
Partitioning of Proteins and Anti-Nutrients in Cassava (Manihot esculenta Crantz) Leaf Processing Fractions after Mechanical Extraction and Ultrafiltration



Abstract
:1. Introduction
2. Materials and Methods
2.1. Leaf Processing
2.2. Ultrafiltration Unit and Procedures
2.3. Mass Fractions and Dry Matter
2.4. Crude Protein (CP)
2.5. Total Phenolic Content
2.6. Amino Acid Profile
2.7. SDS-PAGE Analysis
2.8. Neutral Detergent Fiber, Acid Detergent Fiber, Acid Detergent Lignin, and Ash Content
2.9. Statistical Analysis
3. Results
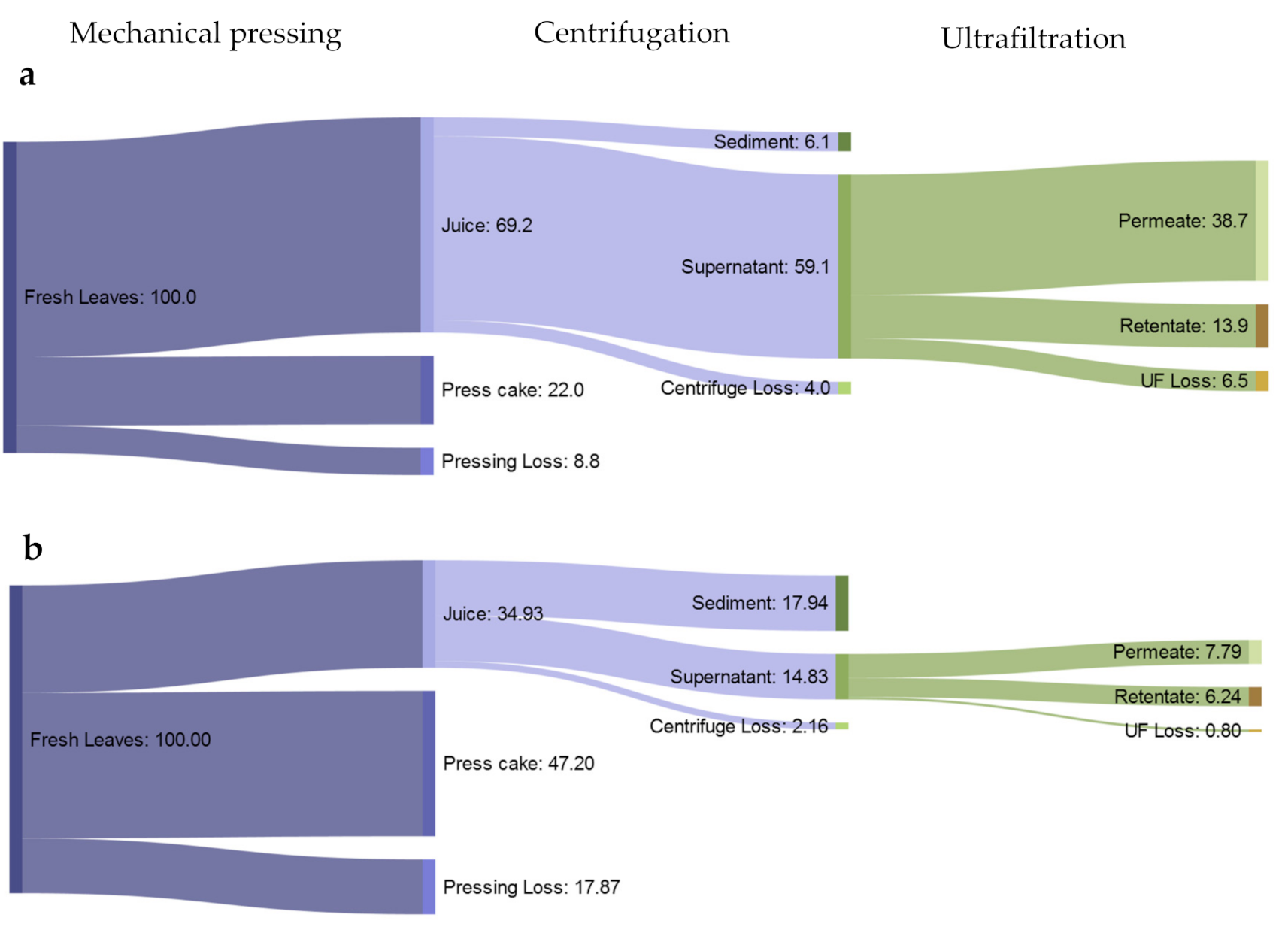
3.1. Mass Balance
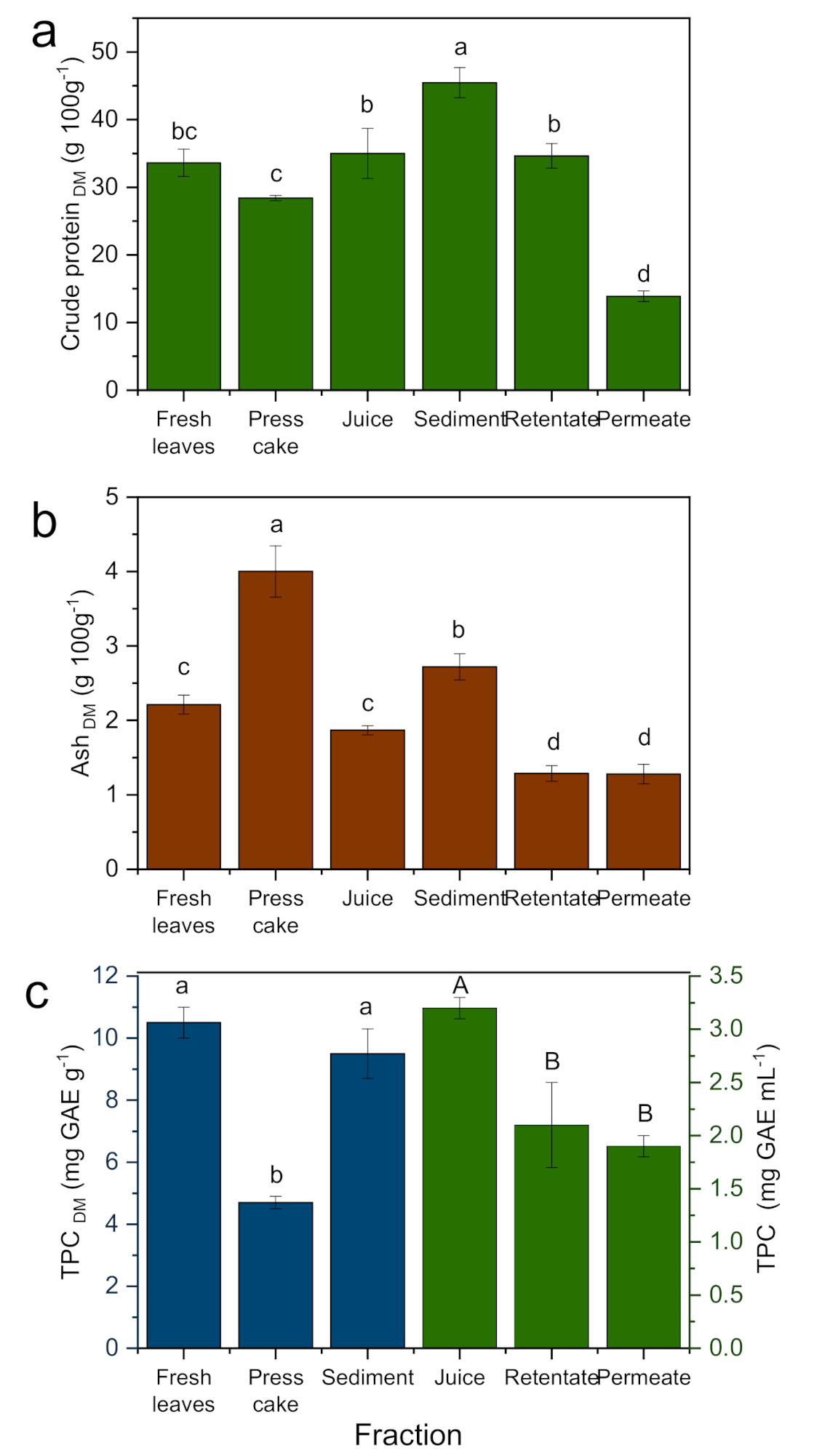
3.2. Crude Protein
3.3. Ash
3.4. Total Phenolic Content
3.5. Amino Acid Profile
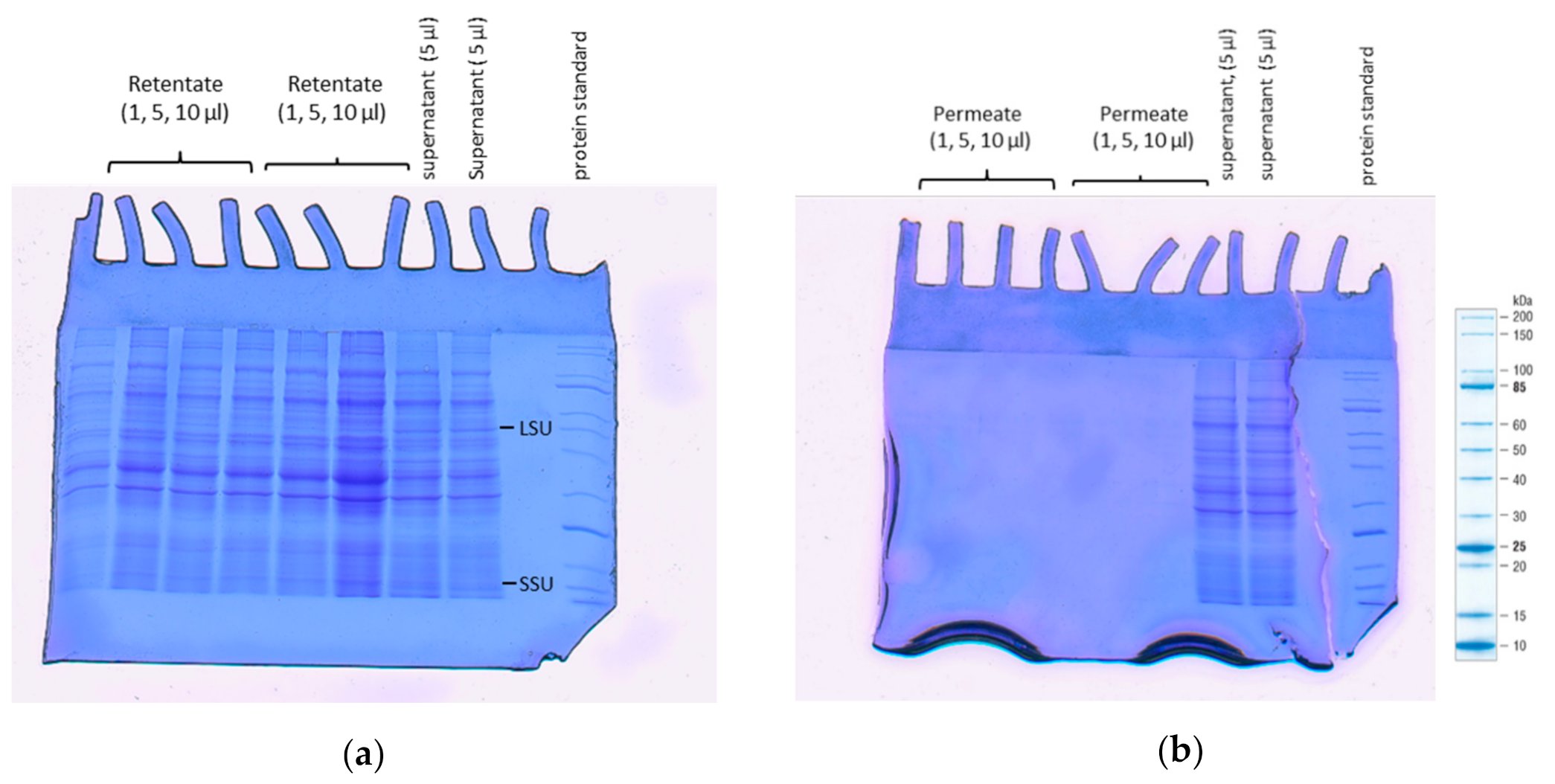
3.6. SDS-PAGE Analysis
3.7. Neutral Detergent Fiber (NDF), Acid Detergent Fiber (ADF), and Acid Detergent Lignin (ADL)
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Bouis, H.E.; Welch, R.M. Biofortification—A Sustainable Agricultural Strategy for Reducing Micronutrient Malnutrition in the Global South. Crop Sci. 2010, 50, S-20–S-32. [Google Scholar] [CrossRef] [Green Version]
- Morales, E.M.; Domingos, R.N.; Angelis, D.F. Improvement of Protein Bioavailability by Solid-State Fermentation of Babassu Mesocarp Flour and Cassava Leaves. Waste Biomass Valorization 2016, 9, 581–590. [Google Scholar] [CrossRef]
- FAOSTAT. Food and Agricultural Organization of the United Nations; FAOSTAT Database: Rome, Italy; Available online: http://faostat3.fao.org/home/E (accessed on 30 March 2021).
- Jarvis, A.; Ramirez-Villegas, J.; Campo, B.V.H.; Navarro-Racines, C. Is cassava the answer to African climate change adaptation? Trop. Plant Biol. 2012, 5, 9–29. [Google Scholar] [CrossRef]
- Achidi, A.U.; Ajayi, O.A.; Bokanga, M.; Maziya-Dixon, B. The Use of Cassava Leaves as Food in Africa. Ecol. Food Nutr. 2005, 44, 423–435. [Google Scholar] [CrossRef]
- Zhao, P.; Liu, P.; Shao, J.; Li, C.; Wang, B.; Guo, X.; Yan, B.; Xia, Y.; Peng, M. Analysis of different strategies adapted by two cassava cultivars in response to drought stress: Ensuring survival or continuing growth. J. Exp. Bot. 2015, 66, 1477–1488. [Google Scholar] [CrossRef] [Green Version]
- Latif, S.; Müller, J. Potential of cassava leaves in human nutrition: A review. Trends Food Sci. Technol. 2015, 44, 147–158. [Google Scholar] [CrossRef]
- Fasuyi, A.O. Nutrient Composition and Processing Effects on Cassava Leaf (Manihot esculenta, Crantz) Antinutrients. Pak. J. Nutr. 2005, 4, 37–42. [Google Scholar]
- Popoola, J.O.; Egwari, L.O.; Bilewu, Y.; Omonigbehin, E.; Ogunlana, O.O.; Daramola, F. Proximate analysis and SDS-PAGE protein profiling of cassava leaves: Utilization as leafy vegetable in Nigeria. MOJ Ecol. Environ. Sci. 2019, 4, 5. [Google Scholar] [CrossRef]
- Ravindran, G.; Ravindran, V. Changes in the nutritional composition of cassava (Manihot esculenta Crantz) leaves during maturity. Food Chem. 1988, 27, 299–309. [Google Scholar] [CrossRef]
- Hue, K.T.; Do Thi Thanh Van, I.L.; Wredle, E.; Sporndly, E. Effect of harvesting frequency, variety and leaf maturity on nutrient composition, hydrogen cyanide content and cassava foliage yield. Asian-Australas. J. Anim. Sci. 2012, 25, 1691–1700. [Google Scholar] [CrossRef]
- Gomez, G.; Valdivieso, M. Cassava for animal feeding: Effect of variety and plant age on production of leaves and roots. Anim. Feed Sci. Technol. 1984, 11, 49–55. [Google Scholar] [CrossRef]
- Padmaja, G. Cyanide detoxification in cassava for food and feed uses. Crit. Rev. Food Sci. Nutr. 1995, 35, 299–339. [Google Scholar] [CrossRef]
- Baiao, D.D.S.; de Freitas, C.S.; Gomes, L.P.; da Silva, D.; Correa, A.; Pereira, P.R.; Aguila, E.M.D.; Paschoalin, V.M.F. Polyphenols from Root, Tubercles and Grains Cropped in Brazil: Chemical and Nutritional Characterization and Their Effects on Human Health and Diseases. Nutrients 2017, 9, 1044. [Google Scholar] [CrossRef]
- Ravindran, V. Cassava leaves as animal feed: Potential and limitations. J. Sci. Food Agric. 1993, 61, 141–150. [Google Scholar] [CrossRef]
- Coldebella, P.F.; Gomes, S.D.; Evarini, J.A.; Cereda, M.P.; Coelho, S.R.; Coldebella, A. Evaluation of protein extraction methods to obtain protein concentrate from cassava leaf. Eng. Agrícola 2013, 33, 1223–1233. [Google Scholar] [CrossRef] [Green Version]
- Urribarrí, L.; Chacón, D.; González, O.; Ferrer, A. Protein Extraction and Enzymatic Hydrolysis of Ammonia-Treated Cassava Leaves (Manihot esculenta Crantz). Appl. Biochem. Biotechnol. 2009, 153, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Alamu, E.O.; Prisca, C.; Olaniyan, B.; Omosebi, M.O.; Adegunwa, M.O.; Chikoye, D.; Maziya-Dixon, B. Evaluation of nutritional properties, and consumer preferences of legume-fortified cassava leaves for low-income households in Zambia. Cogent Food Agric. 2021, 7, 1885796. [Google Scholar] [CrossRef]
- Montagnac, J.A.; Davis, C.R.; Tanumihardjo, S.A. Processing Techniques to Reduce Toxicity and Antinutrients of Cassava for Use as a Staple Food. Compr. Rev. Food Sci. Food Saf. 2009, 8, 17–27. [Google Scholar] [CrossRef]
- Aletor, O. Comparative, nutritive and physico-chemical evaluation of cassava (Manihot esculenta) leaf protein concentrate and fish meal. J. Food Agric. Environ. 2010, 8, 39–43. [Google Scholar]
- Bradbury, J.H.; Denton, I.C. Mild method for removal of cyanogens from cassava leaves with retention of vitamins and protein. Food Chem. 2014, 158, 417–420. [Google Scholar] [CrossRef]
- Latif, S.; Zimmermann, S.; Barati, Z.; Müller, J. Detoxification of Cassava Leaves by Thermal, Sodium Bicarbonate, Enzymatic, and Ultrasonic Treatments. J. Food Sci. 2019, 84, 1986–1991. [Google Scholar] [CrossRef] [PubMed]
- Diasolua Ngudi, D.; Kuo, Y.-H.; Lambein, F. Amino acid profiles and protein quality of cooked cassava leaves or ‘saka-saka’. J. Sci. Food Agric. 2003, 83, 529–534. [Google Scholar] [CrossRef]
- Tenorio, A.T.; Gieteling, J.; De Jong, G.A.; Boom, R.M.; Van Der Goot, A.J. Recovery of protein from green leaves: Overview of crucial steps for utilisation. Food Chem. 2016, 203, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Nynäs, A.-L. White Proteins from Green Leaves in Food Applications; Faculty of Landscape Architecture, Horticulture and Crop Production Science: Alnarp, Sweden, 2018; p. 39. [Google Scholar]
- Latif, S.; Romuli, S.; Barati, Z.; Müller, J. CFD assisted investigation of mechanical juice extraction from cassava leaves and characterization of the products. Food Sci. Nutr. 2020, 8, 3089–3098. [Google Scholar] [CrossRef] [Green Version]
- Lamsal, B.P.; Koegel, R.G.; Boettcher, M.E. Separation of protein fractions in alfalfa juice: Effects of some pre–treatment methods. Trans. ASAE 2003, 46, 715. [Google Scholar] [CrossRef]
- Tenorio, A.T. Sugar Beet Leaves for Functional Ingredients; Wageningen University: Wageningen, The Netherlands, 2017. [Google Scholar]
- Koschuh, W.; Povoden, G.; Thang, V.H.; Kromus, S.; Kulbe, K.D.; Novalin, S.; Krotscheck, C. Production of leaf protein concentrate from ryegrass (Lolium perenne × multiflorum) and alfalfa (Medicago sauva subsp. sativa). Comparison between heat coagulation/centrifugation and ultrafiltration. Desalination 2004, 163, 253–259. [Google Scholar] [CrossRef]
- Saxena, A.; Tripathi, B.P.; Kumar, M.; Shahi, V.K. Membrane-based techniques for the separation and purification of proteins: An overview. Adv. Colloid Interface Sci. 2009, 145, 1–22. [Google Scholar] [CrossRef]
- Fernández, S.S.; Menéndez, C.; Mucciarelli, S.; Padilla, A.P. Saltbush (Atriplex lampa) leaf protein concentrate by ultrafiltration for use in balanced animal feed formulations. J. Sci. Food Agric. 2007, 87, 1850–1857. [Google Scholar] [CrossRef]
- Kinsella, J.E. Leaf proteins for foods. J. Am. Oil Chem. Soc. 1979, 56, 471. [Google Scholar] [CrossRef]
- Castellanos, R.; Altamirano, S.; Moretti, R. Nutritional characteristics of cassava (Manihot esculenta Crantz) leaf protein concentrates obtained by ultrafiltration and acidic thermocoagulation. Plant Foods Hum. Nutr. 1994, 45, 357–363. [Google Scholar] [CrossRef]
- Shukla, R.; Cheryan, M. Stability and performance of ultrafiltration membranes in aqueous ethanol. Sep. Sci. Technol. 2003, 38, 1533–1547. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis of AOAC; Association of Official Analytical Chemist International: Washington, DC, USA, 1990. [Google Scholar]
- Thaipong, K.; Boonprakob, U.; Crosby, K.; Cisneros-Zevallos, L.; Hawkins Byrne, D. Comparison of ABTS, DPPH, FRAP, and ORAC assays for estimating antioxidant activity from guava fruit extracts. J. Food Compos. Anal. 2006, 19, 669–675. [Google Scholar] [CrossRef]
- Commission, E. Commission Regulation (EC) No 152/2009 of 27 January 2009 laying down the methods of sampling and analysis for the official control of feed. Off. J. Eur. 2009, L54, 130. [Google Scholar]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Colas, D.; Doumeng, C.; Pontalier, P.Y.; Rigal, L. Green crop fractionation by twin-screw extrusion: Influence of the screw profile on alfalfa (Medicago sativa) dehydration and protein extraction. Chem. Eng. Process. Process Intensif. 2013, 72, 1–9. [Google Scholar] [CrossRef]
- Goodrich, E. Protein Concentration and Diafiltration by Tangential Flow Filtration. Millipore Lit. Number TB032. 1999. Available online: http://wolfson.huji.ac.il/purification/PDF/dialysis/MILLIPORE_TFF.pdf (accessed on 15 February 2021).
- Van Reis, R.; Goodrich, E.; Yson, C.; Frautschy, L.; Whiteley, R.; Zydney, A. Constant Cwall ultrafiltration process control. J. Membr. Sci. 1997, 130, 123–140. [Google Scholar] [CrossRef]
- Teo, C.R.P.A.; Prudencio, S.H.; Coelho, S.R.M.; Teo, M.S. Obtenção e caracterização físico-química de concentrado protéico de folhas de mandioca. Rev. Bras. Eng. Agrícola E Ambient. 2010, 14, 993–999. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Grimi, N.; Jaffrin, M.Y.; Ding, L. Leaf protein concentration of alfalfa juice by membrane technology. J. Membr. Sci. 2015, 489, 183–193. [Google Scholar] [CrossRef]
- Oresegun, A.; Fagbenro, O.A.; Ilona, P.; Bernard, E.; Yildiz, F. Nutritional and anti-nutritional composition of cassava leaf protein concentrate from six cassava varieties for use in aqua feed. Cogent Food Agric. 2016, 2. [Google Scholar] [CrossRef]
- Aletor, O.; Oshodi, A.A.; Ipinmoroti, K. Chemical composition of common leafy vegetables and functional properties of their leaf protein concentrates. Food Chem. 2002, 78, 63–68. [Google Scholar] [CrossRef]
- Achidi, A.U.; Ajayi, O.A.; Maziya-Dixon, B.; Bokanga, M. The Effect of Processing on the Nutrient Content of Cassava (Manihot Esculenta Crantz) Leaves. J. Food Process. Preserv. 2008, 32, 486–502. [Google Scholar] [CrossRef]
- Nur, F.O.; Siti, A.H.; Umi, K.Y. Comparative evaluation of organic and inorganic fertilizers on total phenolic, total flavonoid, antioxidant activity and cyanogenic glycosides in cassava (Manihot esculenta). Afr. J. Biotechnol. 2013, 12, 2414–2421. [Google Scholar]
- Cirkovic Velickovic, T.D.; Stanic-Vucinic, D.J. The role of dietary phenolic compounds in protein digestion and processing technologies to improve their antinutritive properties. Compr. Rev. Food Sci. Food Saf. 2018, 17, 82–103. [Google Scholar] [CrossRef] [Green Version]
- Abdelnour, S.; Abd El-Hack, M.; Ragni, M. The Efficacy of High-Protein Tropical Forages as Alternative Protein Sourcesfor Chickens: A Review. Agriculture 2018, 8, 86. [Google Scholar] [CrossRef] [Green Version]
- Díaz-Reinoso, B.; Moure, A.; Domínguez, H.; Parajó, J.C. Membrane concentration of antioxidants from Castanea sativa leaves aqueous extracts. Chem. Eng. J. 2011, 175, 95–102. [Google Scholar] [CrossRef]
- Hasim, S.F.; Lia Kusuma, D. Effect of Boiled Cassava Leaves (Manihot esculenta Crantz) on Total Phenolic, Flavonoid and its Antioxidant Activity. Curr. Biochem. 2016, 3, 12. [Google Scholar]
- Eggum, B.O. The protein quality of cassava leaves. Br. J. Nutr. 1970, 24, 761–768. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. The United Nations University. Protein and Amino Acid Requirements in Human Nutrition: Report of a Joint FAO/WHO/UNU Expert Consultation; World Health Organization: Geneva, Switzerland, 2007; Volume 935. [Google Scholar]
- Mariotti, F.; Tomé, D.; Mirand, P.P. Converting Nitrogen into Protein—Beyond 6.25 and Jones’ Factors. Crit. Rev. Food Sci. Nutr. 2008, 48, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Oni, A.O.; Arigbede, O.M.; Oni, O.O.; Onwuka, C.F.I.; Anele, U.Y.; Oduguwa, B.O.; Yusuf, K.O. Effects of feeding different levels of dried cassava leaves (Manihot esculenta, Crantz) based concentrates with Panicum maximum basal on the performance of growing West African Dwarf goats. Livest. Sci. 2010, 129, 24–30. [Google Scholar] [CrossRef]
- Wargiono, J.; Sudaryanto, B. Cassava leaves and forage crops for ruminant feed in the establishment of sustainable cassava farming system in Indonesia. In Proceedings of the National Workshop-Seminar on Sustainable Livestock Production on Local Feed Resources, Ho Chi Minh City, Vietnam, 18–20 January 2000; pp. 496–503. [Google Scholar]
- Verheyden-Tixier, H.; Renaud, P.-C.; Morellet, N.; Jamot, J.; Besle, J.-M.; Dumont, B. Selection for nutrients by red deer hinds feeding on a mixed forest edge. Oecologia 2008, 156, 715–726. [Google Scholar] [CrossRef]
- Cherdthong, A.; Pornjantuek, B.; Wachirapakorn, C. Effect of feeding cassava bioethanol waste on nutrient intake, digestibility, and rumen fermentation in growing goats. Trop. Anim. Health Prod. 2016, 48, 1369–1374. [Google Scholar] [CrossRef]
- Karri, V.R.; Nalluri, N. Cassava: Meeting the global protein need. Plant Sci. Today 2016, 3, 304–311. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amino Acid | Fractions | Recommended Amount * | |||||
|---|---|---|---|---|---|---|---|
| Fresh Leaves | Press Cake | Juice | Sediment | Retentate | Permeate | ||
| Tryptophan | 2.0 ± 0.1 a | 2.2 ± 0.1 a | 0.5 ± 0.0 bc | 2.5 ± 0.3 a | 0.7 ± 0.1 b | 0.0 c | 0.6 |
| Threonine | 4.5 ± 0.2 a | 4.7 ± 0.0 a | 4.8 ± 0.3 a | 4.8 ± 0.3 a | 5.2 ± 0.6 a | 1.4 ± 0.0 b | 2.3 |
| Valine | 5.7 ± 0.3 a | 6.1 ± 0.3 a | 5.8 ± 0.2 a | 6.3 ± 0.0 a | 6.5 ± 0.0 a | 2.1 ± 0.2 b | 3.9 |
| Methionine | 1.9 ± 0.1 ab | 1.9 ± 0.1 ab | 1.4 ± 0.0 bc | 2.1 ± 0.0 a | 1.1 ± 0.3 c | 0.0 d | 1.6 |
| Isoleucine | 4.7 ± 0.2 a | 5.2 ± 0.3 a | 4.6 ± 0.2 a | 5.3 ± 0.0 a | 5.0 ± 0.6 a | 1.4 ± 0.0 b | 3.0 |
| Leucine | 8.9 ± 0.2 ab | 9.9 ± 0.3 ab | 8.9 ± 0.2 ab | 10.3 ± 0.2 a | 8.3 ± 1.1 b | 1.4 ± 0.0 c | 5.9 |
| Phenylalanine | 5.8 ± 0.4 bc | 6.7 ± 0.1 ab | 5.8 ± 0.2 bc | 7.1 ± 0.1 a | 5.0 ± 0.6 c | 1.4 ± 0.0 d | N/A |
| Histidine | 2.2 ± 0.1 a | 2.2 ± 0.1 a | 2.1 ± 0.0 a | 2.2 ± 0.0 a | 1.8 ± 0.0 b | 0.0 c | 1.5 |
| Lysine | 6.3 ± 0.3 a | 6.4 ± 0.3 a | 6.5 ± 0.2 a | 6.4 ± 0.1 a | 7.9 ± 1.0 a | 2.8 ± 0.0 b | 4.5 |
| Aspartic acid | 10.1 ± 0.3 a | 9.7 ± 0.3 ab | 10.2 ± 0.3 a | 10.1 ± 0.5 a | 11.4 ± 0.4 a | 5.6 ± 2.0 b | N/A |
| Glutamic acid | 12.6 ± 0.3 a | 11.5 ± 0.4 a | 12.6 ± 0.3 a | 12.1 ± 0.3 a | 14.2 ± 1.6 a | 7.1 ± 2.0 b | N/A |
| Alanine | 6.6 ± 0.2 a | 6.9 ± 0.2 a | 7.0 ± 0.2 a | 7.1 ± 0.2 a | 7.4 ± 1.0 a | 3.5 ± 1.0 b | N/A |
| Tyrosine | 3.5 ± 0.3 a | 3.6 ± 0.1 a | 4.3 ± 0.0 a | 4.1 ± 0.1 a | 4.1 ± 0.6 a | 1.4 ± 0.0 b | N/A |
| Serine | 4.2 ± 0.0 a | 4.4 ± 0.1 a | 4.4 ± 0.2 a | 4.3 ± 0.3 a | 5.4 ± 0.6 a | 1.4 ± 0.0 b | N/A |
| Glycine | 5.8 ± 0.2 a | 6.2 ± 0.2 a | 6.1 ± 0.2 a | 6.5 ± 0.2 a | 5.9 ± 0.6 a | 2.1 ± 0.1 b | N/A |
| Cysteine | 0.8 ± 0.0 d | 0.8 ± 0.0 d | 1.2 ± 0.0 c | 0.7 ± 0.0 d | 1.8 ± 0.0 a | 1.4 ± 0.0 b | 0.6 |
| Arginine | 5.9 ± 0.2 ab | 5.7 ± 0.1 ab | 6.1 ± 0.2 ab | 6.5 ± 0.2 a | 5.0 ± 0.2 b | 1.4 ± 0.0 c | N/A |
| Proline | 5.1 ± 0.1 a | 5.7 ± 0.0 a | 5.0 ± 0.0 a | 5.5 ± 0.1 a | 5.0 ± 0.2 a | 1.4 ± 0.0 b | N/A |
| AAA | 9.2 | 10.3 | 10.1 | 11.2 | 9.2 | 2.8 | 3.8 |
| Total amino acid | 96.4 | 99.6 | 97.3 | 103.8 | 101.5 | 36.0 | N/A |
| Fractions | NDF (g 100 g−1DM) | ADF (g 100 g−1DM) | ADL (g 100 g−1DM) |
|---|---|---|---|
| Fresh leaves | 19.5 ± 3.0 b | 13.8 ± 0.3 b | 1.4 ±0.1 b |
| Press cake | 28.0 ± 0.5 a | 25.3 ± 0.2 a | 4.0 ± 0.4 a |
| Juice | 9.3 ± 2.1 c | 1.0 ± 0.0 c | 0.0 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ayele, H.H.; Latif, S.; Bruins, M.E.; Müller, J. Partitioning of Proteins and Anti-Nutrients in Cassava (Manihot esculenta Crantz) Leaf Processing Fractions after Mechanical Extraction and Ultrafiltration. Foods 2021, 10, 1714. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10081714
Ayele HH, Latif S, Bruins ME, Müller J. Partitioning of Proteins and Anti-Nutrients in Cassava (Manihot esculenta Crantz) Leaf Processing Fractions after Mechanical Extraction and Ultrafiltration. Foods. 2021; 10(8):1714. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10081714
Chicago/Turabian StyleAyele, Haimanot Hailegiorigs, Sajid Latif, Marieke E. Bruins, and Joachim Müller. 2021. "Partitioning of Proteins and Anti-Nutrients in Cassava (Manihot esculenta Crantz) Leaf Processing Fractions after Mechanical Extraction and Ultrafiltration" Foods 10, no. 8: 1714. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10081714