
Increasing the Versatility of Durum Wheat through Modifications of Protein and Starch Composition and Grain Hardness

 , , ,
, , , 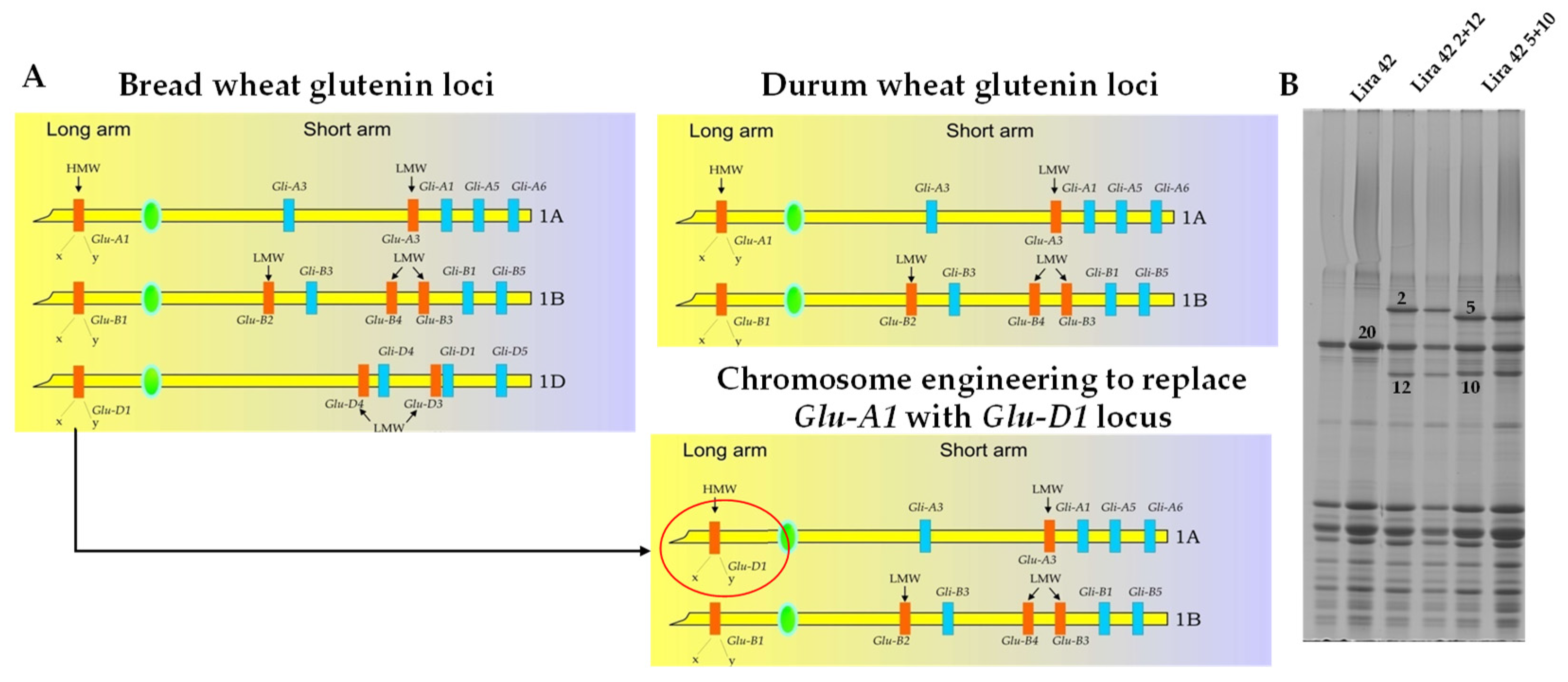{kind=link}
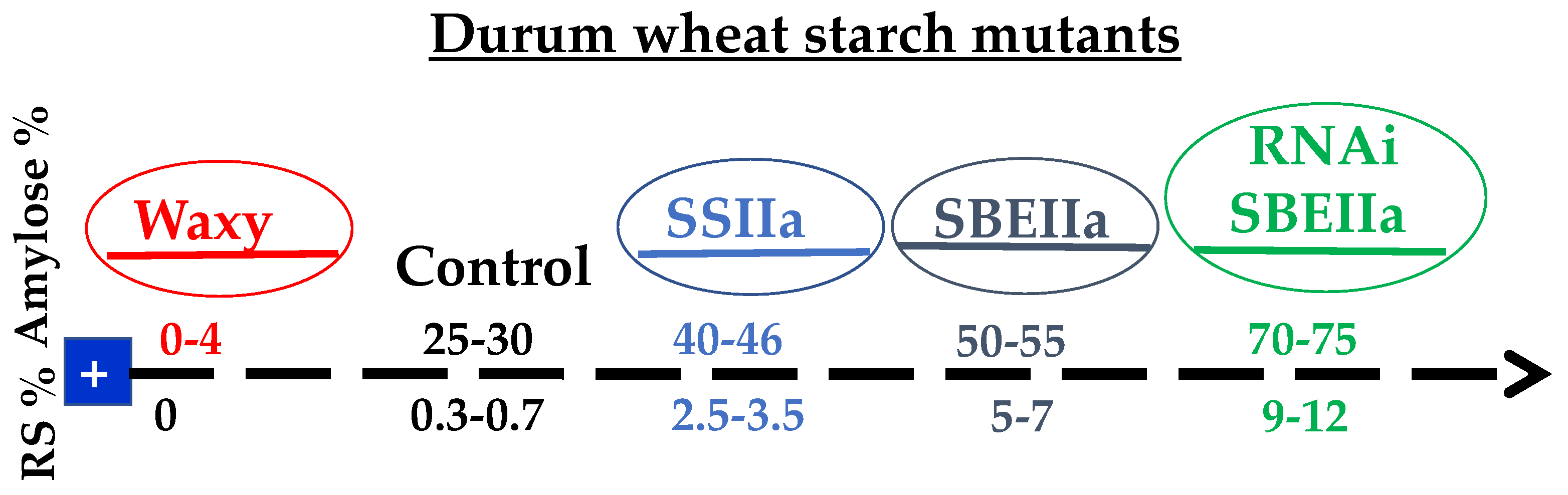{kind=link}
Abstract
:1. Introduction
2. Introduction of D Genome Gluten Proteins in Durum Wheat
3. Manipulation of Starch Composition
3.1. Low Amylose Durum Wheat
3.2. High Amylose Durum Wheat
4. Characterization of Soft Durum Wheat
5. Introduction of the HMW-GS 1Dx2 + 1Dy12, 1Dx5 + 1Dy10 and (Gpc-B1) Allele in Soft Durum
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Abecassis, J.; Cuq, B.; Boggini, G.; Namoune, H. Other traditional durum-derived products, Chapter 10. In Durum Wheat Chemistry and Technology, 2nd ed.; Sissons, M., Abecassis, J., Marchylo, B., Carcea, M., Eds.; AACC International Inc.: St. Paul, MN, USA, 2012; pp. 177–199. [Google Scholar]
- Pasqualone, A. Italian durum wheat breads. In Bread Consumption and Health; Silva Clerici, M.T.P., Ed.; Nova Biomedical: New York, NY, USA, 2012; pp. 57–80. [Google Scholar]
- Quaglia, G.B. Other durum wheat products. In Durum Wheat: Chemistry and Technology; Fabriani, G., Lintas, C., Eds.; American Association of Cereal Chemistry: St. Paul, MN, USA, 1988; pp. 263–282. [Google Scholar]
- Ammar, K.; Kronstad, W.E.; Morris, C.F. Breadmaking quality of selected durum wheat genotypes and its relationship with high molecular weight glutenin subunits allelic variation and gluten protein polymeric composition. Cereal Chem. 2000, 77, 230–236. [Google Scholar] [CrossRef]
- Boggini, G.; Tusa, P.; Pogna, N.E. Bread making quality of durum genotypes with some novel glutenin compositions. J. Cereal Sci. 1995, 22, 105–113. [Google Scholar] [CrossRef]
- Dexter, J.E.; Matsuo, R.R.; Preston, K.R.; Kilborn, R.H. Comparison of gluten strength, mixing properties, baking quality and spaghetti quality of some Canadian durum and common wheats. Can. Inst. Food Technol. J. 1981, 14, 108–111. [Google Scholar] [CrossRef]
- Guzman, C.; Autrique, J.E.; Mondal, S.; Singh, R.P.; Govindan, V.; Morales-Dorantes, A.; Pena, R.J. Response to drought and heat stress on wheat quality, with special emphasis on breadmaking quality, in durum wheat. Field Crops Res. 2016, 186, 157–165. [Google Scholar] [CrossRef]
- Hareland, G.A.; Puhr, D.P. Baking performance of durum and soft wheat flour in a sponge-dough breadmaking procedure. Cereal Chem. 1999, 75, 830–835. [Google Scholar] [CrossRef]
- Sissons, M.J. Role of durum wheat composition on the quality of pasta and bread. Food 2008, 2, 75–90. [Google Scholar]
- Shewry, P.R.; Halford, N.G. Cereal seed storage proteins: Structures, properties and role in grain utilization. J. Exp. Bot. 2002, 53, 947–958. [Google Scholar] [CrossRef] [Green Version]
- Kiszonas, A.M.; Morris, C.F. Wheat breeding for quality: A historical review. Cereal Chem. 2018, 95, 17–34. [Google Scholar] [CrossRef] [Green Version]
- Payne, P.L. Genetics of wheat storage proteins and the effect of allelic variation on bread making quality. Plant Physiol. 1987, 38, 141–153. [Google Scholar] [CrossRef]
- Ceoloni, C.; Forte, P.; Gennaro, A.; Micali, S.; Carozza, R.; Bitti, A. Recent developments in durum wheat chromosome engineering. Cytogenet. Genome Res. 2005, 109, 328–334. [Google Scholar] [CrossRef]
- Payne, P.I.; Jackson, E.A.; Holt, L.M. The association between -gliadin 45 and gluten strength in durum wheat varieties: A direct causal effect or the result of g netic linkage? J. Cereal Sci. 1984, 2, 73–81. [Google Scholar] [CrossRef]
- Pogna, N.E.; Autran, J.-C.; Mellini, F.; Lafiandra, D.; Feillet, P. Chromosome 1B encoded gliadins and glutenin subunits in durum wheat: Genetics and relationship to gluten strength. J. Cereal Sci. 1990, 11, 15–34. [Google Scholar] [CrossRef]
- Ruiz, M.; Giraldo, P. The influence of allelic variability of prolamins on gluten quality in durum wheat: An overview. J. Cereal Sci. 2021, 101, 103304. [Google Scholar] [CrossRef]
- Shevkani, K.; Singh, N.; Bajaj, R.; Kaur, A. Wheat starch production, structure, functionality and applications—A review. Int. J. Food Sci. Technol. 2017, 52, 38–58. [Google Scholar] [CrossRef]
- Hardy, K.; Brand-Miller, J.; Brown, K.D.; Thomas, M.G.; Copeland, L. The importance of dietary carbohydrate in human evolution. Q. Rev. Biol. 2015, 90, 251–268. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Xu, X. Modified starches and the stability of frozen foods. In Starch in Food; Woodhead Publishing: Sawston, UK, 2018; pp. 581–593. [Google Scholar]
- McCallum, C.M.; Comai, L.; Greene, E.A.; Henikoff, S. Targeting induced local lesions in genomes (TILLING) for plant functional genomics. Plant Physiol. 2000, 123, 439–442. [Google Scholar] [CrossRef] [Green Version]
- Slade, A.J.; McGuire, C.; Loeffler, D.; Mullenberg, J.; Skinner, W.; Fazio, G.; Holm, A.; Brandt, K.M.; Steine, M.N.; Goodstal, J.F.; et al. Development of high amylose wheat through TILLING. BMC Plant Biol. 2012, 12, 69. [Google Scholar] [CrossRef] [Green Version]
- Dong, C.; Vincent, K.; Sharp, P. Simultaneous mutation detection of three homoeologous genes in wheat by High Resolution Melting analysis and Mutation Surveyor. BMC Plant Biol. 2009, 9, 143. [Google Scholar] [CrossRef] [Green Version]
- Hazard, B.; Zhang, X.; Colasuonno, P.; Uauy, C.; Beckles, D.M.; Dubcovsky, J. Induced mutations in the Starch Branching Enzyme II (SBEII) genes increase amylose and resistant starch content in durum wheat. Crop Sci. 2012, 52, 1754–1766. [Google Scholar]
- Sestili, F.; Janni, M.; Doherty, A.; Botticella, E.; D’Ovidio, R.; Masci, S.; Jones, H.D.; Lafiandra, D. Increasing the amylose content of durum wheat through silencing of the SBEIIa genes. BMC Plant Biol. 2010, 10, 144. [Google Scholar] [CrossRef] [Green Version]
- Botticella, E.; Sestili, F.; Hernandez-Lopez, A.; Phillips, A.; Lafiandra, D. High resolution melting analysis for the detection of EMS induced mutations in wheat Sbella genes. BMC Plant Biol. 2011, 11, 156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabriani, G.; Lintas, C. Durum wheat. In Chemistry and Technology; American Association of Cereal Chemists: St. Paul, MN, USA, 1988. [Google Scholar]
- Posner, E.S.; Hibbs, A.N. Milling of soft and durum wheats. In Wheat Flour Milling; AACC International: St. Paul, MN, USA, 1997; pp. 257–283. [Google Scholar]
- Bhave, M.; Morris, C.F. Molecular genetics of puroindolines and related genes: Regulation of expression, membrane binding properties and applications. Plant Mol. Biol. 2008, 66, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Bhave, M.; Morris, C.F. Molecular genetics of puroindolines and related genes: Allelic diversity in wheat and other grasses. Plant Mol. Biol. 2008, 66, 205–219. [Google Scholar] [CrossRef] [PubMed]
- Morris, C.F. 2016 Cereals—Overview of uses: Accent on wheat grain. In Encyclopedia of Food Grains, 2nd ed.; Wrigley, C., Corke, H., Seetharaman, K., Faubion, J., Eds.; Academic Press: Oxford, UK, 2016; pp. 1–7. [Google Scholar]
- Gazza, L.; Sgrulletta, D.; Cammerata, A.; Gazzelloni, G.; Perenzin, M.; Pogna, N.E. Pastamaking and breadmaking quality of soft-textured durum wheat lines. J. Cereal Sci. 2011, 54, 481–487. [Google Scholar] [CrossRef]
- Morris, C.F.; Simeone, M.C.; King, G.E.; Lafiandra, D. Transfer of soft kernel texture from Triticum aestivum to durum wheat, Triticum turgidum ssp. durum. Crop Sci. 2011, 51, 114–122. [Google Scholar] [CrossRef]
- Barro, F.; Rooke, L.; Bekes, F.; Gras, P.; Tatham, A.S.; Fido, R.; Barcelo, P. Transformation of wheat with high molecular weight subunit genes results in improved functional properties. Nat. Biotechnol. 1997, 15, 1295–1299. [Google Scholar] [CrossRef]
- Blechl, A.; Lin, J.; Nguyen, S.; Chan, R.; Anderson, O.D.; Dupont, F.M. Transgenic wheats with elevated levels of Dx5 and/or Dy10 high-molecular-weight glutenin subunits yield doughs with increased mixing strength and tolerance. J. Cereal Sci. 2007, 45, 172–183. [Google Scholar] [CrossRef]
- Butow, B.J.; Tatham, A.S.; Savage, A.W.J.; Gilbert, S.M.; Shewry, F.M.; Solomon, R.G.; Bekes, F. Creating a balance—The incorporation of a HMW glutenin subunit into transgenic wheat lines. J. Cereal Sci. 2003, 38, 181–187. [Google Scholar] [CrossRef]
- Graybosch, R.A.; Seabourn, B.; Chen, Y.R.; Blechl, A.E. Quality and agronomic effects of three high-molecular-weight glutenin subunit transgenic events in winter wheat. Cereal Chem. 2011, 88, 95–102. [Google Scholar] [CrossRef]
- He, G.Y.; Rooke, L.; Steele, S.; Bekes, F.; Gras, P.; Tatham, A.S.; Lazzeri, P.A. Transformation of pasta wheat (Triticum turgidum L. var. durum) with high-molecular weight glutenin subunit genes and modification of dough functionality. Mol. Breed. 1999, 5, 377–386. [Google Scholar] [CrossRef]
- Gadaleta, A.; Blechl, A.E.; Nguyen, S.; Cardone, M.F.; Ventura, M.; Quick, J.S.; Blanco, A. Stably expressed D-genome derived HMW glutenin subunit genes transformed into different durum wheat genotypes change dough mixing properties. Mol. Breed. 2008, 22, 267–279. [Google Scholar] [CrossRef]
- Joppa, L.R.; Klindworth, D.L.; Hareland, G.A. Transfer of high molecular weight glutenins from spring wheat to durum wheat. In Proceedings of the 9th International Wheat Genetics Symposium, Saskatoon, SK, Canada, 2–8 August 1998; Slinkard, A.E., Ed.; University of Saskatchewan Extension Press: Saskatoon, SK, Canada, 1998; pp. 257–260. [Google Scholar]
- Klindworth, D.L.; Hareland, G.A.; Elias, E.M.; Ohm, J.-B.; Puhr, D.; Xu, S.S. Interactions of genotype and glutenin subunit composition on breadmaking quality of durum 1AS.1AL.-1DL translocation lines. Cereal Chem. 2014, 91, 211–217. [Google Scholar] [CrossRef]
- Lukaszewski, A.J. Registration of six germplasms of durum wheat with introgressions of the Glu-D1 locus. Crop Sci. 2003, 43, 1138–1139. [Google Scholar] [CrossRef]
- Sissons, M.; Pleming, D.; Margiotta, B.; D’Egidio, M.G.; Lafiandra, D. Effect of the introduction of D-genome related gluten proteins on durum wheat pasta and bread making quality. Crop Pasture Sci. 2014, 65, 27–37. [Google Scholar] [CrossRef]
- Vitellozzi, F.; Ciaffi, M.; Dominici, L.; Ceoloni, C. Isolation of a chromosomally engineered durum wheat line carrying the common wheat Glu-D1d allele. Agronomie 1997, 17, 413–419. [Google Scholar] [CrossRef] [Green Version]
- Morris, C.F. Bread-baking quality and the effects of Glu-D1 Gene introgressions in durum wheat (Triticum turgidum ssp. durum). Cereal Chem. 2021, 98, 1151–1158. [Google Scholar] [CrossRef]
- Klindworth, D.L.; Hareland, G.A.; Elias, E.M.; Xu, S.S. Agronomic and quality characteristics of 1as. 1al-1dl translocation lines of durum wheat carrying glutenin allele Glu-D1d. Crop Sci. 2005, 45, 77–84. [Google Scholar]
- Ammar, K.; Lukaszewsky, A.J.; Banowetz, G.M. Effect of Glu-D1 (5+10) on gluten strength and polymeric protein composition in durum wheat. Cereal Foods World 1997, 42, 610. [Google Scholar]
- Lafiandra, D.; Margiotta, B.; Colaprico, G.; Masci, S.I.; Roth, M.R.; MacRitchie, F. Introduction of the D-genome related high-and low-Mr glutenin subunits into durum wheat and their effect on technological properties. In Proceedings of the 7th International Gluten Workshop, Bristol, UK, 2–6 April 2000; Shewry, P.R., Tatham, A.S., Eds.; Royal Society of Chemistry: London, UK, 2000; pp. 51–54. [Google Scholar]
- Sissons, M.; Pleming, D.; Sestili, F.; Lafiandra, D. Effect of Glu-D1 introgression and amylose content on breadmaking potential of blends of durum and hexaploid wheat. Cereal Chem. 2019, 96, 193–206. [Google Scholar] [CrossRef]
- Sissons, M.; Sestili, F.; Lafiandra, D. Effect of Glu-D1 introgression on dough and pasta making quality of durum wheat lines with different glutenin composition and amylose content. Cereal Chem. 2019, 96, 207–219. [Google Scholar] [CrossRef]
- Rao, B.N. A Study of the Rheological Properties and Gluten Protein Components Associated with Enhanced Baking Quality in Durum Wheat (Triticum turgidum L. var. durum). Master’s Thesis, University of Saskatchewan, Saskatoon, SK, Canada, 2008. Available online: https://harvest.usask.ca/handle/10388/etd-09142008-143422 (accessed on 20 May 2022).
- Kiszonas, A.M.; Ibba, M.I.; Boehm, J.D., Jr.; Morris, C.F. Effects of Glu-D1 gene introgressions on soft white spring durum wheat (Triticum turgidum ssp. durum) quality. Cereal Chem. 2021, 98, 1112–1122. [Google Scholar] [CrossRef]
- Botticella, E.; Savatin, D.V.; Sestili, F. The triple jags of Dietary Fibers in cereals: How biotechnology is longing for High Fiber grains. Front. Plant Sci. 2021, 12, 745579. [Google Scholar] [CrossRef] [PubMed]
- Ball, S.G.; Morell, M.K. From bacterial glycogen to starch: Understanding the biogenesis of the plant starch granule. Ann. Rev. Plant Biol. 2003, 54, 207–233. [Google Scholar] [CrossRef] [PubMed]
- Sestili, F.; Sparla, F.; Botticella, E.; Janni, M.; D’Ovidio, R.; Giuseppe, F.; Marri, L.; Cuesta-Seijo, J.A.; Trost, P.; Lafiandra, D. The down-regulation of the genes encoding Isoamylase 1 alters the starch composition of the durum wheat grain. Plant Sci. 2016, 252, 230–238. [Google Scholar] [CrossRef] [PubMed]
- Guzmán, C.; Alvarez, J.B. Wheat waxy proteins: Polymorphism, molecular characterization and effects on starch properties. Theor. Appl. Genet. 2016, 129, 1–16. [Google Scholar] [CrossRef]
- Urbano, M.; Margiotta, B.; Colaprico, G.; Lafiandra, D. Waxy proteins in diploid, tetraploid and hexaploid wheats. Plant Breed. 2002, 121, 465–469. [Google Scholar] [CrossRef]
- Nakamura, T.; Kuroda, A. Variations in the content of starch-granule bound protein among several Japanese cultivars of common wheat (Triticum aestivum L.). Euphytica 1992, 64, 215–219. [Google Scholar]
- Lafiandra, D.; Sestili, F.; D’Ovidio, R.; Janni, M.; Botticella, E.; Ferrazzano, G.; DeAmbrogio, E. Approaches for modification of starch composition in durum wheat. Cereal Chem. 2010, 87, 28–34. [Google Scholar] [CrossRef]
- Grant, L.A.; Vignaux, N.; Doehlert, D.C.; McMullen, M.S.; Elias, E.M.; Kianian, S. Starch characteristics of waxy and nonwaxy tetraploid (Triticum turgidum L. var. durum) wheats. Cereal Chem. 2001, 78, 590–595. [Google Scholar] [CrossRef]
- Vignaux, N.; Doehlert, D.C.; Hegstad, J.; Elias, E.M.; McMullen, M.S.; Grant, L.A.; Kianian, S.F. Grain quality characteristics and milling performance of full and partial waxy durum lines. Cereal Chem. 2004, 81, 377–383. [Google Scholar] [CrossRef]
- Vignaux, N.; Doehlert, D.C.; Elias, E.M.; McMullen, M.S.; Grant, L.A.; Kianian, S.F. Quality of spaghetti made from full and partial waxy durum wheat. Cereal Chem. 2005, 82, 93–100. [Google Scholar] [CrossRef]
- Gianibelli, M.C.; Sissons, M.J.; Batey, I.L. Effect of source and proportion of waxy starches on pasta cooking quality. Cereal Chem. 2005, 82, 321–327. [Google Scholar] [CrossRef]
- Sharma, R.; Sissons, M.J.; Rathjen, A.J.; Jenner, C.F. The Null-4A allele at the waxy locus in durum wheat affects pasta cooking quality. J. Cereal Sci. 2002, 35, 287–297. [Google Scholar] [CrossRef]
- Šárka, E.; Dvořáček, V. New processing and applications of waxy starch (a review). J. Food Eng. 2017, 206, 77–87. [Google Scholar] [CrossRef]
- Bhattacharya, M.; Erazo-Castrejón, S.V.; Doehlert, D.C.; McMullen, M.S. Staling of bread as affected by waxy wheat flour blends. Cereal Chem. 2002, 79, 178–182. [Google Scholar] [CrossRef]
- Hayakawa, K.; Tanaka, K.; Nakamura, T.; Endo, S.; Hoshino, T. End use quality of waxy wheat flour in various grain-based foods. Cereal Chem. 2004, 81, 666–672. [Google Scholar] [CrossRef]
- Birt, D.F.; Boylston, T.; Hendrich, S.; Jane, J.L.; Hollis, J.; Li, L.; McClelland, J.; Moore, S.; Phillips, G.J.; Rowling, M.; et al. Resistant starch: Promise for improving human health. Adv. Nutr. 2013, 4, 587–601. [Google Scholar] [CrossRef] [Green Version]
- Sestili, F.; Botticella, E.; Lafiandra, D. TILLING for improved starch composition in wheat. In Genomics of Plant Genetic Resources; Tuberosa, R., Graner, A., Frison, E., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 467–487. [Google Scholar]
- Topping, D. Cereal complex carbohydrates and their contribution to human health. J. Cereal Sci. 2007, 46, 220–229. [Google Scholar] [CrossRef]
- Lockyer, S.; Nugent, A.P. Health effects of resistant starch. Nutr. Bull. 2017, 42, 10–41. [Google Scholar] [CrossRef]
- Carretta, M.D.; Quiroga, J.; López, R.; Hidalgo, M.A.; Burgos, R.A. Participation of short-chain fatty acids and their receptors in gut inflammation and colon cancer. Front. Physiol. 2021, 12, 662739. [Google Scholar] [CrossRef]
- Montroy, J.; Berjawi, R.; Lalu, M.M.; Podolsky, E.; Peixoto, C.; Sahin, L.; Stintzi, A.; Mack, D.; Fergusson, D.A. The effects of resistant starches on inflammatory bowel disease in preclinical and clinical settings: A systematic review and meta-analysis. BMC Gastroenterol. 2020, 20, 372. [Google Scholar] [CrossRef] [PubMed]
- Lafiandra, D.; Riccardi, G.; Shewry, P.R. Improving cereal grain carbohydrates for diet and health. J. Cereal Sci. 2014, 59, 312–326. [Google Scholar] [CrossRef] [Green Version]
- Lattimer, J.M.; Haub, M.D. Effects of dietary fiber and its components on metabolic health. Nutrients 2010, 2, 1266–1289. [Google Scholar] [CrossRef] [Green Version]
- Hogg, A.C.; Gause, K.; Hofer, P.; Martin, J.M.; Graybosch, R.A.; Hansen, L.E.; Giroux, M.J. Creation of a high-amylose durum wheat through mutagenesis of starch synthase II (SSIIa). J. Cereal Sci. 2013, 57, 377–383. [Google Scholar] [CrossRef] [Green Version]
- Sestili, F.; Palombieri, S.; Botticella, E.; Mantovani, P.; Bovina, R.; Lafiandra, D. TILLING mutants of durum wheat result in a high amylose phenotype and provide information on alternative splicing mechanisms. Plant Sci. 2015, 233, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Yamamori, M.; Endo, T.R. Variation of starch granule proteins and chromosome mapping of their coding genes in common wheat. Theor. Appl. Genet. 1996, 93, 275–281. [Google Scholar] [CrossRef]
- Botticella, E.; Sestili, F.; Ferrazzano, G.; Mantovani, P.; Cammerata, A.; D’Egidio, M.G.; Lafiandra, D. The impact of the SSIIa null mutations on grain traits and composition in durum wheat. Breed. Sci. 2016, 16025. [Google Scholar] [CrossRef] [Green Version]
- Martin, J.M.; Hogg, A.C.; Hofer, P.; Manthey, F.A.; Giroux, M.J. Impacts of SSIIa-A null allele on durum wheat noodle quality. Cereal Chem. 2014, 91, 176–182. [Google Scholar] [CrossRef]
- Hogg, A.C.; Martin, J.M.; Manthey, F.A.; Giroux, M.J. Nutritional and quality traits of pasta made from SSIIa null high-amylose durum wheat. Cereal Chem. 2015, 92, 395–400. [Google Scholar] [CrossRef]
- Nugent, A.P. Health properties of resistant starch. Nutr. Bull. 2005, 30, 27–54. [Google Scholar] [CrossRef]
- Regina, A.; Bird, A.; Topping, D.; Bowden, S.; Freeman, J.; Barsby, T.; Kosar-Hashemi, B.; Li, Z.; Rahman, S.; Morell, M. High-amylose wheat generated by RNA interference improves indices of large-bowel health in rats. Proc. Natl Acad. Sci. USA 2006, 103, 3546–3551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hazard, B.; Zhang, X.; Naemeh, M.; Hamilton, M.K.; Rust, B.; Raybould, H.E.; Newman, J.W.; Martin, R.; Dubcovsky, J. Mutations in durum wheat SBEII genes affect grain yield components, quality, and fermentation responses in rats. Crop Sci. 2015, 55, 2813–2825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sissons, M.; Sestili, F.; Botticella, E.; Masci, S.; Lafiandra, D. Can manipulation of durum wheat amylose content reduce the glycaemic index of spaghetti? Foods 2020, 9, 693. [Google Scholar] [CrossRef] [PubMed]
- Soh, H.N.; Sissons, M.J.; Turner, M.A. Effect of starch granule size distribution and elevated amylose content on durum dough rheology and spaghetti cooking quality. Cereal Chem. 2006, 83, 513–519. [Google Scholar] [CrossRef]
- Li, Y.; Wang, Q.; Li, X.Y.; Xiao, X.; Sun, F.S.; Wang, C.; Hu, W.; Feng, Z.; Chang, J.; Chen, M.; et al. Coexpression of the high molecular weight glutenin subunit 1Ax1 and puroindoline improves dough mixing properties in durum wheat (Triticum turgidum L. ssp. durum). PLoS ONE 2012, 7, e50057. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Mao, X.; Wang, Q.; Zhang, J.R.; Li, X.Y.; Ma, F.Y.; Sun, F.; Chang, J.; Chen, M.; Wang, Y.; et al. Overexpression of Puroindoline a gene in transgenic durum wheat (Triticum turgidum ssp. durum) leads to a medium-hard kernel texture. Mol. Breed. 2014, 33, 545–554. [Google Scholar] [CrossRef]
- Murray, J.C.; Kiszonas, A.M.; Wilson, J.; Morris, C.F. Effect of soft kernel texture on the milling properties of soft durum wheat. Cereal Chem. 2016, 93, 513–517. [Google Scholar] [CrossRef]
- Heinze, K.; Kiszonas, A.M.; Murray, J.C.; Morris, C.F.; Lullien-Pellerin, V. Puroindoline genes introduced into durum wheat reduce milling energy and change milling behavior similar to soft common wheats. J. Cereal Sci. 2016, 71, 183–189. [Google Scholar] [CrossRef] [Green Version]
- Murray, J.C.; Kiszonas, A.M.; Morris, C.F. Influence of soft kernel texture on the flour, water absorption, rheology, and baking quality of durum wheat. Cereal Chem. 2017, 94, 215–222. [Google Scholar] [CrossRef]
- Boehm, J.D., Jr.; Ibba, M.I.; Kiszonas, A.M.; Morris, C.F. End-use quality of CIMMYT-derived soft-kernel durum wheat germplasm: I. Grain, milling, and soft wheat quality. Crop Sci. 2017, 57, 1475–1484. [Google Scholar] [CrossRef]
- Kiszonas, A.M.; Ibba, M.I.; Boehm, J.D., Jr.; Morris, C.F. Effects of the functional Gpc-B1 allele on soft durum wheat grain, milling, flour, dough, and breadmaking quality. Cereal Chem. 2021, 98, 1250–1258. [Google Scholar] [CrossRef]
- Murray, J.C.; Kiszonas, A.M.; Morris, C.F. Influence of soft kernel texture on fresh durum pasta. J. Food Sci. 2018, 83, 2812–2818. [Google Scholar] [CrossRef] [PubMed]
- Gu, B.-J.; Kerr, C.J.; Morris, C.F.; Ganjyal, G.M. Soft durum wheat as a potential ingredient for direct expanded extruded products. J. Cereal Sci. 2021, 98, 103184. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lafiandra, D.; Sestili, F.; Sissons, M.; Kiszonas, A.; Morris, C.F. Increasing the Versatility of Durum Wheat through Modifications of Protein and Starch Composition and Grain Hardness. Foods 2022, 11, 1532. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11111532
Lafiandra D, Sestili F, Sissons M, Kiszonas A, Morris CF. Increasing the Versatility of Durum Wheat through Modifications of Protein and Starch Composition and Grain Hardness. Foods. 2022; 11(11):1532. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11111532
Chicago/Turabian StyleLafiandra, Domenico, Francesco Sestili, Mike Sissons, Alecia Kiszonas, and Craig F. Morris. 2022. "Increasing the Versatility of Durum Wheat through Modifications of Protein and Starch Composition and Grain Hardness" Foods 11, no. 11: 1532. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11111532