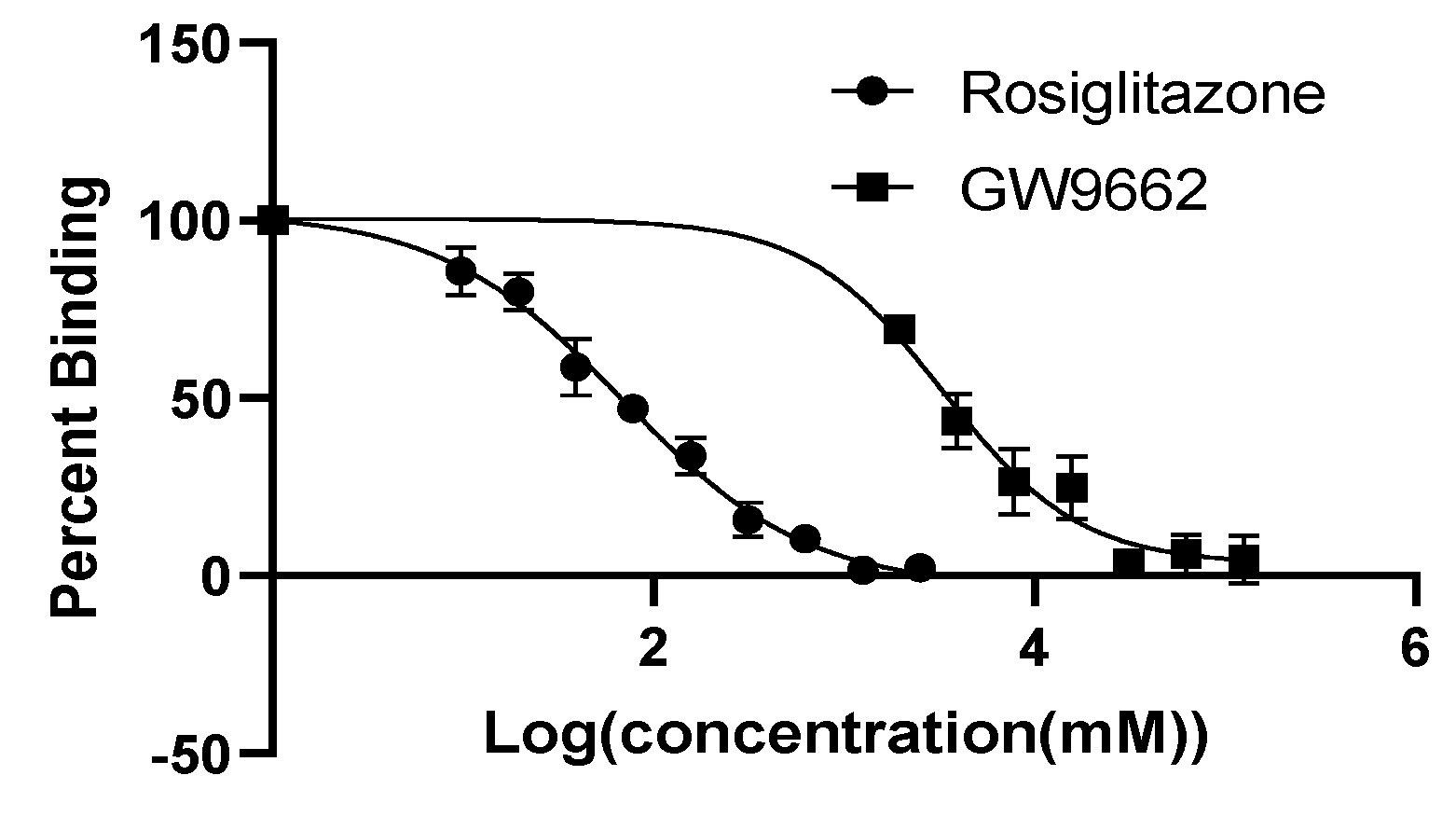
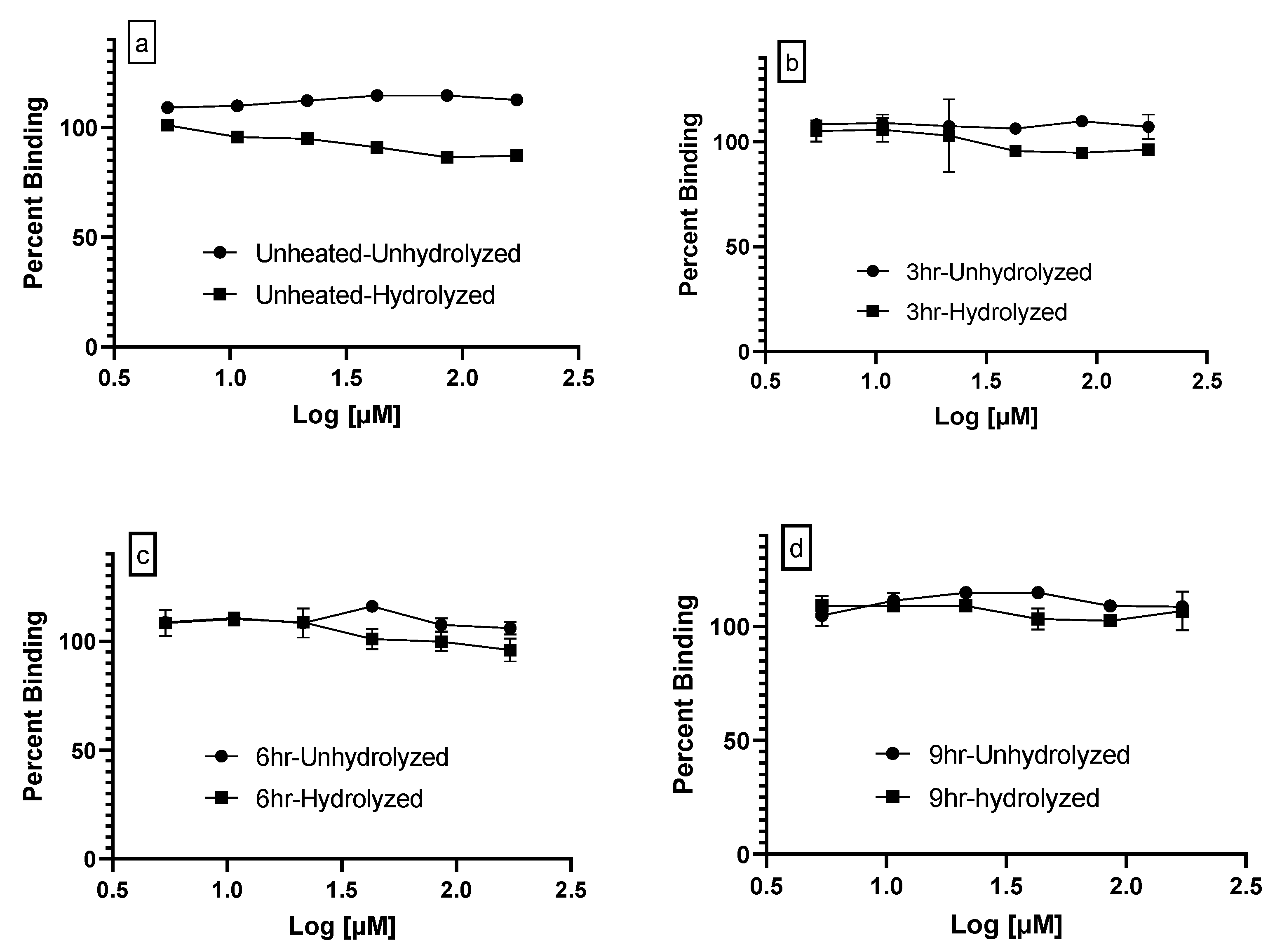
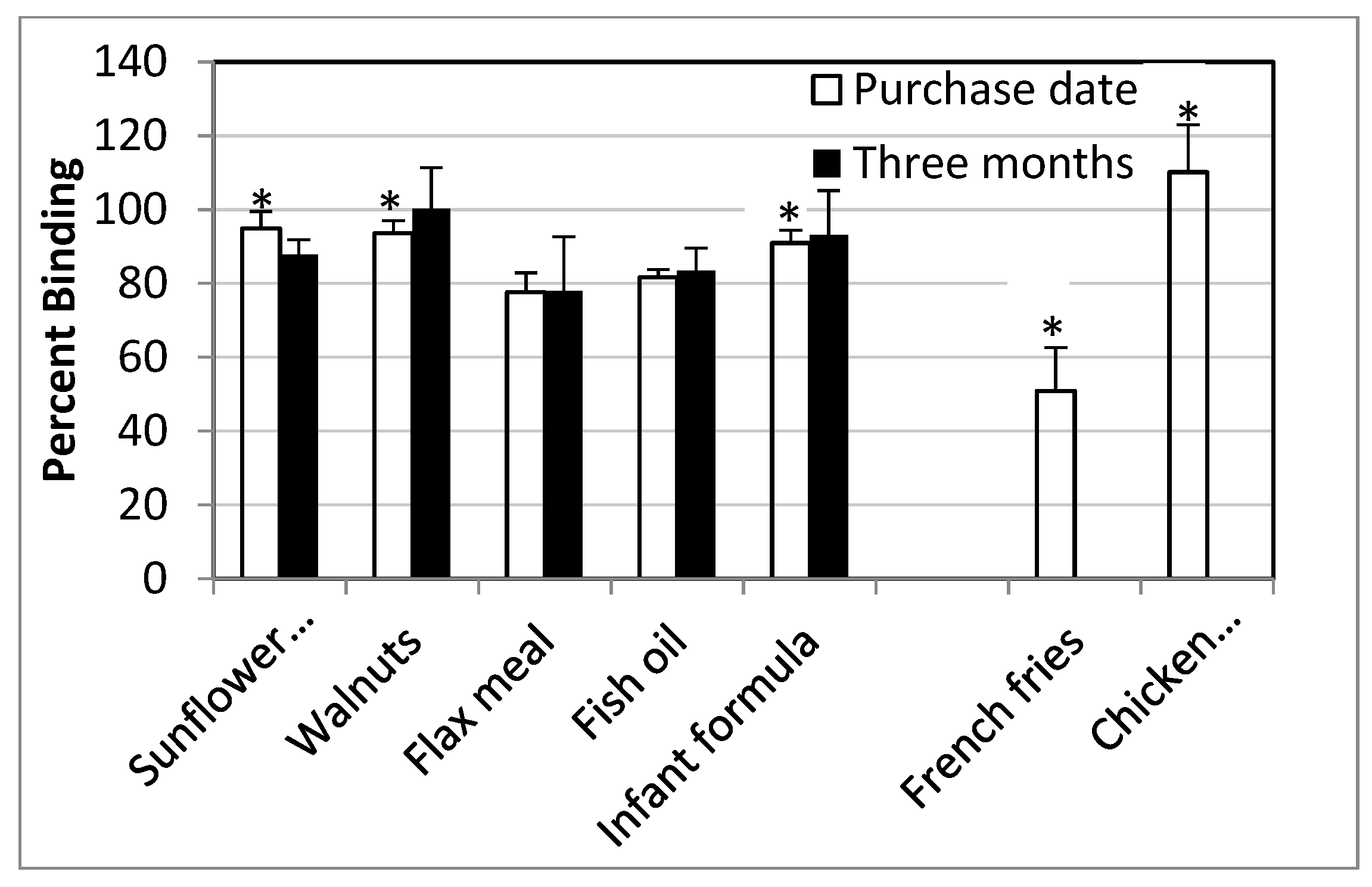
3.3. PPARγ Ligand Binding
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lee, S.; Birukov, K.G.; Romanoski, C.E.; Springstead, J.R.; Lusis, A.J.; Berliner, J.A. Role of phospholipid oxidation products in atherosclerosis. Circ. Res. 2012, 111, 778–799. [Google Scholar] [CrossRef]
- Zhong, S.; Li, L.; Shen, X.; Li, Q.; Xu, W.; Wang, X.; Tao, Y.; Yin, H. An update on lipid oxidation and inflammation in cardiovascular diseases. Free Radic. Biol. Med. 2019, 144, 266–278. [Google Scholar] [CrossRef]
- Florens, N.; Calzada, C.; Lyasko, E.; Juillard, L.; Soulage, C.O. Modified Lipids and Lipoproteins in Chronic Kidney Disease: A New Class of Uremic Toxins. Toxins 2016, 8, 376. [Google Scholar] [CrossRef] [Green Version]
- Sottero, B.; Rossin, D.; Poli, G.; Biasi, F. Lipid Oxidation Products in the Pathogenesis of Inflammation-related Gut Diseases. Curr. Med. Chem. 2018, 25, 1311–1326. [Google Scholar] [CrossRef] [PubMed]
- Sayon-Orea, C.; Martinez-Gonzalez, M.A.; Gea, A.; Flores-Gomez, E.; Basterra-Gortari, F.J.; Bes-Rastrollo, M. Consumption of fried foods and risk of metabolic syndrome: The SUN cohort study. Clin. Nutr. 2014, 33, 545–549. [Google Scholar] [CrossRef] [PubMed]
- Sayon-Orea, C.; Carlos, S.; Martínez-Gonzalez, M.A. Does cooking with vegetable oils increase the risk of chronic diseases?: A systematic review. Br. J. Nutr. 2015, 113, S36–S48. [Google Scholar] [CrossRef] [Green Version]
- Gadiraju, T.; Patel, Y.; Gaziano, J.; Djoussé, L. Fried Food Consumption and Cardiovascular Health: A Review of Current Evidence. Nutrients 2015, 7, 8424–8430. [Google Scholar] [CrossRef] [PubMed]
- Olivero-David, R.; Paduano, A.; Fogliano, V.; Vitaglione, P.; Bastida, S.; González-Muñoz, M.J.; Benedí, J.; Sacchi, R.; Sánchez-Muniz, F.J. Effect of Thermally Oxidized Oil and Fasting Status on the Short-Term Digestibility of Ketolinoleic Acids and Total Oxidized Fatty Acids in Rats. J. Agric. Food Chem. 2011, 59, 4684–4691. [Google Scholar] [CrossRef]
- Penumetcha, M.; Khan, N.; Parthasarathy, S. Dietary oxidized fatty acids: An atherogenic risk? J. Lipid Res. 2000, 41, 1473–1480. [Google Scholar] [CrossRef]
- Staprans, I.; Rapp, J.H.; Pan, X.M.; Kim, K.Y.; Feingold, K.R. Oxidized lipids in the diet are a source of oxidized lipid in chylomicrons of human serum. Arterioscler. Thromb. Vasc. Biol. 1994, 14, 1900–1905. [Google Scholar] [CrossRef] [Green Version]
- Ng, C.-Y.; Leong, X.-F.; Masbah, N.; Adam, S.K.; Kamisah, Y.; Jaarin, K. Heated vegetable oils and cardiovascular disease risk factors. Vascul. Pharmacol. 2014, 61, 1–9. [Google Scholar] [CrossRef]
- Khan-Merchant, N.; Penumetcha, M.; Meilhac, O.; Parthasarathy, S. Oxidized fatty acids promote atherosclerosis only in the presence of dietary cholesterol in low-density lipoprotein receptor knockout mice. J. Nutr. 2002, 132, 3256–3262. [Google Scholar] [CrossRef] [Green Version]
- Staprans, I.; Pan, X.M.; Rapp, J.H.; Feingold, K.R. The role of dietary oxidized cholesterol and oxidized fatty acids in the development of atherosclerosis. Mol. Nutr. Food Res. 2005, 49, 1075–1082. [Google Scholar] [CrossRef]
- Chao, P.-M.; Huang, H.-L.; Liao, C.-H.; Huang, S.-T.; Huang, C. A high oxidised frying oil content diet is less adipogenic, but induces glucose intolerance in rodents. Br. J. Nutr. 2007, 98, 63. [Google Scholar] [CrossRef] [Green Version]
- Rosen, E.D.; Sarraf, P.; Troy, A.E.; Bradwin, G.; Moore, K.; Milstone, D.S.; Spiegelman, B.M.; Mortensen, R.M. PPARγ is required for the differentiation of adipose tissue in vivo and in vitro. Mol. Cell 1999, 4, 611–617. [Google Scholar] [CrossRef]
- Tontonoz, P.; Spiegelman, B.M. Fat and Beyond: The Diverse Biology of PPARγ. Annu. Rev. Biochem. 2008, 77, 289–312. [Google Scholar] [CrossRef] [PubMed]
- Varga, T.; Czimmerer, Z.; Nagy, L. PPARs are a unique set of fatty acid regulated transcription factors controlling both lipid metabolism and inflammation. Biochim. Biophys. Acta BBA Mol. Basis Dis. 2011, 1812, 1007–1022. [Google Scholar] [CrossRef]
- Marion-Letellier, R.; Savoye, G.; Ghosh, S. Fatty acids, eicosanoids and PPAR gamma. Eur. J. Pharmacol. 2016, 785, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Itoh, T.; Fairall, L.; Amin, K.; Inaba, Y.; Szanto, A. Structural basis for the activation of PPARγ by oxidized fatty acids. Nat. Struct. Mol. Biol. 2008, 15, 924–931. [Google Scholar] [CrossRef] [PubMed]
- Hara, A.; Radin, N.S. Lipid extraction of tissues with a low-toxicity solvent. Anal. Biochem. 1978, 90, 420–426. [Google Scholar] [CrossRef] [Green Version]
- Washburn, K.W. A Modification of the Folch Method of Lipid Extraction for Poultry. Poult. Sci. 1989, 68, 1425–1427. [Google Scholar] [CrossRef]
- Woo, K.L.; Kim, M.C.; Ki, J.I. Study on Hydrolysis Method for Extremely Small Amount of Lipids by Organic Basic Sloution, Tetramethylammonium Hydroxide/Methanol and Capillary Gas Chromatographic Analysis of Fatty Acid Composition Depending on Derivatization Methods. J. Appl. Sci. 2002, 2, 804–815. [Google Scholar]
- Ebrahimzadeh, A.; Pirzad, F.; Tahanian, H.; Aghdam, M.S. Influence of Gum Arabic Enriched with GABA Coating on Oxidative Damage of Walnut Kernels. Food Technol. Biotechnol. 2019, 57, 554–560. [Google Scholar] [CrossRef] [PubMed]
- Mehyar, G.F.; Al-Ismail, K.; Han, J.H.; Chee, G.W. Characterization of Edible Coatings Consisting of Pea Starch, Whey Protein Isolate, and Carnauba Wax and their Effects on Oil Rancidity and Sensory Properties of Walnuts and Pine Nuts. J. Food Sci. 2012, 77, E52–E59. [Google Scholar] [CrossRef] [PubMed]
- Nichols, P.; Dogan, L.; Sinclair, A. Australian and New Zealand Fish Oil Products in 2016 Meet Label Omega-3 Claims and Are Not Oxidized. Nutrients 2016, 8, 703. [Google Scholar] [CrossRef] [Green Version]
- Jairoun, A.A.; Shahwan, M.; Zyoud, S.H. Fish oil supplements, oxidative status, and compliance behaviour: Regulatory challenges and opportunities. PLoS ONE 2020, 15, e0244688. [Google Scholar] [CrossRef] [PubMed]
- Imran, M.; Anjum, F.M.; Ahmad, N.; Khan, M.K.; Mushtaq, Z.; Nadeem, M.; Hussain, S. Impact of extrusion processing conditions on lipid peroxidation and storage stability of full-fat flaxseed meal. Lipids Health Dis. 2015, 14, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Li, Y.; Cai, L.; Fang, L. Characteristics on the oxidation stability of infant formula powder with different ingredients during storage. Food Sci. Nutr. 2020, 8, 6392–6400. [Google Scholar] [CrossRef]
- Jackson, V.; Penumetcha, M. Dietary oxidised lipids, health consequences and novel food technologies that thwart food lipid oxidation: An update. Int. J. Food Sci. Technol. 2019, 54, 1981–1988. [Google Scholar] [CrossRef] [Green Version]
- Jasani, B.; Simmer, K.; Patole, S.K.; Rao, S.C. Long chain polyunsaturated fatty acid supplementation in infants born at term. Cochrane Database Syst. Rev. 2017, 3, CD000376. [Google Scholar] [CrossRef]
- Dordevic, D.; Kushkevych, I.; Jancikova, S.; Zeljkovic, S.C.; Zdarsky, M.; Hodulova, L. Modeling the effect of heat treatment on fatty acid composition in home-made olive oil preparations. Open Life Sci. 2020, 15, 606–618. [Google Scholar] [CrossRef] [PubMed]
- Koelmel, J.P.; Aristizabal-Henao, J.J.; Ni, Z.; Fedorova, M.; Kato, S.; Otoki, Y.; Nakagawa, K.; Lin, E.Z.; Godri Pollitt, K.J.; Vasiliou, V.; et al. A Novel Technique for Redox Lipidomics Using Mass Spectrometry: Application on Vegetable Oils Used to Fry Potatoes. J. Am. Soc. Mass Spectrom. 2021, 32, 1798–1809. [Google Scholar] [CrossRef] [PubMed]
- Ciavarella, C.; Motta, I.; Valente, S.; Pasquinelli, G. Pharmacological (or Synthetic) and Nutritional Agonists of PPAR-γ as Candidates for Cytokine Storm Modulation in COVID-19 Disease. Molecules 2020, 25, 2076. [Google Scholar] [CrossRef] [PubMed]
- Bansal, G.; Zhou, W.; Barlow, P.; Joshi, P.S. Review of rapid tests available for measuring the quality changes in frying oils and comparison with standard methods. Crit. Rev. Food Sci. Nutr. 2010, 50, 503–514. [Google Scholar] [CrossRef]
- Ribeiro Filho, H.V.; Bernardi Videira, N.; Bridi, A.V.; Tittanegro, T.H.; Helena Batista, F.A.; de Carvalho Pereira, J.G.; de Oliveira, P.S.L.; Bajgelman, M.C.; Le Maire, A.; Figueira, A.C.M. Screening for PPAR Non-Agonist Ligands Followed by Characterization of a Hit, AM-879, with Additional No-Adipogenic and cdk5-Mediated Phosphorylation Inhibition Properties. Front. Endocrinol. 2018, 9, 778–799. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skinner, J.; Arora, P.; McMath, N.; Penumetcha, M. Determination of Oxidized Lipids in Commonly Consumed Foods and a Preliminary Analysis of Their Binding Affinity to PPARγ. Foods 2021, 10, 1702. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10081702
Skinner J, Arora P, McMath N, Penumetcha M. Determination of Oxidized Lipids in Commonly Consumed Foods and a Preliminary Analysis of Their Binding Affinity to PPARγ. Foods. 2021; 10(8):1702. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10081702
Chicago/Turabian StyleSkinner, Joanna, Payal Arora, Nicole McMath, and Meera Penumetcha. 2021. "Determination of Oxidized Lipids in Commonly Consumed Foods and a Preliminary Analysis of Their Binding Affinity to PPARγ" Foods 10, no. 8: 1702. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10081702