
Developing Qualitative Plasmid DNA Reference Materials to Detect Mechanisms of Quinolone and Fluoroquinolone Resistance in Foodborne Pathogens

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
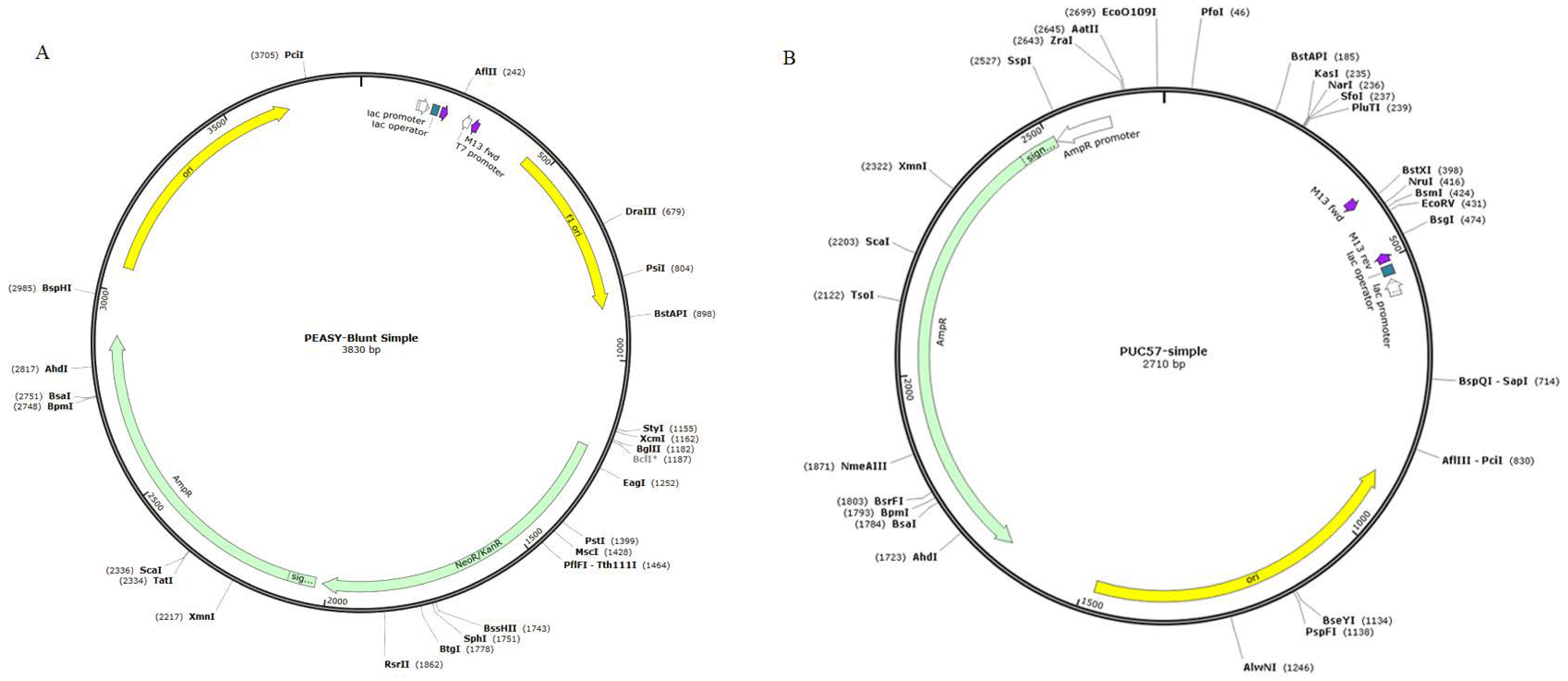
2.1. Target DNA Synthesis and Recombinant Plasmid and Strain Construction
2.2. Genetic Stability Test of the Target DNA in Recombinant Strains
2.3. Extraction of Plasmids Carrying Antibiotic Resistance-Encoding Genes
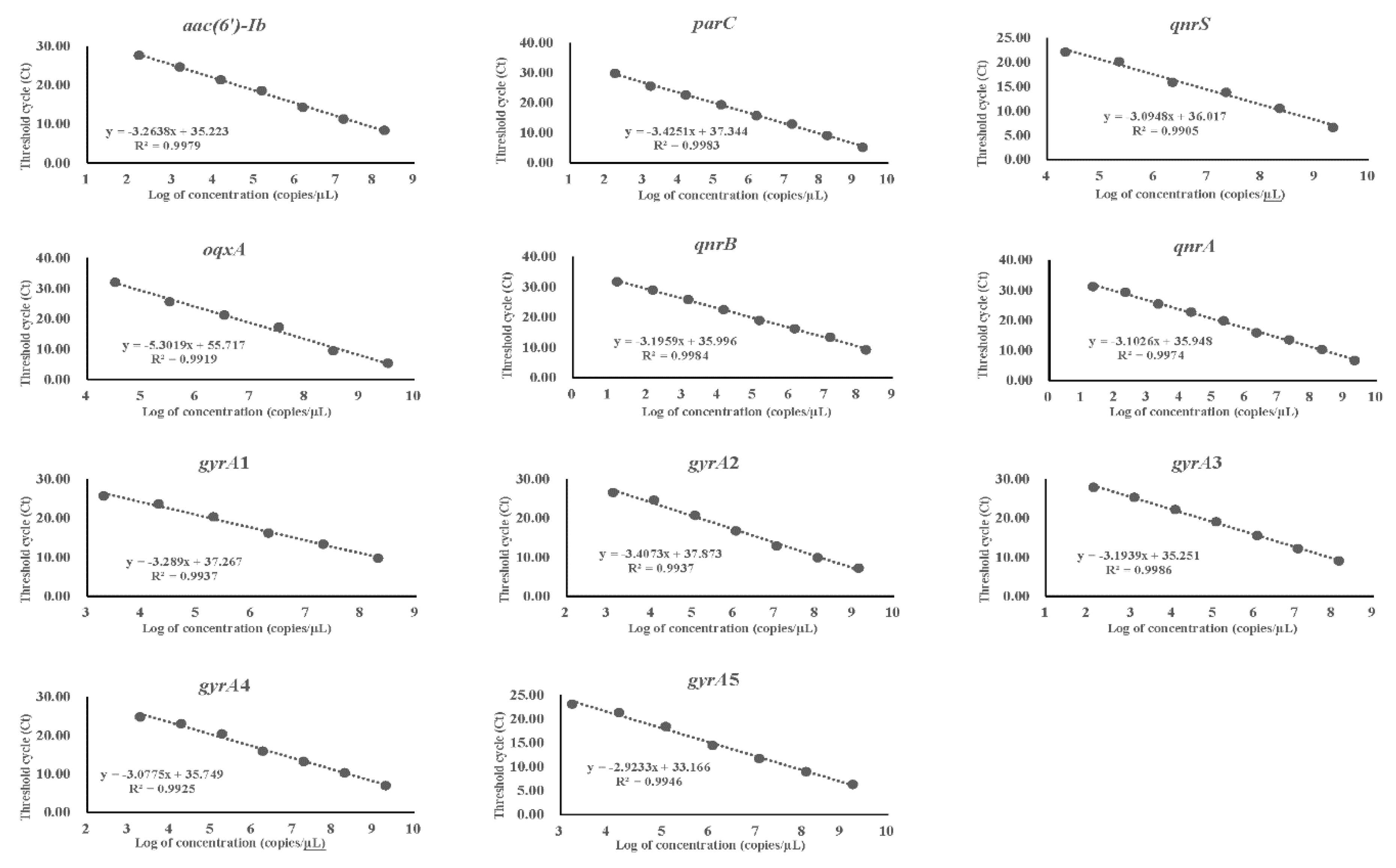
2.4. Assessment of Limit of Detection (LOD) for the Target DNA
2.5. Preparation of Plasmid DNA Reference Materials
2.6. Homogeneity Test of Plasmid DNA Reference Materials
2.7. Storage Stability Test of Plasmid DNA Reference Materials
2.8. Data Analysis
3. Results
3.1. Recombinant Plasmid and Strain Construction
3.2. Genetic Stability
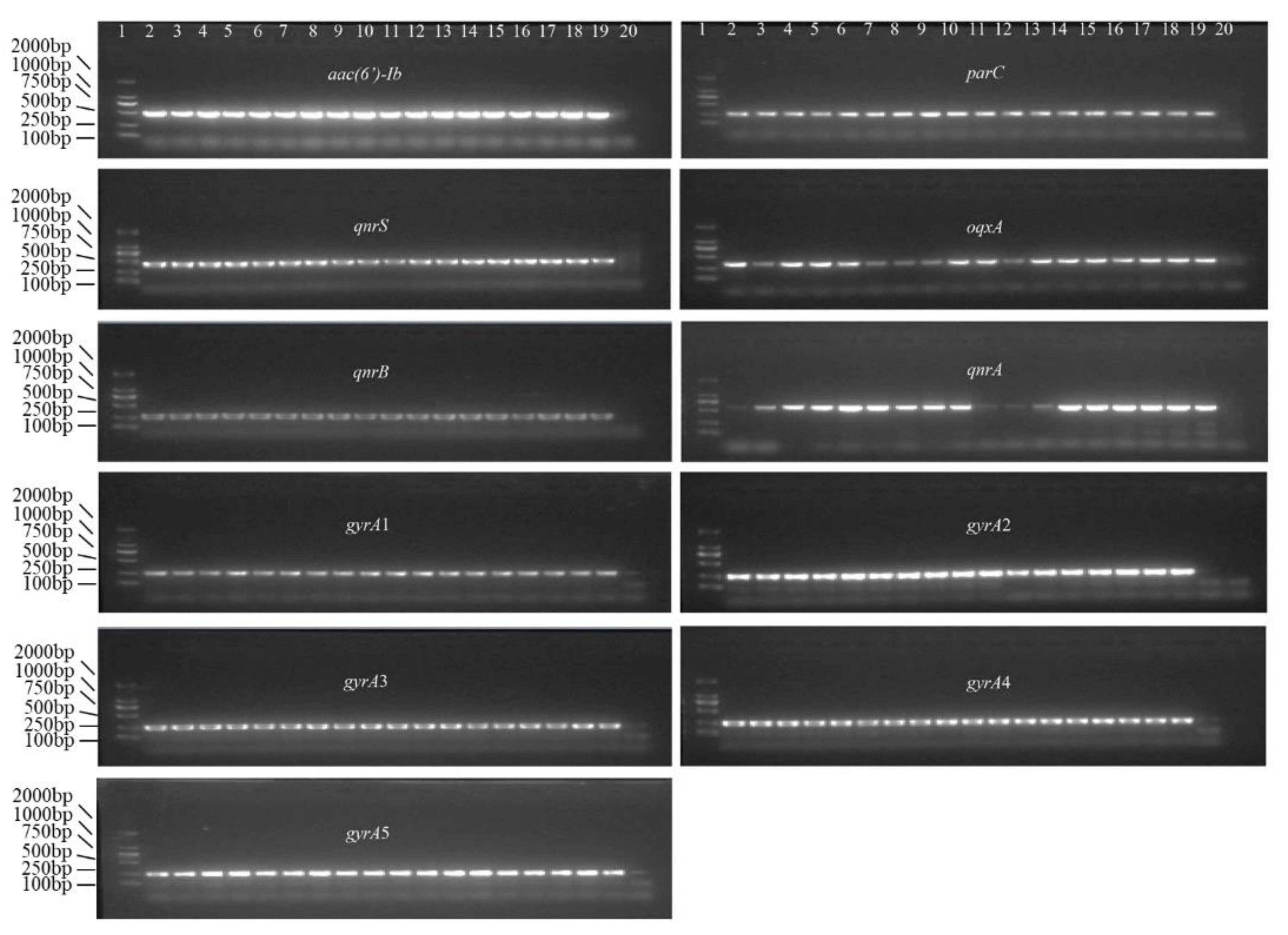
3.3. LOD of PCR and qPCR
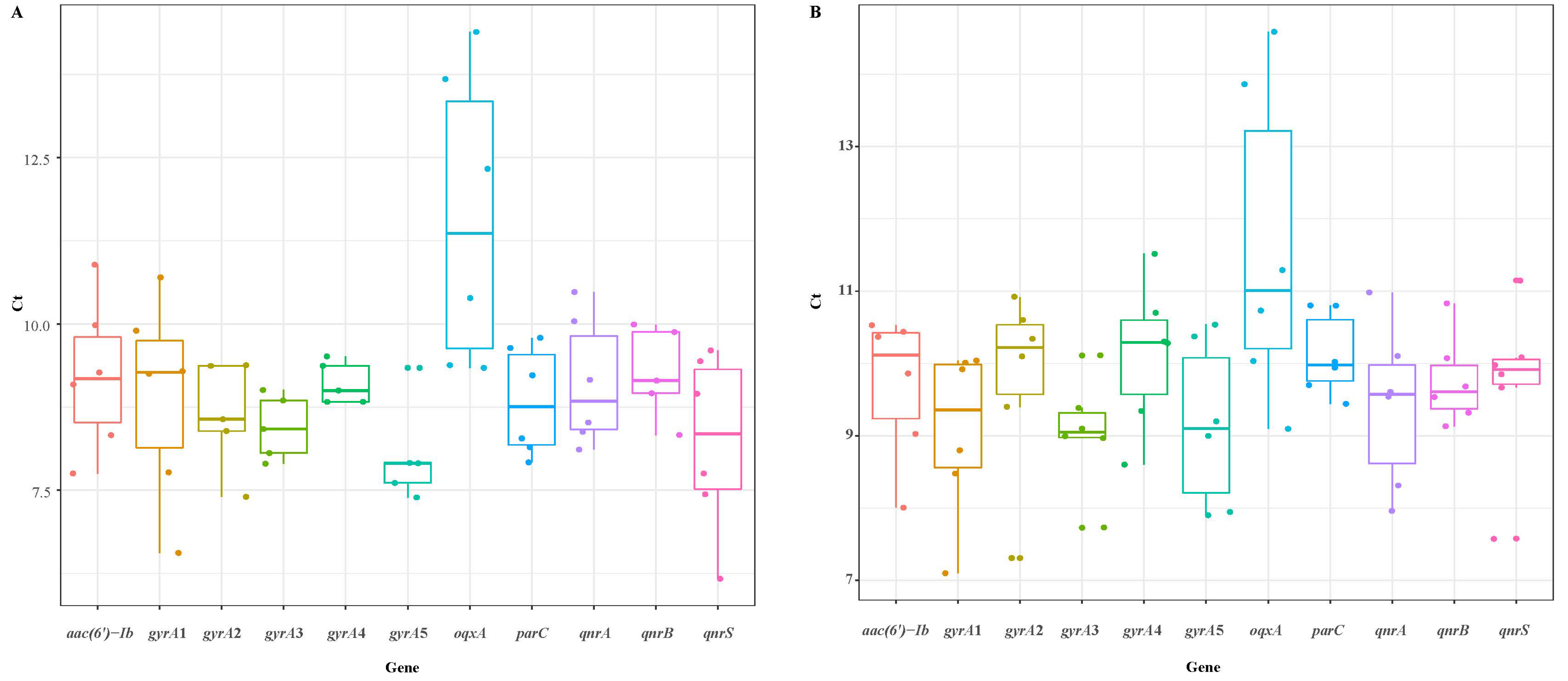
3.4. Homogeneity of Plasmid DNA Reference Materials
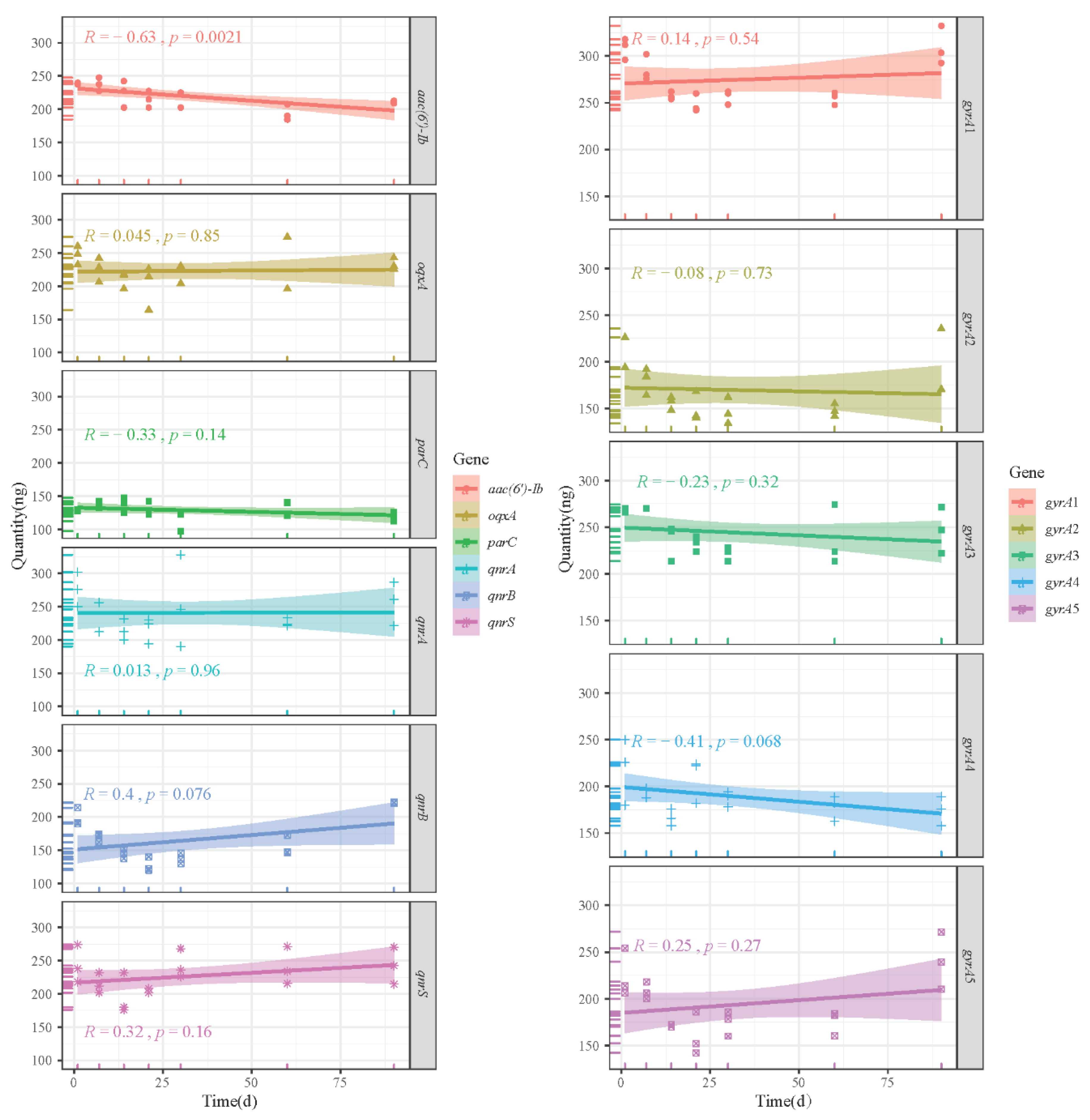
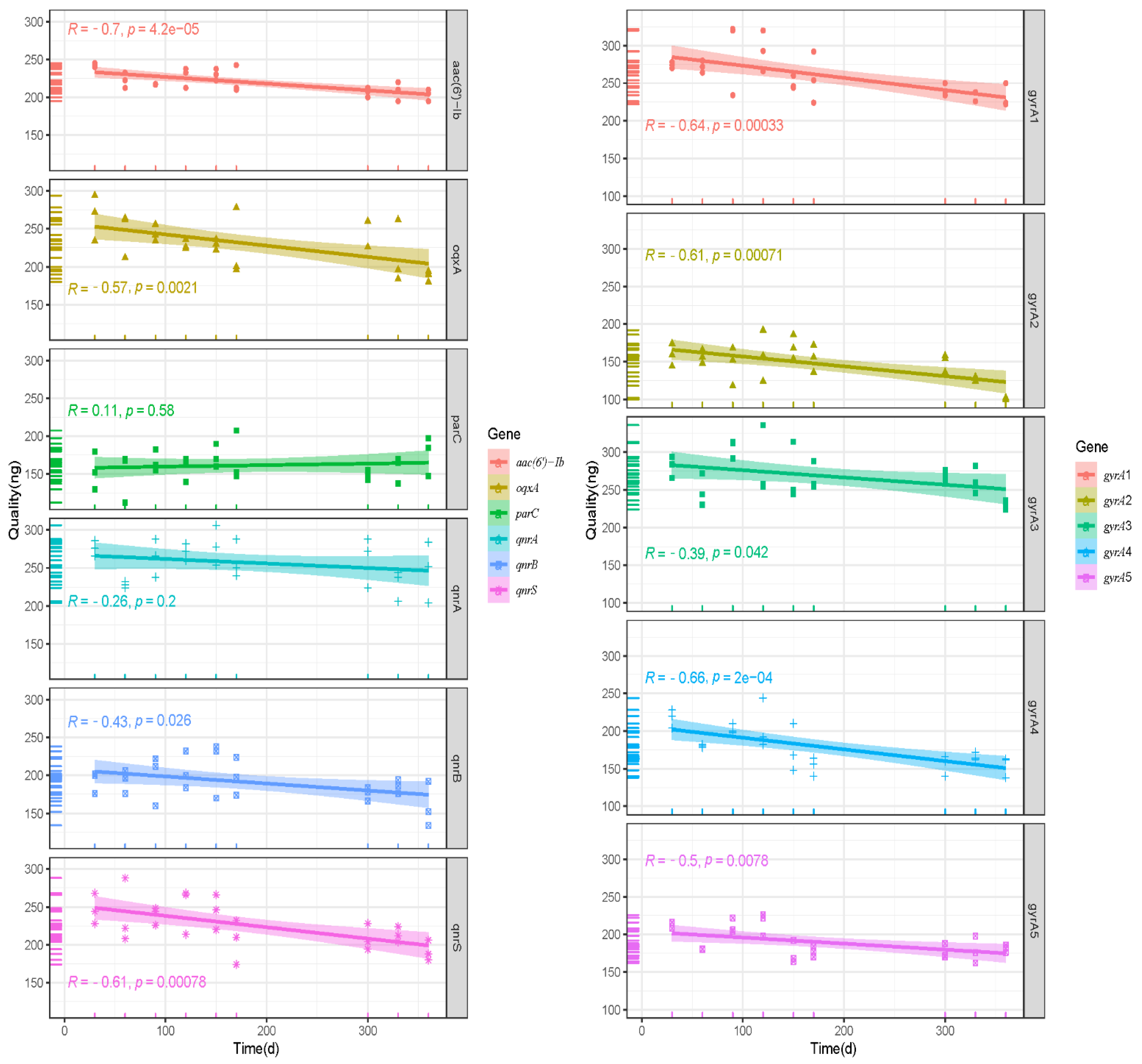
3.5. Storage Stability of Plasmid DNA Reference Materials
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Estimating the Burden of Foodborne Diseases. 2016. Available online: https://www.who.int/activities/estimating-the-burden-of-foodborne-diseases (accessed on 6 November 2020).
- Centers for Disease Control and Prevention [CDC]. Foodborne Diseases Active Surveillance Network FoodNet 2013 Surveillance Report. 2014. Available online: https://www.cdc.gov/foodnet/pdfs/508-compliant-2013-foodnet-annual-report.pdf (accessed on 6 November 2020).
- European Food Safety Authority (EFSA). The European Union Summary Report on Trends and Sources of Zoonoses, Zoonotic Agents and Food-Borne Outbreaks in 2016. 2017. Available online: https://0-efsa-onlinelibrary-wiley-com.brum.beds.ac.uk/doi/epdf/10.2903/j.efsa.2017.5077 (accessed on 6 November 2020).
- Crump, J.A.; Sjölund-Karlsson, M.; Gordon, M.; Parry, C.M. Epidemiology, clinical presentation, laboratory diagnosis, antimicrobial resistance, and antimicrobial management of invasive salmonella infections. Clin. Microbiol. Rev. 2015, 28, 901–937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horiyama, T.; Yamaguchi, A.; Nishino, K. TolC dependency of multidrug efflux systems in Salmonella enterica serovar Typhimurium. J. Antimicrob. Chemother. 2010, 65, 1372–1376. [Google Scholar] [CrossRef] [PubMed]
- Azargun, R.; Barhaghi, M.H.S.; Kafil, H.S.; Oskouee, M.A.; Sadeghi, V.; Memar, M.Y.; Ghotaslou, R. Frequency of DNA gyrase and topoisomerase IV mutations and plasmid-mediated quinolone resistance genes among Escherichia coli and Klebsiella pneumoniae isolated from urinary tract infections in Azerbaijan, Iran. J. Glob. Antimicrob. Resist. 2019, 17, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Yassine, I.; Rafei, R.; Osman, M.; Mallat, H.; Dabboussi, F.; Hamze, M. Plasmid-mediated quinolone resistance: Mechanisms, detection, and epidemiology in the Arab countries. Infect. Genet. Evol. 2019, 76, 104020. [Google Scholar] [CrossRef] [PubMed]
- Jing, R.; Wang, H.; Ju, S.; Cui, M. Reference materials for molecular diagnostics: Current achievements and future strategies. Clin. Biochem. 2018, 56, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Ke, L.; Dai, X.L.; Lin, J.; Huang, C.J.; Zeng, W.Y.; Zhang, W.L. Preparation of reference materials for Shigella. Food Sci. 2015, 36, 253–259. [Google Scholar] [CrossRef]
- Ke, L.; Huang, X.R.; Lin, J.; Dai, X.L.; Zeng, W.Y.; Zhang, W.L. Study on the reference materials of Salmonella Enteritidis nucleic acid. J. Food Sci. Biotechnol. 2015, 34, 891–895. [Google Scholar] [CrossRef]
- Xia, D.D.; Zhao, Y.Y.; Ma, P.P.; Wang, S.L.; Ma, C.Y.; Gao, S.F.; Kang, W.Y. Preparation of plasmid DNA reference material and its application in rapid detection of goji berries (Lycium barbarum) contaminated with Escherichia coli. Food Sci. 2020, 41, 267–274. [Google Scholar] [CrossRef]
- Tikhonova, E.B.; Yamada, Y.; Zgurskaya, H.I. Sequential mechanism of assembly of multidrug efflux pump AcrAB-TolC. Chem. Biol. 2011, 18, 454–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Sheng, H.; Xu, W.; Huang, J.; Meng, L.; Cao, C.; Zeng, J.; Meng, J.; Yang, B. Diversity of serotype, genotype, and antibiotic susceptibility of salmonella prevalent in pickled ready-to-eat meat. Front. Microbiol. 2019, 10, 2577. [Google Scholar] [CrossRef] [PubMed]
- Cattoir, V.; Poirel, L.; Rotimi, V.; Soussy, C.-J.; Nordmann, P. Multiplex PCR for detection of plasmid-mediated quinolone resistance qnr genes in ESBL-producing enterobacterial isolates. J. Antimicrob. Chemother. 2007, 60, 394–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Zhang, W.; Pan, W.; Yin, J.; Pan, Z.; Gao, S.; Jiao, X. Prevalence of qnr, aac(6′)-Ib-cr, qepA, and oqxAB in Escherichia coli isolates from humans, animals, and the environment. Antimicrob. Agents Chemother. 2012, 56, 3423–3427. [Google Scholar] [CrossRef] [Green Version]
- Park, C.H.; Robicsek, A.; Jacoby, G.A.; Sahm, D.; Hooper, D.C. Prevalence in the United States of aac(6′)-Ib-cr encoding a ciprofloxacin-modifying enzyme. Antimicrob. Agents Chemother. 2006, 50, 3953–3955. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.F.; Meng, X.F.; Yang, B.W.; Xia, X.D.; Wang, X.; Xi, M.L. Resistance of chickenborne Salmonella to quinolone and fluoroquinolones and related genes. J. Chin. Food Sci. Technol. 2015, 15, 158–165. [Google Scholar] [CrossRef]
- Eaves, D.J.; Randall, L.; Gray, D.T.; Buckley, A.; Woodward, M.J.; White, A.P.; Piddock, L.J.V. Prevalence of mutations within the quinolone resistance-determining region of gyrA, gyrB, parC, and parE and association with antibiotic resistance in quinolone-resistant salmonella enterica. Antimicrob. Agents Chemother. 2004, 48, 4012–4015. [Google Scholar] [CrossRef] [Green Version]
- Miao, X.; Jin, C.; Zhong, Y.; Feng, J.; Yan, C.; Xia, X.; Zhang, Y.; Peng, X. Data-independent acquisition-based quantitative proteomic analysis reveals the protective effect of apigenin on palmitate-induced lipotoxicity in human aortic endothelial cells. J. Agric. Food Chem. 2020, 68, 8836–8846. [Google Scholar] [CrossRef]
- Verdier, V.; Mosquera, G.; Assigbetse, K. Detection of the cassava bacterial blight pathogen, Xanthomonas axonopodis pv. manihotis, by Polymerase Chain Reaction. Plant Dis. 1998, 82, 79–83. [Google Scholar] [CrossRef] [Green Version]
- Verdier, V.; Mosquera, G. Specific detection of Xanthomonas axonopodis pv. manihotis with a DNA hybridization probe. J. Phytopathol. 1999, 147, 417–423. [Google Scholar] [CrossRef]
- Ogunjobi, A.A.; Fagade, O.E.; Dixon, A.G.O. Comparative analysis of genetic variation among Xanthomonas axonopodis pv. manihotis isolated from the western states of Nigeria using RAPD and AFLP. Indian J. Microbiol. 2010, 50, 132–138. [Google Scholar] [CrossRef] [Green Version]
- David, R.L.; Marta, H. Next-day Salmonella spp. detection method based on Real-Time PCR for foods. Methods Mol. Biol. 2021, 2182, 1–6. [Google Scholar] [CrossRef]
- Domesle, K.J.; Yang, Q.R.; Hammack, T.S.; Ge, B.L. Validation of a Salmonella loop-mediated isothermal amplification assay in animal food. Int. J. Food Microbiol. 2018, 264, 63–76. [Google Scholar] [CrossRef]
- Magnus, U.; Daniel, G.; Leonor, S.B.; Yonatan, G.; Susanne, J.; Makoto, O.; Monica, M.L.; Athena, L.; Aleksandra, E.S.; Teodora, W.; et al. The novel 2016 WHO Neisseria gonorrhoeae reference strains for global quality assurance of laboratory investigations: Phenotypic, genetic and reference genome characterization. J. Antimicrob. Chemother. 2016, 71, 3096–3108. [Google Scholar] [CrossRef] [Green Version]
- Luo, H.P.; Qu, H.R.; Shen, J.Y.; Ren, X.; Chen, Y.W.; Xie, G.D.; Yu, W.; Liu, N.; Ding, H.; Cui, S.H. Development of microbial reference materials for Cronobactersakazakii. J. Food Saf. Qual. 2019, 10, 54–59. [Google Scholar]
- Ke, L.; Lin, J.; Dai, X.L.; Huang, C.J.; Huang, X.R. The reference materials of preparation about Salmonella Enteritidis. Food Res. Dev. 2015, 36, 111–115. [Google Scholar] [CrossRef]
- Ma, P.P. Research and Development of Plasmid DNA Qualitative Reference Material of Listeria Monocytogenes and Its Application in the Detection of Tremella; Henan University: Zhengzhou, China, 2020. [Google Scholar]
- Debode, F.; Janssen, E.; Berben, G. Development of 10 new screening PCR assays for GMO detection targeting promoters (pFMV, pNOS, pSSuAra, pTA29, pUbi, pRice actin) and terminators (t35S, tE9, tOCS, tg7). Eur. Food Res. Technol. 2013, 236, 659–669. [Google Scholar] [CrossRef]
- Dorlass, E.G.; Monteiro, C.O.; Viana, A.O.; Soares, C.P.; Machado, R.R.G.; Thomazelli, L.M.; Araujo, D.B.; Leal, F.B.; Candido, E.D.; Telezynski, B.L.; et al. Lower cost alternatives for molecular diagnosis of COVID-19: Conventional RT-PCR and SYBR Green-based RT-qPCR. Braz. J. Microbiol. 2020, 51, 1117–1123. [Google Scholar] [CrossRef] [PubMed]
- Yoshii, Y.; Shimizu, K.; Morozumi, M.; Chiba, N.; Ubukata, K.; Uruga, H.; Hanada, S.; Wakui, H.; Ito, S.; Takasaka, N.; et al. Identification of pathogens by comprehensive real-time PCR versus conventional methods in community-acquired pneumonia in Japanese adults. Infect. Dis. 2016, 48, 782–788. [Google Scholar] [CrossRef] [PubMed]
- Fábio, C.A.B.; Andréia, Z.D.; Deise, J.K.; Josias, C.F.; Ana, C.M.A. Development of plasmid DNA reference material for the quantification of genetically modified common bean Embrapa 5.1. J. Agric. Food Chem. 2013, 61, 4921–4926. [Google Scholar] [CrossRef]
- Wu, Y.H.; Li, J.; Wang, Y.L.; Li, X.F.; Li, Y.J.; Zhu, L.; Li, J.; Wu, G. Development and application of a general plasmid reference material for GMO screening. Plasmid 2016, 87–88, 28–36. [Google Scholar] [CrossRef]
- Zhang, N.N.; Deng, Y.Q.; Nian, Q.G.; Kang, X.P.; Yang, Y.H.; Qin, C.F. Construction and characterization of recombinant pseudovirus particles carrying specific genomic RNA of St. Louis encephalitis virus. Mil. Med. Sci. 2017, 41, 194–198. [Google Scholar] [CrossRef]
- Junichi, M.; Shuko, H.; Satoshi, F.; Junji, Y.; Hiroki, N.; Shigehiro, N.; Reona, T.; Kazumi, K. Development of a reference material of a single DNA molecule for the quality control of PCR testing. Anal. Chem. 2014, 86, 8621–8627. [Google Scholar] [CrossRef]
- Vallejo, C.V.; Tere, C.P.; Calderon, M.N.; Arias, M.M.; Leguizamon, J.E. Development of a genomic DNA reference material for Salmonella enteritidis detection using polymerase chain reaction. Mol. Cell Probes 2021, 55, 101690. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.G.; Li, Y.P.; Luo, J.; Li, J.H.; Yu, J.P.; Wei, H.P. Preparation and characterization of pseudoviral positive control for nucleic acid detection of MERS-CoV. Chin. J. Front. Health Quar. 2019, 42, 233–238. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Sequence (5′–3′) | Annealing Temperature (°C) | Product Size (bp) | Reference |
|---|---|---|---|---|---|
| qnrA | qnrA-F qnrA-R | AGAGGATTTCTCACGCCAGG TGCCAGGCACAGATCTTGAC | 56 | 579 | [14] |
| qnrB | qnrB-F qnrB-R | GGCATTGAAATTCGCCACTG TTTGCTGCTCGCCAGTCGAA | 56 | 263 | [14] |
| qnrS | qnrS-F qnrS-R | GCAAGTTCATTGAACAGGGT TCTAAACCGTCGAGTTCGGCG | 56 | 427 | [14] |
| oqxA | oqxA-F oqxA-R | GACAGCGTCGCACAGAATG GGAGACGAGGTTGGTATGGA | 56 | 339 | [15] |
| aac(6′)-Ib | aac(6′)-Ib-F aac(6′)-Ib-R | TTGCGATGCTCTATGAGTGGCTA CTCGAATGCCTGGCGTGTTT | 55 | 482 | [16] |
| gyrA | gyrA-F gyrA-R | CCGTACCGTCATAGTTATCC CGTTGGTGACGTAATCGGTA | 56 | 251 | [17] |
| parC | parC-F parC-R | TAACAGCAGCTCGGCGTATT CTATGCGATGTCAGAGCTGG | 54 | 262 | [18] |
| Gene | Primer | Sequence (5′–3′) | Product Size (bp) |
|---|---|---|---|
| qnrA | qnrA-F qnrA-R | TGCTTTGGCATAGAGTTCAGG GGCATTGCTCCAGTTGTTTT | 192 |
| qnrB | qnrB-F qnrB-R | GGCATTGAAATTCGCCACTG TTTGCTGCTCGCCAGTCGAA | 263 |
| qnrS | qnrS-F qnrS-R | TCGTCGCTGCCACTTTGAT ATGCACCCGCTAGGTTCGTT | 296 |
| oqxA | oqxA-F oqxA-R | GACAGCGTCGCACAGAATG GGAGACGAGGTTGGTATGGA | 339 |
| aac(6′)-Ib | aac(6′)-Ib-F aac(6′)-Ib-R | CCGACACTTGCTGACGTACA GTTTCTTCTTCCCACCATCC | 155 |
| gyrA | gyrA-F gyrA-R | CCGTACCGTCATAGTTATCC CGTTGGTGACGTAATCGGTA | 251 |
| parC | parC-F parC-R | TAACAGCAGCTCGGCGTATT CTATGCGATGTCAGAGCTGG | 262 |
| Gene | Difference | SS | Df | MS | F-Value | p-Value | F-Critical Value |
|---|---|---|---|---|---|---|---|
| aac(6′)-Ib | interblock | 8.60 | 11.00 | 0.78 | 2.52 | 0.06 | 2.72 |
| intraclass | 3.73 | 12.00 | 0.31 | ||||
| qnrA | interblock | 4.63 | 11.00 | 0.42 | 2.05 | 0.12 | 2.72 |
| intraclass | 2.47 | 12.00 | 0.21 | ||||
| qnrB | interblock | 11.60 | 11.00 | 1.05 | 2.65 | 0.05 | 2.72 |
| intraclass | 4.77 | 12.00 | 0.40 | ||||
| qnrS | interblock | 11.87 | 11.00 | 1.08 | 2.64 | 0.05 | 2.72 |
| intraclass | 4.91 | 12.00 | 0.41 | ||||
| oqxA | interblock | 8.48 | 11.00 | 0.77 | 1.68 | 0.19 | 2.72 |
| intraclass | 5.50 | 12.00 | 0.46 | ||||
| parC | interblock | 1.66 | 11.00 | 0.15 | 0.86 | 0.60 | 2.72 |
| intraclass | 2.12 | 12.00 | 0.18 | ||||
| gyrA1 | interblock | 4.31 | 11.00 | 0.39 | 1.18 | 0.39 | 2.72 |
| intraclass | 3.98 | 12.00 | 0.33 | ||||
| gyrA2 | interblock | 4.08 | 11.00 | 0.37 | 2.64 | 0.05 | 2.72 |
| intraclass | 1.69 | 12.00 | 0.14 | ||||
| gyrA3 | interblock | 6.05 | 11.00 | 0.55 | 1.41 | 0.28 | 2.72 |
| intraclass | 4.69 | 12.00 | 0.39 | ||||
| gyrA4 | interblock | 5.59 | 11.00 | 0.51 | 2.23 | 0.09 | 2.72 |
| intraclass | 2.74 | 12.00 | 0.23 | ||||
| gyrA5 | interblock | 3.29 | 11.00 | 0.30 | 2.47 | 0.07 | 2.72 |
| intraclass | 1.45 | 12.00 | 0.12 |
| Gene | 1 Day | 7 Days | 13 Days |
|---|---|---|---|
| Ct Value | Ct Value | Ct Value | |
| aac(6’)-Ib | 7.33 ± 0.42 | 8.31 ± 0.57 | 7.79 ± 0.54 |
| parC | 7.66 ± 0.58 | 7.80 ± 0.82 | 9.24 ± 0.90 |
| qnrS | 7.26 ± 0.69 | 6.76 ± 0.25 | 7.36 ± 0.38 |
| oqxA | 7.89 ± 0.87 | 8.39 ± 1.22 | 7.98 ± 1.55 |
| qnrB | 8.62 ± 0.15 | 7.53 ± 0.28 | 7.75 ±0.61 |
| qnrA | 8.00 ± 0.63 | 7.94 ± 0.40 | 7.85 ± 0.86 |
| gyrA1 | 9.12 ± 1.14 | 7.13 ± 0.32 | 8.33 ± 0.23 |
| gyrA2 | 7.55 ± 0.44 | 8.78 ± 0.80 | 8.91 ± 1.33 |
| gyrA3 | 8.44 ± 0.51 | 7.33 ± 0.38 | 6.39 ± 0.47 |
| gyrA4 | 8.55 ± 0.78 | 8.16 ± 0.41 | 8.22 ± 0.85 |
| gyrA5 | 8.42 ± 0.75 | 7.87 ± 0.77 | 7.77 ± 0.63 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niu, Q.; Su, X.; Lian, L.; Huang, J.; Xue, S.; Zhou, W.; Zhao, H.; Lu, X.; Cui, S.; Chen, J.; et al. Developing Qualitative Plasmid DNA Reference Materials to Detect Mechanisms of Quinolone and Fluoroquinolone Resistance in Foodborne Pathogens. Foods 2022, 11, 154. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11020154
Niu Q, Su X, Lian L, Huang J, Xue S, Zhou W, Zhao H, Lu X, Cui S, Chen J, et al. Developing Qualitative Plasmid DNA Reference Materials to Detect Mechanisms of Quinolone and Fluoroquinolone Resistance in Foodborne Pathogens. Foods. 2022; 11(2):154. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11020154
Chicago/Turabian StyleNiu, Qinya, Xiumin Su, Luxin Lian, Jinling Huang, Shutong Xue, Wei Zhou, Hongyang Zhao, Xing’an Lu, Shenghui Cui, Jia Chen, and et al. 2022. "Developing Qualitative Plasmid DNA Reference Materials to Detect Mechanisms of Quinolone and Fluoroquinolone Resistance in Foodborne Pathogens" Foods 11, no. 2: 154. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11020154