
Astragalin and Isoquercitrin Isolated from Aster scaber Suppress LPS-Induced Neuroinflammatory Responses in Microglia and Mice

 , ,
, , 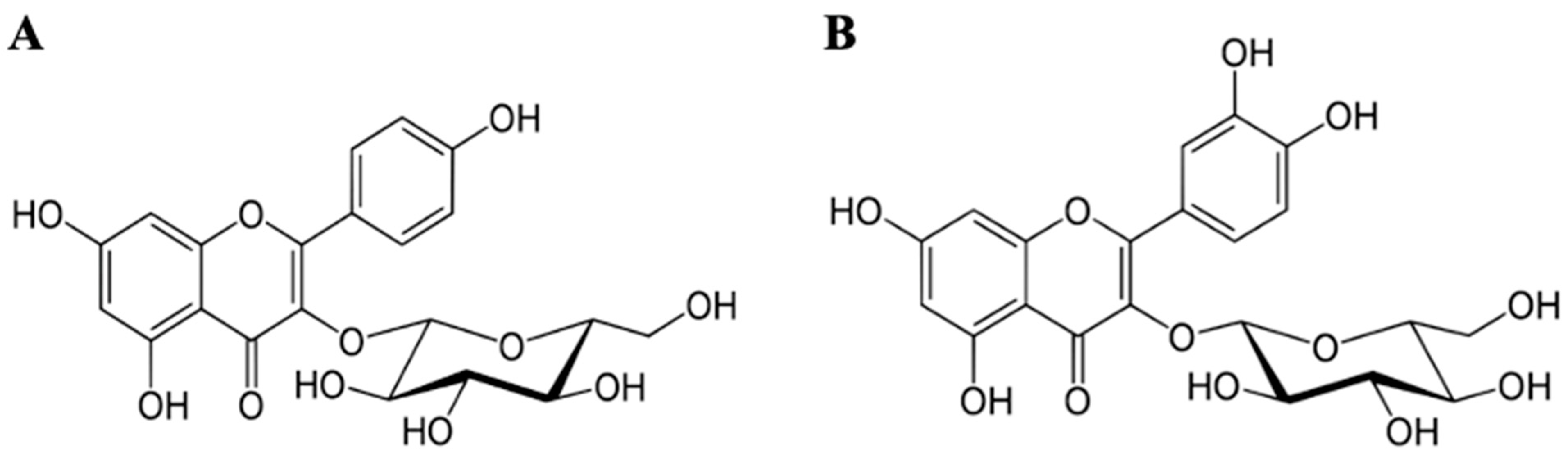{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Reagents and Apparatus
2.3. Isolation of Phytochemical Constituents from A. scaber
2.4. Microglial Cells Culture
2.5. Cell Viability and NO Content
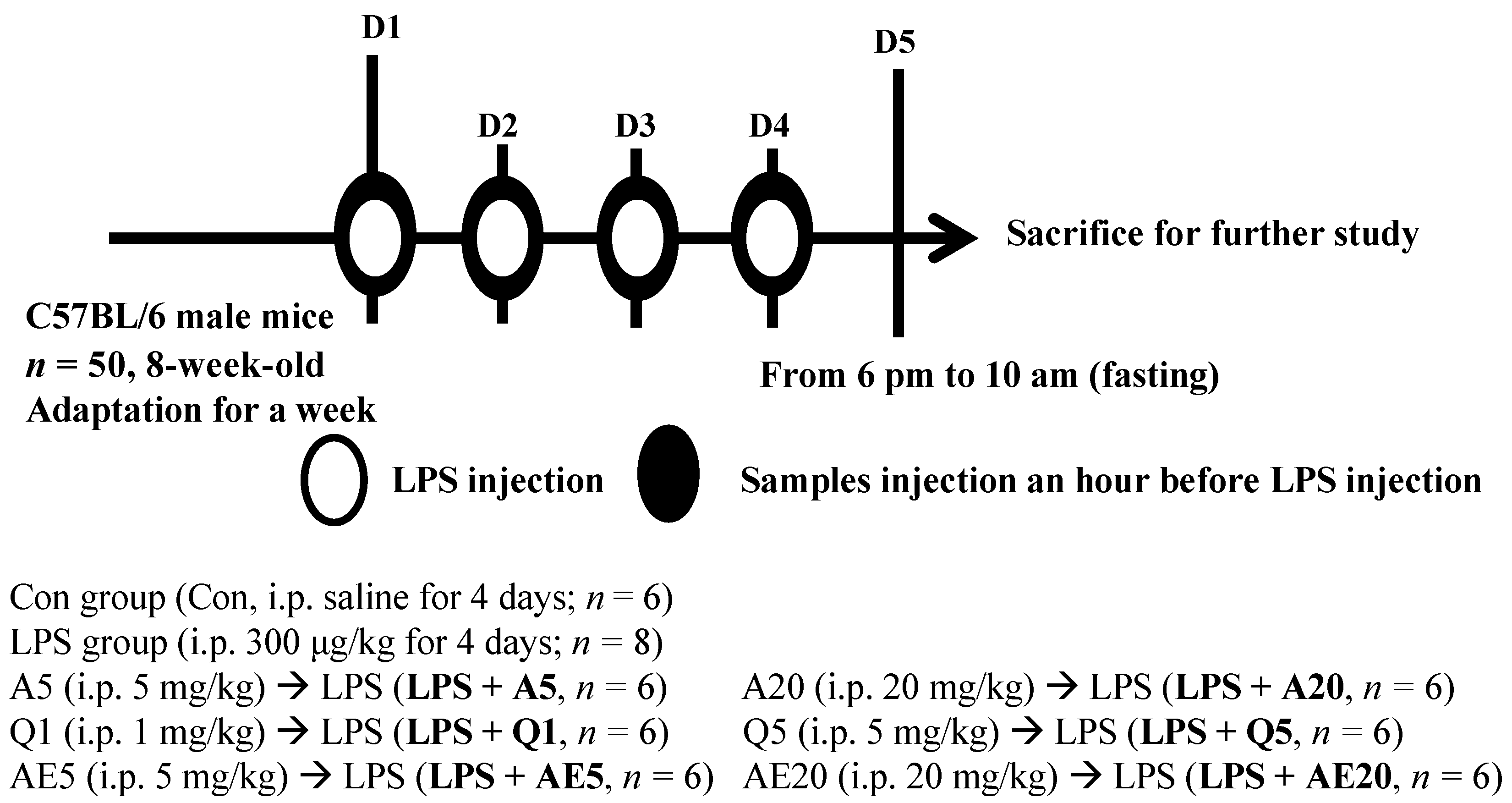
2.6. Animal Treatments
2.7. Total RNA Extraction and RT-PCR
2.8. Western Blot Analysis
2.9. Assay for TNF-α, IL-1β and IL-6 Secretion
2.10. ROS Level Measurement
2.11. DPPH Assay
2.12. Statistical Analysis
3. Results
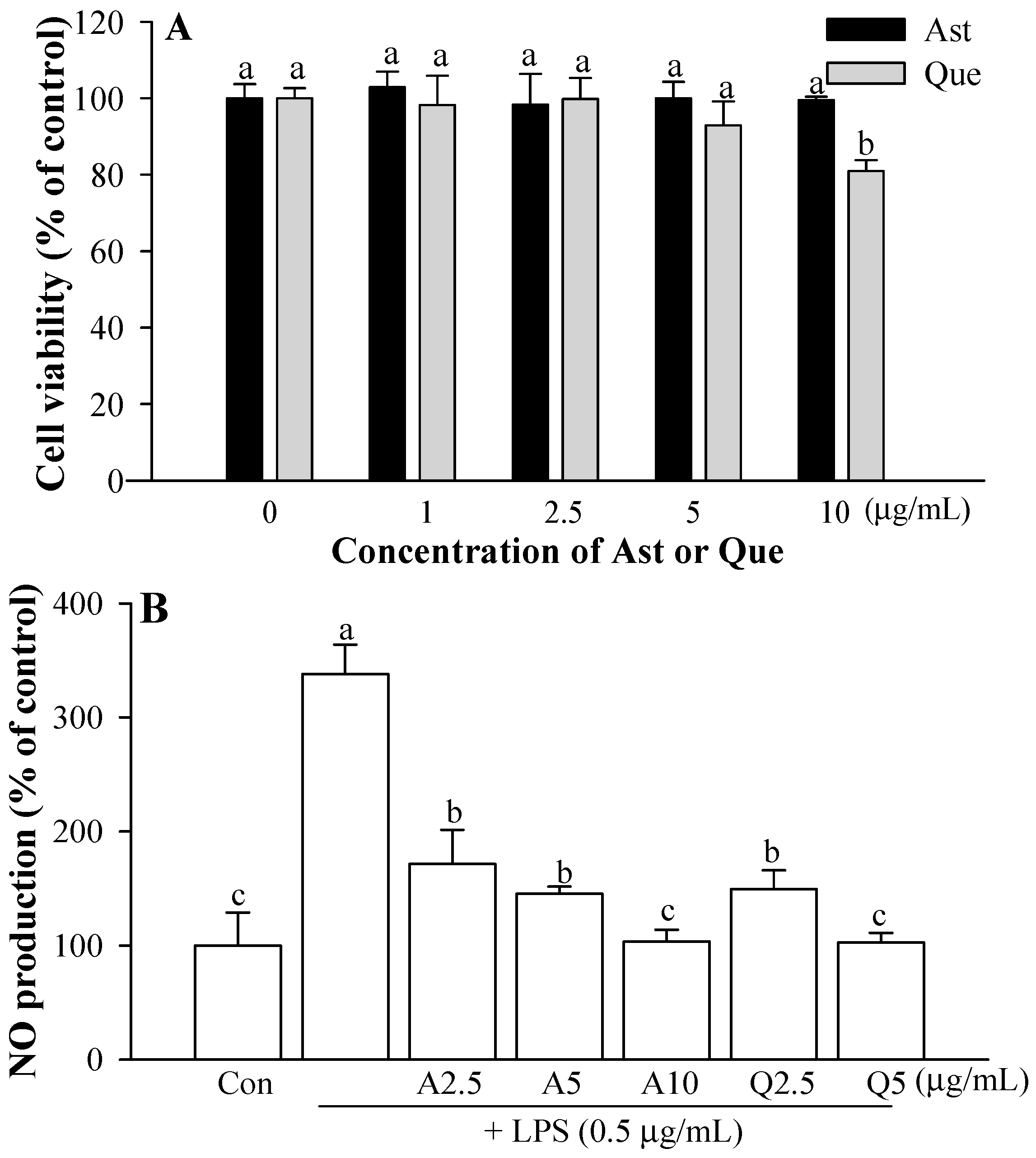
3.1. Cytotoxicity of Ast and Que Isolated from A. scaber
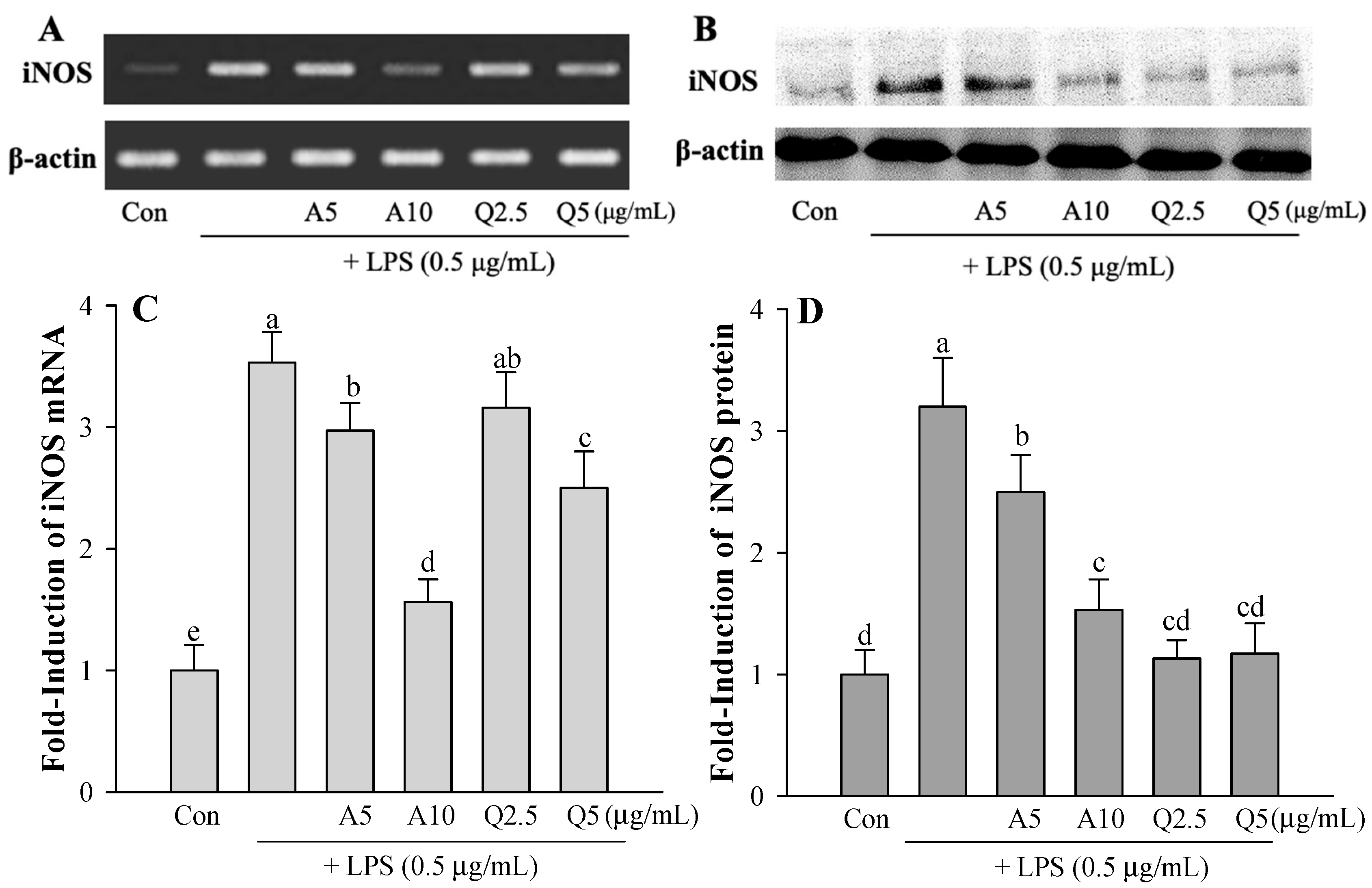
3.2. Effects of Ast and Que on LPS-induced Microglia
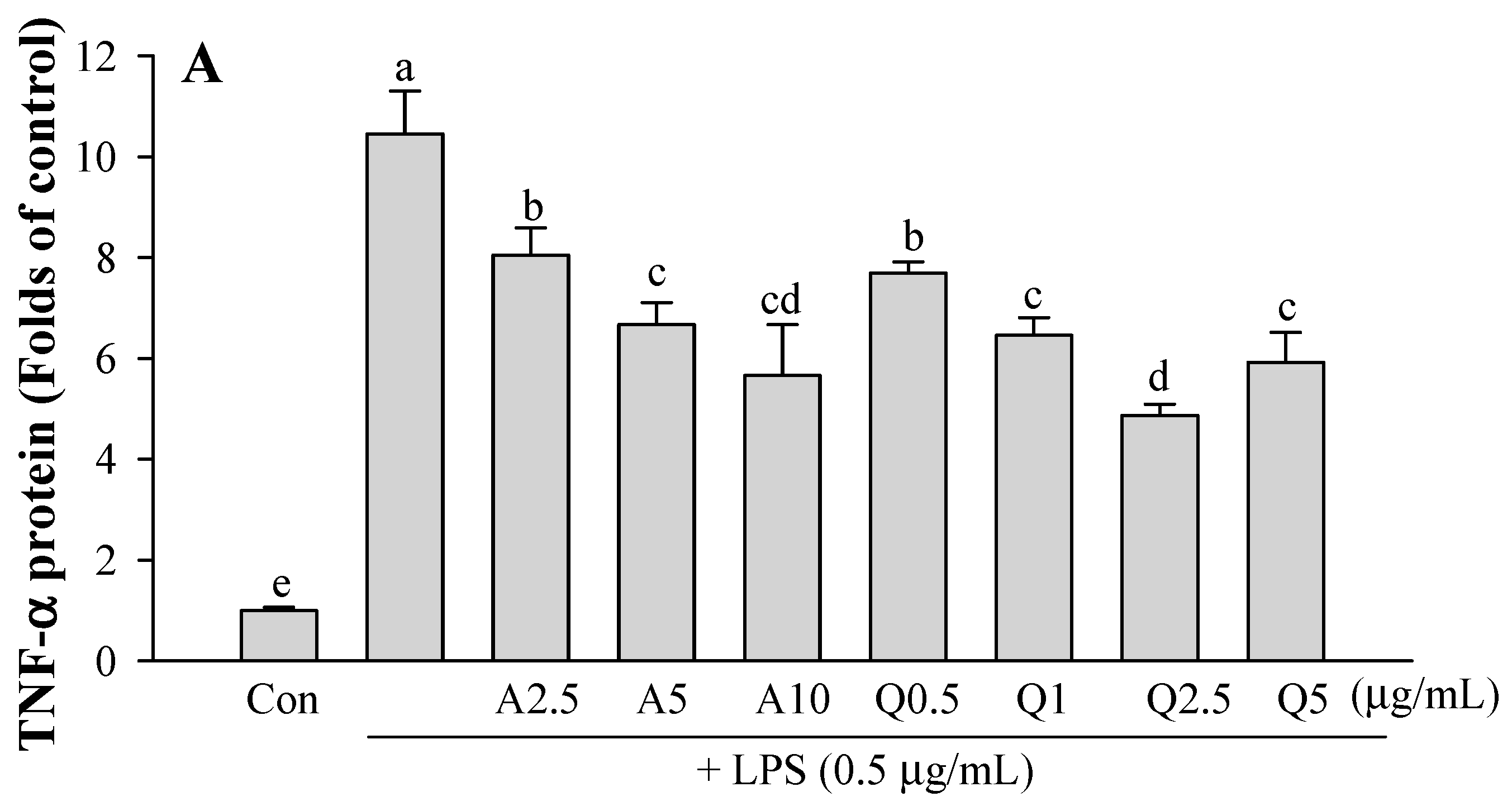
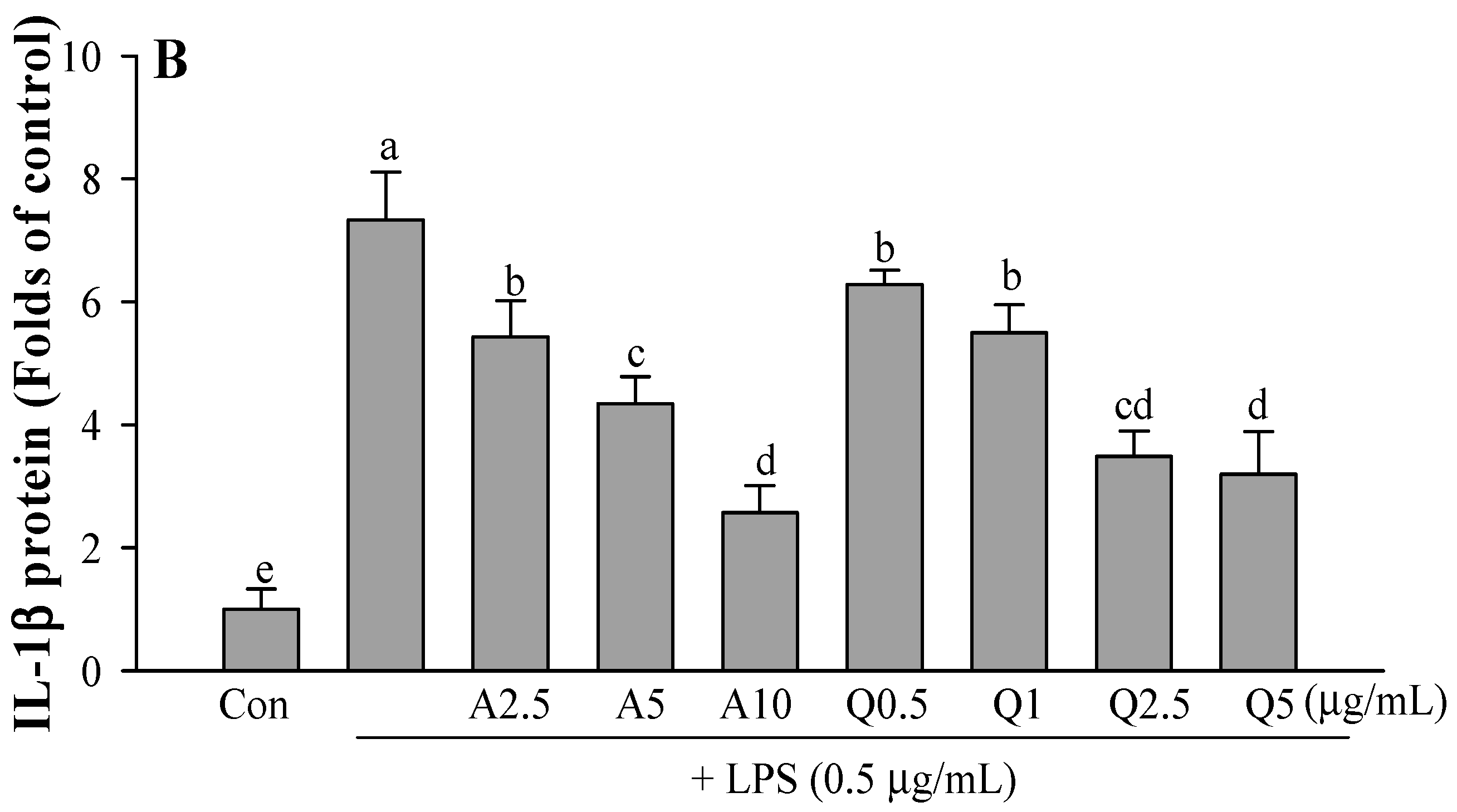
3.3. Effects of Ast and Que on TNF-α and IL-1β Production in LPS-Treated Microglia
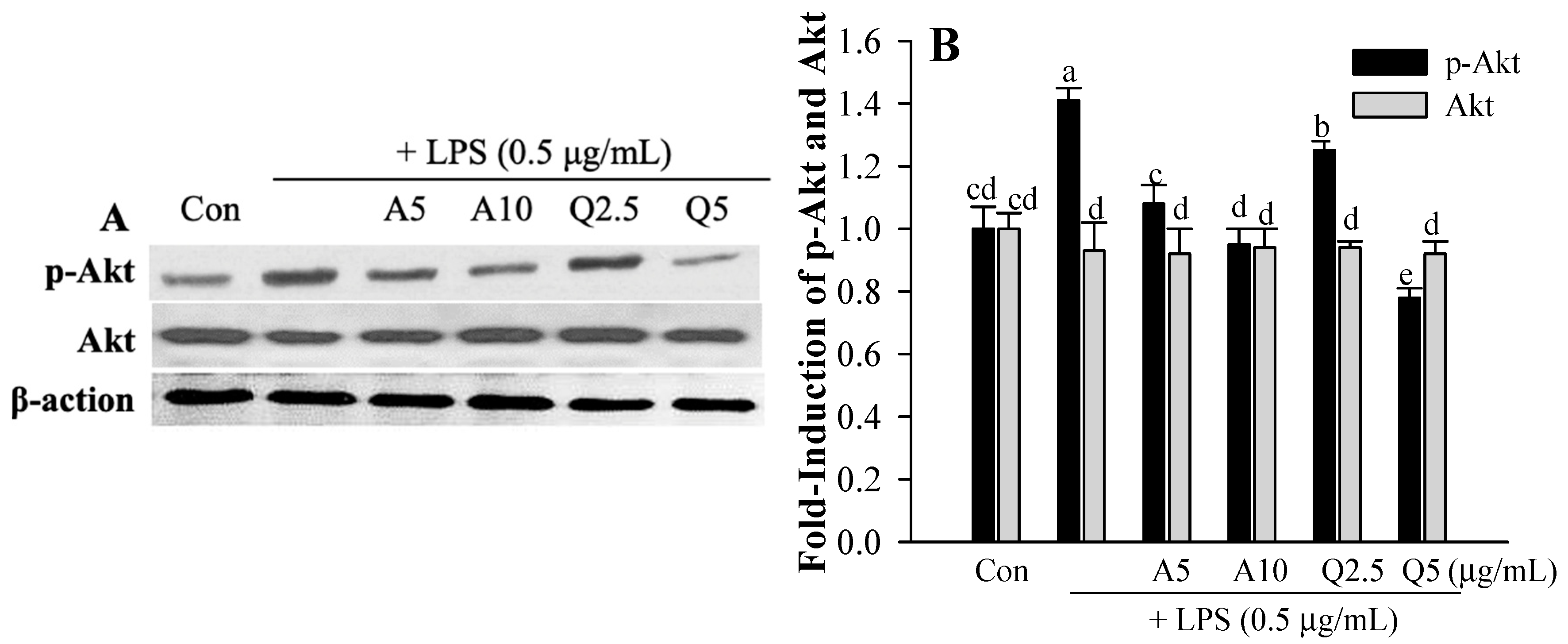
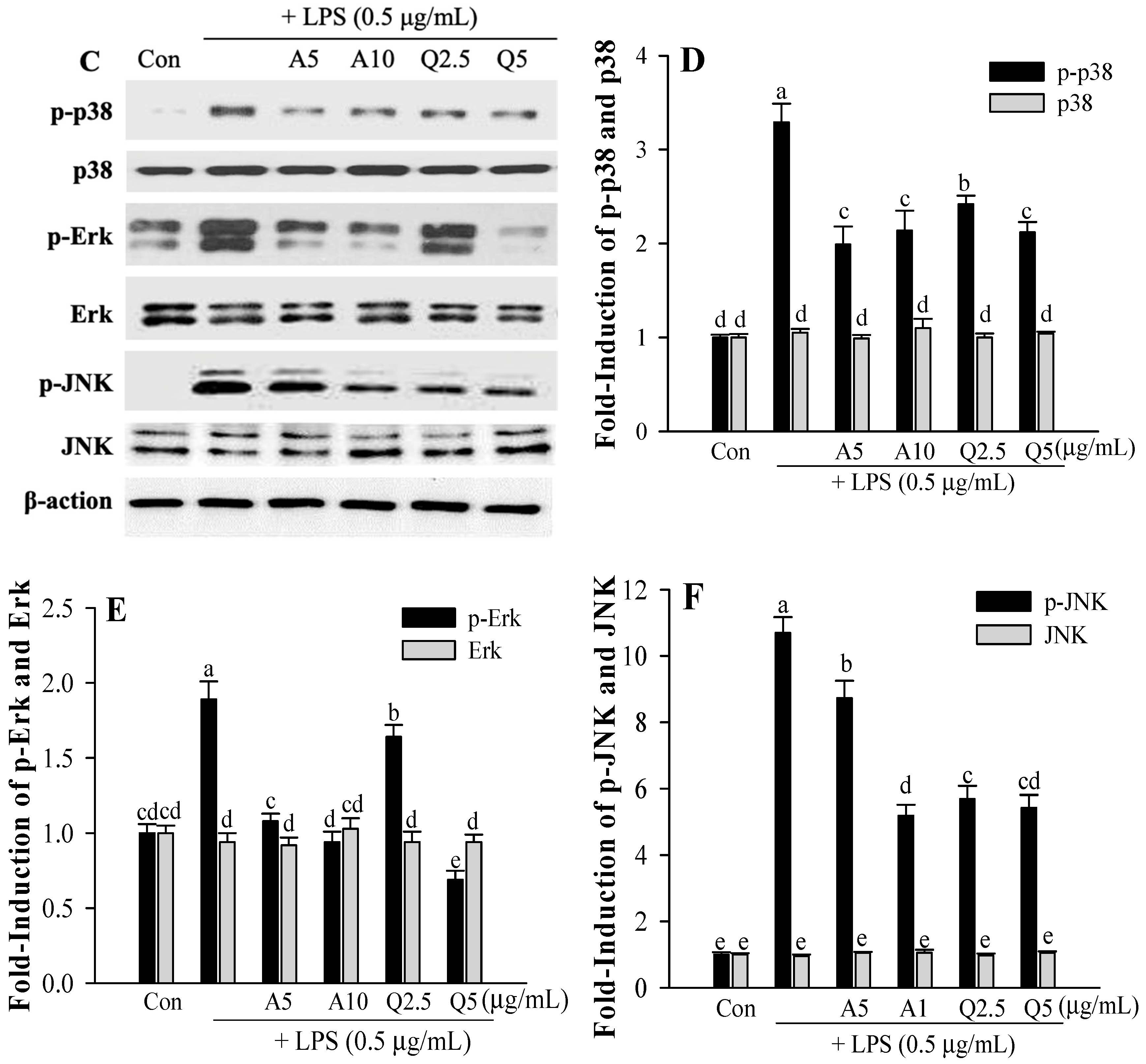
3.4. Ast and Que Inhibit LPS-Induced Phosphorylation of the Akt/MAPK Pathway in Microglia
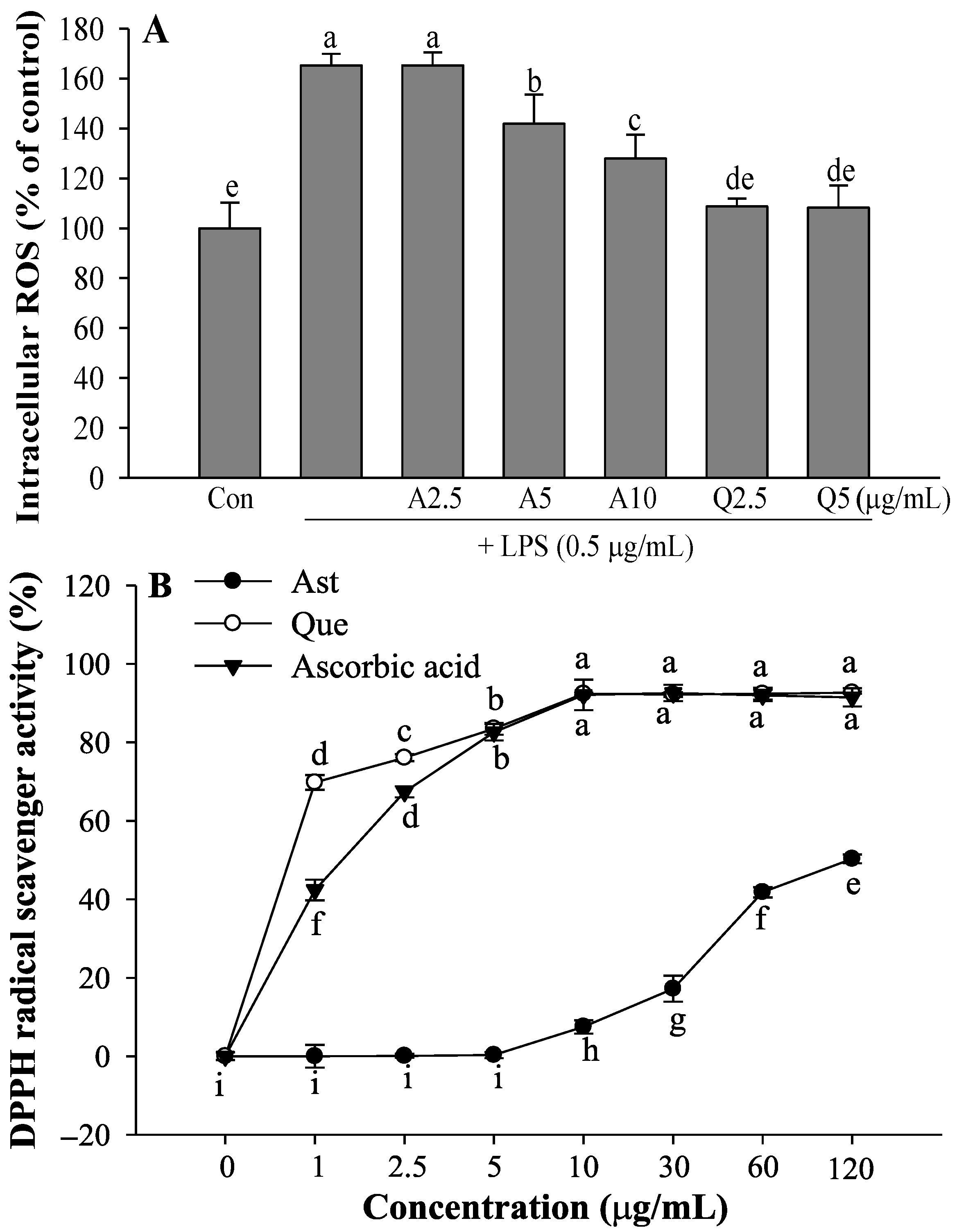
3.5. Effects of Ast and Que on LPS-Induced Intracellular ROS Production and DPPH Radical Scavenging Activity
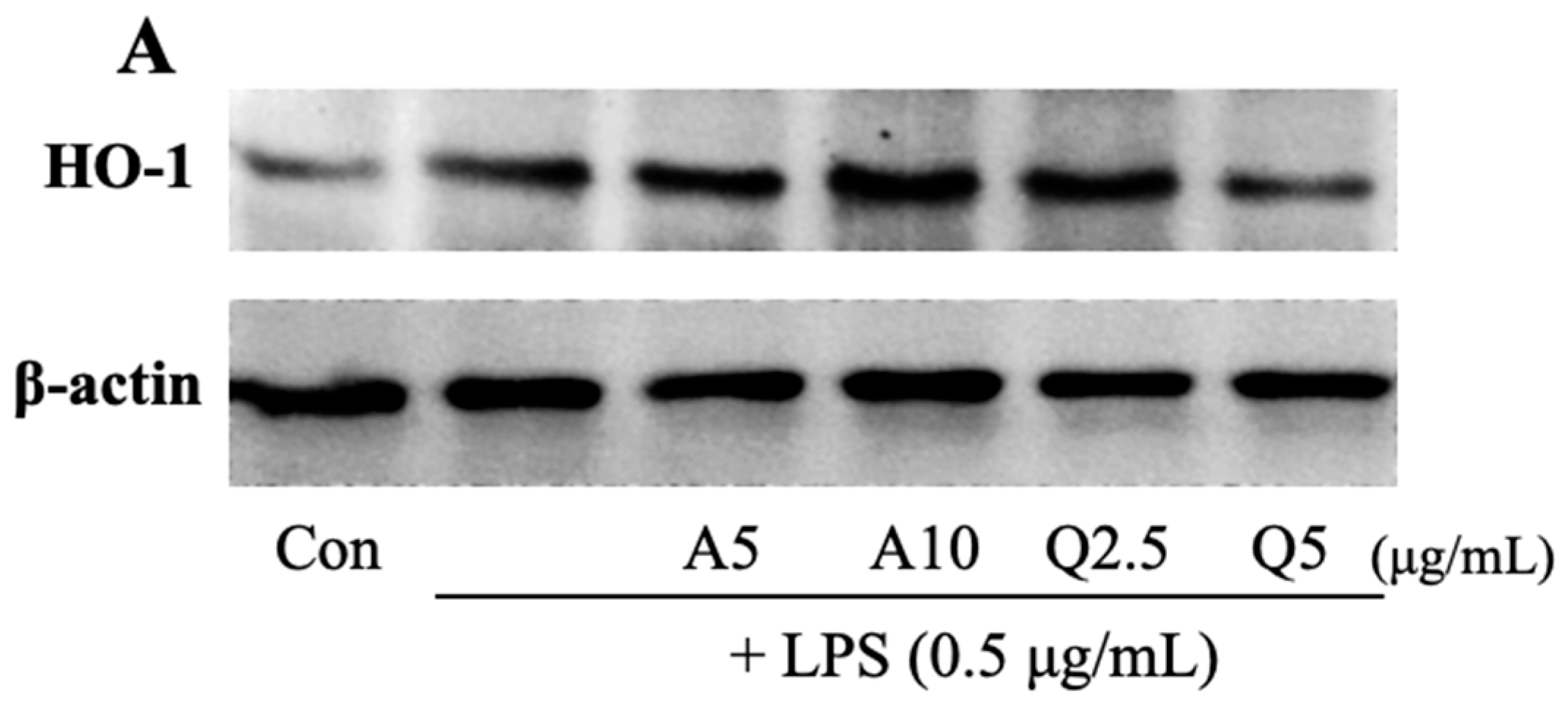
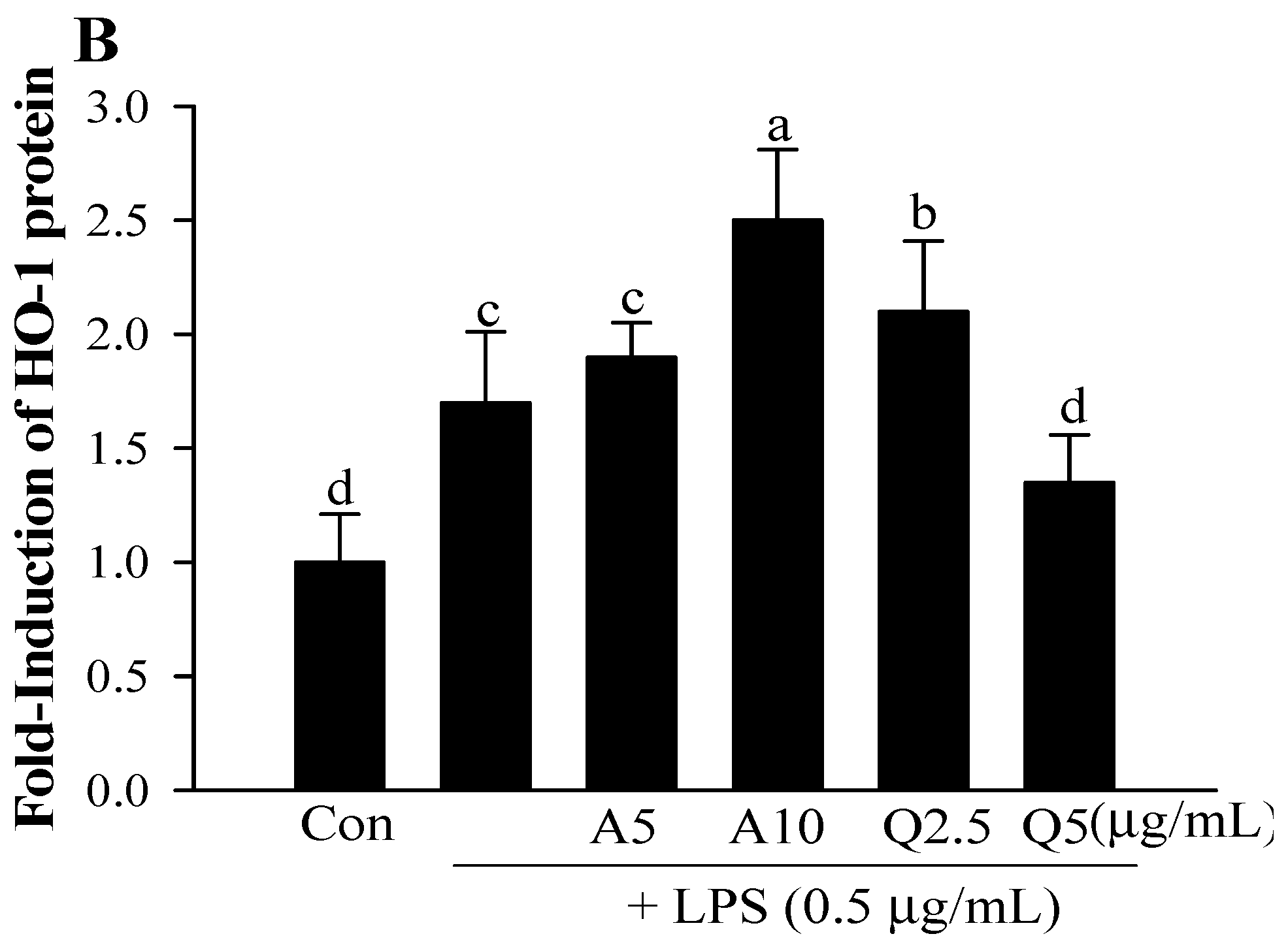
3.6. Effects of Ast and Que on HO-1 Production in LPS-Treated Microglia
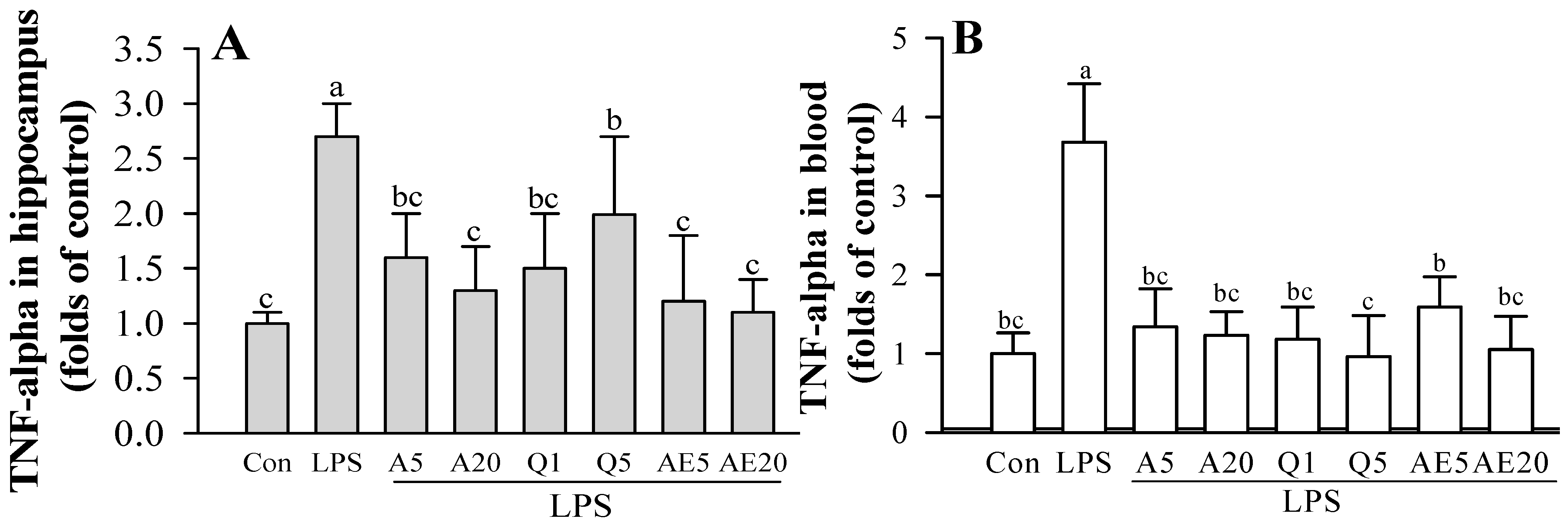
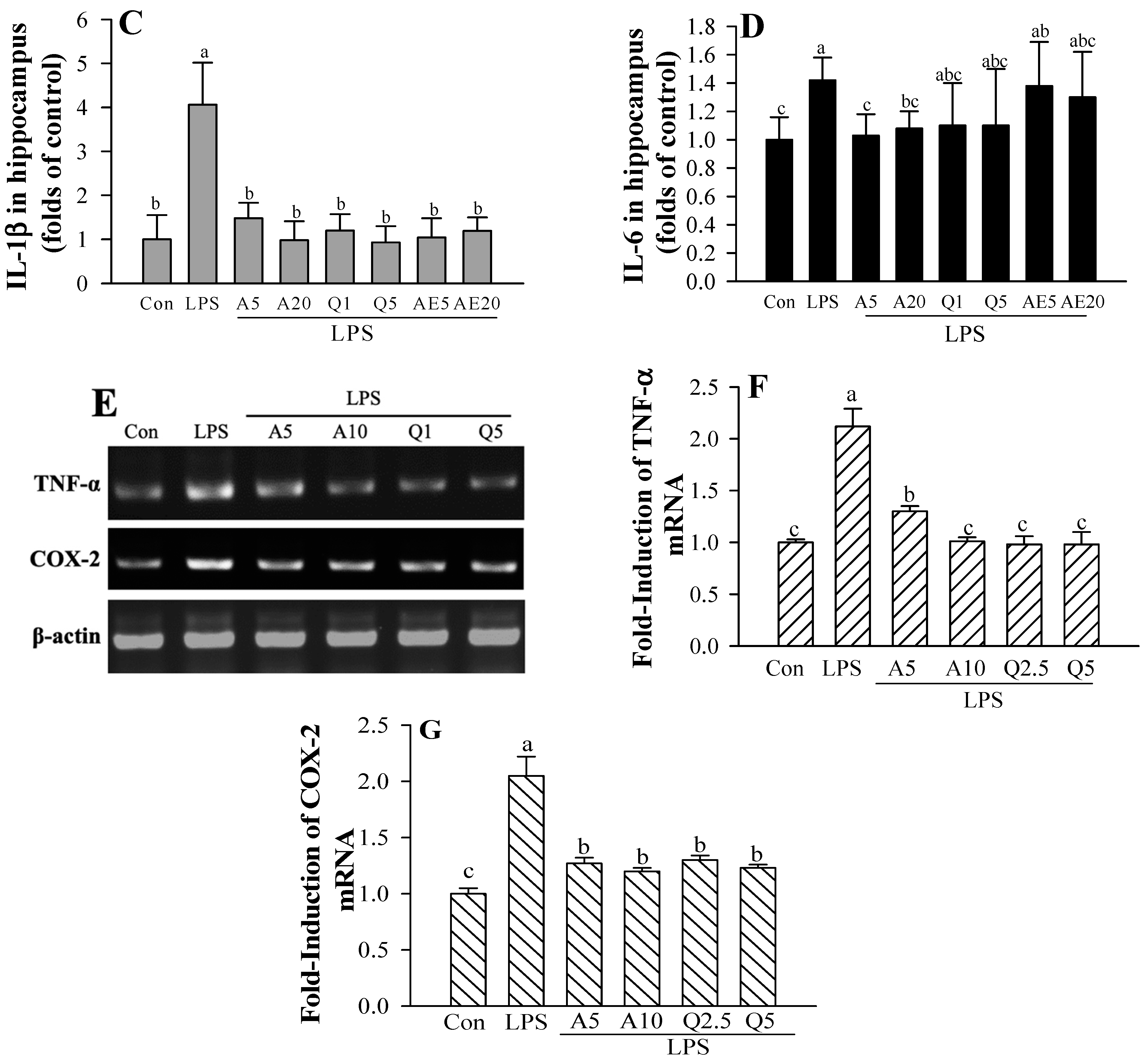
3.7. Effects of Ast and Que on Cytokines Production in the Hippocampus of LPS-Induced Mice
3.8. Ast and Que Inhibit LPS-Induced Phosphorylation of NF-κB/MAPK Pathwas in the Hippocampus of LPS-Induced Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Block, M.L.; Zecca, L.; Hong, J.S. Microglia-mediated neurotoxicity: Uncovering the molecular mechanisms. Nat. Rev. Neurosci. 2007, 8, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Heneka, M.T.; O’Banion, M.K. Inflammatory processes in Alzheimer’s disease. J. Neuroimmunol. 2007, 184, 69–91. [Google Scholar] [CrossRef] [PubMed]
- Lawson, L.J.; Perry, V.H.; Dri, P.; Gordon, S. Heterogeneity in the distribution and morphology of microglia in the normal adult mouse brain. Neuroscience 1990, 39, 151–170. [Google Scholar] [CrossRef]
- Tambuyzer, B.R.; Ponsaerts, P.; Nouwen, E.J. Microglia: Gatekeepers of central nervous system immunology. J. Leukoc. Biol. 2009, 85, 352–370. [Google Scholar] [CrossRef] [PubMed]
- Giulian, D. Microglia and the immune pathology of Alzheimer’s disease. Am. J. Hum. Gen. 1999, 65, 13–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, H.; Liu, W.; Zhang, W.; Hong, J.H.; Liu, B.; Zhang, W.; Hong, J. Novel anti-inflammatory therapy for Parkinson’s disease. Trends Pharmacol. Sci. 2003, 24, 395–401. [Google Scholar] [CrossRef]
- West, M.; Mhatre, M.; Ceballos, A.; Floyd, R.; Grammas, P.; Gabbita, S.; Hamdheydari, L.; Mai, T.; Mou, S.; Pye, Q.; et al. The arachidonic acid 5-lipoxygenase inhibitor nordihydroguaiaretic acid inhibits tumor necrosis factor-alpha activation of microglia and extends survival of G93A-SOD1 transgenic mice. J. Neurochem. 2004, 91, 133–143. [Google Scholar] [CrossRef]
- Wei, J.C.C.; Huang, H.C.; Chen, W.J.; Huang, C.N.; Peng, C.H.; Lin, C.L. Epigallocatechin gallate attenuates amyloid β-induced inflammation and neurotoxicity in EOC 13.31 microglia. Eur. J. Pharmacol. 2016, 770, 16–24. [Google Scholar]
- Kaminska, B. MAPK signalling pathways as molecular targets for anti-inflammatory therapy—From molecular mechanisms to therapeutic benefits. BBA-Proteins Proteom. 2005, 1754, 253–262. [Google Scholar] [CrossRef]
- Sun, G.Y.; Chen, Z.; Jasmer, K.J.; Chuang, D.Y.; Gu, Z.; Hannink, M.; Simony, A. Quercetin attenuate inflammatory responses in BV2 microglial cells: Role of MAPKs on the Nrf2 pathway and induction of heme oxygenase-1. PLoS ONE 2015, 10, e0141509. [Google Scholar]
- Saponaro, C.; Cianciulli, A.; Calvello, R.; Dragone, T.; Iacobazzi, F.; Panaro, M.A. The P13K/Akt pathway is required for LPS activation of microglial cells. Immunopharmacol. Immunotoxicol. 2012, 34, 858–865. [Google Scholar] [CrossRef] [PubMed]
- Cianciulli, P.A.; Calvello, R.; Porro, C.; Trotta, T.; Salvatore, R.; Panaro, A. PI3k/Akt signaling pathway plays a crucial role in the anti–inflammatory effects of curcumin in LPS-activated microglia. Int. Immunopharmacol. 2016, 36, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Spagnuolo, C.; Moccia, S.; Russo, G.L. Anti-inflammatory effects of flavonoids in neurodegenerative disorders. Eur. J. Med. Chem. 2018, 153, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Ryter, S.W.; Choi, A.M. Heme oxygenase-1/carbon monoxide: From metabolism to molecular therapy. Am. J. Respir. Cell Mol. Biol. 2009, 41, 251–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, Q.; Huang, B.; Zhang, X.; Zhu, Y.; Chen, X. Astaxanthin protects against MPP(+)-induced oxidative stress in PC12 cells via the HO-1/NOX2 axis. BMC Neurosci. 2012, 13, 156. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.M.; Chung, M.J.; Ha, T.J.; Choi, H.N.; Jang, S.J.; Kim, S.O.; Chun, M.H.; Do, S.I.; Choo, Y.K.; Park, Y.I. Neuroprotective effects of black soybean anthocyanins via inactivation of ASK1-JNK/p38 pathways and mobilization of cellular sialic acids. Life Sci. 2012, 90, 874–882. [Google Scholar] [CrossRef]
- Chung, M.J.; Lee, S.H.; Park, Y.I.; Lee, J.S.; Kwon, K.H. Neuroprotective effects of phytosterols and flavonoids from Cirsium setidens and Aster scaber in human brain neuroblastoma SK-N-SH cells. Life Sci. 2016, 148, 173–182. [Google Scholar] [CrossRef]
- Chung, T.Y.; Eiserich, J.P.; Shibamoto, T. Volatile compounds isolated from edible Korean chamchwi (Aster scaber Thunb). J. Agric. Food Chem. 1993, 41, 1693–1697. [Google Scholar] [CrossRef]
- Jung, C.M.; Kwon, H.C.; Seo, J.J.; Ohizumi, Y.; Matsunaga, K.; Saito, S.; Lee, K.R. Two new monoterpene peroxide glycoside from Aster scaber. Chem. Pharm. Bull. 2001, 49, 912–914. [Google Scholar] [CrossRef] [Green Version]
- Choi, Y.E.; Choi, S.I.; Han, X.; Men, X.; Jang, G.W.; Kwon, H.Y.; Kang, S.R.; Han, J.S.; Lee, O.H. Radical scavenging-linked anti-adipogenic activity of Aster scaber ethanolic extract and its bioactive compound. Antioxidants 2020, 9, 1290. [Google Scholar] [CrossRef]
- Choi, J.H.; Park, Y.H.; Lee, I.S.; Lee, S.P. Antioxidant activity and inhibitory effect of Aster scaber Thunb. Extract on adipocyte differentiation in 3T3-L1 cells. Korean J. Food Sci. Technol. 2013, 45, 356–363. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.S.; Kim, S.H. Inhibitory effect of astragalin on expression of lipopolysaccharide-induced inflammatory mediators through NF-kB in macrophages. Arch. Pharm. Res. 2011, 34, 2101–2107. [Google Scholar] [CrossRef] [PubMed]
- Kotani, M.; Matsumoto, M.; Fujita, A.; Higa, S.; Wang, W.; Suemura, M.; Kishimoto, T.; Tanaka, T. Persimmon leaf extract and astragalin inhibit development of dermatitis and IgE elevation in NC/Nga mice. J. Allergy Clin. Immunol. 2000, 106, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Qu, D.; Han, J.; Ren, H.; Yang, W.; Zhang, X.; Zheng, Q.; Wang, D. Cardioprotective effects of astragalin against myocardial ischemia/reperfusion injury in isolated rat heart. Oxid. Med. Cell. Longev. 2016, 2016, 8194690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, M.; Li, W.Y.; Xie, J.; Wang, Z.L.; Wen, Y.L.; Zhao, C.C.; Tao, L.; Li, L.F.; Tian, Y.; Sheng, J. Astragalin inhibits the proliferation and migration of human colon cancer HCT116 cells by regulating the NF-kB signaling pathway. Front. Pharmacol. 2021, 12, 639256. [Google Scholar] [CrossRef] [PubMed]
- Soromou, L.W.; Chen, N.; Jiang, L.; Huo, M.; Wei, M.; Chu, X.; Martin, F.; Millimouno, F.M.; Feng, H.; Sidime, Y.; et al. Astragalin attenuates lipopolysaccharide-induced inflammatory responses by down-regulating NF-kB signaling pathway. Biochem. Biophys. Res. Commun. 2012, 419, 256–261. [Google Scholar] [CrossRef] [PubMed]
- Rogerio, A.P.; Kanashiro, A.; Fontanari, C.; Silva, E.V.G.; Lucisano-Valim, Y.M.; Soares, E.G.; Faccioli, L.H. Anti-inflammatory activity of quercetin and isoquercitrin in experimental murine allergic asthma. Inflamm. Res. 2007, 56, 402–408. [Google Scholar] [CrossRef]
- Ll, L.; Zhang, X.H.; Liu, G.R.; Liu, C.; Dong, Y.M. Isoquercitrin suppresses the expression of histamine and pro-inflammatory cytokines by inhibiting the activation of MAP kinases and NF-kB in human KU812 cells. Chin. J. Nat. Med. 2016, 14, 407–412. [Google Scholar]
- Jung, S.H.; Kim, B.J.; Lee, E.H.; Osborne, N.N. Isoquercitrin is the most effective antioxidant in the plant Thuja orientalis and able to counteract oxidative-induced damage to a transformed cell line (RGC-5 cells). Neurochem. Int. 2010, 57, 713–721. [Google Scholar] [CrossRef]
- Xie, W.; Wang, M.; Chen, C.; Zhang, X.; Melzig, M.F. Hepatoprotective effect of isoquercitrin against acetaminophen-induced liver injury. Life Sci. 2016, 152, 180–189. [Google Scholar] [CrossRef]
- Kim, C.H.; Kim, J.E.; Song, Y.J. Antiviral activities of quercetin and isoquercitrin against human herpesviruses. Molecules 2020, 25, 2379. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Wang, W.; Li, H.; Liu, J.; Zhang, P.; Cheng, Y.; Qin, X.; Hu, Y.; Wei, Y. The neuroprotective effects of isoquercitrin purified from apple pomace by high-speed countercurrent chromatography in the MPTP acute mouse model of Parkinson’s disease. Food Funct. 2021, 12, 6091–6101. [Google Scholar] [CrossRef] [PubMed]
- Andy, S.N.; Pandy, V.; Alias, Z.; Kadir, H.A. Deoxyelephantopin ameliorates lipopolysaccharides (LPS)-induced memory impairments in rats: Evidence for its anti-neuroinflammatory properties. Life Sci. 2018, 206, 45–60. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.H.; Chung, M.J.; Jeong, D.Y.; Oh, D.H. Immunostimulary activity of isoflavone-glycosides and ethanol extract from a fermented soybean product in human primary immune cells. J. Med. Food 2014, 17, 1113–1121. [Google Scholar] [CrossRef] [PubMed]
- Chung, M.J.; Sohng, J.K.; Choi, D.J.; Park, Y.I. Inhibitory effect of phloretin and biochanin A on IgE-mediated allergic responses in rat basophilic leukemia RBL-2H3 cells. Life Sci. 2013, 93, 401–408. [Google Scholar] [CrossRef]
- Min, J.S.; Lee, D.S. A screen for dual-protection molecules from a natural product library against neuronal cell death and microglial cell activation. J. Life Sci. 2015, 25, 656–662. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.S.; Ryu, D.K.; Lim, J.H.; Cho, S.R.; Kang, B.Y.; Choi, H.J. Artesunate activates Nrf2 pathway-driven anti-inflammatory potential through ERK signaling in microglial BV2 cells. Neurosci. Lett. 2012, 509, 17–21. [Google Scholar] [CrossRef]
- Sugama, S. Stress-induced microglial activation may facilitate the progression of neurodegenerative disorders. Med. Hypotheses 2009, 73, 1031–1034. [Google Scholar] [CrossRef]
- Vaca, K.; Wendt, E. Divergent effects of astroglial and microglia secretions on neuron growth and survival. Exp. Neurol. 1992, 118, 62–72. [Google Scholar] [CrossRef]
- Zhang, F.X.; Xu, R.S. Juglanin ameliorates LPS-induced neuroinflammation in animal models of Parkinson’s disease and cell culture via inactivating TLR4/NF-kB pathway. Biomed. Pharmacother. 2018, 97, 1011–1019. [Google Scholar] [CrossRef]
- Stansley, B.; Post, J.; Hensley, K. A comparative review of cell culture systems for the study of microglial biology in Alzheimer’s disease. J. Neuroinflamm. 2012, 9, 115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilms, H.; Zecca, L.; Rosenstiel, P.; Sievers, J.; Deuschl, G.; Lucius, R. Inflammation in Parkinson’s diseases and other neurodegenerative diseases: Cause and therapeutic implications. Curr. Pharm. Des. 2007, 13, 1925–1928. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.F.; Malik, A.B. NF-kB activation as pathological mechanism of septic shock and inflammation. Am. J. Physiol. Lung Cell Mol. Physiol. 2006, 290, L622–L645. [Google Scholar] [CrossRef] [PubMed]
- Jekabsone, A.; Neher, J.J.; Borutaite, V.; Brown, G.C. Nitric oxide from neuronal nitric oxide synthase sensitises neurons to hypoxia-induced death via competitive inhibition of cytochrome oxidase. J. Neurochem. 2007, 103, 346–356. [Google Scholar] [CrossRef]
- Min, K.J.; Lee, J.T.; Joe, E.H.; Kwon, T.K. An IκBα phosphorylation inhibitor induces heme oxygenase-1 (HO-1) expression through the activation of reactive oxygen species (ROS)–Nrf2–ARE signaling and ROS–PI3K/Akt signaling in an NF-κB-independent mechanism. Cell. Signal. 2011, 23, 1505–1513. [Google Scholar] [CrossRef]














Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, E.H.; Shim, Y.Y.; Lee, H.I.; Lee, S.; Reaney, M.J.T.; Chung, M.J. Astragalin and Isoquercitrin Isolated from Aster scaber Suppress LPS-Induced Neuroinflammatory Responses in Microglia and Mice. Foods 2022, 11, 1505. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11101505
Kim EH, Shim YY, Lee HI, Lee S, Reaney MJT, Chung MJ. Astragalin and Isoquercitrin Isolated from Aster scaber Suppress LPS-Induced Neuroinflammatory Responses in Microglia and Mice. Foods. 2022; 11(10):1505. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11101505
Chicago/Turabian StyleKim, Eun Hae, Youn Young Shim, Hye In Lee, Sanghyun Lee, Martin J. T. Reaney, and Mi Ja Chung. 2022. "Astragalin and Isoquercitrin Isolated from Aster scaber Suppress LPS-Induced Neuroinflammatory Responses in Microglia and Mice" Foods 11, no. 10: 1505. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11101505