Predicted Release and Analysis of Novel ACE-I, Renin, and DPP-IV Inhibitory Peptides from Common Oat (Avena sativa) Protein Hydrolysates Using in Silico Analysis

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Oat Protein Extraction
2.3. In Silico Digestion
2.4. Bioactivity Prediction In Silico
2.5. Chemical Synthesis of Peptides
2.6. Renin Inhibition Assay
2.7. ACE-I Inhibition Assay
2.8. DPP-IV Inhibition Assay
3. Results
3.1. In Silico Bioactivity Prediction
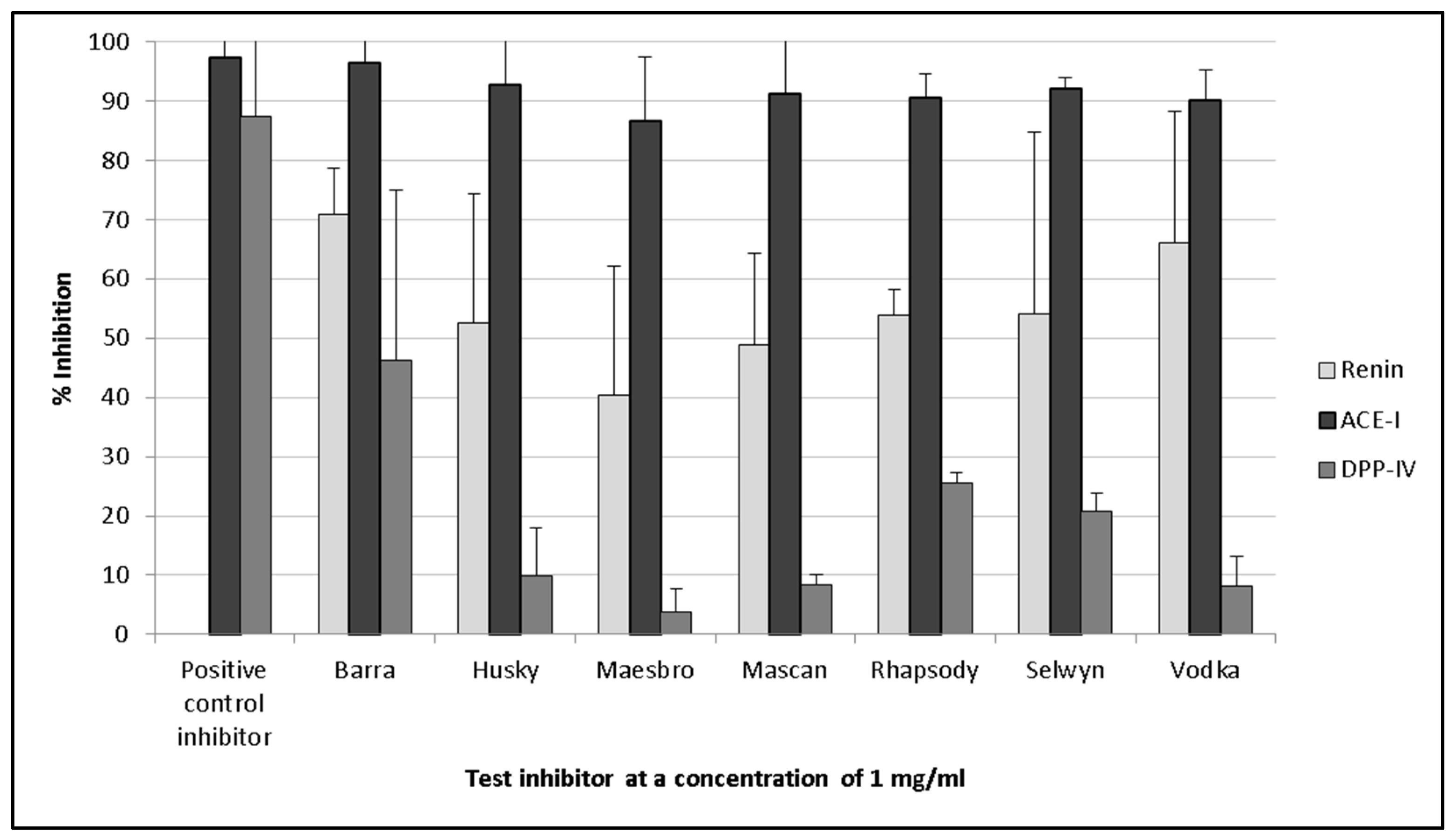
3.2. Renin Inhibition
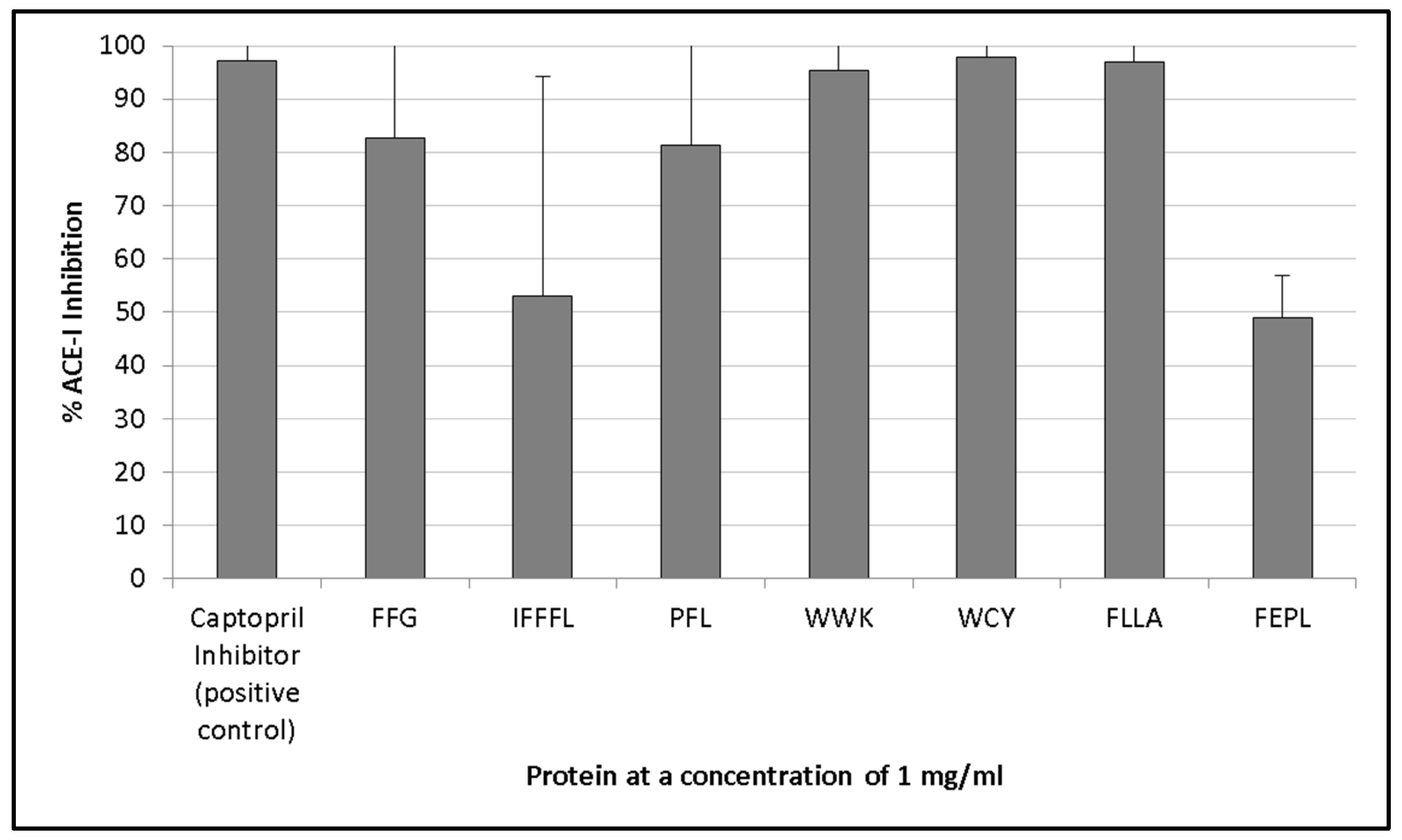
3.3. ACE-I Inhibition
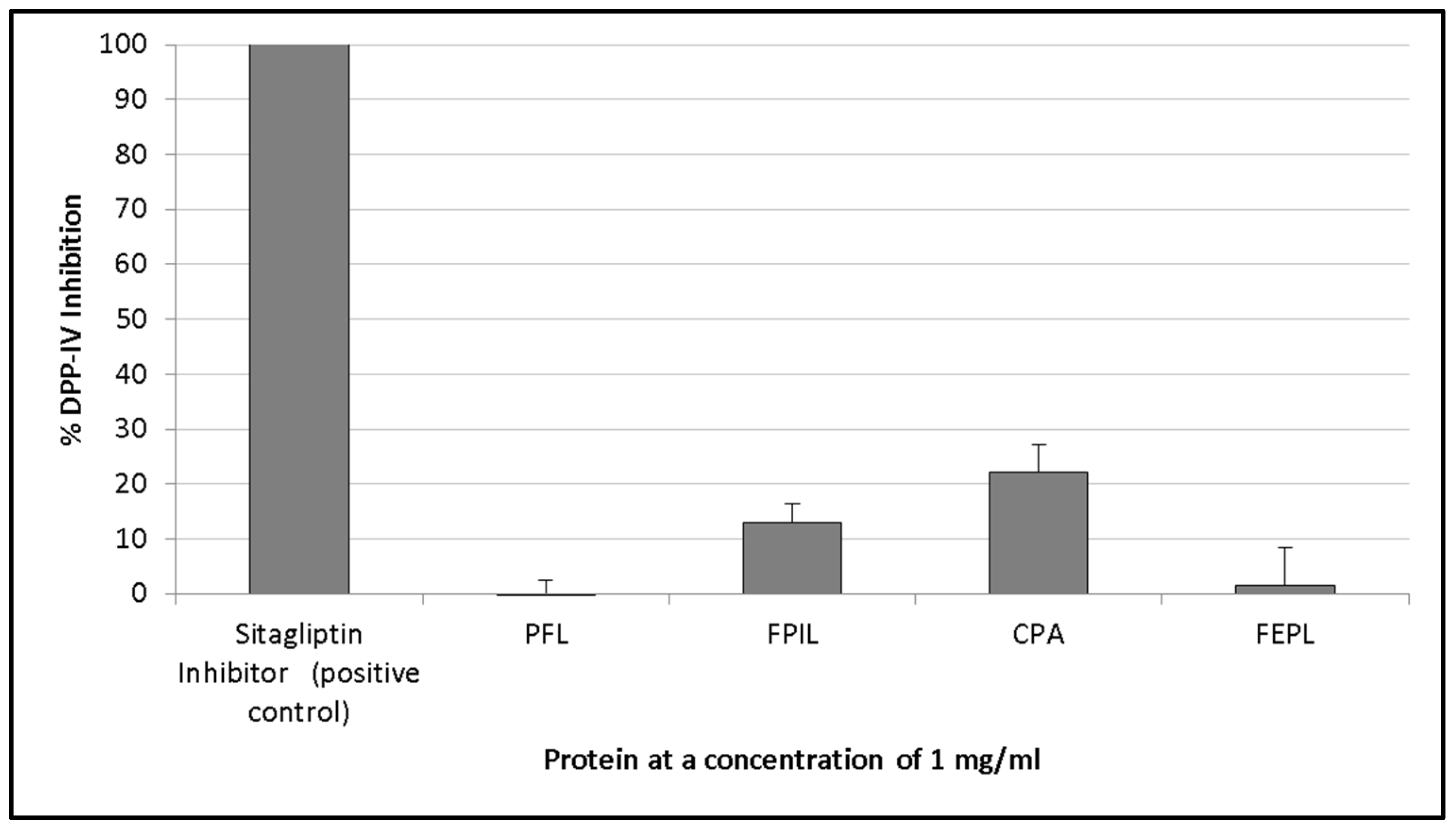
3.4. DPP-IV Inhibition
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide | Predicted Activity | Peptide Ranker Score | Protein | Enzyme | Location | Mass (Da) |
|---|---|---|---|---|---|---|
| FFG * | ACE-I | 1 | Avenin | Papain, Ficin | f(109–111) | 369.45 |
| IFFFL * | ACE-I | 0.99 | Avenin | Ficin | f(3–7) | 685.93 |
| WCY * | ACE-I | 0.98 | 11S globulin/12S globulin | Papain, Ficin/Papain | f(172–174)/f(172–174) | 470.57 |
| PFL * | ACE-I | 0.98 | Avenin-3/Avenin-E | Ficin/Ficin | f(84–86)/f(70–72) | 375.5 |
| WWK * | ACE-I | 0.98 | Avenin/Tryptophanins | Papain, Ficin/Papain, Ficin | f(70–72)/f(65–67) | 518.65 |
| IFFFLA | ACE-I | 0.95 | Avenin | Papain | f(3–8) | 757.02 |
| FPIL * | DPP-IV, ACE-I | 0.92 | 11S globulin | Ficin | f(364–367) | 488.68 |
| IWR | ACE-I | 0.91 | Avenin/Tryptophanins | Papain/Papain | f(98–100)/f(94–96) | 473.61 |
| FPTL | DPP-IV, | 0.84 | 12S globulin | Ficin | f(356–359) | 476.62 |
| PFV | DPP | 0.82 | 12S globulin | Ficin | f(264–266) | 361.47 |
| CPA * | DPP-IV | 0.79 | 11S globulin/12S globulin/Avenin-3/Avenin-E | Papain, Ficin/Papain, Ficin/Papain/Papain | f(121–123)/f(121–123)/f(116–118)/f(102–104) | 289.37 |
| FLLA * | DPP | 0.79 | 11S globulin/12S globulin | Papain/Papain | f(203–206)/f(203–206) | 462.62 |
| FEPL * | ACE-I | 0.75 | 12S globulin | Papain | f(53–56) | 504.63 |
| PLR | ACE-I | 0.74 | 11S globulin/12S globulin | Papain/Papain | f(55–57) | 384.51 |
References
- WHO. Global Health Risks: Mortality and Burden of Disease Attributable to Selected Major Risks; World Health Organization: Geneva, Switzerland, 2009. [Google Scholar]
- Heidenreich, P.A.; Trogdon, J.G.; Khavjou, O.A.; Butler, J.; Dracup, K.; Ezekowitz, M.D.; Finkelstein, E.A.; Hong, Y.; Johnston, S.C.; Khera, A.; et al. Forecasting the future of cardiovascular disease in the united states: A policy statement from the American heart association. Circulation 2011, 123, 933–944. [Google Scholar] [CrossRef] [PubMed]
- Ondetti, M.A.; Cushman, D.W. Enzymes of the renin-angiotensin system and their inhibitors. Annu. Rev. Biochem. 1982, 51, 283–308. [Google Scholar] [CrossRef] [PubMed]
- Turk, B. Targeting proteases: Successes, failures and future prospects. Nat. Rev. Drug Discov. 2006, 5, 785–799. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, C.; Gallagher, E.; Tasdemir, D.; Hayes, M. Heart health peptides from macroalgae and their potential use in functional foods. J. Agric. Food Chem. 2011, 59, 6829–6836. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.-Y.; Peng, C.; Jiao, R.; Wong, Y.M.; Yang, N.; Huang, Y. Anti-hypertensive nutraceuticals and functional foods. J. Agric. Food Chem. 2009, 57, 4485–4499. [Google Scholar] [CrossRef] [PubMed]
- Korhonen, H.; Pihlanto, A. Bioactive peptides: Production and functionality. Int. Dairy J. 2006, 16, 945–960. [Google Scholar] [CrossRef]
- Clare, D.A.; Swaisgood, H.E. Bioactive milk peptides: A prospectus. J. Dairy Sci. 2000, 83, 1187–1195. [Google Scholar] [CrossRef]
- Nakamura, Y.; Yamamoto, N.; Sakai, K.; Okubo, A.; Yamazaki, S.; Takano, T. Purification and characterization of angiotensin I-converting enzyme inhibitors from sour milk. J. Dairy Sci. 1995, 78, 777–783. [Google Scholar] [CrossRef]
- Hatanaka, T.; Inoue, Y.; Arima, J.; Kumagai, Y.; Usuki, H.; Kawakami, K.; Kimura, M.; Mukaihara, T. Production of dipeptidyl peptidase iv inhibitory peptides from defatted rice bran. Food Chem. 2012, 134, 797–802. [Google Scholar] [CrossRef] [PubMed]
- Wilson, J.; Hayes, M.; Carney, B. Angiotensin-I-converting enzyme and prolyl endopeptidase inhibitory peptides from natural sources with a focus on marine processing by-products. Food Chem. 2011, 129, 235–244. [Google Scholar] [CrossRef]
- Pellegrini, A. Antimicrobial peptides from food proteins. Curr. Pharm. Des. 2003, 9, 1225–1238. [Google Scholar] [CrossRef] [PubMed]
- Chabance, B.; Marteau, P.; Rambaud, J.C.; Migliore-Samour, D.; Boynard, M.; Perrotin, P.; Guillet, R.; Jollès, P.; Fiat, A.M. Casein peptide release and passage to the blood in humans during digestion of milk or yogurt. Biochimie 1998, 80, 155–165. [Google Scholar] [CrossRef]
- Suetsuna, K.; Ukeda, H.; Ochi, H. Isolation and characterization of free radical scavenging activities peptides derived from casein. J. Nutr. Biochem. 2000, 11, 128–131. [Google Scholar] [CrossRef]
- Nagaoka, S.; Futamura, Y.; Miwa, K.; Awano, T.; Yamauchi, K.; Kanamaru, Y.; Tadashi, K.; Kuwata, T. Identification of novel hypocholesterolemic peptides derived from bovine milk beta-lactoglobulin. Biochem. Biophys. Res. Commun. 2001, 281, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, M. Food-derived peptides and intestinal functions. Biofactors 2004, 21, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Yvon, M.; Beucher, S.; Guilloteau, P.; Le Huerou-Luron, I.; Corring, T. Effects of caseinomacropeptide (cmp) on digestion regulation. Reprod. Nutr. Dev. 1994, 34, 527–537. [Google Scholar] [CrossRef] [PubMed]
- Teschemacher, H. Opioid receptor ligands derived from food proteins. Curr. Pharm. Des. 2003, 9, 1331–1344. [Google Scholar] [CrossRef] [PubMed]
- Gill, H.S.; Doull, F.; Rutherfurd, K.J.; Cross, M.L. Immunoregulatory peptides in bovine milk. Br. J. Nutr. 2000, 84 (Suppl. S1), S111–S117. [Google Scholar] [CrossRef]
- Meisel, H.; FitzGerald, R.J. Biofunctional peptides from milk proteins: Mineral binding and cytomodulatory effects. Curr. Pharm. Des. 2003, 9, 1289–1295. [Google Scholar] [PubMed]
- Saito, T. Antihypertensive peptides derived from bovine casein and whey proteins. In Bioactive Components of Milk; Bösze, Z., Ed.; Springer: New York, NY, USA, 2008; pp. 295–317. [Google Scholar]
- Li, G.H.; Qu, M.R.; Wan, J.Z.; You, J.M. Antihypertensive effect of rice protein hydrolysate with in vitro Angiotensin-I-converting enzyme inhibitory activity in spontaneously hypertensive rats. Asia Pac. J. Clin. Nutr. 2007, 16 (Suppl. S1), 275–280. [Google Scholar]
- Rho, S.J.; Lee, J.S.; Chung, Y.I.; Kim, Y.W.; Lee, H.G. Purification and identification of an Angiotensin I-converting enzyme inhibitory peptide from fermented soybean extract. Process Biochem. 2009, 44, 490. [Google Scholar] [CrossRef]
- Motoi, H.; Kodama, T. Isolation and characterization of angiotensin I-converting enzyme inhibitory peptides from wheat gliadin hydrolysate. Nahrung 2003, 47, 354–358. [Google Scholar] [CrossRef] [PubMed]
- Suetsuna, K.; Nakano, T. Identification of an antihypertensive peptide from peptic digest of wakame (Undaria pinnatifida). J. Nutr. Biochem. 2000, 11, 450–454. [Google Scholar] [CrossRef]
- Aluko, R.E. Determination of nutritional and bioactive properties of peptides in enzymatic pea, chickpea, and mung bean protein hydrolysates. J. AOAC Int. 2008, 91, 947–956. [Google Scholar] [PubMed]
- Lee, J.-E.; Bae, I.Y.; Lee, H.G.; Yang, C.-B. Tyr-Pro-Lys, an Angiotensin-I-converting enzyme inhibitory peptide derived from Broccoli (Brassica oleracea italica). Food Chem. 2006, 99, 143–148. [Google Scholar] [CrossRef]
- Suetsuna, K. Isolation and characterization of Angiotensin-I-converting enzyme inhibitor dipeptides derived from Allium sativum L (garlic) isolation and characterization of angiotensin I-converting enzyme inhibitor dipeptides derived from. J. Nutr. Biochem. 1998, 9, 415. [Google Scholar] [CrossRef]
- Miguel, M.; Aleixandre, A. Antihypertensive peptides derived from egg proteins. J. Nutr. 2006, 136, 1457–1460. [Google Scholar] [PubMed]
- Vercruysse, L.; Van Camp, J.; Smagghe, G. Ace inhibitory peptides derived from enzymatic hydrolysates of animal muscle protein: A review. J. Agric. Food Chem. 2005, 53, 8106–8115. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Hu, J.; Miyaguchi, Y.; Bai, X.; Du, Y.; Lin, B. Isolation and characterization of angiotensin i-converting enzyme inhibitory peptides derived from porcine hemoglobin. Peptides 2006, 27, 2950–2956. [Google Scholar] [CrossRef] [PubMed]
- Jung, W.K.; Mendis, E.; Je, J.Y.; Park, P.J.; Son, B.W.; Kim, H.C. Angiotensin-I-converting enzyme inhibitory peptide from yellowfin sole (Limanda aspera) frame protein and its antihypertensive effect in spontaneously hypertensive rats Angiotensin I-converting enzyme inhibitory peptide from yellowfin sole (Limanda aspera) frame protein and its antihypertensive effect in spontaneously hypertensive rats. Food Chem. 2006, 94, 26. [Google Scholar]
- Cavazos, A.; Gonzalez de Mejia, E. Identification of bioactive peptides from cereal storage proteins and their potential role in prevention of chronic diseases. Compr. Rev. Food Sci. Food Saf. 2013, 12, 364–380. [Google Scholar] [CrossRef]
- Biel, W.; Bobko, K.; Maciorowski, R. Chemical composition and nutritive value of husked and naked oats grain. J. Cereal Sci. 2009, 49, 413–418. [Google Scholar] [CrossRef]
- Minkiewicz, P.; Dziuba, J.; Iwaniak, A.; Dziuba, M.; Darewicz, M. Biopep database and other programs for processing bioactive peptide sequences. J. AOAC Int. 2008, 91, 965–980. [Google Scholar] [PubMed]
- Lafarga, T.; O’Connor, P.; Hayes, M. Identification of novel dipeptidyl peptidase-IV and angiotensin-I-converting enzyme inhibitory peptides from meat proteins using in silico analysis. Peptides 2014, 59, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Mooney, C.; Haslam, N.J.; Pollastri, G.; Shields, D.C. Towards the improved discovery and design of functional peptides: Common features of diverse classes permit generalized prediction of bioactivity. PLoS ONE 2012, 7, e45012. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Aluko, R.E. Identification and inhibitory properties of multifunctional peptides from pea protein hydrolysate. J. Agric. Food Chem. 2010, 58, 11471–11476. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Aluko, R.E.; Nakai, S. Structural requirements of angiotensin-I-converting enzyme inhibitory peptides: Quantitative structure-activity relationship study of di- and tripeptides. J. Agric. Food Chem. 2006, 54, 732–738. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, J.Á.G.; Ramos, M.; Recio, I. Angiotensin converting enzyme-inhibitory activity of peptides isolated from Manchego cheese. Stability under simulated gastrointestinal digestion. Int. Dairy J. 2004, 14, 1075–1080. [Google Scholar]
- Ondetti, M.A.; Rubin, B.; Cushman, D.W. Design of specific inhibitors of Angiotensin-converting enzyme: New class of orally active antihypertensive agents. Science 1977, 196, 441–444. [Google Scholar] [CrossRef] [PubMed]
- Nongonierma, A.B.; Mooney, C.; Shields, D.C.; FitzGerald, R.J. In silico approaches to predict the potential of milk protein-derived peptides as dipeptidyl peptidase-IV (DPP-IV) inhibitors. Peptides 2014, 57, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Nongonierma, A.B.; FitzGerald, R.J. Inhibition of dipeptidyl peptidase IV (DPP-IV) by proline containing casein-derived peptides. J. Funct. Foods 2013, 5, 1909–1917. [Google Scholar] [CrossRef]
- Picariello, G.; Ferranti, P.; Fierro, O.; Mamone, G.; Caira, S.; Di Luccia, A.; Monica, S.; Addeo, F. Peptides surviving the simulated gastrointestinal digestion of milk proteins: Biological and toxicological implications. J. Chromatogr. B 2010, 878, 295–308. [Google Scholar] [CrossRef] [PubMed]
- Homayouni, A.; Azizi, A.; Ehsani, M.; Yarmand, M.; Razavi, S. Effect of microencapsulation and resistant starch on the probiotic survival and sensory properties of synbiotic ice cream. Food Chem. 2008, 111, 50–55. [Google Scholar] [CrossRef]
- Cheung, H.S.; Wang, F.L.; Ondetti, M.A.; Sabo, E.F.; Cushman, D.W. Binding of peptide substrates and inhibitors of angiotensin-converting enzyme. Importance of the COOH-terminal dipeptide sequence. J. Biol. Chem. 1980, 255, 401–407. [Google Scholar] [PubMed]
- Lan, V.T.; Ito, K.; Ohno, M.; Motoyama, T.; Ito, S.; Kawarasaki, Y. Analyzing a dipeptide library to identify human dipeptidyl peptidase iv inhibitor. Food Chem. 2015, 175, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Yano, S.; Suzuki, K.; Funatsu, G. Isolation from alpha-zein of thermolysin peptides with Angiotensin-I-converting enzyme inhibitory activity. Biosci. Biotechnol. Biochem. 1996, 60, 661–663. [Google Scholar] [CrossRef] [PubMed]
- Bella, A.M.; Erickson, R.H.; Kim, Y.S. Rat intestinal brush border membrane Dipeptidyl aminopeptidase IV: Kinetic properties and substrate specificities of the purified enzyme. Arch. Biochem. Biophys. 1982, 218, 156–162. [Google Scholar] [CrossRef]
- Ashmarin, I.; Karazeeva, E.; Lyapina, L.; Samonina, G. The simplest proline-containing peptides PG, GP, PGP, and GPGG: Regulatory activity and possible sources of biosynthesis. BioChemistry 1998, 63, 119–124. [Google Scholar] [PubMed]
- Byun, H.-G.; Kim, S.-K. Structure and activity of angiotensin I converting enzyme inhibitory peptides derived from Alaskan pollack skin. J. Biochem. Mol. Biol. 2002, 35, 239–243. [Google Scholar] [CrossRef] [PubMed]
- Gallego, M.; Aristoy, M.-C.; Toldrá, F. Dipeptidyl peptidase IV inhibitory peptides generated in Spanish dry-cured ham. Meat Sci. 2014, 96, 757–761. [Google Scholar] [CrossRef] [PubMed]
- Saito, Y.; Wanezaki, K.; Kawato, A.; Imayasu, S. Structure and activity of Angiotensin I converting enzyme inhibitory peptides from sake and sake lees. Biosci. Biotechnol. Biochem. 1994, 58, 1767–1771. [Google Scholar] [CrossRef] [PubMed]
- Gangopadhyay, N.; Wynne, K.; O’Connor, P.; Gallagher, E.; Brunton, N.P.; Rai, D.K.; Hayes, M. In silico and in vitro analyses of the angiotensin-I-converting enzyme inhibitory activity of hydrolysates generated from crude barley (Hordeum vulgare) protein concentrates. Food Chem. 2016, 203, 367–374. [Google Scholar] [CrossRef] [PubMed]
- Sica, D.A.; Gehr, T. Angiotensin-Converting Enzyme Inhibitors. In Hypertension; Elsevier-Saunders: Philadelphia, PA, USA, 2005; pp. 669–682. [Google Scholar]
- Fein, A. ACE inhibitors worsen inflammatory pain. Med. Hypotheses 2009, 72, 757. [Google Scholar] [CrossRef] [PubMed]
- Sidorenkov, G.; Navis, G. Safety of ace inhibitor therapies in patients with chronic kidney disease. Expert Opin. Drug Saf. 2014, 13, 1383–1395. [Google Scholar] [CrossRef] [PubMed]
- Li, G.-H.; Le, G.-W.; Shi, Y.-H.; Shrestha, S. Angiotensin-I–converting enzyme inhibitory peptides derived from food proteins and their physiological and pharmacological effects. Nutr. Res. 2004, 24, 469–486. [Google Scholar] [CrossRef]
- FitzGerald, R.J.; Meisel, H. Milk protein-derived peptide inhibitors of angiotensin-I-converting enzyme. Br. J. Nutr. 2000, 84, 33–37. [Google Scholar] [CrossRef]
- Fisher, N.D.; Hollenberg, N.K. Renin inhibition: What are the therapeutic opportunities? J. Am. Soc. Nephrol. 2005, 16, 592–599. [Google Scholar] [CrossRef] [PubMed]
- Lawandi, J.; Gerber-Lemaire, S.; Juillerat-Jeanneret, L.; Moitessier, N. Inhibitors of prolyl oligopeptidases for the therapy of human diseases: Defining diseases and inhibitors. J. Med. Chem. 2010, 53, 3423–3438. [Google Scholar] [CrossRef] [PubMed]





| Protein | UniProt ID | Sequence ** | Amino Acid Length | Molecular Mass (Da) |
|---|---|---|---|---|
| 11S globulin | Q38780 | MATTSFPSMLFYFCIFLLFHGSMAQLFGQSSTPWQSSRQGGLRGCRFDRLQAFEPLRQVRSQAGITEYFDEQNEQFRCTGVSVIRRVIEPQGLVLPQYHNAPALVYILQGRGFTGLTFPGCPATFQQQFQPFDQSQFAQGQRQSQTIKDEHQRVQRFKQGDVVALPAGIVHWCYNDGDAPIVAIYVFDVNNNANQLEPRQKEFLLAGNNKREQQSGNNIFSGLSVQLLSEALGISQQAAQRIQSQNDQRGEIIRVSQGLQFLKPIVSQQVPGEQQVYQPIQTQEGQATQYQVGQSTQYQVGKSTPYQGGQSSQYQAGQSWDQSFNGLEENFCSLEARKNIENPQHADTYNPRAGRITRLNSKNFPILNIVQMSATRVNLYQNAILSPFWNINAHSVIYMIQGHARVQVVNNNGQTVFNDILRRGQLLIVPQHFVVLKKAEREGCQYISFKTNPNSMVSHIAGKSSILRALPIDVLANAYRISRQEARNLKNNRGEEFGAFTPKLTQKGFQSYQDIEEGSSSPVRASE | 527 | 59,406 |
| 12S globulin | P12615 | MATTRFPSLLFYSCIFLLCNGSMAQLFGQSFTPWQSSRQGGLRGCKFDRLQAFEPLRQVRSQAGITEYFDEQNEQFRCAGVSVIRRVIEPQGLLLPQYHNAPGLVYILQGRGFTGLTFPGCPATFQQQFQQFDQARFAQGQSKSQNLKDEHQRVHHIKQGDVVALPAGIVHWCYNDGDAPIVAVYVFDVNNNANQLEPRQKEFLLAGNNKREQQFGQNIFSGFSVQLLSEALGISQQAAQKIQSQNDQRGEIIRVSQGLQFLKPFVSQQGPVEHQAYQPIQSQQEQSTQYQVGQSPQYQEGQSTQYQSGQSWDQSFNGLEENFCSLEARQNIENPKRADTYNPRAGRITHLNSKNFPTLNLVQMSATRVNLYQNAILSPYWNINAHSVMHMIQGRARVQVVNNHGQTVFNDILRRGQLLIIPQHYVVLKKAEREGCQYISFKTTPNSMVSYIAGKTSILRALPVDVLANAYRISRQESQNLKNNRGEEFGAFTPKFAQTGSQSYQDEGESSSTEKASE | 518 | 58,545 |
| Avenin-3 | P80356 | MKTFLIFALLAMAATMATAQFDPSEQYQPYPEQQQPILQQQQMLLQQQQQMLLQQQPLLQVLQQQLNPCRQFLVQQCSPVAVVPFLRSQILQQSSCQVMRQQCCRQLEQIPEQLRCPAIHSVVQAIIMQQQQFFQPQMQQQFFQPQMQQVTQGIFQPQMQQVTQGIFQPQLQQVTQGIFQPQMQGQIEGMRAFALQALPAMCDVYVPPHCPVATAPLGGF | 220 | 25,275 |
| Avenin-E | Q09114 | TTTVQYNPSEQYQPYPEQQEPFVQQQPFVQQQQQPFVQQQQMFLQPLLQQQLNPCKQFLVQQCSPVAVVPFLRSQILRQAICQVARQQCCRQLAQIPEQLRCPAIHSVVQAIILQQQQQQQFFQPQLQQQVFQPQLQQVFNQPQQQAQFEGMRAFALQALPAMCDVYVPPQCPVATAPLGGF | 182 | 21,036 |
| Avenin-F | Q09097 | TTTVQYDPSEQYQPYPEQQEPFVQQQPPFVQQQQPFVQQQEPF | 43 | 5214 |
| Avenin-A | Q09095 | PSEQYQPYPEQQQPFLQQQPLELQQQQXXLVLFLQK | 36 | 4393 |
| Avenin | P27919 | MKIFFFLALLALVVSATFAQYAESDGSYEEVEGSHDRCQQHQMKLDSCREYVAERCTTMRDFPITWPWKWWKGGCEELRNECCQLLGQMPSECRCDAIWRSIQRELGGFFGTQQGLIGKRLKIAKSLPTQSTWALSAISPNSMVSHIAGKSSILRALPVDVLANAYRISRQEARNLKNNRGQESGVFTPKFTQTSFQPYPEGEDESSLINKASE | 214 | 24,230 |
| Tryptophanin/2S albumin | A7U440 | MKALFLLAFLALAASAAFAQQYADTGVGGWDGCMPEKARLNSCKDYVVERCLTLKDIPITWPWKWWKGGCESEVRSQCCMELNQIAPHCRCKAIWRAVQGELGGFLGFQQSEIMKQVHVAQSLPSRCNMGPNCNFPTNLGYY | 142 | 15,901 |
| Location | Group | Amino Acids | Reference | |
|---|---|---|---|---|
| Renin inhibitory peptide characteristics | N1/N-terminus | Hydrophobic | Ala, Gly, Val, Leu, Ile, Pro, Phe, Met, Trp | [38] |
| N2 | N/A | |||
| N3/C-terminus | Bulky | Trp, Val, Ile, Leu, Tyr, Met, Phe, | [38] | |
| ACE-I inhibitory peptide characteristics | N1/N-terminus | Hydrophobic (Small with low lipophilicity) | Val, Ile, Leu | [39] |
| N2 | Positively charged (Large with high lipophilicity & low electronic properties) | Leu, Arg | [39] | |
| N3/C-terminus | Aromatic acids (Small with low lipophilicity & high electronic properties) | Pro, Phe, Trp | [39] | |
| - | Leu | [40] | ||
| Positively charged | Lys, Arg | [41] | ||
| - | Pro | [41] | ||
| DPP-IV inhibitory peptide characteristics | N1/N-terminus | Hydrophobic or aromatic | Leu, Ile, Val, Phe, Trp, Try | [42] |
| N2 | N/A | |||
| N3/C-terminus | - | Pro, Ala |
| Peptide | Peptide Ranker Score | BIOPEP ID | Activity Description | IC50 | Mass (Da) | Reference |
|---|---|---|---|---|---|---|
| FG | 0.99 | 7605 | ACE-I inhibitor | 3700.0 μM | 222.229 | [43,44,45,46] |
| PF | 0.99 | 8854 | DPP-IV inhibitor | N/A | 262.294 | [47] |
| FL | 0.99 | 8555 | DPP-IV inhibitor | 399.58 μM | 278.337 | [43] |
| FY | 0.98 | 3556 | ACE-I inhibitor | 25 μM | 328.347 | [48] |
| FA | 0.96 | 3176 | DPP-IV inhibitor | N/A | 236.256 | [49] |
| FN | 0.95 | 8778 | DPP-IV inhibitor | N/A | 279.281 | [47] |
| MG | 0.94 | 7609 | ACE-I inhibitor | 4800 μM | 206.25 | [46] |
| PG | 0.88 | 7625 | ACE-I inhibitor | 17,000 μM | 172.169 | [46] |
| PG | 0.88 | 8855 | DPP-IV inhibitor | N/A | 172.169 | [47] |
| PG | 0.88 | 3285 | N/A | N/A | 172.169 | [50] |
| MR | 0.85 | 8836 | DPP-IV inhibitor | N/A | 305.386 | [47] |
| PL | 0.81 | 7513 | ACE-I inhibitor | 337.32 μM | 228.277 | [51] |
| PL | 0.81 | 8638 | DPP-IV inhibitor | N/A | 228.277 | [52] |
| PLG | 0.8 | 7510 | ACE-I inhibitor | 4.74 μM | 285.329 | [51] |
| PR | 0.79 | 3537 | ACE-I inhibitor | 4.10 μM | 271.305 | [53] |
| RG | 0.74 | 8882 | DPP-IV inhibitor | N/A | 231.24 | [47] |
| LG | 0.72 | 7619 | ACE-I inhibitor | 8800 μM | 188.212 | [46] |
| MA | 0.69 | 3173 | DPP-IV inhibitor | N/A | 220.277 | [49] |
| RL | 0.63 | 8886 | DPP-IV inhibitor | N/A | 287.348 | [47] |
| Peptide | Solubility in Water | Resistance to Digestion | Toxicity | Allergenicity Probability |
|---|---|---|---|---|
| FFG | Poor | No | Non-toxin | Non-allergen |
| IFFFL | Poor | No | Non-toxin | Non-allergen |
| PFL | Poor | No | Non-toxin | Non-allergen |
| WWK | Good | No | Non-toxin | 33.3% |
| WCY | Poor | No | Non-toxin | 33.3% |
| FPIL | Poor | No | Non-toxin | Non-allergen |
| CPA | Poor | Yes | Non-toxin | Non-allergen |
| FLLA | Poor | No | Non-toxin | Non-allergen |
| FEPL | Good | No | Non-toxin | Non-allergen |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bleakley, S.; Hayes, M.; O’ Shea, N.; Gallagher, E.; Lafarga, T. Predicted Release and Analysis of Novel ACE-I, Renin, and DPP-IV Inhibitory Peptides from Common Oat (Avena sativa) Protein Hydrolysates Using in Silico Analysis. Foods 2017, 6, 108. https://0-doi-org.brum.beds.ac.uk/10.3390/foods6120108
Bleakley S, Hayes M, O’ Shea N, Gallagher E, Lafarga T. Predicted Release and Analysis of Novel ACE-I, Renin, and DPP-IV Inhibitory Peptides from Common Oat (Avena sativa) Protein Hydrolysates Using in Silico Analysis. Foods. 2017; 6(12):108. https://0-doi-org.brum.beds.ac.uk/10.3390/foods6120108
Chicago/Turabian StyleBleakley, Stephen, Maria Hayes, Nora O’ Shea, Eimear Gallagher, and Tomas Lafarga. 2017. "Predicted Release and Analysis of Novel ACE-I, Renin, and DPP-IV Inhibitory Peptides from Common Oat (Avena sativa) Protein Hydrolysates Using in Silico Analysis" Foods 6, no. 12: 108. https://0-doi-org.brum.beds.ac.uk/10.3390/foods6120108