
A Response Surface Methodology (RSM) Approach for Optimizing the Attenuation of Human IgE-Reactivity to β-Lactoglobulin (β-Lg) by Hydrostatic High Pressure Processing


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Instrument
2.3. HHP Treatments
2.4. Experimental Design and Data Analysis
2.5. Determination of Optimal Operating Conditions and Validation of the Model
2.6. The Bicinchoninic Acid (BCA) Protein Assay
2.7. SDS-PAGE
2.8. Enzyme-Linked Immuno-Sorbent Assay (ELISA)
2.9. Spectral Measurements
2.10. Dot Blotting with Rabbit IgG and Human IgE
2.11. Statistical Analysis
3. Results and Discussion
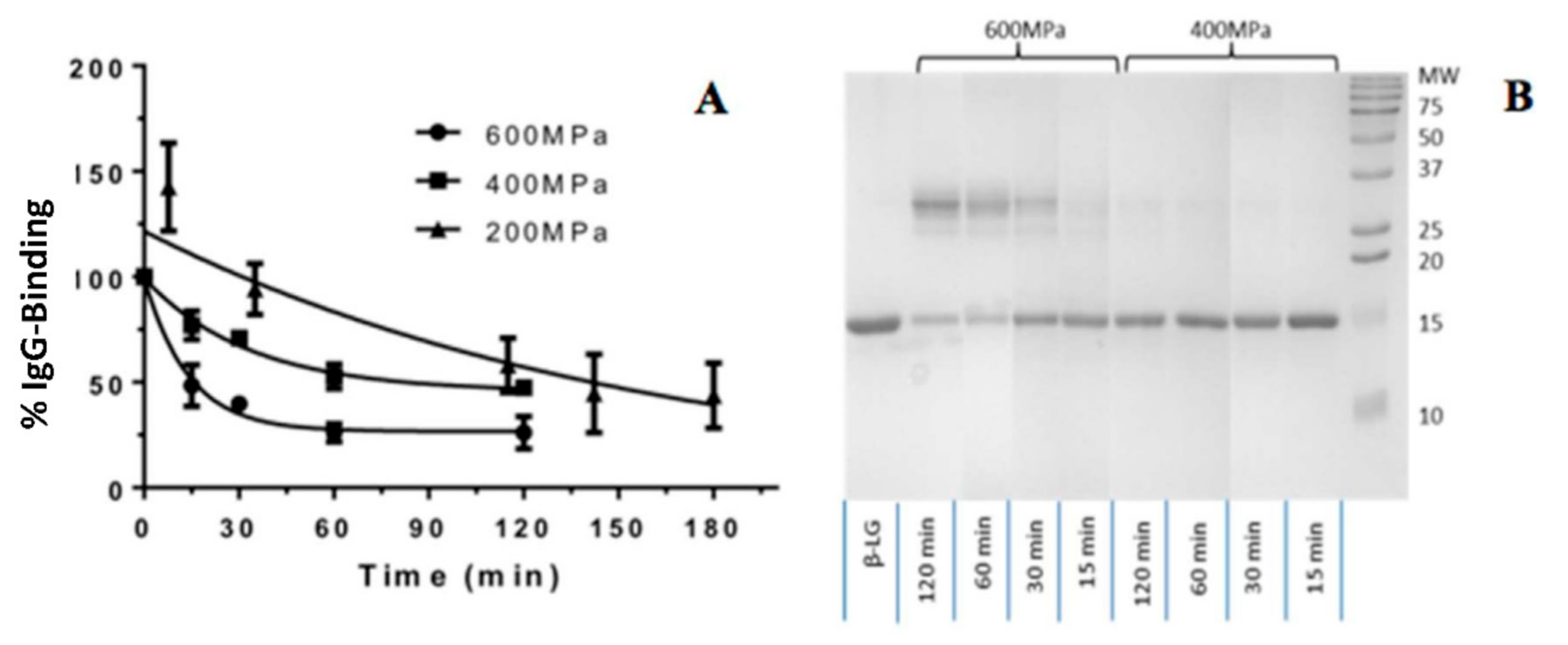
3.1. Preliminary Study of Pressure, Holding Time and IgG Binding
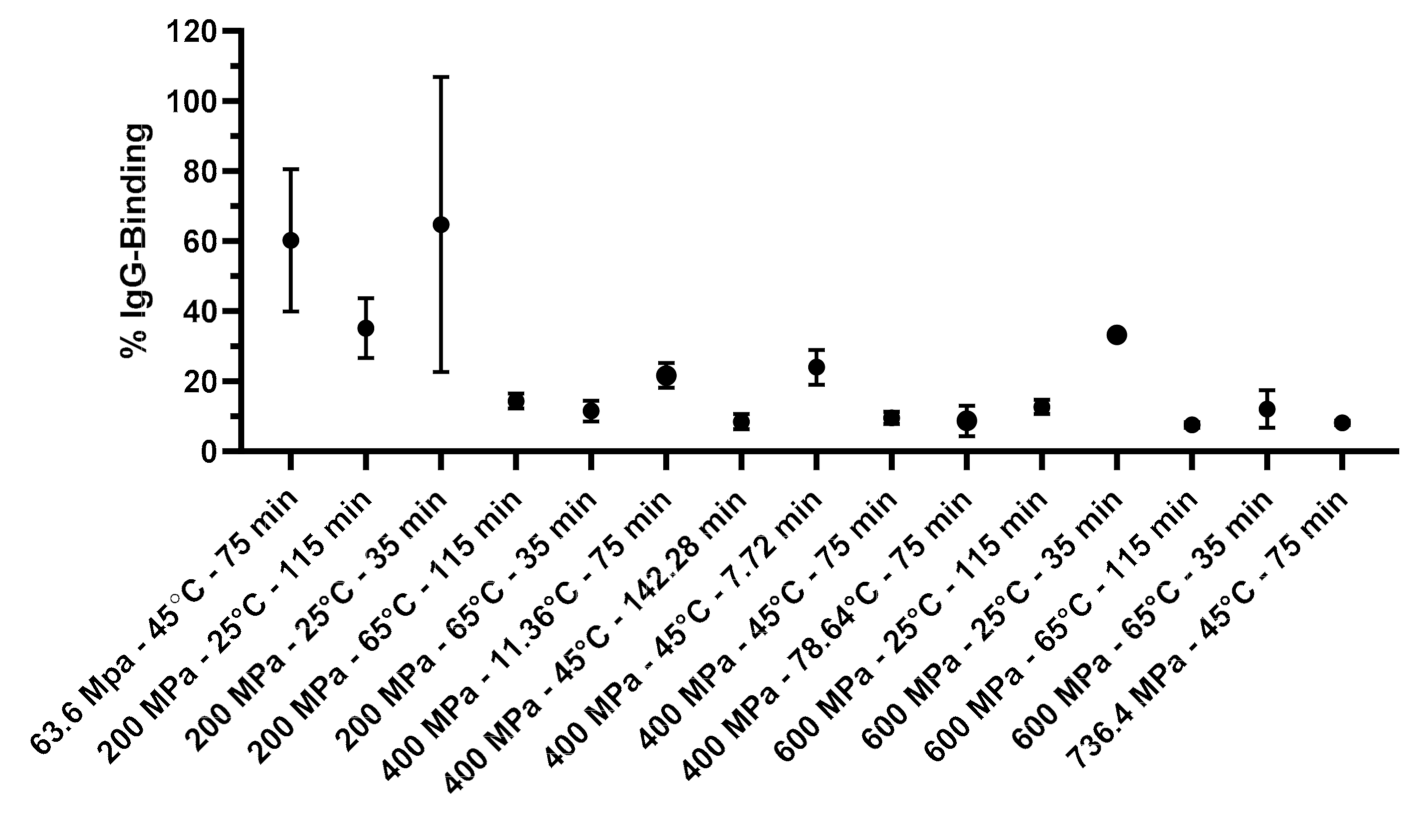
3.2. Effects of Process Variables on the Molecular Changes of β-Lactoglobulin Determined by IgG Binding
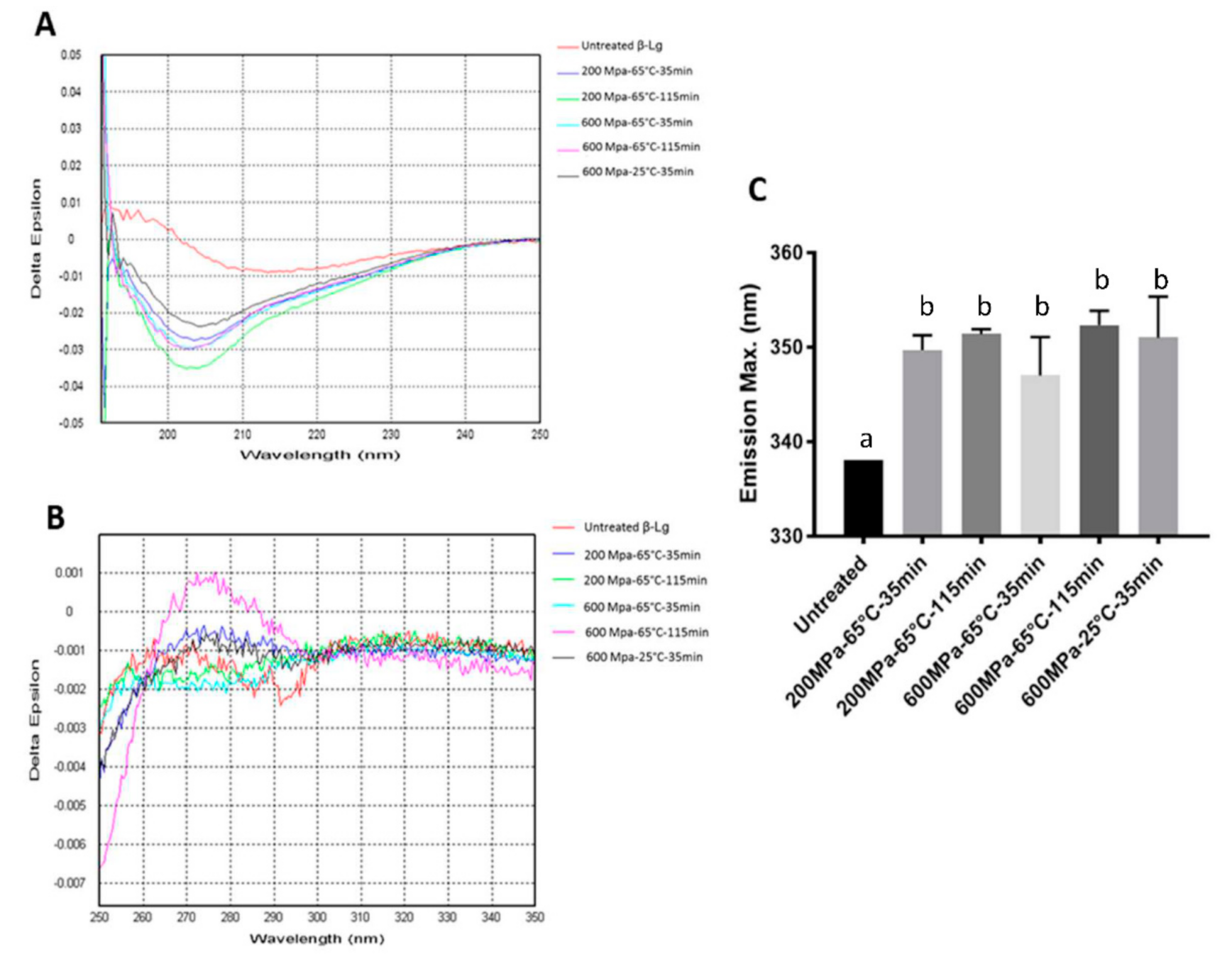
3.3. The CD and Intrinsic Fluorescence Analyses of β-Lg Treated with Selected HHP Conditions
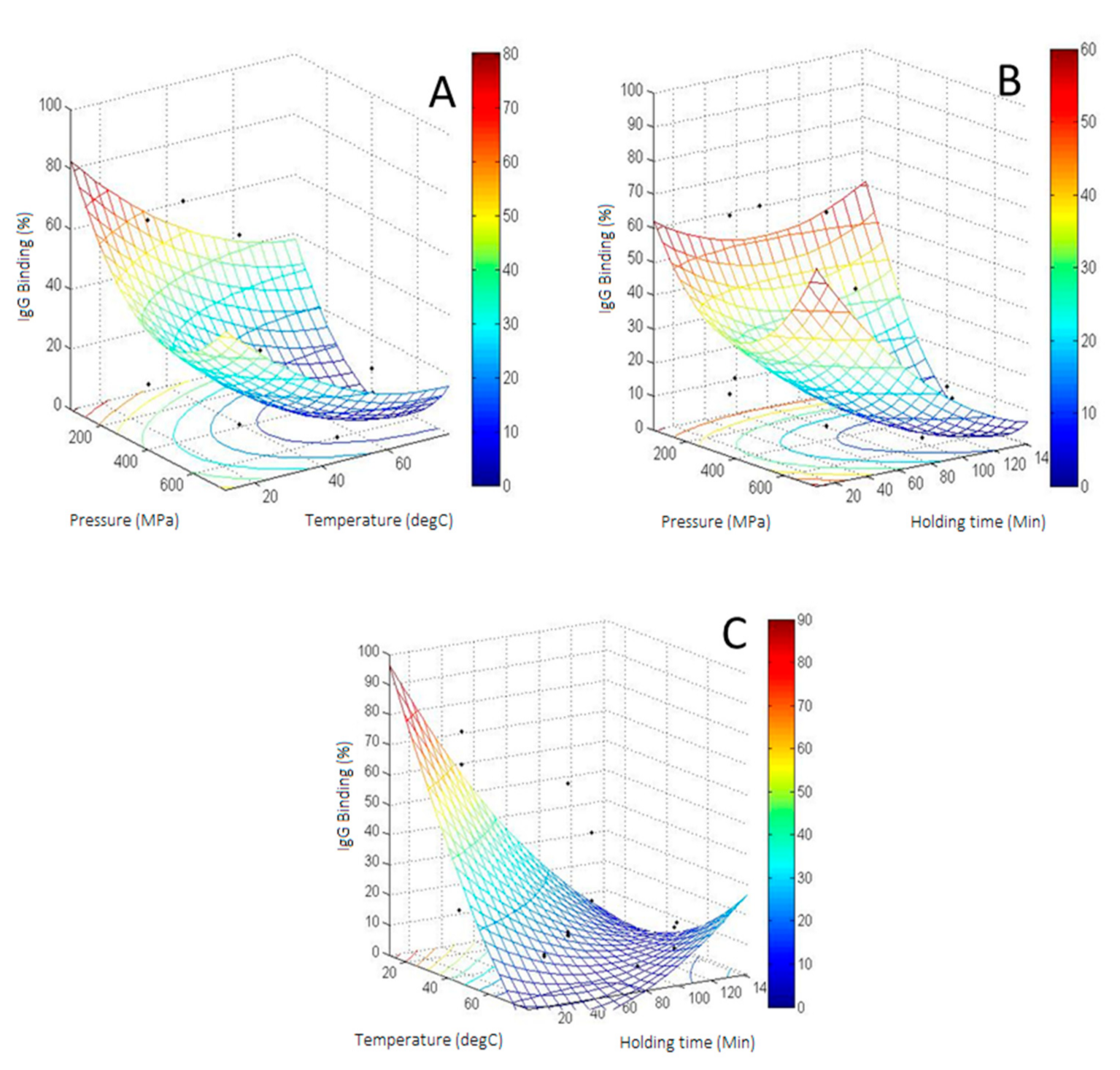
3.4. CCD of IgG Binding of HHP Treated β-Lactoglobulin
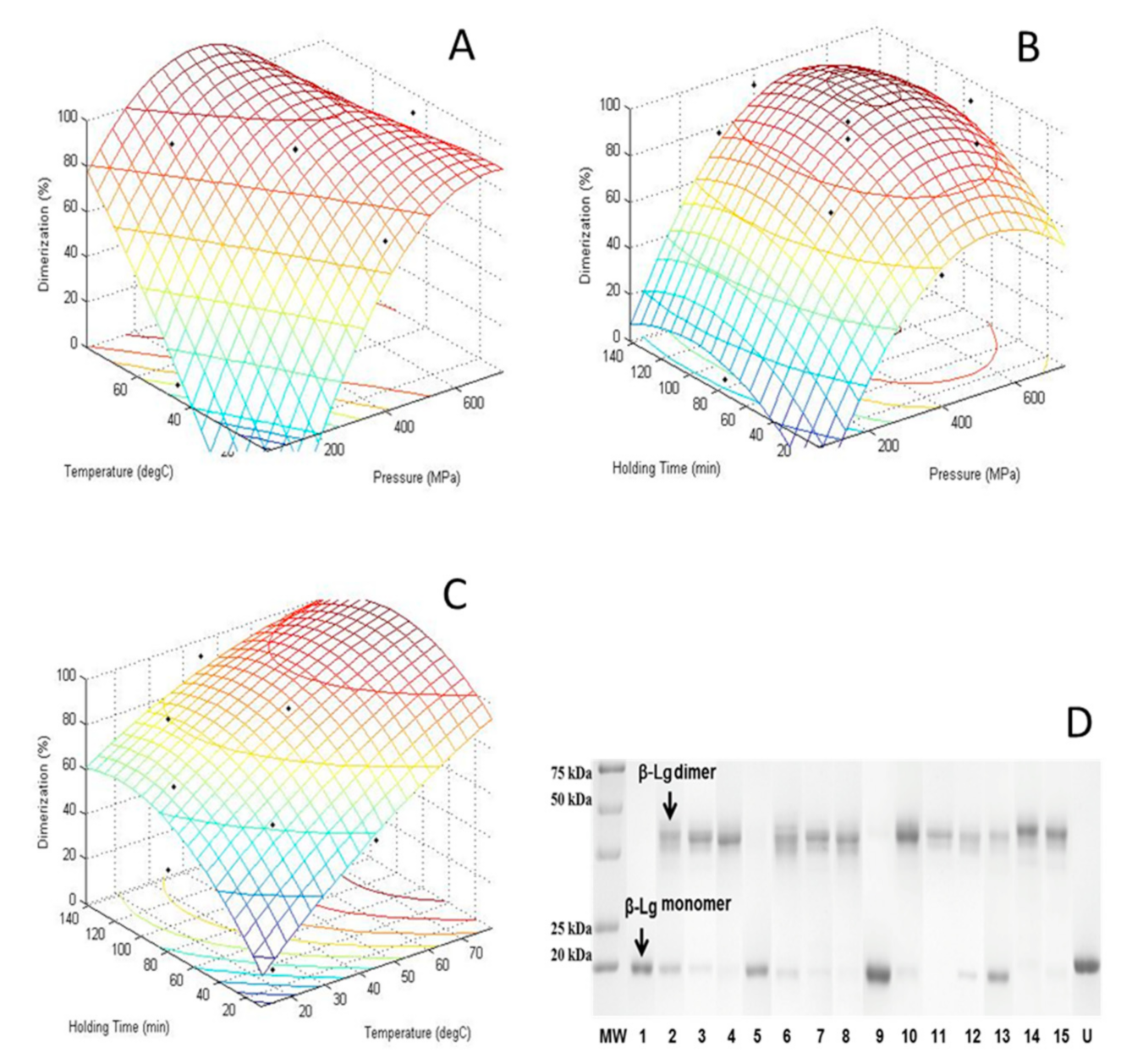
3.5. CCD of Dimerization of HHP Treated β-Lg
3.6. Relationship between IgG Binding and Dimerization
3.7. Determination and Validation of Calculated Optimum Response
3.8. Effects of Thermal and HHP Treatments on α-La
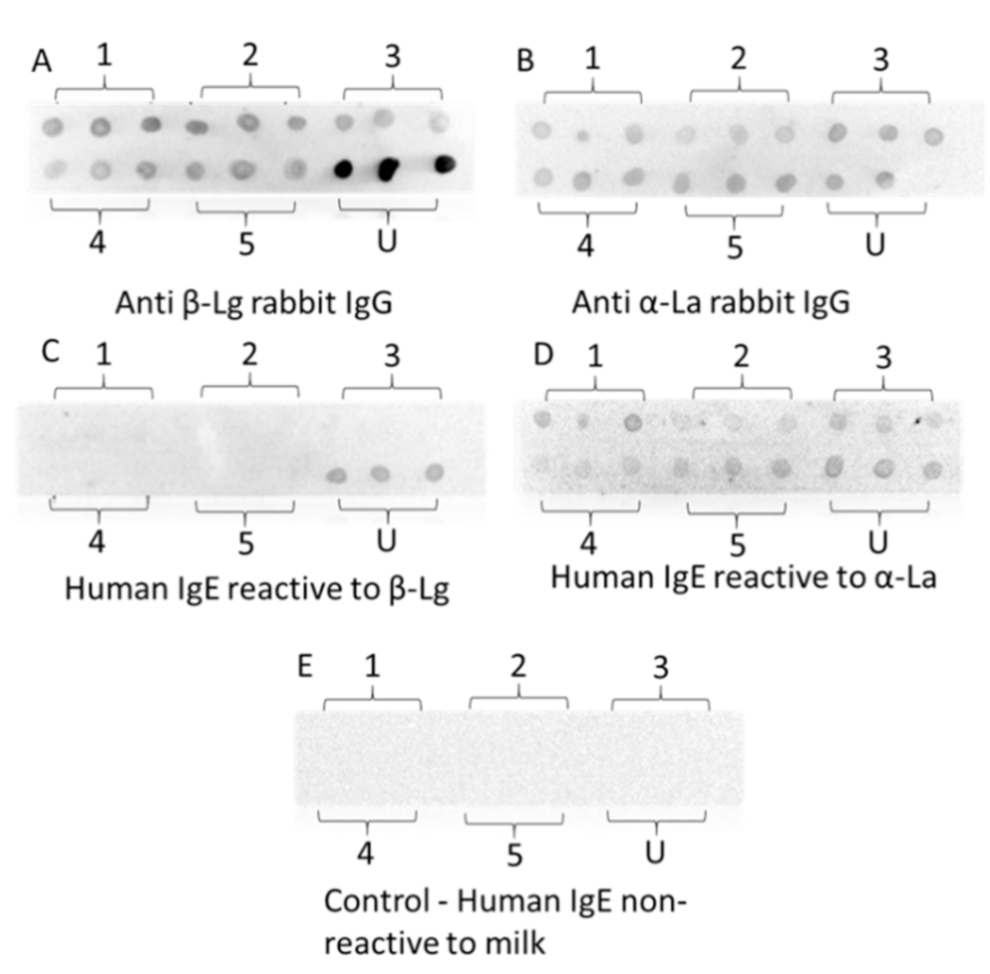
3.9. Dot-Blotting with Rabbit IgG and Human IgE
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Mullins, R.J.; Wainstein, B.K.; Barnes, E.H.; Liew, W.K.; Campbell, D.E. Increases in anaphylaxis fatalities in Australia from 1997 to 2013. Clin. Exp. Allergy 2016, 46, 1099–1110. [Google Scholar] [CrossRef]
- Schmidt, D.; Meijer, R.; Slangen, C.; Van Beresteijn, E. Raising the pH of the pepsin-catalysed hydrolysis of bovine whey proteins increases the antigenicity of the hydrolysates. Clin. Exp. Allergy 1995, 25, 1007–1017. [Google Scholar] [CrossRef] [PubMed]
- Mills, S.; Ross, R.; Hill, C.; Fitzgerald, G.; Stanton, C. Milk intelligence: Mining milk for bioactive substances associated with human health. Int. Dairy J. 2011, 21, 377–401. [Google Scholar] [CrossRef]
- Fox, A.T.; Thomson, M. Adverse reactions to cows’ milk. Paediatr. Child Health 2007, 17, 288–294. [Google Scholar] [CrossRef]
- Breiteneder, H.; Mills, E.C. Molecular properties of food allergens. J. Allergy Clin. Immunol. 2005, 115, 14–23. [Google Scholar] [CrossRef]
- Wal, J. Structure and function of milk allergens. Allergy 2001, 56, 35–38. [Google Scholar] [CrossRef]
- Hinrichs, J.; Rademacher, B. Kinetics of combined thermal and pressure-induced whey protein denaturation in bovine skim milk. Int. Dairy J. 2005, 15, 315–323. [Google Scholar] [CrossRef]
- Maier, I.; Okun, V.M.; Pittner, F.; Lindner, W. Changes in peptic digestibility of bovine β-lactoglobulin as a result of food processing studied by capillary electrophoresis and immunochemical methods. J. Chromatogr. B. 2006, 841, 160–167. [Google Scholar] [CrossRef]
- Metcalfe, D.D.; Astwood, J.D.; Townsend, R.; Sampson, H.A.; Taylor, S.L.; Fuchs, R.L. Assessment of the allergenic potential of foods derived from genetically engineered crop plants. Crit. Rev. Food Sci. Nutr. 1996, 36, 165–186. [Google Scholar] [CrossRef] [PubMed]
- Mozhaev, V.V.; Heremans, K.; Frank, J.; Masson, P.; Balny, C. High pressure effects on protein structure and function. Proteins Struct. Funct. Bioinform. 1996, 24, 81–91. [Google Scholar] [CrossRef]
- Silva, J.L.; Weber, G. Pressure stability of proteins. Annu. Rev. Phys. Chem. 1993, 44, 89–113. [Google Scholar] [CrossRef]
- Balny, C.; Masson, P. Effects of high pressure on proteins. Food Rev. Int. 1993, 9, 611–628. [Google Scholar] [CrossRef]
- Considine, T.; Patel, H.A.; Singh, H.; Creamer, L.K. Influence of binding conjugated linoleic acid and myristic acid on the heat-and high-pressure-induced unfolding and aggregation of β-lactoglobulin B. Food Chem. 2007, 102, 1270–1280. [Google Scholar] [CrossRef]
- Tolgyesi, F.; Bode, C.; Smeller, L.; Kim, D.; Kim, K.; Heremans, K.; Fidy, J. Pressure activation of the chaperone function of small heat shock proteins. Cell. Mol. Biol. 2004, 50, 361–370. [Google Scholar]
- Hajós, G.; Polgár, M.; Farkas, J. High-pressure effects on IgE immunoreactivity of proteins in a sausage batter. Innov. Food Sci. Emerg. Technol. 2004, 5, 443–449. [Google Scholar] [CrossRef]
- Dhakal, S.; Liu, C.; Zhang, Y.; Roux, K.H.; Sathe, S.K.; Balasubramaniam, V.M. Effect of high pressure processing on the immunoreactivity of almond milk. Food Res. Int. 2014, 62, 215–222. [Google Scholar] [CrossRef]
- Kleber, N.; Maier, S.; Hinrichs, J. Antigenic response of bovine β-lactoglobulin influenced by ultra-high pressure treatment and temperature. Innov. Food Sci. Emerg. Technol. 2007, 8, 39–45. [Google Scholar] [CrossRef]
- Cuadrado, C.; Sanchiz, A.; Vicente, F.; Ballesteros, I.; Linacero, R. Changes Induced by Pressure Processing on Immunoreactive Proteins of Tree Nuts. Molecules 2020, 25, 954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Yang, W.; Chung, S.Y.; Chen, H.; Ye, M.; Teixeira, A.A.; Gregory, J.F.; Welt, B.A.; Shriver, S. Effect of pulsed ultraviolet light and high hydrostatic pressure on the antigenicity of almond protein extracts. Food Bioprocess. Technol. 2013, 6, 431–440. [Google Scholar] [CrossRef]
- Husband, F.A.; Aldick, T.; Van der Plancken, I.; Grauwet, T.; Hendrickx, M.; Skypala, I.; Mackie, A.R. High-pressure treatment reduces the immunoreactivity of the major allergens in apple and celeriac. Mol. Nutr. Food Res. 2011, 55, 1087–1095. [Google Scholar] [CrossRef] [Green Version]
- Johnson, P.E.; Van der Plancken, I.; Balasa, A.; Husband, F.A.; Grauwet, T.; Hendrickx, M.; Knorr, D.; Mills, E.N.; Mackie, A.R. High pressure, thermal and pulsed electric-field-induced structural changes in selected food allergens. Mol. Nutr. Food Res. 2010, 54, 1701–1710. [Google Scholar] [CrossRef]
- Somkuti, J.; Bublin, M.; Breiteneder, H.; Smeller, L. Pressure—temperature stability, Ca2+ binding, and pressure—temperature phase diagram of cod parvalbumin: Gad m 1. Biochemistry 2012, 51, 5903–5911. [Google Scholar] [CrossRef]
- Liu, C.; Tao, S.; Liu, R.; Chen, F.; Xue, W. Is high pressure treatment able to modify the allergenicity of the largemouth bass allergens? High. Press. Res. 2021, 32, 551–556. [Google Scholar] [CrossRef]
- López-Fandiño, R. Functional improvement of milk whey proteins induced by high hydrostatic pressure. Crit. Rev. Food Sci. Nutr. 2006, 46, 351–363. [Google Scholar] [CrossRef]
- Rivalain, N.; Roquain, J.; Demazeau, G. Development of high hydrostatic pressure in biosciences: Pressure effect on biological structures and potential applications in biotechnologies. Biotechnol. Adv. 2010, 28, 659–672. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Namkung, H.; Ahn, H.; Paik, I. Production of iron enriched eggs of laying hens. Asian-Australas. J. Anim. Sci. 2004, 17, 1725–1728. [Google Scholar] [CrossRef]
- De Maria, S.; Ferrari, G.; Maresca, P. Effect of high hydrostatic pressure on the enzymatic hydrolysis of bovine serum albumin. J. Sci. Food Agric. 2017, 97, 3151–3158. [Google Scholar] [CrossRef] [PubMed]
- Chicón, R.; López-Fandiño, R.; Alonso, E.; Belloque, J. Proteolytic pattern, antigenicity, and serum immunoglobulin E binding of β-lactoglobulin hydrolysates obtained by pepsin and high-pressure treatments. J. Dairy Sci. 2008, 91, 928–938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boukil, A.; Suwal, S.; Chamberland, J.; Pouliot, Y.; Doyen, A. Ultrafiltration performance and recovery of bioactive peptides after fractionation of tryptic hydrolysate generated from pressure-treated β-lactoglobulin. J. Membr. Sci. 2018, 556, 42–53. [Google Scholar] [CrossRef]
- Olsen, K.; Kristiansen, K.R.; Skibsted, L.H. Effect of high hydrostatic pressure on the steady-state kinetics of tryptic hydrolysis of β-lactoglobulin. Food Chem. 2003, 80, 255–260. [Google Scholar] [CrossRef]
- Zeece, M.; Huppertz, T.; Kelly, A. Effect of high-pressure treatment on in-vitro digestibility of β-lactoglobulin. Innov. Food Sci. Emerg. Technol. 2008, 9, 62–69. [Google Scholar] [CrossRef]
- Izquierdo, F.J.; Alli, I.; Gómez, R.; Ramaswamy, H.S.; Yaylayan, V. Effects of high pressure and microwave on pronase and α-chymotrypsin hydrolysis of β-lactoglobulin. Food Chem. 2005, 92, 713–719. [Google Scholar] [CrossRef]
- Blayo, C.; Vidcoq, O.; Lazennec, F.; Dumay, E. Effects of high pressure processing (hydrostatic high pressure and ultra-high pressure homogenisation) on whey protein native state and susceptibility to tryptic hydrolysis at atmospheric pressure. Food Res. Int. 2016, 79, 40–53. [Google Scholar] [CrossRef]
- Beran, M.; Klubal, R.; Molik, P.; Strohalm, J.; Urban, M.; Klaudyova, A.A.; Prajzlerova, K. Influence of high-hydrostatic pressure on tryptic and chymotryptic hydrolysis of milk proteins. High. Press. Res. 2009, 29, 23–27. [Google Scholar] [CrossRef]
- Bamdad, F.; Shin, S.H.; Suh, J.-W.; Nimalaratne, C.; Sunwoo, H. Anti-inflammatory and antioxidant properties of casein hydrolysate produced using high hydrostatic pressure combined with proteolytic enzymes. Molecules 2017, 22, 609. [Google Scholar] [CrossRef]
- Knudsen, J.; Otte, J.; Olsen, K.; Skibsted, L. Effect of high hydrostatic pressure on the conformation of β-lactoglobulin A as assessed by proteolytic peptide profiling. Int. Dairy J. 2002, 12, 791–803. [Google Scholar] [CrossRef]
- Maynard, F.; Weingand, A.; Hau, J.; Jost, R. Effect of high-pressure treatment on the tryptic hydrolysis of bovine β-lactoglobulin AB. Int. Dairy J. 1998, 8, 125–133. [Google Scholar] [CrossRef]
- Lozano-Ojalvo, D.; Pérez-Rodríguez, L.; Pablos-Tanarro, A.; López-Fandiño, R.; Molina, E. Pepsin treatment of whey proteins under high pressure produces hypoallergenic hydrolysates. Innov. Food Sci. Emerg. Technol. 2017, 43, 154–162. [Google Scholar] [CrossRef]
- Marciniak, A.; Suwal, S.; Brisson, G.; Britten, M.; Pouliot, Y.; Doyen, A.J.F.H. Studying a chaperone-like effect of beta-casein on pressure-induced aggregation of beta-lactoglobulin in the presence of alpha-lactalbumin. Food Hydrocoll. 2018, 84, 9–15. [Google Scholar] [CrossRef]
- Zhao, Y.; Sun, X.; Marquis, C.P.; Lee, N.A. Development of a sensitive sandwich ELISA specific to 2S albumin (Ana o 3) as a stable protein marker for cashew nut residue detection in pre-packaged food products. Food Control 2019, 96, 432–440. [Google Scholar] [CrossRef]
- Yang, J.; Dunker, A.K.; Powers, J.R.; Clark, S.; Swanson, B.G. β-Lactoglobulin molten globule induced by high pressure. J. Agric. Food Chem. 2001, 49, 3236–3243. [Google Scholar] [CrossRef]
- Karamonová, L.; Fukal, L.; Kodíček, M.; Rauch, P.; Mills, E.C.; Morgan 1, M.R. Immunoprobes for thermally-induced alterations in whey protein structure and their application to the analysis of thermally-treated milks. Food Agric. Immunol. 2003, 15, 77–91. [Google Scholar] [CrossRef]
- Manderson, G.A.; Creamer, L.K.; Hardman, M.J. Effect of heat treatment on the circular dichroism spectra of bovine β-lactoglobulin A, B, and C. J. Agric. Food Chem. 1999, 47, 4557–4567. [Google Scholar] [CrossRef]
- Meng, X.; Bai, Y.; Gao, J.; Li, X.; Chen, H. Effects of high hydrostatic pressure on the structure and potential allergenicity of the major allergen bovine β-lactoglobulin. Food Chem. 2017, 219, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Svenning, C.; Brynhildsvold, J.; Molland, T.; Langsrud, T.; Vegarud, G.E. Antigenic response of whey proteins and genetic variants of β-lactoglobulin–The effect of proteolysis and processing. Int. Dairy J. 2000, 10, 699–711. [Google Scholar] [CrossRef]
- Considine, T.; Patel, H.A.; Anema, S.G.; Singh, H.; Creamer, L.K. Interactions of milk proteins during heat and high hydrostatic pressure treatments–A Review. Innov. Food Sci. Emerg. Technol. 2007, 8, 1–23. [Google Scholar]
- Clement, G.; Boquet, D.; Frobert, Y.; Bernard, H.; Negroni, L.; Chatel, J.-M.; Adel-Patient, K.; Creminon, C.; Wal, J.-M.; Grassi, J. Epitopic characterization of native bovine β-lactoglobulin. J. Immunol. Methods 2002, 266, 67–78. [Google Scholar] [CrossRef]
- Chikenji, G.; Kikuchi, M. What is the role of non-native intermediates of β-lactoglobulin in protein folding? Proc. Natl. Acad. Sci. USA 2000, 97, 14273–14277. [Google Scholar] [CrossRef] [Green Version]
- Jegouic, M.; Grinberg, V.Y.; Guingant, A.; Haertlé, T. Thiol-induced oligomerization of α-lactalbumin at high pressure. J. Protein Chem. 1996, 15, 501–509. [Google Scholar] [CrossRef]
- Hong, Y.-H.; Creamer, L.K. Changed protein structures of bovine β-lactoglobulin B and α-lactalbumin as a consequence of heat treatment. Int. Dairy J. 2002, 12, 345–359. [Google Scholar] [CrossRef]
- Dalgleish, D.G.; Senaratne, V.; Francois, S. Interactions between α-lactalbumin and β-lactoglobulin in the early stages of heat denaturation. J. Agric. Food Chem. 1997, 45, 3459–3464. [Google Scholar] [CrossRef]
- Calvo, M.M.; Leaver, J.; Banks, J.M. Influence of other whey proteins on the heat-induced aggregation of α-lactalbumin. Int. Dairy J. 1993, 3, 719–727. [Google Scholar] [CrossRef]
- Hochwallner, H.; Schulmeister, U.; Swoboda, I.; Balic, N.; Geller, B.; Nystrand, M.; Härlin, A.; Thalhamer, J.; Scheiblhofer, S.; Niggemann, B. Microarray and allergenic activity assessment of milk allergens. Clin. Exp. Allergy 2010, 40, 1809–1818. [Google Scholar] [CrossRef] [PubMed]
- Alessandri, S.; Sancho, A.; Vieths, S.; Mills, C.E.; Wal, J.-M.; Shewry, P.R.; Rigby, N.; Hoffmann-Sommergruber, K. High-throughput NMR assessment of the tertiary structure of food allergens. PLoS ONE 2012, 7, e39785. [Google Scholar] [CrossRef] [Green Version]
- Wal, J.-M. Cow’s milk proteins/allergens. Ann. Allergy Asthma Immunol. 2002, 89, 3–10. [Google Scholar] [CrossRef]
- Patel, H.A.; Singh, H.; Anema, S.G.; Creamer, L.K. Effects of heat and high hydrostatic pressure treatments on disulfide bonding interchanges among the proteins in skim milk. J. Agric. Food Chem. 2006, 54, 3409–3420. [Google Scholar] [CrossRef]
- Huppertz, T.; Vasiljevic, T.; Zisu, B.; Deeth, H. Novel processing technologies: Effects on whey protein structure and functionality. In Whey Proteins; Academic Press: London, UK, 2019; pp. 281–334. [Google Scholar]
- McSweeney, P.L.; O’Mahony, J.A. Advanced Dairy Chemistry: Volume 1B: Proteins: Applied Aspects; Springer: New York, NY, USA, 2016. [Google Scholar]
- Rattray, W.; Jelen, P. Thermal stability of skim milk/whey protein solution blends. Food Res. Int. 1997, 30, 327–334. [Google Scholar] [CrossRef]
- Chaplin, L.C.; Lyster, R.L. Irreversible heat denaturation of bovine α-lactalbumin. J. Dairy Res. 1986, 53, 249–258. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code-Level of Variables | Experimental Level of Variables | |||||
|---|---|---|---|---|---|---|
| Test Number | x1 | x2 | x3 | P (MPa) | T (°C) | t (min) |
| 1 | −1 | −1 | −1 | 200 | 25 | 35 |
| 2 | +1 | −1 | −1 | 600 | 25 | 35 |
| 3 | −1 | +1 | −1 | 200 | 65 | 35 |
| 4 | +1 | +1 | −1 | 600 | 65 | 35 |
| 5 | −1 | −1 | +1 | 200 | 25 | 115 |
| 6 | +1 | −1 | +1 | 600 | 25 | 115 |
| 7 | −1 | +1 | +1 | 200 | 65 | 115 |
| 8 | +1 | +1 | +1 | 600 | 65 | 115 |
| 9 | −1.68 | 0 | 0 | 63.6 | 45 | 75 |
| 10 | +1.68 | 0 | 0 | 736.4 | 45 | 75 |
| 11 | 0 | −1.68 | 0 | 400 | 11.36 | 75 |
| 12 | 0 | +1.68 | 0 | 400 | 78.64 | 75 |
| 13 | 0 | 0 | −1.68 | 400 | 45 | 7.72 |
| 14 | 0 | 0 | +1.68 | 400 | 45 | 142.28 |
| 15 | 0 | 0 | 0 | 400 | 45 | 75 |
| 16 | 0 | 0 | 0 | 400 | 45 | 75 |
| 17 | 0 | 0 | 0 | 400 | 45 | 75 |
| 18 | 0 | 0 | 0 | 400 | 45 | 75 |
| 19 | 0 | 0 | 0 | 400 | 45 | 75 |
| 20 | 0 | 0 | 0 | 400 | 45 | 75 |
| Variables | %IgG Binding | Dimerization | % IgG Binding | % IgG Binding | |||
|---|---|---|---|---|---|---|---|
| Test Number | P (MPa) | T (°C) | t (min) | Exp. Value | Exp. Value | Cal. Value a | Cal. Value b |
| 1 | 505 | 56 | 102 | 21.4 ± 6.4 | 84.6 | 1.4 | 12.6 |
| 2 | 505 | 51 | 102 | 27.2 ± 0.5 | 71.2 | 1.6 | 16.5 |
| 3 | 505 | 61 | 102 | 29.2 ± 0.6 | 74.9 | 1.5 | 15.2 |
| 4 | 455 | 56 | 102 | 19.4 ± 1.8 | 74.9 | 2.1 | 15.2 |
| 5 | 555 | 56 | 102 | 32.6 ± 7.3 | 76.4 | 2.0 | 14.8 |
| 6 | 505 | 56 | 92 | 34.5 ± 8.6 | 71.5 | 1.7 | 16.4 |
| 7 | 505 | 56 | 112 | 27.4 ± 12.5 | 75.4 | 1.6 | 15.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, X.; Vivien Chua, J.; Le, Q.A.; Trujillo, F.J.; Oh, M.-H.; Campbell, D.E.; Mehr, S.; Lee, N.A. A Response Surface Methodology (RSM) Approach for Optimizing the Attenuation of Human IgE-Reactivity to β-Lactoglobulin (β-Lg) by Hydrostatic High Pressure Processing. Foods 2021, 10, 1741. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10081741
Sun X, Vivien Chua J, Le QA, Trujillo FJ, Oh M-H, Campbell DE, Mehr S, Lee NA. A Response Surface Methodology (RSM) Approach for Optimizing the Attenuation of Human IgE-Reactivity to β-Lactoglobulin (β-Lg) by Hydrostatic High Pressure Processing. Foods. 2021; 10(8):1741. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10081741
Chicago/Turabian StyleSun, Xin, Jialing Vivien Chua, Quynh Anh Le, Francisco J. Trujillo, Mi-Hwa Oh, Dianne E. Campbell, Sam Mehr, and Nanju Alice Lee. 2021. "A Response Surface Methodology (RSM) Approach for Optimizing the Attenuation of Human IgE-Reactivity to β-Lactoglobulin (β-Lg) by Hydrostatic High Pressure Processing" Foods 10, no. 8: 1741. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10081741