1. Introduction
Ultra-high temperature (UHT) processing of milk is utilized to ensure product safety during long shelf-life storage. However, the severe heat treatment induces unwanted changes to the dairy matrix and its components, which can produce molecular processing-induced changes in the proteins and affect their physiochemical properties. These protein modifications occur upon heat treatment; however, their severity extends during storage. The induced protein changes can influence the nutritional value of the products, as well as lead to unwanted off-flavors and color changes, and can potentially contribute to sedimentation or age gelation [
1]. Age gelation is the formation of an irreversible three-dimensional protein network induced by proteolytic enzymes (either endogenous or exogenous) and/or physicochemical changes occurring during storage. On the other hand, sedimentation is the formation of a compact protein-rich layer at the bottom of the product. Depending on fat content, fat aggregation can also occur in the top layer. These phenomena have been well-documented as occurring during the storage of UHT milk, but the underlying molecular mechanisms are still not fully understood. Recently, it was found that UHT milk prepared from raw milk containing large casein micelles generated more sediment during a 6-month storage period at 40 °C than UHT milk prepared from milk with smaller casein micelles or stored at a lower temperature [
2,
3]. It was suggested that rearrangements involving dissociation of β-lactoglobulin-κ-casein complexes from the micelles in relation to UHT treatment and storage could be one driving force, leading to sedimentation of the shaved micelles no longer repulsed by protruding, negative κ-casein tails [
3,
4]. In addition, proteolysis resulting in κ-casein liberation from the casein micelle was also thought to play a role in sediment formation [
3]. However, these studies also suggested that factors other than protein degradation might be involved. Differences in sedimentation between milk pools from raw milk with different casein micelle sizes could indicate an influence of casein micelle density on sedimentation, while a higher sedimentation at high storage temperatures might be due to an increase of protein modifications due to cross-linking of proteins and Maillard reaction [
2]. Hence, it was suggested that further study of protein modifications was necessary, in order to fully understand the differences in sedimentation between small and large native casein micelles [
2,
3].
There are currently two main pathways identified as causing formation of protein cross-linking in processed milk: the Maillard reaction and the dehydroalanine (DHA) pathway. The Maillard reaction is a complex chain of reactions that is initiated between an amine group on a protein/peptide and a reducing sugar, resulting in the formation of different aroma compounds, advanced glycation end products (AGEs) and a large group of cross-linked products, known as melanoidins [
5]. In the early stage of Maillard reaction, a condensation reaction is initiated between a carbonyl group in the open chain form of a sugar and a nucleophilic amino group (lysine residues or free N-terminals), resulting in the formation of a Schiff-base, which then rearranges into the more stable Amadori product. In the advanced stage of the Maillard reaction, the Amadori product can be degraded to a reactive α-dicarbonyl compound, which can later react with free amino acids to form Strecker aldehydes, or with nucleophilic side chains of amino acids bound in peptides or proteins to form different AGEs. Major AGEs previously identified in UHT milk are N-ε-(carboxyethyl)lysine (CEL) and N-ε-(carboxymethyl)lysine (CML) which are peptide- or protein-bound lysine modifications derived from the reaction of the α-dicarbonyls glyoxal and methylglyoxal with a single lysine residue [
6]. The reaction of glyoxal and methylglyoxal with two lysine residues bound in the sequence of peptides or proteins will form the inter or intra-protein cross-links, known as glyoxal-lysine dimers (GOLD) and methylglyoxal-lysine dimers (MOLD). Instead of degradation, compounds derived from the Amadori product can suffer cyclation and enolization, forming β-pyranone and 3-furanone. Further isomerization of β-pyranone, and its reaction with proteins, is speculated to be an alternative protein cross-linking formation mechanism [
7,
8].
On the other hand, the sugar-independent DHA pathway can lead to aggregate formation by covalent cross-linking between specific residues. During the initial stage, either serine, phosphoserine, glycoserine, cysteine or cystine residues within protein chains can undergo a heat- or alkali-induced β-elimination reaction to form the unstable DHA compound. After rearrangement, the alkene of DHA can further react with lysine, cysteine or histidine interchain or intrachain amino acid residues to form covalent cross-links known as lysinoalanine (LAL), lanthionine (LAN) or histidinoalanine (HAL), respectively [
9]. Both Maillard reaction and DHA-driven protein cross-linking are known to increase with the intensity of the heat treatment [
10], and during storage [
5,
6].
New methods for quantification of processing-induced markers have been developed in recent years. The use of a liquid chromatography triple quadrupole based method (LC-MS triple Q) with multiple reaction monitoring (MRM) has proven to be a fast and reliable method for LAL and LAN quantification [
11]. The prospect of using this novel, quantitative method for simultaneous quantification of multiple processing-induced markers is a new milestone in the study of protein cross-linking in foods. Recently, targeted MS analysis has been successfully used in the study of lactose influence on Maillard reaction and DHA-mediated protein cross-linking in caseins and whey proteins [
10]. It has also been used to study these processing-induced changes in different food products and plasma [
12]. This method enables simultaneous quantification of the development of both sugar-dependent and sugar-independent covalent processing-induced markers during storage of UHT-treated milk. This information is key to the research of the main mechanisms driving aggregate formation and sedimentation during milk storage.
In the present study, it is hypothesized that processing-induced markers are higher in milk with increased levels of sedimentation. To investigate this, samples from a previously conducted storage study [
2,
3] were analyzed by the MRM-based method for absolute quantification of processing-induced protein modifications.
4. Discussion
A targeted MS analysis for absolute quantification of processing-induced protein modifications was applied to a storage experiment carried out for UHT-treated milk prepared from raw milk pools representing small or large casein micelles. The level of sedimentation was previously found to be higher in UHT milk prepared from raw milk representing large casein micelles, and to increase with storage [
2,
3]. The present study therefore investigated whether the level of processing-induced markers was higher in milk with increased levels of sedimentation. The quantification of processing-induced protein modifications in mol per mol of protein used in this study allowed for a more precise comparison between the modifications. The use of this unit enabled a fairer comparison, since it standardized the individual molecular weight of each compound, therefore avoiding processing-markers with a high molecular weight (e.g., furosine) to be overrepresented.
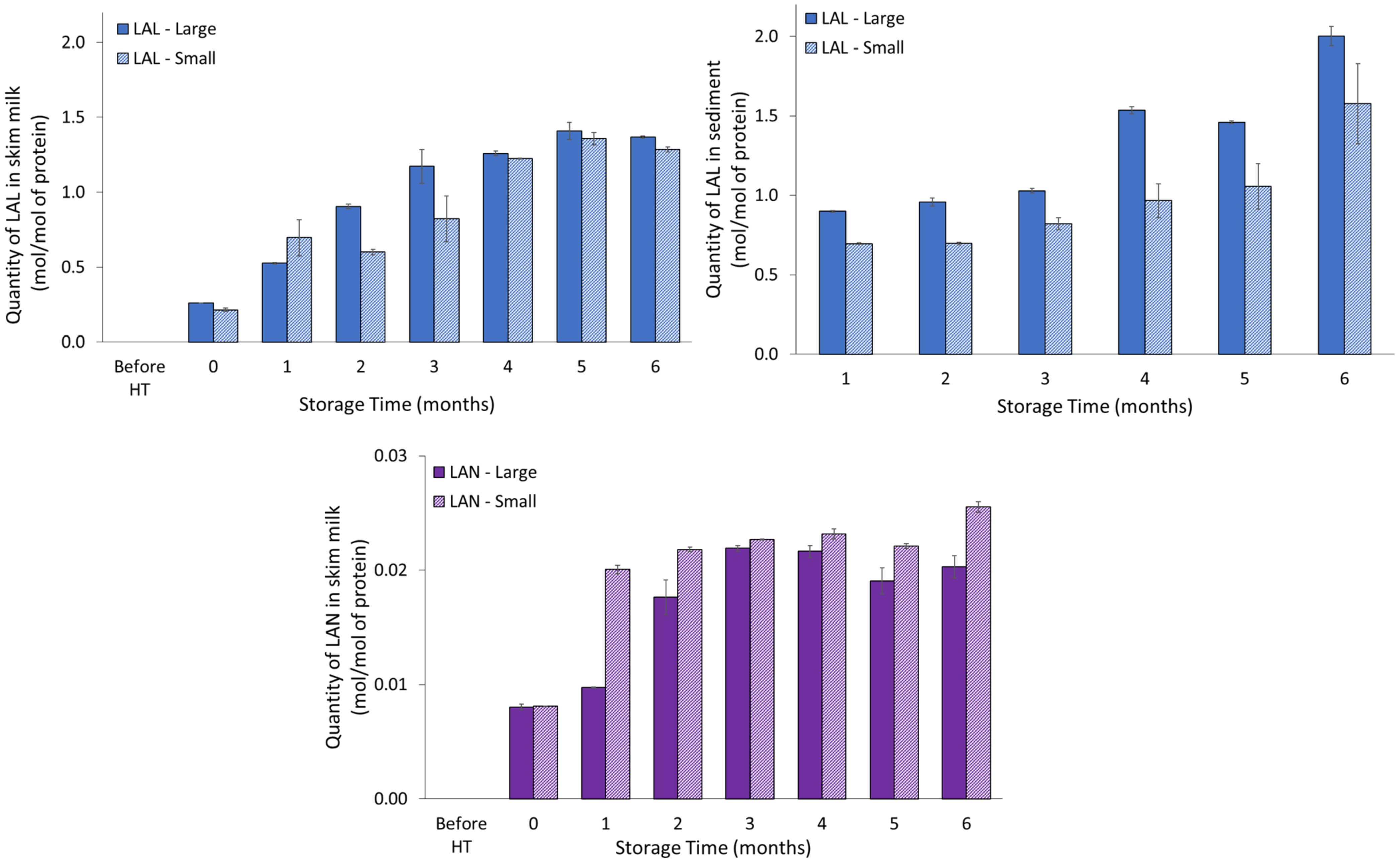
It was found that the molar proportion of furosine increased in both skim milk and sediment with storage, and that it differed in sediment relative to milk pool, with higher proportion in sediment made from UHT milk prepared from raw milk with large casein micelles. However, the furosine level at a molar basis relative to moles of protein was much higher in skim milk as compared with sediment. Both CEL and CML increased in skim milk during storage, though CEL levels in skim milk were very low. In sediment, the level of CEL was fairly constant, and higher than that of skim milk. Furthermore, from three to six months of storage, the level of CML in sediment increased, reaching levels twice as high as those observed in skim milk. The levels of both CEL and CML were higher in sediment derived from milk with large casein micelles compared to milk with small casein micelles. As furosine is an early marker for Maillard reaction, it is also possible that the furosine formed had been converted into later Maillard reaction products, as represented by the quantity of CEL and CML. These results indicate that Maillard-related processes, as represented by both CEL and CML, can be either a contributing driving force, through generation of protein-protein cross-links within or between milk proteins, or a consequence of sedimentation and protein up-concentration in the sediment [
4]. These two mechanisms cannot be discriminated by the design of the present study.
The molar level of LAL was somewhat higher in the sediment compared to the skim milk, and UHT milk representing milk with larger casein micelles showed a more pronounced degree of LAL formation compared to UHT milk representing milk with smaller casein micelles. Therefore, DHA-mediated cross-linking represented by LAL could, in addition to Maillard reaction, be a potential contributing factor to sediment formation, through inter-protein crosslinking or the formation of LAL could be a direct consequence of the unique characteristics of the sedimentation. LAL was much more pronounced in skim milk compared to LAN, and LAN was only detected below the limit of quantification in sedimentation. As LAN is the cross-linking product of DHA and cysteine, the result may reflect that the sediments are predominantly composed of α
S1- and β-casein, which do not contain any cysteine residues and only small amounts of whey proteins [
3,
4,
14].
Since Akkerman, et al., [
3] observed that milk with large casein micelles produced higher amounts of sediment during storage, a correlation between quantity of processing-induced markers and sediment formation is suggested by the present study. The effect of protein cross-linking in milk spoilage has been previously discussed. Andrews & Cheeseman [
15] suggested that cross-linked proteins due to Maillard reactions aggregate into high-molecular-weight protein aggregates, and drive gelation of UHT milk during storage. On the other hand, some studies have shown that the level of reducing sugars has no effect on milk age gelation [
1,
4]. However, in this study it was found that milk sediment was associated with late-stage Maillard reaction, which is known to be related to cross-linking [
7,
8]. Additionally, while Maillard reaction has been widely studied and discarded as the solely causative agent of gelation, none of the previously mentioned approaches considered the influence of sugar-independent cross-link products, such as LAL, which is suggested by the present study to play a role as well.
Other factors partly contributing to sedimentation have also previously been discussed [
2]. One such account relates to Stokes’ law [
16], stating that particles will move according to their size and density over time. However, it has been suggested that this phenomenon may not easily be applied to the complex system of homogenized and UHT-treated milk [
2]. Another account related to the dissociation rate of β-lactoglobulin-κ-casein complexes, as the relative distribution of total κ-casein in the untreated milk was not different between milk representing large and small casein micelles [
2] which was also observed in other studies [
17,
18,
19]. Presumably, this means that the surface of micelles in milk with large casein micelles are less covered by κ-casein compared to milk with smaller casein micelles. A looser structure of the casein micelles can ease proteolysis and dissociation of the of β-lactoglobulin-κ-casein complexes, and as a result lead to increased level of sediment in milk with large casein micelles. Furthermore plasmin activity in the milk has been related to casein micelle size, which supports a previous study showing that c-terminal-derived κ-casein peptides correlated with the level of sediment in the milk [
3]. This indicates that dissociation of κ-casein from the casein micelle may destabilize the casein micelle, causing formation of sediment [
3]. The level of sediment could also relate to pH and ionic calcium levels, as a previous study observed that ionic calcium above 1.5 mM and pH below 6.7 resulted in increased levels of sediment. However, a relationship between pH and ionic calcium could not be confirmed in our previous study [
2]. Sedimentation in UHT milk is likely induced through the synergy of several mechanisms, where Maillard reaction and DHA-mediated crosslinking could potentially contribute.
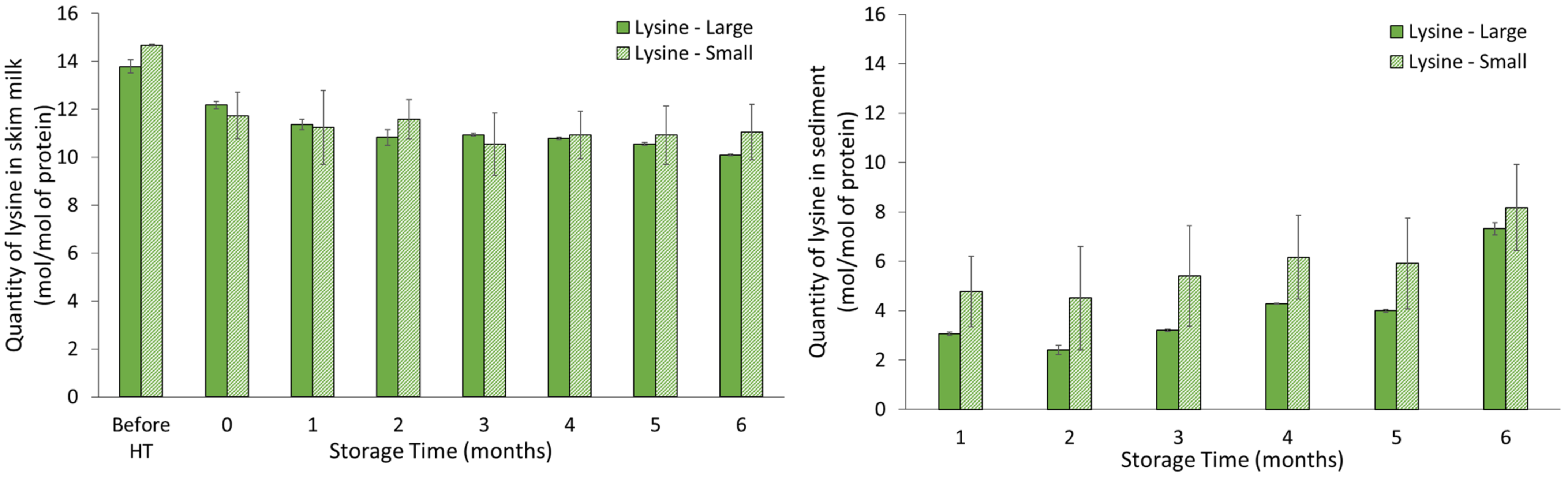
The level of lysine, determined by the MRM-based MS method, represented lysine originating from proteins, peptides and free lysine, as the method relies on complete hydrolysis of proteins into amino acid residues and their derivatives using acid hydrolysis. The result showed that the percentage of lysine did not show any significant difference during storage. However, a large difference in concentration between skim milk and sediment was observed, although the molar concentration was adjusted for protein concentration. One explanation could be that a much higher formation of processing-induced protein modifications existed in the sediment. These could include modifications not analyzed in the MRM method applied in this study, such as the late-stage Maillard reaction products GOLD and MOLD, which have previously been identified in UHT milk [
6], and which use lysine as a substrate for their reaction. The level of lysine was lower in sediment from milk representing larger casein micelles, and therefore supports the higher level of processing-induced protein modification in milk with larger casein micelles compared with milk with smaller casein milk.
The analytical method used in the present study is highly sensitive. However, there is a limitation to both detection and quantification of the compounds in these samples. Although both CEL and LAN was detected in sediment samples, they were not able to be quantified, as their concentration levels were below the quantitative range of the MS instrument, which was 3.9 ng/mL. Below this, the quantitative accuracy of the analytical standard was outside the defined threshold of 15% [
10].
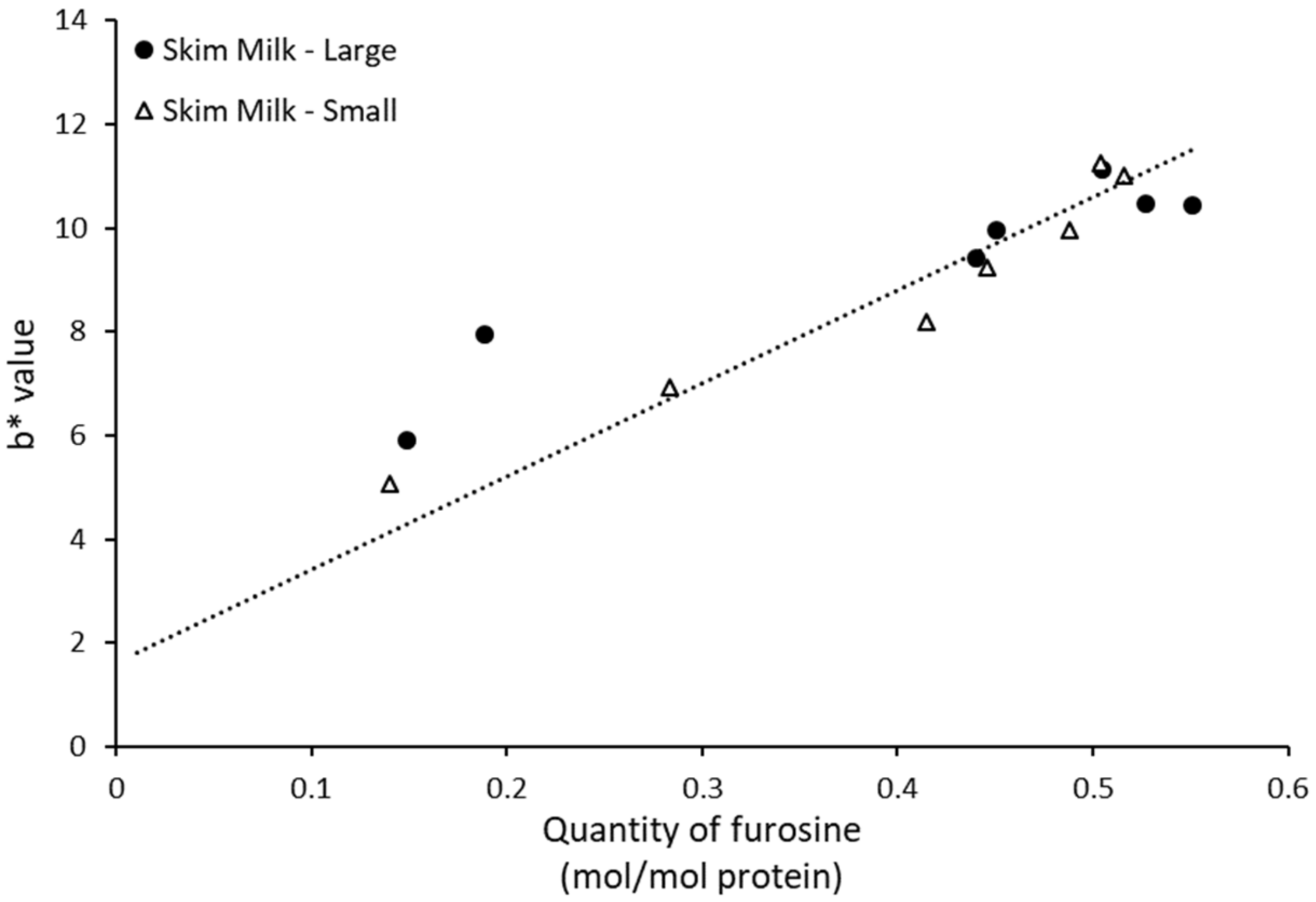
As furosine is a relevant indicator for discerning the onset of Maillard reaction, methods to quickly and reliably measure furosine formation have been widely discussed. Some studies have showcased a potential correlation between furosine and browning, more specifically measurement of b* value [
20]. Lysine-derived compounds (such as furosine) have been shown in previous studies to have a higher impact on browning than other amino acid products [
21]. In this study, it was possible to corroborate a direct relation between b* value and furosine concentration, especially after the second month of storage. These results are in accordance with those observed in
Figure 1, where furosine content was shown to be the main Maillard reaction product in skim milk, especially for the last months of storage, hence being the main driver of browning.


 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}