Phytochemicals and Traditional Use of Two Southernmost Chilean Berry Fruits: Murta (Ugni molinae Turcz) and Calafate (Berberis buxifolia Lam.)


Abstract
:1. Introduction
2. Traditional Knowledge around Murta and Calafate
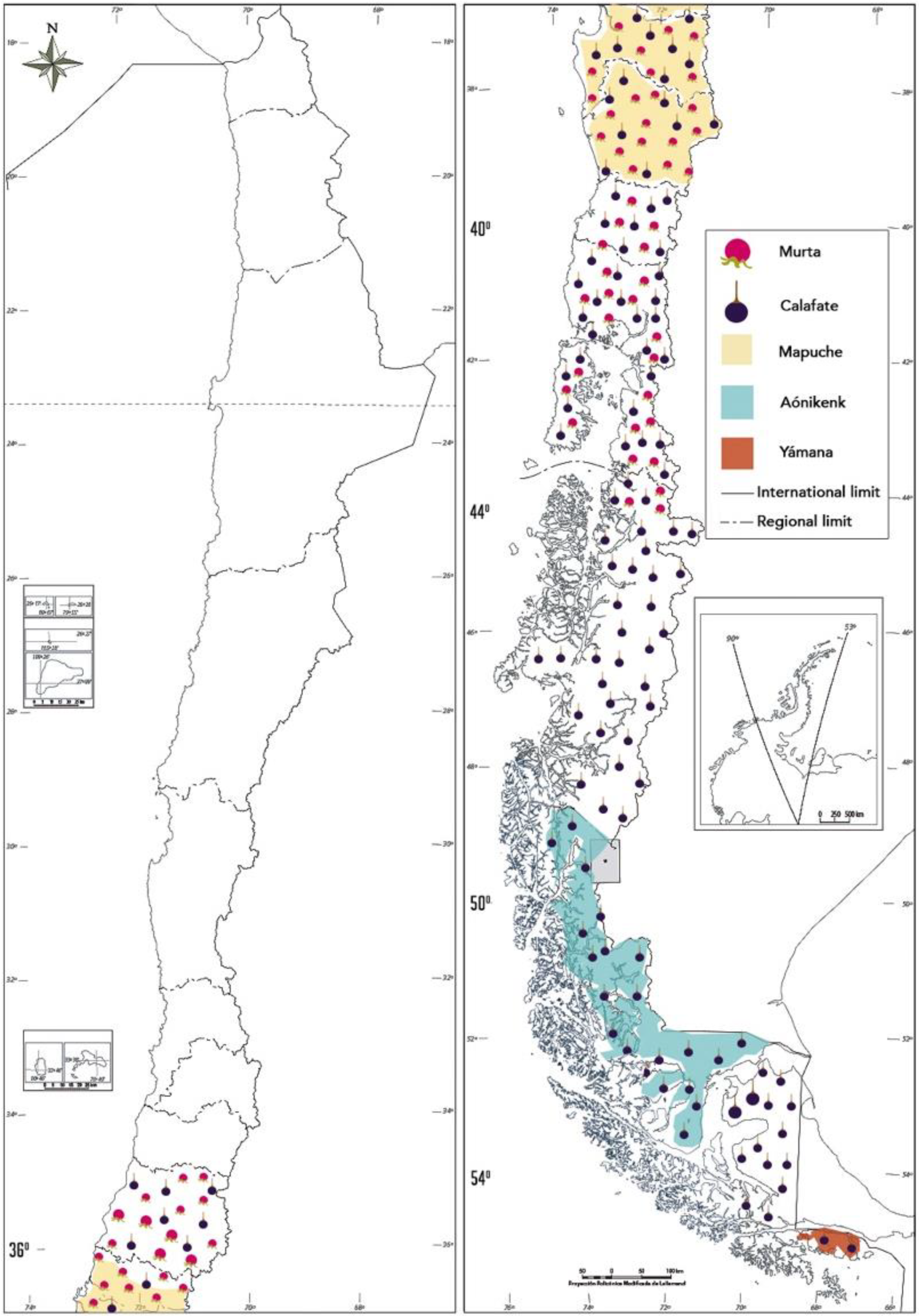
2.1. Murta and Calafate in the Mapuche and Rural Culture
2.2. Calafate in the Extreme South Communities of Chile
3. Scientific Knowledge around Murta and Calafate
3.1. Nutritional Content of Murta and Calafate Fruits
3.2. Phytochemicals in Murta and Calafate Fruits
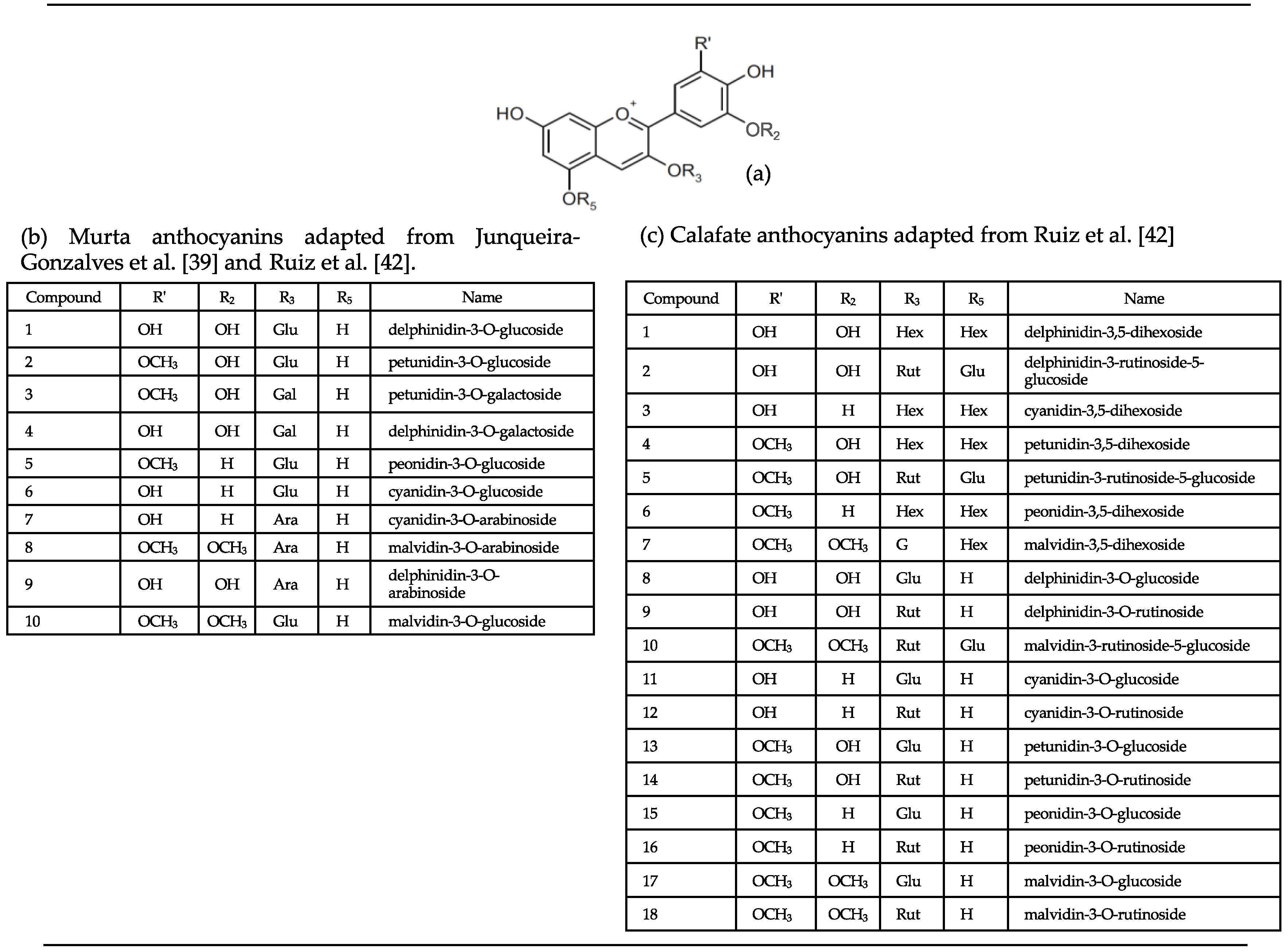
3.2.1. Anthocyanins in Murta and Calafate Fruit
3.2.2. Phenolic Acids and Other Flavonoids in Murta and Calafate Fruit
3.3. Phytochemical Changes in Murta and Calafate
3.3.1. Genotype vs. Environment
3.3.2. Stage at Fruit Harvest
3.3.3. Storage and Fruit Processing
3.4. Validation of Traditional Use and New Insights in the Research of Murta and Calafate
3.4.1. Antioxidant Capacity
3.4.2. Anti-Inflammatory Activity
3.4.3. Antimicrobial Activity
3.4.4. Analgesic Activity
3.4.5. New Insights in the Research of Murta and Calafate
3.5. Potential Health Benefits Associated with Murta and Calafate Fruit Consumption
4. Conclusions and Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wilhem De Mösbach, E. Botánica Indígena; Editorial Andrés Bello: Santiago, Chile, 1992; pp. 26–33. [Google Scholar]
- Estomba, D.; Ladio, A.; Lozada, M. Medicinal wild plant knowledge and gathering patterns in a Mapuche community from North-western Patagonia. J. Ethnopharmacol. 2006, 103, 109–119. [Google Scholar] [CrossRef]
- Lozada, M.; Ladio, A.; Weigandt, M. Cultural transmission of ethnobotanical knowledge in a rural community of northwestern Patagonia, Argentina. Econ. Bot. 2006, 60, 374–385. [Google Scholar] [CrossRef]
- Darwin, C. Viaje de un Naturalista Alrededor del Mundo; Joaquín Gil: Buenos Aires, Argentina, 1945; p. 618. [Google Scholar]
- Seguel, I.; Torralbo, L. Murtilla: El berry nativo del Sur de Chile. Rev. Tierra Adentro 2004, 57, 20–25. [Google Scholar]
- Hoffmann, A. Flora Silvestre de Chile. Zona Araucana. Árboles, Arbustos y Enredaderas Leñosas; Fundación Claudio Gay: Santiago, Chile, 1982; p. 258. [Google Scholar]
- Seeram, N.P. Berry fruits: Compositional elements, biochemical activities, and the impact of their intake on human health, performance, and disease. J. Agric. Food Chem. 2008, 56, 627–629. [Google Scholar] [CrossRef] [PubMed]
- Halvorsen, B.L.; Holte, K.; Myhrstad, M.C.W.; Barikmo, I.; Hvattum, E.; Remberg, S.F.; Wold, A.-B.; Haffner, K.; Baugerod, H.; Andersen, L.F.; et al. A systematic screening of total antioxidants in dietary plants. J. Nutr. 2002, 132, 461–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fredes, C.; Yousef, G.G.; Robert, P.; Grace, M.H.; Lila, M.A.; Gómez, M.; Gebauer, M.; Montenegro, G. Anthocyanin profiling of wild maqui berries (Aristotelia chilensis [Mol.] Stuntz) from different geographical regions in Chile. J. Sci. Food Agric. 2014, 94, 2639–2648. [Google Scholar] [CrossRef] [PubMed]
- Lee, S. Strategic design of delivery systems for nutraceuticals. In Nanotechnology Applications in Food: Flavor, Stability, Nutrition, and Safety; Oprea, O.E., Grumezescu, A.M., Eds.; Academic Press, Elsevier Inc.: London, UK, 2017; pp. 65–86. [Google Scholar]
- Molina, J.I. Compendio de la Historia Geográfica, Natural y Civil del Reyno de Chile; Don Antonio de Sancha: Madrid, España, 1788; p. 418. [Google Scholar]
- Gay, C. Historia Física y Política de Chile. Botánica. Tomo Primero; Museo de Historia Natural de Santiago: Santiago, Chile, 1845; p. 535. [Google Scholar]
- Gay, C. Historia Física y Política de Chile. Botánica. Tomo Segundo; Museo de Historia Natural de Santiago: Santiago, Chile, 1846; p. 372. [Google Scholar]
- Montenegro, G. Chile Nuestra Flora Útil; Universidad Católica de Chile: Santiago, Chile, 2002; p. 267. [Google Scholar]
- Fredes, C.; Montenegro, G. Chilean plants as a source of polyphenols. In Natural Antioxidants and Biocides from Wild Medicinal Plants; Céspedes, C., Sampietro, D., Seigler, D., Rai, M., Eds.; CAB International: Wallingford, UK, 2013; pp. 116–136. [Google Scholar]
- Minsal-Ministerio de Salud. Medicamentos Herbarios Tradicionales. 2009. Available online: https://www.minsal.cl/wp-content/uploads/2018/02/Libro-MHT-2010.pdf (accessed on 10 April 2018).
- Muñoz, O.; Montes, M.; Wilkomirsky, T. Plantas Medicinales de Uso en Chile. Química y Farmacología; Editorial Universitaria: Santiago, Chile, 2001; p. 330. [Google Scholar]
- Martinic, M. Los Aónikenk: Historia y Cultura; Universidad de Magallanes: Punta Arenas, Chile, 1995; p. 387. [Google Scholar]
- Barros, A. Aborígenes Australes de América; Editorial Lord Cochrane: Santiago, Chile, 1975; pp. 15–25. [Google Scholar]
- Cárdenas, R. Poemas Migratorios; Armando Menedin: Santiago, Chile, 1974; pp. 29–30. [Google Scholar]
- Torres, A.; Seguel, I.; Contreras, G.; Castro, M. Physico-chemical characterization of murta (murtilla) fruits Ugni molinae Turcz. Agric. Técnica 1999, 59, 260–270. [Google Scholar]
- Schreckinger, M.E.; Lotton, J.; Lila, M.A.; De Mejia, E.G. Berries from South America: A comprehensive review on chemistry, health potential, and commercialization. J. Med. Food 2010, 13, 233–246. [Google Scholar] [CrossRef]
- Scheuermann, E.; Seguel, I.; Montenegro, A.; Bustos, R.; Hormazabal, E.; Quiroz, A. Evolution of aroma compounds of murtilla fruits (Ugni molinae Turcz) during storage. J. Sci. Food Agric. 2008, 88, 485–492. [Google Scholar] [CrossRef]
- Ah-Hen, K.S.; Vega-Gálvez, A.; Moraga, N.O.; Lemus-Mondaca, R. Modelling of rheological behaviour of pulps and purées from fresh and frozen-thawed murta (Ugni molinae Turcz) berries. Int. J. Food Eng. 2012, 8. [Google Scholar] [CrossRef]
- Arancibia-Avila, P.; Namiesnik, J.; Toledo, F.; Werner, E.; Martinez-Ayala, A.M.; Rocha-Guzmán, N.E.; Gallegos-Infante, J.A.; Gorinstein, S. The influence of different time durations of thermal processing on berries quality. Food Control 2012, 26, 587–593. [Google Scholar] [CrossRef]
- Arena, M.E.; Zuleta, A.; Dyner, L.; Constenla, D.; Ceci, L.; Curvetto, N. Berberis buxifolia fruit growth and ripening: Evolution in carbohydrate and organic acid contents. Sci. Hortic. 2013, 158, 52–58. [Google Scholar] [CrossRef]
- Fredes, C.; Montenegro, G.; Zoffoli, J.; Santander, F.; Robert, P. Comparison of total phenolic, total anthocyanin and antioxidant activity of polyphenol-rich fruits grown in Chile. Cienc. Investig. Agrar. 2014, 41, 49–60. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, A.; Hermosin-Gutierrez, I.; Mardones, C.; Vergara, C.; Herlitz, E.; Vega, M.; Dorau, C.; Winterhalter, P.; Von Baer, D. Polyphenols and antioxidant activity of calafate (Berberis microphylla) fruits and other native berries from southern Chile. J. Agric. Food Chem. 2010, 58, 6081–6089. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.H. Potential synergy of phytochemicals in cancer prevention: Mechanism of action. J. Nutr. 2004, 134, 3479S–3485S. [Google Scholar] [CrossRef] [Green Version]
- Reilly, K. On-farm and fresh produce management. In Handbook of Plant Food Phytochemicals: Sources Stability and Extraction; Tiwari, B.K., Brunton, N., Brennan, C., Eds.; Wiley-Blackwell Publishing Co.: Hoboken, NJ, USA, 2013; pp. 201–235. [Google Scholar]
- Springob, K.; Kutchan, T.M. Introduction to the different classes of natural products. In Plant-Derived Natural Products; Osbourn, A.E., Lanzotti, V., Eds.; Springer: New York, NY, USA, 2009; pp. 1–50. [Google Scholar]
- Robert, P.; Fredes, C. The encapsulation of anthocyanins from berry-type fruits: Trends in foods. Molecules 2015, 20, 5875–5888. [Google Scholar] [CrossRef]
- Manach, C.; Scalbert, A.; Morand, C.; Rémésy, C.; Jiménez, L. Polyphenols: Food sources and bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [CrossRef] [Green Version]
- Castañeda-Ovando, A.; Pacheco-Hernández, M.L.; Páez-Hernández, M.A.; Rodríguez, J.A.; Galán-Vidal, C.A. Chemical studies of anthocyanins: A review. Food Chem. 2009, 113, 859–871. [Google Scholar] [CrossRef]
- Khoo, H.E.; Azlan, A.; Tang, S.T.; Lim, S.M. Anthocyanindins and anthocyanins: Colored pigments as food, pharmaceutical ingredients, and the potential health benefits. Food Nutr. Res. 2017, 61, 1361779. [Google Scholar] [CrossRef] [Green Version]
- Robert, P.; García, P.; Fredes, C. Drying and preservation of polyphenols. In Advances in Technologies for Producing Food-Relevant Polyphenols; Cuevas-Valenzuela, J., Vergara Salinas, J.R., Pérez-Correa, J.R., Eds.; CRC Press, Taylor and Francis Group: Boca Raton, FL, USA, 2017; pp. 281–302. [Google Scholar]
- Brito, A.; Areche, C.; Sepulveda, B.; Kennelly, E.J.; Simirgiotis, M.J. Anthocyanin characterization, total phenolic quantification and antioxidant features of some Chilean edible berry extracts. Molecules 2014, 19, 10936–10955. [Google Scholar] [CrossRef] [Green Version]
- Ramirez, J.E.; Zambrano, R.; Sepulveda, B.; Kennelly, E.J.; Simirgiotis, M.J. Anthocyanins and antioxidant capacities of six Chilean berries by HPLC-HR-ESI-ToF-MS. Food Chem. 2015, 176, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Junqueira-Goncalves, M.P.; Yanez, L.; Morales, C.; Navarro, M.; Contreras, R.A.; Zuniga, G.E. Isolation and characterization of phenolic compounds and anthocyanins from murta (Ugni molinae Turcz) fruits. Assessment of antioxidant and antibacterial activity. Molecules 2015, 20, 5698–5713. [Google Scholar] [CrossRef] [PubMed]
- Augusto, T.R.; Scheuerman, E.S.; Alencar, S.M.; D’Arce, M.A.; Costa De Camargo, A.; Vieira, T.M. Phenolic compounds and antioxidant activity of hydroalcoholic extracts of wild and cultivated murtilla (Ugni molinae Turcz). Food Sci. Technol. 2014, 34, 667–673. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, A.; Mardones, C.; Vergara, C.; Hermosin-Gutierrez, I.; Von Baer, D.; Hinrichsen, P.; Rodriguez, R.; Arribillaga, D.; Dominguez, E. Analysis of hydroxycinnamic acids derivatives in calafate (Berberis microphylla G. Forst) berries by liquid chromatography with photodiode array and mass spectrometry detection. J. Chromatogr. A 2013, 1281, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, A.; Mardones, C.; Vergara, C.; Von Baer, D.; Gomez-Alonso, S.; Gomez, M.V.; Hermosin-Gutierrez, I. Isolation and structural elucidation of anthocyanidin 3,7-beta-o-diglucosides and caffeoyl-glucaric acids from calafate berries. J. Agric. Food Chem. 2014, 62, 6918–6925. [Google Scholar] [CrossRef] [PubMed]
- Meyer, R.S.; DuVal, A.E.; Jensen, H.R. Patterns and processes in crop domestication: An historical review and quantitative analysis of 203 global food crops. New Phytol. 2012, 196, 29–48. [Google Scholar] [CrossRef] [PubMed]
- Chacon-Fuentes, M.; Parra, L.; Rodriguez-Saona, C.; Seguel, I.; Ceballos, R.; Quiroz, A. Domestication in murtilla (Ugni molinae) reduced defensive flavonol levels but increased resistance against a native herbivorous insect. Environ. Entomol. 2015, 44, 627–637. [Google Scholar] [CrossRef]
- Chacon-Fuentes, M.; Parra, L.; Lizama, M.; Seguel, I.; Urzua, A.; Quiroz, A. Plant flavonoid content modified by domestication. Environ. Entomol. 2017, 46, 1080–1089. [Google Scholar] [CrossRef]
- Alfaro, S.; Mutis, A.; Palma, R.; Quiroz, A.; Seguel, I.; Scheuermann, E. Influence of genotype and harvest year on polyphenol content and antioxidant activity in murtilla (Ugni molinae Turcz) fruit. J. Soil Sci. Plant Nutr. 2013, 13, 67–78. [Google Scholar] [CrossRef] [Green Version]
- Mariangel, E.; Reyes-Diaz, M.; Lobos, W.; Bensch, E.; Schalchli, H.; Ibarra, P. The antioxidant properties of calafate (Berberis microphylla) fruits from four different locations in southern Chile. Cienc. Investig. Agrar. 2013, 40, 161–170. [Google Scholar] [CrossRef] [Green Version]
- Arena, M.E.; Postemsky, P.D.; Curvetto, N.R. Changes in the phenolic compounds and antioxidant capacity of Berberis microphylla G. Forst berries in relation to light intensity and fertilization. Sci. Hortic. 2017, 218, 63–71. [Google Scholar] [CrossRef]
- Vvedenskaya, I.O.; Vorsa, N. Flavonoid composition over fruit development and maturation in American cranberry, Vaccinium macrocarpon Ait. Plant Sci. 2004, 167, 1043–1054. [Google Scholar] [CrossRef]
- Kulkarni, A.P.; Aradhya, S.M. Chemical changes and antioxidant activity in pomegranate arils during fruit development. Food Chem. 2005, 93, 319–324. [Google Scholar] [CrossRef]
- Wang, S.Y.; Chen, C.-T.; Wang, C.Y. The influence of light and maturity on fruit quality and flavonoid content of red raspberries. Food Chem. 2009, 112, 676–684. [Google Scholar] [CrossRef]
- Parr, A.J.; Bolwell, G.P. Phenols in the plant and in man. The potential for possible nutritional enhancement of the diet by modifying the phenols content or profile. J. Sci. Food Agric. 2000, 80, 985–1012. [Google Scholar] [CrossRef]
- Fredes, C.; Montenegro, G.; Zoffoli, J.P.; Gómez, M.; Robert, P. Polyphenol content and antioxidant activity of maqui (Aristotelia chilensis [Mol.] Stuntz) during fruit development and maturation in Central Chile. Chil. J. Agric. Res. 2012, 72, 582–589. [Google Scholar] [CrossRef] [Green Version]
- Arena, M.; Curvetto, N. Berberis buxifolia fruiting: Kinetic growth behavior and evolution of chemical properties during the fruiting period and different growing seasons. Sci. Hortic. 2008, 118, 120–127. [Google Scholar] [CrossRef]
- Wang, S.Y.; Chen, H.; Camp, M.J.; Ehlenfeldt, M.K. Genotype and growing season influence blueberry antioxidant capacity and other quality attributes. Int. J. Food Sci. Technol. 2012, 47, 1540–1549. [Google Scholar] [CrossRef]
- Patras, A.; Brunton, N.P.; O’Donnell, C.; Tiwari, B.K. Effect of thermal processing on anthocyanin stability in foods; mechanisms and kinetics of degradation. Trends Food Sci. Technol. 2010, 21, 3–11. [Google Scholar] [CrossRef]
- De Pascual-Teresa, S.; Sanchez-Ballesta, M.T. Anthocyanins: From plant to health. Phytochem. Rev. 2008, 7, 281–299. [Google Scholar] [CrossRef]
- Rhim, J.-W. Kinetics of thermal degradation of anthocyanin pigment solutions driven from red flower cabbage. Food Sci. Biotech. 2002, 11, 361–364. [Google Scholar]
- Mundombe Sinela, A.; Mertz, C.; Achir, N.; Rawat, N.; Vidot, K.; Fulcrand, H.; Dornier, M. Exploration of reaction mechanisms of anthocyanin degradation in a roselle extract through kinetic studies on formulated model media. Food Chem. 2017, 235, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Sharma, H.P.; Sugandha, P.H. Enzymatic added extraction and clarification of fruit juices-A review. Crit. Rev. Food Sci. Nutr. 2017, 57, 1215–1227. [Google Scholar] [CrossRef] [PubMed]
- Reyes, A.; Bubnovich, V.; Bustos, R.; Vásquez, M.; Vega, R.; Scheuermann, E. Comparative study of different process conditions of freeze drying of ‘Murtilla’ berry. Dry. Technol. 2010, 28, 1416–1425. [Google Scholar] [CrossRef]
- López, J.; Ah-Hen, K.S.; Vega-Gálvez, A.; Morales, A.; García-Segovia, P.; Uribe, E. Effects of drying methods on quality attributes of murta (Ugni molinae Turcz) berries: Bioactivity, nutritional aspects, texture profile, microstructure and functional properties. J. Food Process Eng. 2017, 40, e12511. [Google Scholar] [CrossRef]
- Puente-Díaz, L.; Ah-Hen, K.; Vega-Gálvez, A.; Lemus-Mondaca, R.; Di Scala, K. Combined infrared-convective drying of murta (Ugni molinae Turcz) berries: Kinetic modeling and quality assessment. Dry. Technol. 2013, 31, 329–338. [Google Scholar] [CrossRef]
- Ah-Hen, K.; Zambra, C.E.; Aguëro, J.E.; Vega-Gálvez, A.; Lemus-Mondaca, R. Moisture diffusivity coefficient and convective drying modelling of murta (Ugni molinae Turcz): Influence of temperature and vacuum on drying kinetics. Food Bioprocess Technol. 2013, 6, 919–930. [Google Scholar] [CrossRef]
- López, J.; Vega-Gálvez, A.; Bilbao-Sainz, C.; Chiou, B.-S.; Uribe, E.; Quispe-Fuentes, I. Influence of vacuum drying temperature on: Physico-chemical composition and antioxidant properties of murta berries. J. Food Process Eng. 2017, 40, e12569. [Google Scholar]
- Avello, M.; Pastene, E. Actividad antioxidante de infusos de Ugni molinae Turcz (“Murtilla”). Bol. Latinoam. Caribe Plantas Med. Aromat. 2005, 4, 33–39. [Google Scholar]
- Rubilar, M.; Pinelo, M.; Ihl, M.; Scheuermann, E.; Sineiro, J.; Nuñez, M.J. Murta leaves (Ugni molinae Turcz) as a source of antioxidant polyphenols. J. Agric. Food Chem. 2006, 54, 59–64. [Google Scholar] [CrossRef]
- Delporte, C.; Backhouse, N.; Inostroza, V.; Aguirre, M.C.; Peredo, N.; Silva, X.; Negrete, R.; Miranda, H.F. Analgesic activity of Ugni molinae (murtilla) in mice models of acute pain. J. Ethnopharmacol. 2007, 112, 162–165. [Google Scholar] [CrossRef] [PubMed]
- Aguirre, M.C.; Delporte, C.; Backhouse, N.; Erazo, S.; Letelier, M.E.; Cassels, B.K.; Silva, X.; Alegría, S.; Negrete, R. Topical anti-inflammatory activity of 2α-hydroxy pentacyclic triterpene acids from the leaves of Ugni molinae. Bioorg. Med. Chem. 2006, 14, 5673–5677. [Google Scholar] [CrossRef] [PubMed]
- Goity, L.E.; Queupil, M.J.; Jara, D.; Alegria, S.E.; Pena, M.; Barriga, A.; Aguirre, M.C.; Delporte, C. An HPLC-UV and HPLC-ESI-MS based method for identification of anti-inflammatory triterpenoids from the extracts of Ugni molinae. Bol. Latinoam. Caribe Plantas Med. Aromat. 2013, 12, 108–116. [Google Scholar]
- Arancibia-Radich, J.; Pena-Cerda, M.; Jara, D.; Valenzuela-Bustamante, P.; Goity, L.; Valenzuela-Barra, G.; Silva, X.; Garrido, G.; Delporte, C.; Seguel, I. Comparative study of anti-inflammatory activity and qualitative-quantitative composition of triterpenoids from ten genotypes of Ugni molinae. Bol. Latinoam. Caribe Plantas Med. Aromat. 2016, 15, 274–287. [Google Scholar]
- Avello, M.; Valdivia, R.; Mondaca, M.A.; Ordoñez, J.L.; Bittner, M.; Becerra, J. Activity of Ugni molinae Turcz against microorganisms with clinical importance. Bol. Latinoam. Caribe Plantas Med. Aromat. 2009, 8, 141–144. [Google Scholar]
- Shene, C.; Reyes, A.K.; Villarroel, M.; Sineiro, J.; Pinelo, M.; Rubilar, M. Plant location and extraction procedure strongly alter the antimicrobial activity of murta extracts. Eur. Food Res. Technol. 2009, 228, 467–475. [Google Scholar] [CrossRef]
- Shene, C.; Canquil, N.; Jorquera, M.; Pinelo, M.; Rubilar, M.; Acevedo, F.; Vergara, C.; Von Baer, D.; Mardones, C. In vitro activity on human gut bacteria of murta leaf extracts (Ugni molinae Turcz), a native plant from Southern Chile. J. Food Sci. 2012, 77, M323–M329. [Google Scholar] [CrossRef]
- De Dicastillo, C.L.; Bustos, F.; Valenzuela, X.; Lopez-Carballo, G.; Vilarino, J.M.; Galotto, M.J. Chilean berry Ugni molinae Turcz fruit and leaves extracts with interesting antioxidant, antimicrobial and tyrosinase inhibitory properties. Food Res. Int. 2017, 102, 119–128. [Google Scholar] [CrossRef]
- Jofre, I.; Pezoa, C.; Cuevas, M.; Scheuermann, E.; Freires, I.A.; Rosalen, P.L.; De Alencar, S.M.; Romero, F. Antioxidant and vasodilator activity of Ugni molinae Turcz. (murtilla) and its modulatory mechanism in hypotensive response. Oxid. Med. Cell. Longev. 2016, 6513416. [Google Scholar] [CrossRef] [Green Version]
- Calfío, C.; Huidobro-Toro, J.P. Potent vasodilator and cellular antioxidant activity of endemic patagonian calafate berries (Berberis microphylla) with nutraceutical potential. Molecules 2019, 24, 2700. [Google Scholar] [CrossRef] [Green Version]
- Manosalva, L.; Mutis, A.; Urzua, A.; Fajardo, V.; Quiroz, A. Antibacterial activity of alkaloid fractions from Berberis microphylla G. Forst and study of synergism with ampicillin and cephalothin. Molecules 2016, 21, 76. [Google Scholar] [CrossRef] [Green Version]
- Manosalva, L.; Mutis, A.; Diaz, J.; Urzua, A.; Fajardo, V.; Quiroz, A. Identification of isoquinoline alkaloids from Berberis microphylla by HPLC ESI-MS/MS. Bol. Latinoam. Caribe Plantas Med. Aromat. 2014, 13, 324–335. [Google Scholar]
- Furrianca, M.C.; Alvear, M.; Zambrano, T.; Fajardo, V.; Salazar, L.A. Hypoglycemic effect of Berberis microphylla G. Forst root extract. Trop. J. Pharm. Res. 2017, 16, 2179–2184. [Google Scholar] [CrossRef] [Green Version]
- Pitta-Alvarez, S.I.; Medina-Bolivar, F.; Alvarez, M.A.; Scambatto, A.A.; Marconi, P.L. In vitro shoot culture and antimicrobial activity of Berberis buxifolia Lam. Vitr. Cell. Dev. Biol. Plant 2008, 44, 502–507. [Google Scholar] [CrossRef]
- Albrecht, C.; Pellarin, G.; Rojas, M.J.; Albesa, I.; Eraso, A.J. Beneficial effect of Berberis buxifolia Lam, Zizyphus mistol Griseb and Prosopis alba extracts on oxidative stress induced by chloramphenicol. Medicina (Buenos Aires) 2010, 70, 65–70. [Google Scholar]
- Reyes-Farias, M.; Vasquez, K.; Ovalle-Marin, A.; Fuentes, F.; Parra, C.; Quitral, V.; Jimenez, P.; Garcia-Diaz, D.F. Chilean native fruit extracts inhibit inflammation linked to the pathogenic interaction between adipocytes and macrophages. J. Med. Food. 2015, 18, 601–608. [Google Scholar] [CrossRef] [Green Version]
- Chang, H.C.; Peng, C.H.; Yeh, D.M.; Kao, E.S.; Wang, C.J. Hibiscus sabdariffa extract inhibits obesity and fat accumulation, and improves liver steatosis in humans. Food Funct. 2014, 5, 734–739. [Google Scholar] [CrossRef]
- De Ferrars, R.M.; Czank, C.; Zhang, Q. The pharmacokinetics of anthocyanins and their metabolites in humans. Br. J. Pharmacol. 2014, 171, 3268–3282. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Huang, H.; Zhao, X. Effects of flavonoids-rich Chinese bayberry (Myrica rubra Sieb. et Zucc.) pulp extracts on glucose consumption in human HepG2 cells. J. Funct. Foods 2015, 14, 144–153. [Google Scholar] [CrossRef]
- Pavlidou, E.; Giaginis, C.; Fasoulas, A.; Petridis, D. Evaluation of the effect of blueberries consumption on chronic diseases, illness prevention and health promotion. Nat. Prod. J. 2018, 8, 45–53. [Google Scholar] [CrossRef]
- Siddiq, M.; Dolan, K.D.; Perkins-Veazie, O.; Collins, J.K. Effect of pectinolytic and cellulytic enzymes on the physical, chemical, and antioxidant properties of blueberry (Vaccinium corymbosum L.) juice. LWT-Food Sci. Technol. 2018, 92, 127–132. [Google Scholar] [CrossRef]
- Speisky, H.; López-Alarcón, C.; Gómez, M.; Fuentes, J.; Sandoval-Vicuña, C. First web-based database on total phenolics and oxygen radical absorbance capacity (ORAC) of fruits produced and consumed within the South Andes Region of South America. J. Agric. Food Chem. 2012, 60, 8851–8859. [Google Scholar] [CrossRef]
- Lila, M.; Ribnicky, D.; Rojo, L.; Rojas-Silva, P.; Oren, A.; Havenaar, R.; Janle, E.; Raskin, I.; Yousef, G.; Grace, M. Complementary approaches to gauge the bioavailability and distribution of ingested berry polyphenolics. J. Agric. Food Chem. 2012, 60, 5763–5771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flores, F.; Singh, R.; Kerr, W.; Pegg, R.; Kong, F. Total phenolics content and antioxidant capacities of microencapsulated blueberry anthocyanins during in vitro digestion. Food Chem. 2014, 153, 272–278. [Google Scholar] [CrossRef] [PubMed]
- Mosele, J.I.; Macià, A.; Romero, M.P.; Motilva, M.J. Stability and metabolism of Arbutus unedo bioactive compounds (phenolics and antioxidants) under in vitro digestion and colonic fermentation. Food Chem. 2016, 201, 120–130. [Google Scholar] [CrossRef] [PubMed]
- Ah-Hen, K.S.; Mathias-Rettig, K.; Gomez-Perez, L.S.; Riquelme-Asenjo, G.; Lemus-Mondaca, R.; Munoz-Farina, O. Bioaccessibility of bioactive compounds and antioxidant activity in murta (Ugni molinae T.) berries juices. J. Food Meas. Charact. 2018, 12, 602–615. [Google Scholar] [CrossRef]
- Kay, C.D.; Mazza, G.J.; Holub, B.J. Anthocyanins exist in the circulation primarily as metabolites in adult men. J. Nutr. 2005, 135, 2582–2588. [Google Scholar] [CrossRef] [Green Version]
- Bustamante, L.; Pastene, E.; Duran-Sandoval, D.; Vergara, C.; Von Baer, D.; Mardones, C. Pharmacokinetics of low molecular weight phenolic compounds in gerbil plasma after the consumption of calafate berry (Berberis microphylla) extract. Food Chem. 2018, 268, 347–354. [Google Scholar] [CrossRef]
- Amin, H.P.; Czank, C.; Raheem, S.; Zhang, Q.; Botting, N.P.; Cassidy, A.; Kay, C.D. Anthocyanins and their physiologically relevant metabolites alter the expression of IL-6 and VCAM-1 in CD40L and oxidized LDL challenged vascular endothelial cells. Mol. Nutr. Food Res. 2015, 59, 1095–1106. [Google Scholar] [CrossRef] [Green Version]
- Warner, E.F.; Zhang, Q.; Raheem, Q.S.; O’Hagan, D.; O’Connell, M.A.; Kay, C.D. Common phenolic metabolites of flavonoids, but not their unmetabolized precursors, reduce the secretion of vascular cellular adhesion molecules by human endothelial cells. J. Nutr. 2016, 146, 465–473. [Google Scholar] [CrossRef] [Green Version]
- Warner, E.F.; Smith, M.J.; Zhang, Q.; Raheem, K.S.; O’Hagan, D.; O’Connell, M.A.; Kay, C.D. Signatures of anthocyanin metabolites identified in humans inhibit biomarkers of vascular inflammation in human endothelial cells. Mol. Nutr. Food Res. 2017, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abe, L.T.; Lajolo, F.M.; Genovese, M.I. Potential dietary sources of ellagic acid and other antioxidants among fruits consumed in Brazil: Jabuticaba (Myrciaria jaboticaba (Vell.) Berg). J. Sci. Food Agric. 2012, 92, 1679–1687. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Organ | Traditional Use | Biological Activity | Phytochemicals | Reference |
|---|---|---|---|---|
| Murta leaf | Urinary and throat infection | Antioxidant | NR | [66] |
| Antioxidant | Phenolic acids, hydrolyzable tannins, epicatechin, myricetin, quercetin | [67] | ||
| Analgesic | Flavonoids and triterpenoids | [68] | ||
| Anti-inflammatory | Triterpenoids | [69] | ||
| Anti-inflammatory | Triterpenoids and phenolic compounds | [70,71] | ||
| Antimicrobial | Catechin, rutin, isoquercitrin, ellagic acid, quercitrin, narcissin, isorhamnetin-3-O-glucoside | [72,73,74,75] | ||
| Murta fruit | Astringent | Antimicrobial | Isoquinoline alkaloids | [39] |
| Vasodilator | Gallic acid, catechin, quercetin-3-β-D-glucoside, myricetin, quercetin, and kaempferol | [76,77] | ||
| Calafate root | Control fever, anti-inflammatory, stomach pain, indigestion, colitis | Antimicrobial | Isoquinoline alkaloids | [78,79] |
| Hypoglycaemic | NR | [80] | ||
| Calafate shoot | NR | Antimicrobial | Isoquinoline alkaloids | [78,79,81] |
| Calafate fruit | NR | Antioxidant | NR | [82] |
| Anti-inflammatory | Phenolic compounds | [83] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fredes, C.; Parada, A.; Salinas, J.; Robert, P. Phytochemicals and Traditional Use of Two Southernmost Chilean Berry Fruits: Murta (Ugni molinae Turcz) and Calafate (Berberis buxifolia Lam.). Foods 2020, 9, 54. https://0-doi-org.brum.beds.ac.uk/10.3390/foods9010054
Fredes C, Parada A, Salinas J, Robert P. Phytochemicals and Traditional Use of Two Southernmost Chilean Berry Fruits: Murta (Ugni molinae Turcz) and Calafate (Berberis buxifolia Lam.). Foods. 2020; 9(1):54. https://0-doi-org.brum.beds.ac.uk/10.3390/foods9010054
Chicago/Turabian StyleFredes, Carolina, Alejandra Parada, Jaime Salinas, and Paz Robert. 2020. "Phytochemicals and Traditional Use of Two Southernmost Chilean Berry Fruits: Murta (Ugni molinae Turcz) and Calafate (Berberis buxifolia Lam.)" Foods 9, no. 1: 54. https://0-doi-org.brum.beds.ac.uk/10.3390/foods9010054