
Analysis of Related Metabolites Affecting Taste Values in Rice under Different Nitrogen Fertilizer Amounts and Planting Densities

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Determination of Rice Quality and Taste Value
2.3. Metabolite Extraction
2.4. Quality Control Sample
2.5. Ultra−High Performance Liquid Chromatography–Mass Spectrometry (UHPLC−MS/MS) Analysis
2.6. Experimental Design and Statistical Analysis
3. Results
3.1. Analysis of Rice Quality and Taste Value
3.2. PCA and PLS−DA
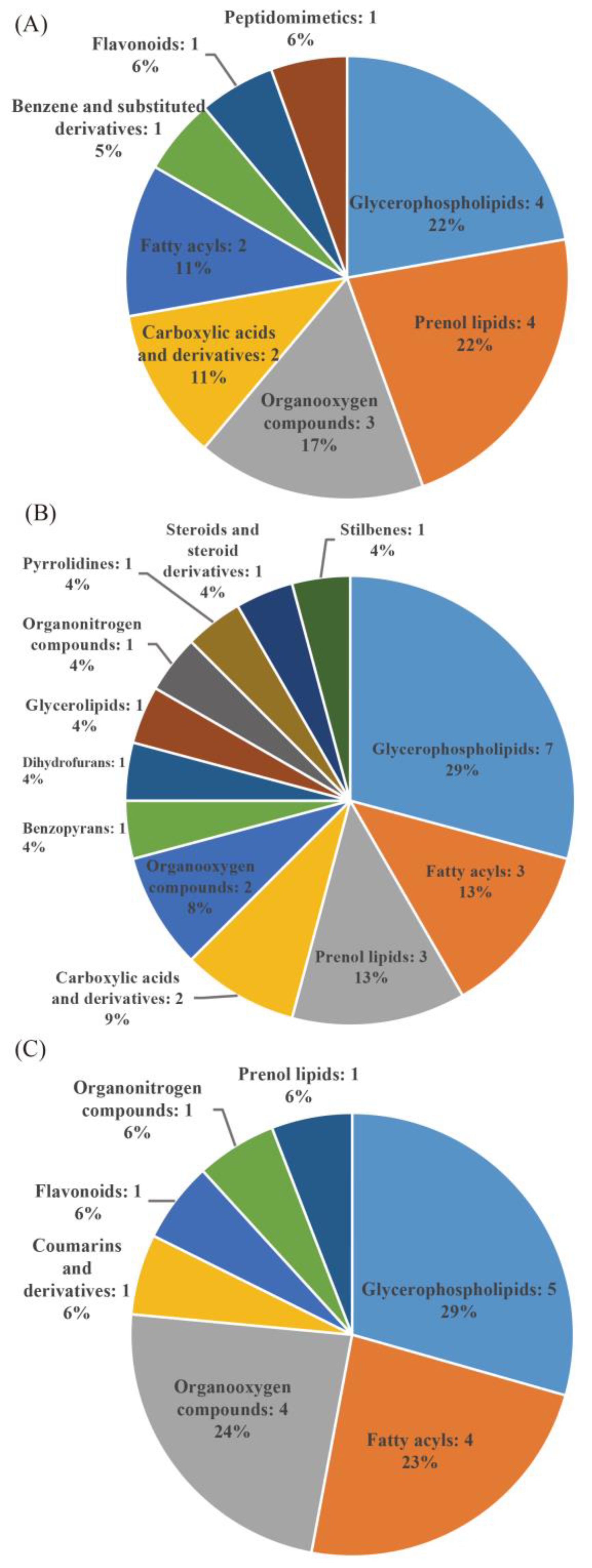
3.3. Differential Metabolite Analysis
3.4. KEGG Analysis
3.5. Correlation Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Pan, Y.Y.; Chen, Y.B.; Wang, C.R.; Li, H.; Huang, D.Q.; Zhou, D.G.; Wang, Z.D.; Zhao, L.; Gong, R.; Zhou, S.C. Metabolism of γ-aminobutyrate and 2-acetyl-1-pyrroline analyses at various grain developmental stages in rice (Oryza sativa L.). Chin. J. Rice Sci. 2021, 35, 121–129. [Google Scholar]
- Zhang, Y.; Zhang, W.; Wu, M.; Liu, G.; Zhang, Z.; Yang, J. Effects of irrigation schedules and phosphorus fertilizer rates on grain yield and quality of upland rice and paddy rice. Environ. Exp. Bot. 2021, 186, 1044652. [Google Scholar] [CrossRef]
- Cheng, B.; Jiang, Y.; Cao, C. Balance rice yield and eating quality by changing the traditional nitrogen management for sustainable production in China. J. Clean. Prod. 2021, 312, 127793. [Google Scholar] [CrossRef]
- Zhou, C.; Huang, Y.; Jia, B.; Wang, Y.; Wang, Y.; Xu, Q.; Li, R.; Wang, S.; Dou, F. Effects of cultivar, nitrogen rate, and planting density on rice-grain quality. Agronomy 2018, 8, 246. [Google Scholar] [CrossRef] [Green Version]
- Wei, H.; Meng, T.; Ge, J.; Zhang, X.; Shi, T.; Ding, E.; Lu, Y.; Li, X.; Tao, Y.; Chen, Y.; et al. Reduced nitrogen application rate with dense planting improves rice grain yield and nitrogen use efficiency: A case study in east China. Crop J. 2021, 9, 954–961. [Google Scholar] [CrossRef]
- Dong, N.; Sun, Y.; Guo, T.; Shi, C.; Zhang, Y.; Kan, Y.; Xiang, Y.; Zhang, H.; Li, Y.; Zhao, H.; et al. UDP-glucosyltransferase regulates grain size and abiotic stress tolerance associated with metabolic flux redirection in rice. Nat. Commun. 2020, 11, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Shen, S.; Zhou, S.; Li, Y.; Mao, Y.; Zhou, J.; Shi, Y.; An, L.; Zhou, Q.; Peng, W.; et al. Rice metabolic regulatory network spanning the entire life cycle. Mol. Plant 2022, 15, 258–275. [Google Scholar] [CrossRef]
- Kusano, M.; Yang, Z.; Okazaki, Y.; Nakabayashi, R.; Fukushima, A.; Saito, K. Using metabolomic approaches to explore chemical diversity in rice. Mol. Plant 2015, 8, 58–67. [Google Scholar] [CrossRef] [Green Version]
- Xiong, Q.; Shen, T.; Zhong, L.; Zhu, C.; Peng, X.; He, X.; Fu, J.; Ouyang, L.; Bian, J.; Hu, L.; et al. Comprehensive metabolomic, proteomic and physiological analyses of grain yield reduction in rice under abrupt drought-flood alternation stress. Physiol. Plant. 2019, 167, 564–584. [Google Scholar] [CrossRef]
- Chen, W.; Wang, W.; Peng, M.; Gong, L.; Gao, Y.; Wan, J.; Wang, S.; Shi, L.; Zhou, B.; Li, Z.; et al. Comparative and parallel genome-wide association studies for metabolic and agronomic traits in cereals. Nat. Commun. 2016, 7, 1–10. [Google Scholar] [CrossRef]
- Chen, Z.; Chen, H.; Jiang, Y.; Wang, J.; Khan, A.; Li, P.; Cao, C. Metabolomic analysis reveals metabolites and pathways involved in grain quality traits of high-quality rice cultivars under a dry cultivation system. Food Chem. 2020, 326, 126845. [Google Scholar] [CrossRef] [PubMed]
- Chaves, F.C.; Broeckling, C.D. Metabolomics for rice grain quality. In The Future of Rice Demand: Quality beyond Productivity; Springer: Cham, Switzerland, 2020; pp. 495–531. [Google Scholar]
- Jang, Y.; Shin, G.; Jung, E.; Lee, S.; Lee, S.; Singh, D.; Jang, E.; Shin, D.; Kim, H.; Hye, W.S.; et al. Process specific differential metabolomes for industrial gochujang types (pepper paste) manufactured using white rice, brown rice, and wheat. Food Chem. 2017, 234, 416–424. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Guo, Y.; Wang, Y.; Li, M.; Li, K.; Liu, X.; Fang, C.; Luo, J. Metabolomic analysis reveals nutritional diversity among three staple crops and three fruits. Foods 2022, 11, 550. [Google Scholar] [CrossRef]
- Li, H.; Lv, Q.; Liu, A.; Wang, J.; Sun, X.; Deng, J.; Chen, Q.; Wu, Q. Comparative metabolomics study of Tartary (Fagopyrum tataricum L.) Gaertn) and common (Fagopyrum esculentum Moench) buckwheat seeds. Food Chem. 2022, 371, 131125. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Q.; Sun, C.; Li, A.; Zhang, J.; Shi, Q.; Zhang, Y.; Hu, J.; Zhou, N.; Wei, H.; Liu, B.; et al. Metabolomics and biochemical analyses revealed metabolites important for the antioxidant properties of purple glutinous rice. Food Chem. 2022, 389, 133080. [Google Scholar] [CrossRef]
- Chen, Z.; Li, P.; Xiao, J.; Jiang, Y.; Cai, M.; Wang, J.; Li, C.; Zhan, M.; Cao, C. Dry cultivation with ratoon system impacts rice quality using rice flour physicochemical traits, fatty and amino acids contents. Food Res. Int. 2021, 150, 110764. [Google Scholar] [CrossRef]
- Qiu, X.; Yang, J.; Zhang, F.; Niu, Y.; Zhao, X.; Shen, C.; Xu, J. Genetic dissection of rice appearance quality and cooked rice elongation by genome-wide association study. Crop J. 2021, 9, 1470–1480. [Google Scholar] [CrossRef]
- Zhou, H.; Xia, D.; Zhao, D.; Li, Y.; Li, P.; Wu, B.; Gao, G.; Zhang, Q.; Wang, G.; Xiao, J.; et al. The origin of Wxla provides new insights into the improvement of grain quality in rice. J. Integr. Plant Biol. 2021, 63, 878–888. [Google Scholar] [CrossRef]
- Zhang, Q.; Guo, B.; Hu, Y.; Zhang, H.; Xu, Y.; Xu, X.; Zhu, B.; Xu, J.; Niu, Z.; Tu, R. Differences in yield and rice quality of soft japonica rice with high quality and high yield under different nitrogen levels. Chin. J. Rice Sci. 2021, 35, 606–616. [Google Scholar]
- Lan, Y.; Guo, X.; Li, M.; Zhao, Y.; Li, X.; Jiang, H.; Wang, H.; Xu, L.; Zhang, X.; Lv, Y. Effects of Interaction between nitrogen application rate and transplanting density on Kenjing 7 rice quality. J. Agric. Sci. Technol. 2021, 23, 136–145. [Google Scholar]
- Wang, L.; Xie, B.; Gu, C.; Zang, J.; Du, J.; Na, Y. Effects of different nitrogen application amount and transplanting density on rice growth and qualities in cold region. Heilongjiang Agric. Sci. 2019, 7, 46–52. [Google Scholar]
- Ning, H.; Qiao, J.; Liu, Z.; Lin, Z.; Li, G.; Wang, Q.; Wang, S.; Ding, Y. Distribution of proteins and amino acids in milled and brown rice as affected by nitrogen fertilization and genotype. J. Cereal Sci. 2010, 52, 90–95. [Google Scholar] [CrossRef]
- Zhang, J.; Luo, W.; Zhao, Y.; Xu, Y.; Song, S.; Chong, K. Comparative metabolomic analysis reveals a reactive oxygen species-dominated dynamic model underlying chilling environment adaptation and tolerance in rice. New Phytol. 2016, 211, 1295–1310. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Chen, X.; Zhu, S.; Sun, M.; Zhou, X. Understanding the flavor signature of the rice grown in different regions of China via metabolite profiling. J. Sci. Food Agric. 2021, 102, 11641. [Google Scholar] [CrossRef]
- Verma, D.K.; Srivastav, P.P. Extraction, identification and quantification methods of rice aroma compounds with emphasis on 2-acetyl-1-pyrroline (2-AP) and its relationship with rice quality: A comprehensive review. Food Rev. Int. 2022, 38, 111–162. [Google Scholar] [CrossRef]
- Concepcion, J.C.T.; Calingacion, M.; Garson, M.J.; Fitzgerald, M.A. Lipidomics reveals associations between rice quality traits. Metabolomics 2020, 16, 1–16. [Google Scholar] [CrossRef]
- Wang, W.; Tu, Q.; Chen, R.; Lv, P.; Xu, Y.; Xie, Q.; Song, Z.; He, Y.; Cai, D.; Zhang, X. Polyploidization increases the lipid content and improves the nutritional quality of rice. Plants 2022, 11, 132. [Google Scholar] [CrossRef]
- Xu, G.; Liang, C.; Huang, P.; Liu, Q.; Xu, Y.; Ding, C.; Li, T. Optimization of rice lipid production from ultrasound-assisted extraction by response surface methodology. J. Cereal Sci. 2016, 70, 23–28. [Google Scholar] [CrossRef]
- Zarei, I.; Brown, D.G.; Nealon, N.J.; Ryan, E.P. Rice bran metabolome contains amino acids, vitamins & cofactors, and phytochemicals with medicinal and nutritional properties. Rice 2017, 10, 1–21. [Google Scholar]
- Chen, Y.; Zhou, S.; Wang, Z.; Wang, C.; Li, H.; Huang, D.; Zhou, D.; Zhao, L.; Gong, R.; Pan, Y. Difference of metabolites of grain and milled rice between ‘Meixiangzhan2’ and ‘Qixinzhan’ with good eating quality. Mol. Plant Breed. 2021, 19, 8255–8264. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Treatment | Brown Rice Rate (%) | Milled Rice Rate (%) | Head Milled Rice Rate (%) | Amylose Content (%) | Appearance | Hardness | Viscosity | Balance Degree | Taste Value |
|---|---|---|---|---|---|---|---|---|---|
| LNHD | 84.46a | 63.54b | 44.72b | 10.71a | 8.93a | 5.33b | 9.03a | 8.93a | 86.30a |
| HNLD | 84.38a | 72.34a | 59.23a | 10.45a | 8.43a | 5.73a | 8.67a | 8.50a | 82.33b |
| MNMD | 84.87a | 71.00a | 55.67a | 9.60a | 8.80a | 5.30b | 8.70a | 8.77a | 84.80ab |
| Pathway Description | Pathway ID | Ratio_in_Pop | p-Value |
|---|---|---|---|
| KEGG pathways between HNLD and LNHD | |||
| Beta-alanine metabolism | map00410 | 32/4803 | 0.0264 |
| Alpha-linolenic acid metabolism | map00592 | 44/4803 | 0.0362 |
| Linoleic acid metabolism | map00591 | 28/4803 | 0.0231 |
| Fatty acid biosynthesis | map00061 | 58/4803 | 0.0474 |
| Ether lipid metabolism | map00565 | 25/4803 | 0.0207 |
| Glycerophospholipid metabolism | map00564 | 52/4803 | 0.0007 |
| KEGG pathways between HNLD and MNMD | |||
| Phenylalanine, tyrosine, and tryptophan biosynthesis | map00400 | 34/4803 | 0.0553 |
| Alpha-linolenic acid metabolism | map00592 | 44/4803 | 0.071 |
| Ether lipid metabolism | map00565 | 25/4803 | 0.0409 |
| Sphingolipid metabolism | map00600 | 25/4803 | 0.0409 |
| KEGG pathways between LNHD and MNMD | |||
| Phosphatidylinositol signaling system | map04070 | 29/4803 | 0.0357 |
| Inositol phosphate metabolism | map00562 | 47/4803 | 0.0573 |
| Sphingolipid metabolism | map00600 | 25/4803 | 0.0308 |
| Galactose metabolism | map00052 | 46/4803 | 0.0561 |
| Fatty acid biosynthesis | map00061 | 58/4803 | 0.0021 |
| Linoleic acid metabolism | map00591 | 28/4803 | 0.0345 |
| Alpha-linolenic acid metabolism | map00592 | 44/4803 | 0.0537 |
| Glycerophospholipid metabolism | map00564 | 52/4803 | 0.0017 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiong, Q.; Sun, C.; Shi, H.; Cai, S.; Xie, H.; Liu, F.; Zhu, J. Analysis of Related Metabolites Affecting Taste Values in Rice under Different Nitrogen Fertilizer Amounts and Planting Densities. Foods 2022, 11, 1508. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11101508
Xiong Q, Sun C, Shi H, Cai S, Xie H, Liu F, Zhu J. Analysis of Related Metabolites Affecting Taste Values in Rice under Different Nitrogen Fertilizer Amounts and Planting Densities. Foods. 2022; 11(10):1508. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11101508
Chicago/Turabian StyleXiong, Qiangqiang, Changhui Sun, Hong Shi, Shuo Cai, Hengwang Xie, Fangping Liu, and Jinyan Zhu. 2022. "Analysis of Related Metabolites Affecting Taste Values in Rice under Different Nitrogen Fertilizer Amounts and Planting Densities" Foods 11, no. 10: 1508. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11101508