
Protein Recovery of Tra Catfish (Pangasius hypophthalmus) Protein-Rich Side Streams by the pH-Shift Method

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
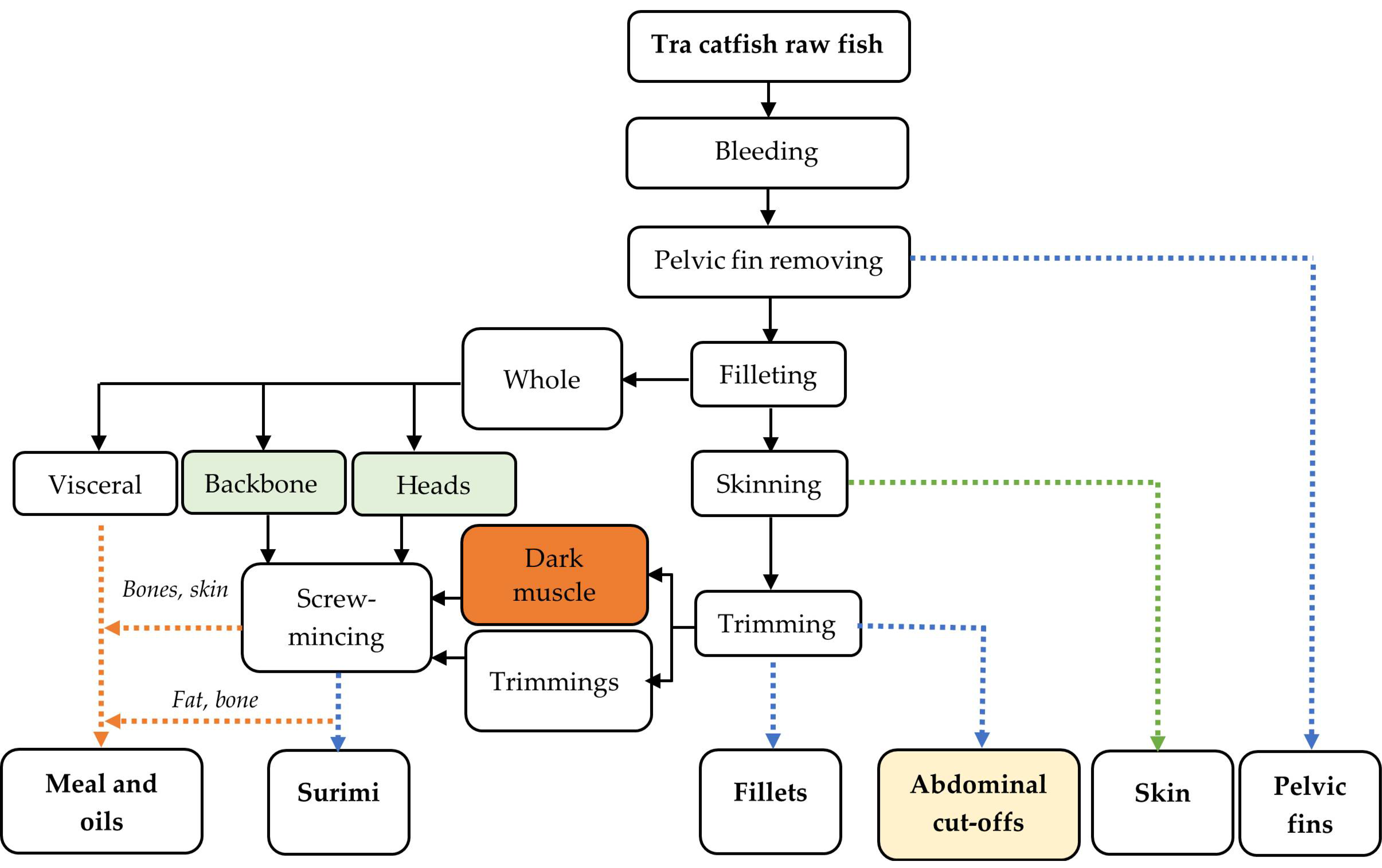
2.1. Raw Materials and Sampling
2.1.1. Collection of Side Streams
2.1.2. Chemicals
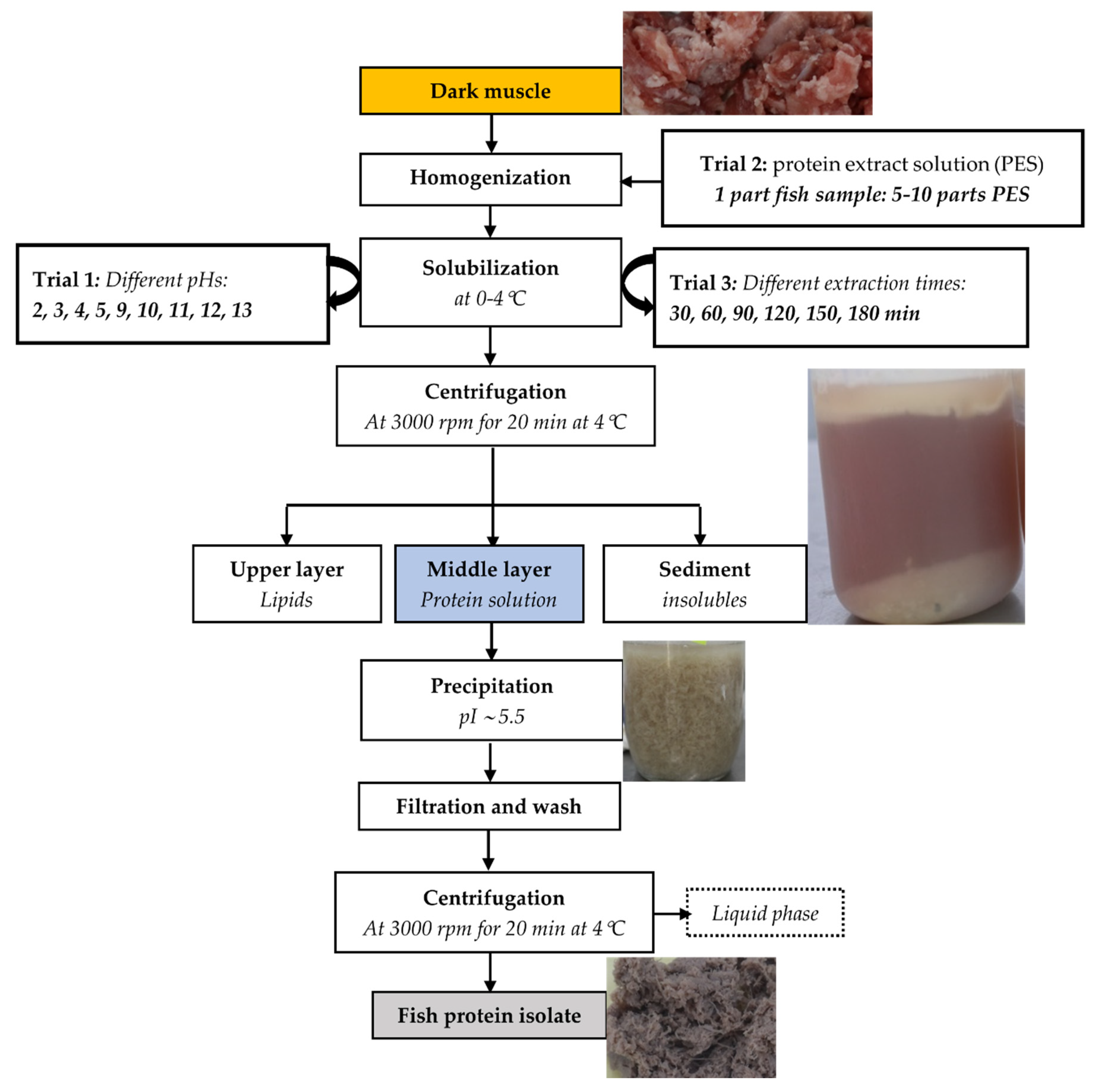
2.1.3. Preparation of Protein Isolate from the Dark Muscle
2.1.4. FPI Preparation from Different Side Streams at the Optimal Conditions
2.2. Analyses
2.2.1. Proximate Composition
2.2.2. Protein Extractable Recovery (PER), FPI Protein, and FPI Dry Matter Recoveries
2.2.3. Lipid and Ash Removal during FPI Production
2.2.4. Amino Acid Analysis
2.2.5. SDS-PAGE Pattern
2.2.6. Colour
2.2.7. Statistical Analysis
3. Results and Discussion
3.1. Optimization of Protein Recovery from Dark Muscle
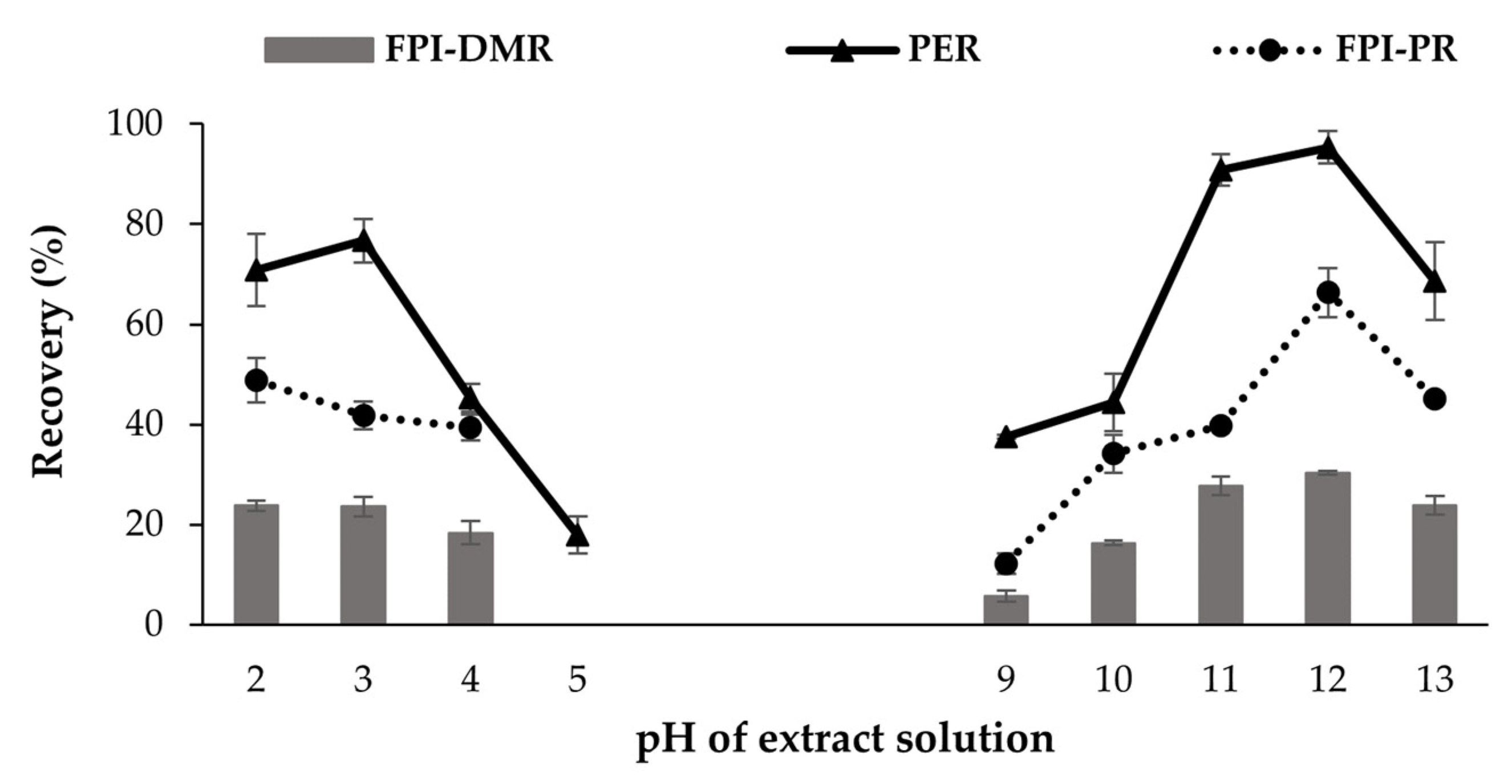
3.1.1. Effects of pH on Extraction Levels (Trial 1)
3.1.2. Effects of Using Different Proportions of Extract Solution (Trial 2)
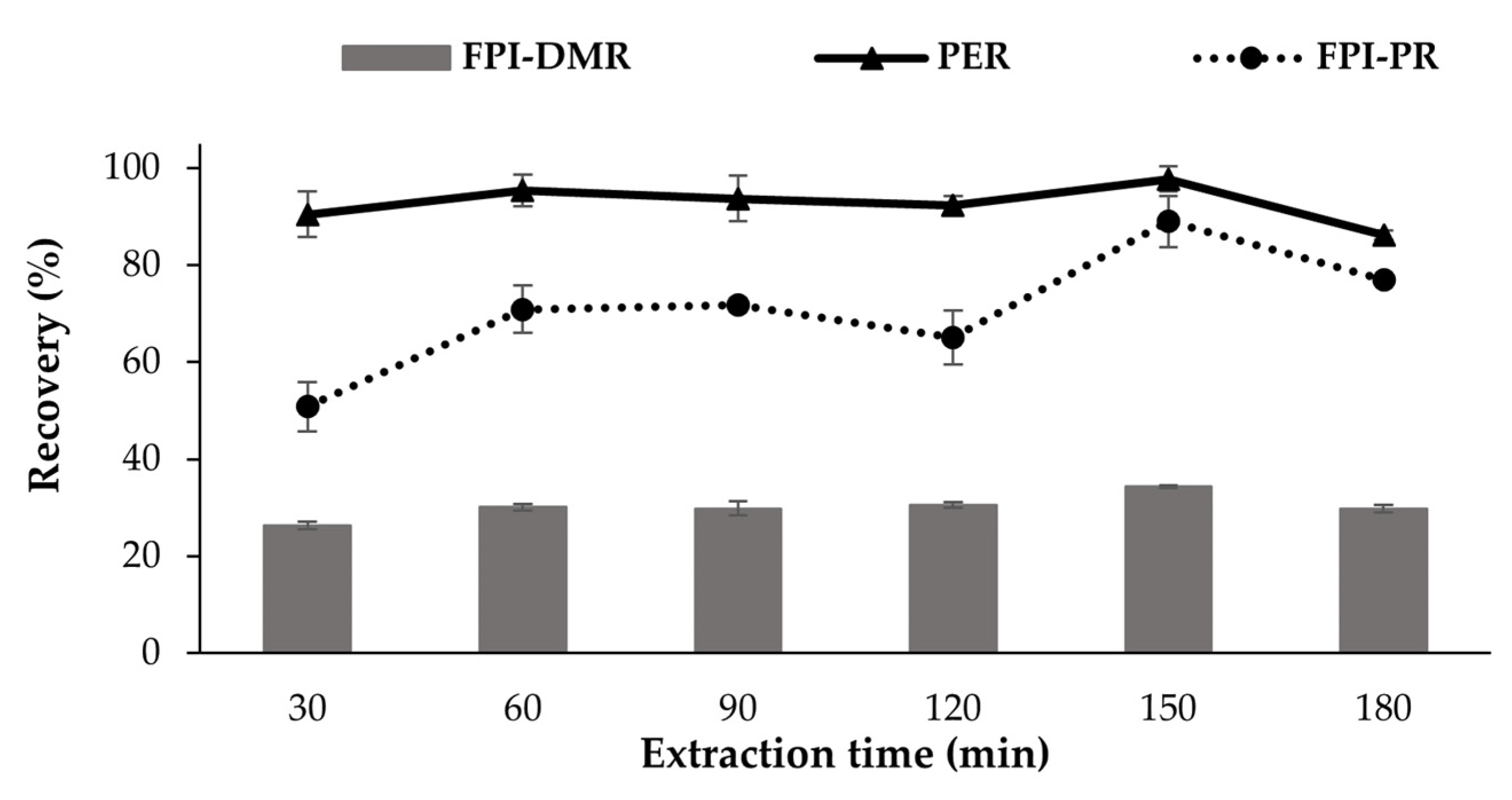
3.1.3. The Effects of Extraction Time (Trial 3)
3.2. Comparison of the FPI Produced from Different Rest Raw Materials and Surimi
3.2.1. FPI Processing Effectivity
3.2.2. Proximate Composition
3.2.3. Amino Acid Profiles
3.2.4. SDS-PAGE
3.2.5. Colour
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Worm, B.; Branch, T.A. The future of fish. Trends Ecol. Evol. 2012, 27, 594–599. [Google Scholar] [CrossRef] [PubMed]
- Costello, C.; Ovando, D.; Clavelle, T.; Strauss, C.K.; Hilborn, R.; Melnychuk, M.C.; Branch, T.A.; Gaines, S.D.; Szuwalski, C.S.; Cabral, R.B. Global fishery prospects under contrasting management regimes. Proc. Natl. Acad. Sci. USA 2016, 113, 5125–5129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McClanahan, T.; Allison, E.H.; Cinner, J.E. Managing fisheries for human and food security. Fish Fish. 2015, 16, 78–103. [Google Scholar] [CrossRef]
- Arason, S.; Karlsdottir, M.; Valsdottir, T.; Slizyte, R.; Rustad, T.; Falch, E.; Eysturskard, J.; Jakobsen, G. Maximum Resource Utilisation-Value Added Fish by-Products; Nordic Innovation Centre: Oslo, Norway, 2010. [Google Scholar]
- Shahidi, F.; Varatharajan, V.; Peng, H.; Senadheera, R. Utilization of marine by-products for the recovery of value-added products. J. Food Bioact. 2019, 6, 6–61. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.-K.; Perera, U.; Rajapakse, N.; Kim, S. Seafood Processing by-Products; Springer: New York, NY, USA, 2014. [Google Scholar]
- Wu, T.H.; Nigg, J.D.; Stine, J.J.; Bechtel, P.J. Nutritional and chemical composition of by-product fractions produced from wet reduction of individual red salmon (Oncorhynchus nerka) heads and viscera. J. Aquat. Food Prod. Technol. 2011, 20, 183–195. [Google Scholar] [CrossRef]
- Stevens, J.R.; Newton, R.W.; Tlusty, M.; Little, D.C. The rise of aquaculture by-products: Increasing food production, value, and sustainability through strategic utilisation. Mar. Policy 2018, 90, 115–124. [Google Scholar] [CrossRef] [Green Version]
- Ghaly, A.; Ramakrishnan, V.; Brooks, M.; Budge, S.; Dave, D. Fish processing wastes as a potential source of proteins, amino acids and oils: A critical review. J. Microb. Biochem. Technol. 2013, 5, 107–129. [Google Scholar]
- Rustad, T.; Storrø, I.; Slizyte, R. Possibilities for the utilisation of marine by-products. Int. J. Food Sci. 2011, 46, 2001–2014. [Google Scholar] [CrossRef]
- Oliveira, A.; O’Keefe, S.; Balaban, M. Fillet yields and proximate composition of cultured Gulf of Mexico sturgeon (Ancipenser oxyrynchus desotoi). J. Aquat. Food Prod. Technol. 2005, 14, 5–16. [Google Scholar] [CrossRef]
- Undeland, I. Chapter 11-Oxidative Stability of Seafood. In Oxidative Stability and Shelf Life of Foods Containing Oils and Fats; Hu, M., Jacobsen, C., Eds.; AOCS Press: Urbana, IL, USA, 2016; pp. 391–460. [Google Scholar] [CrossRef]
- Tokur, B.; Korkmaz, K. The effects of an iron-catalyzed oxidation system on lipids and proteins of dark muscle fish. Food Chem. 2007, 104, 754–760. [Google Scholar] [CrossRef]
- Dang, H.T.T.; Gudjónsdóttir, M.; Karlsdóttir, M.G.; Van Nguyen, M.; Romotowska, P.E.; Tómasson, T.; Arason, S. Influence of temperature stress on lipid stability of Atlantic herring (Clupea harengus) muscle during frozen storage. J. Am. Oil Chem. Soc. 2017, 94, 1439–1449. [Google Scholar] [CrossRef]
- Elena, S.Z.; Mahassine, A.; Rosa, O.; Casilda, N.; Juana, F.L.; Esther, S.; Estrella, S.; José Angel, P.A. Quality characteristics of dark muscle from yellowfin tuna Thunnus albacares to its potential application in the food industry. Food Sci. Nutr. 2011, 2, 20–30. [Google Scholar]
- Herpandi, N.H.; Rosma, A.; Wan Nadiah, W. The tuna fishing industry: A new outlook on fish protein hydrolysates. Compr. Rev. Food Sci. Food Saf. 2011, 10, 195–207. [Google Scholar] [CrossRef]
- Kristinsson, H.G.; Lanier, T.C.; Halldorsdottir, S.M.; Geirsdottir, M.; Park, J.W. Fish protein isolate by pH shift. In Surimi and Surimi Seafood, 3rd ed.; Park, J.W., Ed.; CRC Press: Boca Raton, FL, USA, 2013; pp. 169–192. [Google Scholar]
- Saidi, S.; Deratani, A.; Belleville, M.-P.; Amar, R.B. Antioxidant properties of peptide fractions from tuna dark muscle protein by-product hydrolysate produced by membrane fractionation process. Food Res. Int. 2014, 65, 329–336. [Google Scholar] [CrossRef]
- Nam, P.V.; Van Hoa, N.; Anh, T.T.L.; Trung, T.S. Towards Zero-Waste Recovery of Bioactive Compounds from Catfish (Pangasius Hypophthalmus) By-Products Using an Enzymatic Method. Waste Biomass Valorization 2020, 11, 4195–4206. [Google Scholar] [CrossRef]
- FAO. The State of World Fisheries and Aquaculture 2020. Sustainability in Action; FAO: Rome, Italy, 2020; p. 75. [Google Scholar]
- FAO. The State of World Fisheries and Aquaculture 2018. Meeting the Suitable Development Goals; FAO: Rome, Italia, 2018. [Google Scholar]
- Thong, N.T.; Ankamah-Yeboah, I.; Bronnmann, J.; Nielsen, M.; Roth, E.; Schulze-Ehlers, B. Price transmission in the pangasius value chain from Vietnam to Germany. Aquacult. Rep. 2020, 16, 100266. [Google Scholar] [CrossRef]
- Thuy, C.X.; Lam, T.B.; Mc. Commick, K. Biochemical and functional properties of fish protein isolate (FPI) from pangasius hypophthalmus byproducts as influenced by time and degree of hydrolysis (DH). Int. Food Res. J. 2015, 22, 337–343. [Google Scholar]
- Bechtel, P.J.; Watson, M.A.; Lea, J.M.; Bett-Garber, K.L.; Bland, J.M. Properties of bone from Catfish heads and frames. Food Sci. Nutr. 2019, 7, 1396–1405. [Google Scholar] [CrossRef] [Green Version]
- Thuy, N.T.; Loc, N.T.; Lindberg, J.; Ogle, B. Survey of the production, processing and nutritive value of catfish by-product meals in the Mekong Delta of Vietnam. Livest. Res. Rural. Dev. 2007, 19, 124. [Google Scholar]
- Sørensen, N.K. Slaughtering Processes for Farmed Pangasius in Vietnam. Consultancy Surveying Pangasius Fillet Quality and by-Products Handling in Vietnamese Industry; 8272515628; Norwegian Institute of Fisheries and Aquaculture Research: Tromso, Norway, 2005. [Google Scholar]
- Thuy, N.T. Effects of adding herbal powders to diets containing Tra catfish (Pangasius hypophthalmus) by-products on performance and health status of local chickens. Livest. Res. Rural. Dev. 2019, 31, 0. [Google Scholar]
- Thuy, N.T.; Ha, N.C. Effect of replacing marine fish meal with catfish (Pangasius hypophthalmus) by-product protein hydrolyzate on the growth performance and diarrhoea incidence in weaned piglets. Trop. Anim. Health Prod. 2016, 48, 1435–1442. [Google Scholar] [CrossRef] [PubMed]
- Rurangwa, E.; Baumgartner, U.; Nguyen, H.; van de Vis, J. Aquaculture Innovation in Vietnam; Wageningen Marine Research: Yerseke, The Netherlands, 2016. [Google Scholar]
- Nhu, T.T.; Schaubroeck, T.; Henriksson, P.J.; Bosma, R.; Sorgeloos, P.; Dewulf, J. Environmental impact of non-certified versus certified (ASC) intensive Pangasius aquaculture in Vietnam, a comparison based on a statistically supported LCA. Environ. Pollut. 2016, 219, 156–165. [Google Scholar] [CrossRef] [PubMed]
- Guenneugues, P.; Ianelli, J. Surimi resources and market. In Surimi and Surimi Seafood, 3rd ed.; Park, J.W., Ed.; CRC Press: Boca Raton, FL, USA, 2013; pp. 25–53. [Google Scholar]
- Tanuja, S.; Viji, P.; Zynudheen, A.; Joshy, C. Composition, functional properties and antioxidative activity of hydrolysates prepared from the frame meat of Striped catfish (Pangasianodon hypophthalmus). Egypt. J. Exp. Biol. 2012, 14, 27–35. [Google Scholar] [CrossRef]
- Hassan, M.A.; Xavier, M.; Gupta, S.; Nayak, B.B.; Balange, A.K. Antioxidant properties and instrumental quality characteristics of spray dried Pangasius visceral protein hydrolysate prepared by chemical and enzymatic methods. Environ. Sci. Pollut. Res. 2019, 26, 8875–8884. [Google Scholar] [CrossRef] [PubMed]
- Tanuja, S.; Haridas, A.; Zynudheen, A.; Joshy, C. Functional and antioxidative properties of fish protein hydrolysate (FPH) produced from the frame meat of striped catfish Pangasianodon hypophthalmus (Sauvage, 1878) using alkaline protease alcalase. Indian J. Fish. 2014, 61, 82–89. [Google Scholar]
- Baehaki, A.; Nopianti, R.; Anggraeni, S. Antioxidant activity of skin and bone collagen hydrolyzed from striped catfish (Pangasius pangasius) with papain enzyme. J. Chem. Pharm. Res. 2015, 7, 131–135. [Google Scholar]
- Singh, P.; Benjakul, S.; Maqsood, S.; Kishimura, H. Isolation and characterisation of collagen extracted from the skin of striped catfish (Pangasianodon hypophthalmus). Food Chem. 2011, 124, 97–105. [Google Scholar] [CrossRef]
- Hultin, H.; Kristinsson, H.; Lanier, T.; Park, J. Process for recovery of functional proteins by pH shifts. In Surimi and Surimi Seafood; Park, J.W., Ed.; CRC Press: Boca Raton, FL, USA, 2005; pp. 107–139. [Google Scholar]
- Shaviklo, G.R. Evaluation and Utilisation of Fish Protein Isolate Products. Master’s Thesis, University of Iceland, Reykjavik, Iceland, 2008. [Google Scholar]
- Hultin, H.O.; Kelleher, S.D. High Efficiency Alkaline Protein Extraction. U.S. Patent No 6,136,959, 24 October 2000. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Surasani, V.K.R.; Kudre, T.; Ballari, R.V. Recovery and characterization of proteins from pangas (Pangasius pangasius) processing waste obtained through pH shift processing. Environ. Sci. Pollut. Res. 2018, 25, 11987–11998. [Google Scholar] [CrossRef]
- Tadpitchayangkoon, P.; Yongsawatdigul, J. Comparative study of washing treatments and alkali extraction on gelation characteristics of striped catfish (Pangasius hypophthalmus) muscle protein. J. Food Sci. 2009, 74, C284–C291. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Mæhre, H.K.; Dalheim, L.; Edvinsen, G.K.; Elvevoll, E.O.; Jensen, I.-J. Protein determination-Method matters. Foods 2018, 7, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Abdollahi, M.; Marmon, S.; Chaijan, M.; Undeland, I. Tuning the pH-shift protein-isolation method for maximum hemoglobin-removal from blood rich fish muscle. Food Chem. 2016, 212, 213–224. [Google Scholar] [CrossRef]
- Surasani, V.K.R.; Tyagi, A.; Kudre, T. Recovery of proteins from rohu processing waste using pH shift method: Characterization of isolates. J. Aquat. Food Prod. Technol. 2017, 26, 356–365. [Google Scholar] [CrossRef]
- Zayas, J.F. Functionality of Proteins in Food; Springer: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Tian, Y.; Wang, W.; Yuan, C.; Zhang, L.; Liu, J.; Liu, J. Nutritional and digestive properties of protein isolates extracted from the muscle of the common carp using pH-shift processing. J. Food Process. Preserv. 2017, 41, e12847. [Google Scholar] [CrossRef] [Green Version]
- Abdollahi, M.; Undeland, I. Physicochemical and gel-forming properties of protein isolated from salmon, cod and herring by-products using the pH-shift method. LWT-Food Sci. Technol. 2019, 101, 678–684. [Google Scholar] [CrossRef]
- Tang, Z.X.; Ying, R.F.; Shi, L.E. Physicochemical and functional characteristics of proteins treated by a pH-shift process: A review. Int. J. Food Sci. 2021, 56, 515–529. [Google Scholar] [CrossRef]
- Kristinsson, H.G.; Theodore, A.E.; Demir, N.; Ingadottir, B. A comparative study between acid-and alkali-aided processing and surimi processing for the recovery of proteins from channel catfish muscle. J. Food Sci. 2005, 70, C298–C306. [Google Scholar] [CrossRef]
- Chomnawang, C.; Yongsawatdigul, J. Protein recovery of tilapia frame by-products by pH-shift method. J. Aquat. Food Prod. Technol. 2013, 22, 112–120. [Google Scholar] [CrossRef]
- Zhang, Y.; Chang, S.K. Protein extraction from catfish byproducts and physicochemical properties of the protein isolates. J. Food Sci. 2021, 86, 3061–3074. [Google Scholar] [CrossRef] [PubMed]
- Batista, I. Recovery of proteins from fish waste products by alkaline extraction. Eur. Food Res. Technol. 1999, 210, 84–89. [Google Scholar] [CrossRef]
- Kristinsson, H.G.; Liang, Y. Effect of pH-shift processing and surimi processing on Atlantic croaker (Micropogonias undulates) muscle proteins. J. Food Sci. 2006, 71, C304–C312. [Google Scholar] [CrossRef]
- Visschers, R.W.; de Jongh, H.H. Disulphide bond formation in food protein aggregation and gelation. Biotechnol. Adv. 2005, 23, 75–80. [Google Scholar] [CrossRef]
- Tadpitchayangkoon, P.; Park, J.W.; Yongsawatdigul, J. Conformational changes and dynamic rheological properties of fish sarcoplasmic proteins treated at various pHs. Food Chem. 2010, 121, 1046–1052. [Google Scholar] [CrossRef]
- Azadian, M.; Moosavi-Nasab, M.; Abedi, E. Comparison of functional properties and SDS-PAGE patterns between fish protein isolate and surimi produced from silver carp. Eur. Food Res. Technol. 2012, 235, 83–90. [Google Scholar] [CrossRef]
- Gehring, C.K.; Gigliotti, J.C.; Moritz, J.S.; Tou, J.C.; Jaczynski, J. Functional and nutritional characteristics of proteins and lipids recovered by isoelectric processing of fish by-products and low-value fish: A review. Food Chem. 2011, 124, 422–431. [Google Scholar] [CrossRef]
- Al Khawli, F.; Martí-Quijal, F.J.; Ferrer, E.; Ruiz, M.-J.; Berrada, H.; Gavahian, M.; Barba, F.J.; de la Fuente, B. Aquaculture and its by-products as a source of nutrients and bioactive compounds. In Advances in Food and Nutrition Research; Lorenzo, J.M., Barba, F.J., Eds.; Academic Press: Cambridge, MA, USA, 2020; Volume 92, pp. 1–33. [Google Scholar]
- Undeland, I.; Kristinsson, H.G.; Hultin, H.O. Hemoglobin-mediated oxidation of washed minced cod muscle phospholipids: Effect of pH and hemoglobin source. J. Agric. Food Chem. 2004, 52, 4444–4451. [Google Scholar] [CrossRef]
- Maqsood, S.; Benjakul, S.; Kamal-Eldin, A. Haemoglobin-mediated lipid oxidation in the fish muscle: A review. Trends. Food Sci. Technol. 2012, 28, 33–43. [Google Scholar] [CrossRef]
- Matak, K.E.; Tahergorabi, R.; Jaczynski, J. A review: Protein isolates recovered by isoelectric solubilization/precipitation processing from muscle food by-products as a component of nutraceutical foods. Food Res. Int. 2015, 77, 697–703. [Google Scholar] [CrossRef]
- Chaijan, M.; Panpipat, W. Removal of lipids, cholesterol, nucleic acids and haem pigments during production of protein isolates from broiler meat using pH-shift processes. Int. J. Food Eng. 2017, 13, 20160187. [Google Scholar] [CrossRef]
- Abdollahi, M.; Olofsson, E.; Zhang, J.; Alminger, M.; Undeland, I. Minimizing lipid oxidation during pH-shift processing of fish by-products by cross-processing with lingonberry press cake, shrimp shells or brown seaweed. Food Chem. 2020, 327, 127078. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.C.; Tou, J.C.; Jaczynski, J. Amino acid, fatty acid, and mineral profiles of materials recovered from rainbow trout (Oncorhynchus mykiss) processing by-products using isoelectric solubilization/precipitation. J. Food Sci. 2007, 72, C528–C536. [Google Scholar] [CrossRef]
- Xiong, Y. Muscle proteins. In Proteins in Food Processing, 2nd ed.; Rickey, Y.Y., Ed.; Woodhead Publishing: Duxford, UK, 2018; pp. 127–148. [Google Scholar]
- Marmon, S.K.; Undeland, I. Protein isolation from gutted herring (Clupea harengus) using pH-shift processes. J. Agric. Food Chem. 2010, 58, 10480–10486. [Google Scholar] [CrossRef] [PubMed]
- FAO. Protein and Amino Acid Requirements in Human Nutrition: Report of a Joint FAO/WHO/UNU Expert Consultation; No. 935; The World Health Organization: Geneva, Switzerland, 2007. [Google Scholar]
- Stone, A.; Stanley, D. Mechanisms of fish muscle gelation. Int. Food Res. J. 1992, 25, 381–388. [Google Scholar] [CrossRef]
- Benjakul, S.; Visessanguan, W.; Thongkaew, C.; Tanaka, M. Effect of frozen storage on chemical and gel-forming properties of fish commonly used for surimi production in Thailand. Food Hydrocoll. 2005, 19, 197–207. [Google Scholar] [CrossRef]
- Moosavi-Nasab, M.; Alli, I.; Ismail, A.A.; Ngadi, M.O. Protein structural changes during preparation and storage of surimi. J. Food Sci. 2005, 70, c448–c453. [Google Scholar] [CrossRef] [Green Version]
- Chanarat, S.; Benjakul, S.; Xiong, Y.L. Physicochemical changes of myosin and gelling properties of washed tilapia mince as influenced by oxidative stress and microbial transglutaminase. J. Food Sci. Technol. 2015, 52, 3824–3836. [Google Scholar] [CrossRef] [Green Version]
- Chaijan, M.; Panpipat, W.; Benjakul, S. Physicochemical and gelling properties of short-bodied mackerel (Rastrelliger brachysoma) protein isolate prepared using alkaline-aided process. Food Bioprod. Process. 2010, 88, 174–180. [Google Scholar] [CrossRef]
- Dawson, P.; Acton, J. Impact of proteins on food color. In Proteins in Food Processing, 2nd ed.; Rickey, Y.Y., Ed.; Woodhead Publishing: Duxford, UK, 2018; pp. 599–638. [Google Scholar]
- Acton, J.C.; Dawson, P.L. Color as a functional property of proteins. In Protein functionality in Food Systems; Navam, S.H., Gregory, R.Z., Eds.; Marcel Dekker: New York, NY, USA, 1994; pp. 357–381. [Google Scholar]
- Mazorra-Manzano, M.; Ramírez-Suárez, J.; Moreno-Hernández, J.; Pacheco-Aguilar, R. Seafood proteins. In Proteins in Food Processing; Rickey, Y.Y., Ed.; Woodhead Publishing: Duxford, UK, 2018; pp. 445–475. [Google Scholar]
- Richards, M.P.; Hultin, H.O. Contributions of blood and blood components to lipid oxidation in fish muscle. J. Agric. Food Chem. 2002, 50, 555–564. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Production | FPI-PR (%) | FPI-DMR (%) | Lipid Removal (%) | Ash Removal (%) |
|---|---|---|---|---|
| DM-FPI | 88.9 ± 5.3 a | 34.3 ± 0.2 a | 90.4 ± 1.0 b | 86.3 ± 0.8 b |
| HBB-FPI | 68.2 ± 4.8 b | 19.1 ± 1.4 b | 95.2 ± 0.7 a | 91.5 ± 0.6 a |
| ACO-FPI | 83.0 ± 2.9 a | 32.7 ± 2.7 a | 93.6 ± 0.2 a | 87.2 ± 0.5 b |
| Water Content | Crude Protein | Lipid Content | Ash Content | ||
|---|---|---|---|---|---|
| Raw material | Dark muscle | 66.5 ± 1.0 An | 14.7 ± 0.2 Bn | 17.6 ± 1.5 Bm | 0.8 ± 0.1 Bm |
| HBB | 54.8 ± 1.0 Cn | 15.2 ± 0.1 An | 21.9 ± 0.6 Am | 7.8 ± 0.4 Am | |
| ACO | 60.8 ± 2.0 Bn | 15.3 ± 0.1 An | 23.5 ± 1.1 Am | 1.0 ± 0.1 Bm | |
| FPIs and surimi | DM-FPI | 73.9 ± 0.7 bcm | 23.5 ± 0.9 am | 3.2 ± 0.0 bn | 0.1 ± 0.0 bn |
| HBB-FPI | 77.3 ± 0.8 am | 20.4 ± 0.5 bm | 2.8 ± 0.4 bn | 0.1 ± 0.0 bn | |
| ACO-FPI | 73.1 ± 2.1 cm | 24.4 ± 1.4 am | 3.1 ± 0.1 bn | 0.0 ± 0.0 bn | |
| Surimi | 77.0 ± 0.1 ab | 17.5 ± 0.3 c | 5.4 ± 0.1 a | 0.2 ± 0.0 a |
| Amino Acids | DM-FPI | HBB-FPI | ACO-FPI | FAO/WHO/UNU [70] * |
|---|---|---|---|---|
| Hydroxyproline | ND | ND | 0.6 | |
| Alanine b | 6.3 | 6.5 | 6.4 | |
| Arginine a | 6.6 | 6.9 | 6.3 | |
| Aspartic acid | 11.2 | 12.1 | 10.5 | |
| Cysteine/Cystine | 1.1 | 1.1 | 1.0 | |
| γ-Aminobutyric acid | ND | ND | ND | |
| Glutamic acid | 18.4 | 19.4 | 17.2 | |
| Glycine b | 3.7 | 3.9 | 5.1 | |
| Histidine a | 2.6 | 2.6 | 2.8 | 1.5 |
| Isoleucine ab | 4.7 | 5.0 | 4.6 | 3.0 |
| Leucine ab | 8.8 | 9.1 | 7.9 | 5.9 |
| Lysine a | 9.3 | 10.1 | 8.8 | 4.5 |
| Methionine ab | 3.3 | 3.5 | 2.9 | 2.2 |
| Phenylalanine ab | 3.8 | 4.2 | 3.8 | 3.8 |
| Proline b | 4.6 | 4.3 | 5.0 | |
| Serine | 4.2 | 4.4 | 4.1 | |
| Threonine a | 4.6 | 4.9 | 4.2 | 2.3 |
| Tyrosine | 2.2 | 3.5 | 3.1 | |
| Valine ab | 5.0 | 5.3 | 4.8 | 3.9 |
| Total amino acids | 100.5 | 106.8 | 99.1 | |
| Total essential amino acids | 48.8 | 51.7 | 46.2 | |
| Total Hydrophobic amino acid | 40.3 | 41.8 | 40.6 |
| Lightness (L*) | Redness (a*) | Yellowness (b*) | Whiteness | ||
|---|---|---|---|---|---|
| FPIs | DM-FPI | 47.8 ± 1.0 cA | 1.8 ± 0.1 bA | 11.4 ± 0.5 aA | 46.6 ± 1.1 cB |
| HBB-FPI | 34.0 ± 4.1 dC | 3.9 ± 1.6 aC | 5.4 ± 2.0 cB | 33.6 ± 4.0 dC | |
| ACO-FPI | 64.9 ± 0.3 bB | −1.2 ± 0.1 cB | 11.7 ± 0.2 aA | 63.0 ± 0.4 bA | |
| Industrial surimi | 73.5 ± 0.6 a | −0.2 ± 0.1 c | 4.2 ± 0.5 c | 73.2 ± 0.6 a | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nguyen, H.T.; Bao, H.N.D.; Dang, H.T.T.; Tómasson, T.; Arason, S.; Gudjónsdóttir, M. Protein Recovery of Tra Catfish (Pangasius hypophthalmus) Protein-Rich Side Streams by the pH-Shift Method. Foods 2022, 11, 1531. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11111531
Nguyen HT, Bao HND, Dang HTT, Tómasson T, Arason S, Gudjónsdóttir M. Protein Recovery of Tra Catfish (Pangasius hypophthalmus) Protein-Rich Side Streams by the pH-Shift Method. Foods. 2022; 11(11):1531. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11111531
Chicago/Turabian StyleNguyen, Hang Thi, Huynh Nguyen Duy Bao, Huong Thi Thu Dang, Tumi Tómasson, Sigurjón Arason, and María Gudjónsdóttir. 2022. "Protein Recovery of Tra Catfish (Pangasius hypophthalmus) Protein-Rich Side Streams by the pH-Shift Method" Foods 11, no. 11: 1531. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11111531