Impact of Water Activity on the Inactivation and Gene Expression of Listeria monocytogenes during Refrigerated Storage of Pressurized Dry-Cured Ham


Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms
2.2. Adjustment of aw Values in Sliced-Dry Cured Ham
2.3. Dry-Cured Ham Inoculation and HPP Treatments
2.4. Microbiological Analysis
2.5. Physicochemical Analysis
2.6. RNA Extraction and cDNA Synthesis
2.7. Quantitative PCR
2.8. Data and Statistical Analysis
3. Results
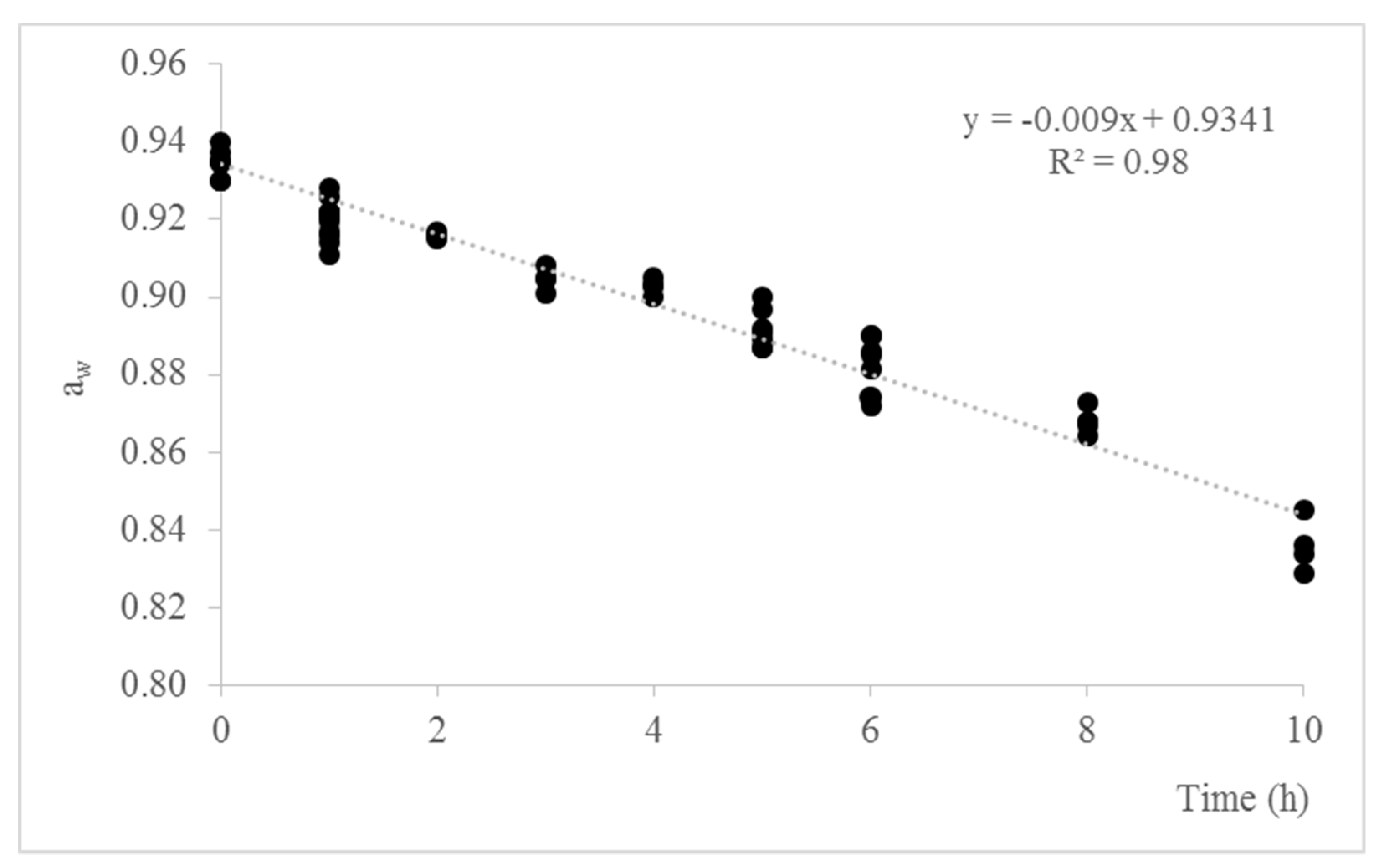
3.1. Adjustment of aw in Sliced Dry-Cured Ham
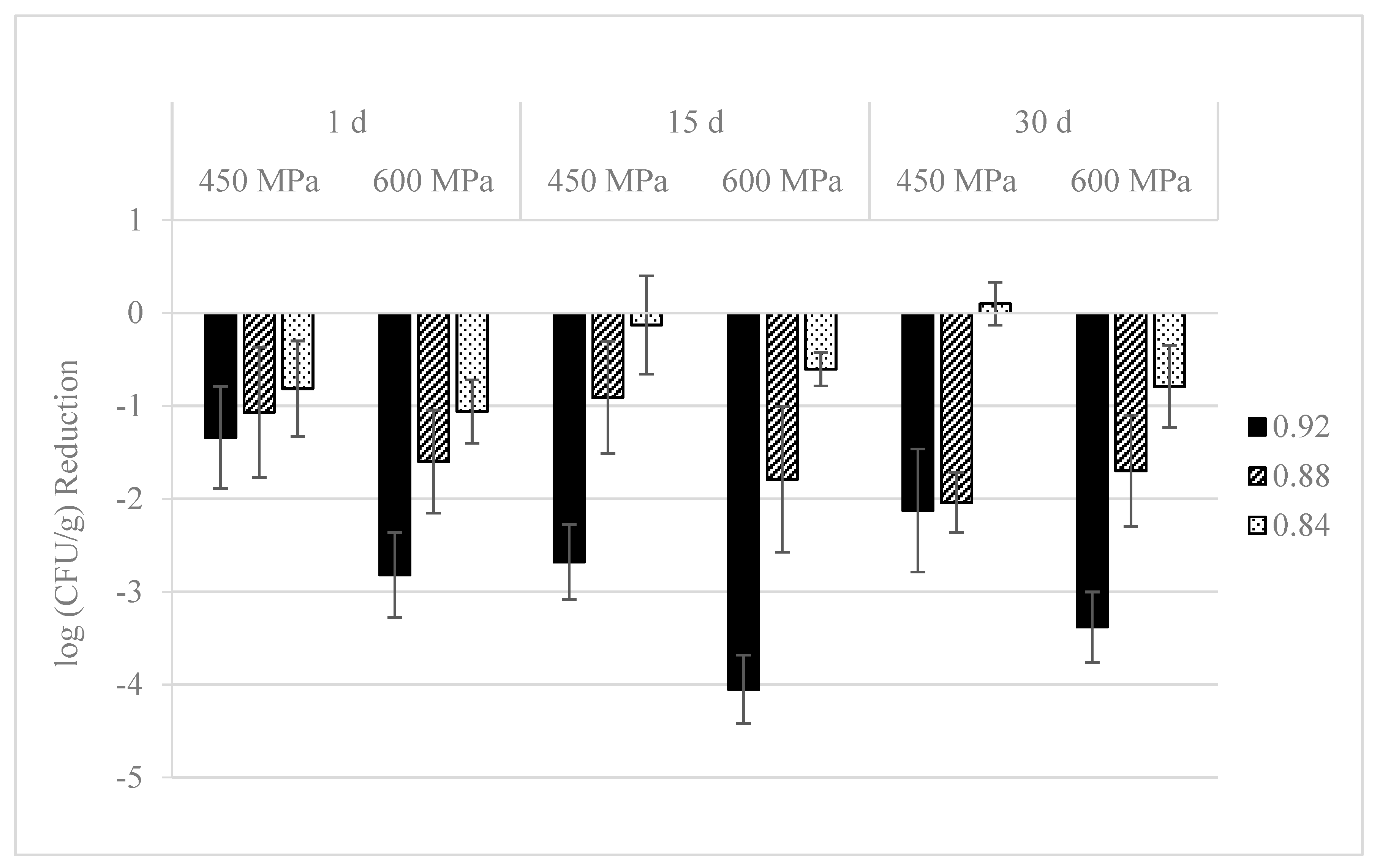
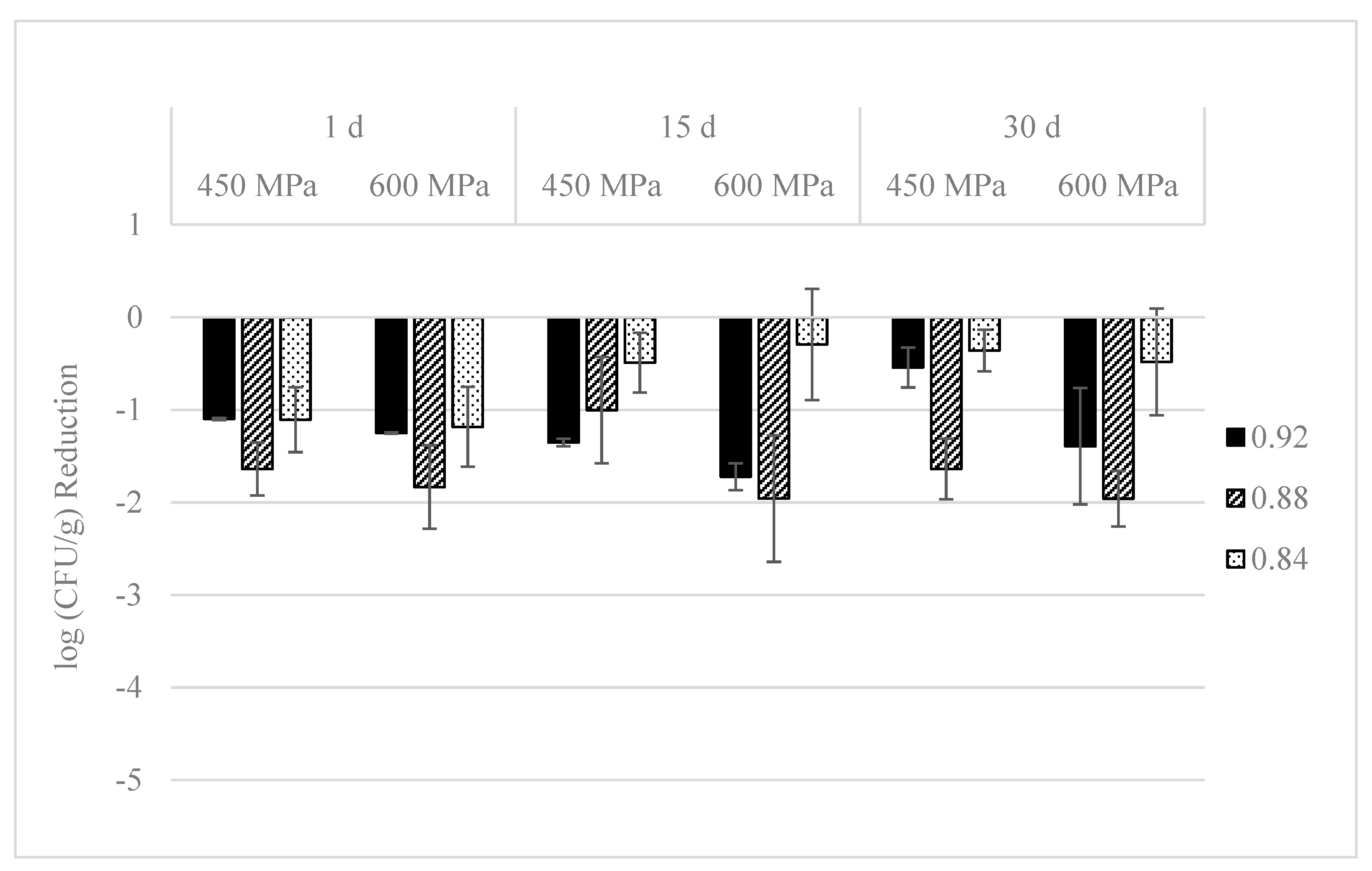
3.2. Effect on L. monocytogenes Population
3.3. Effect on TVC and pH
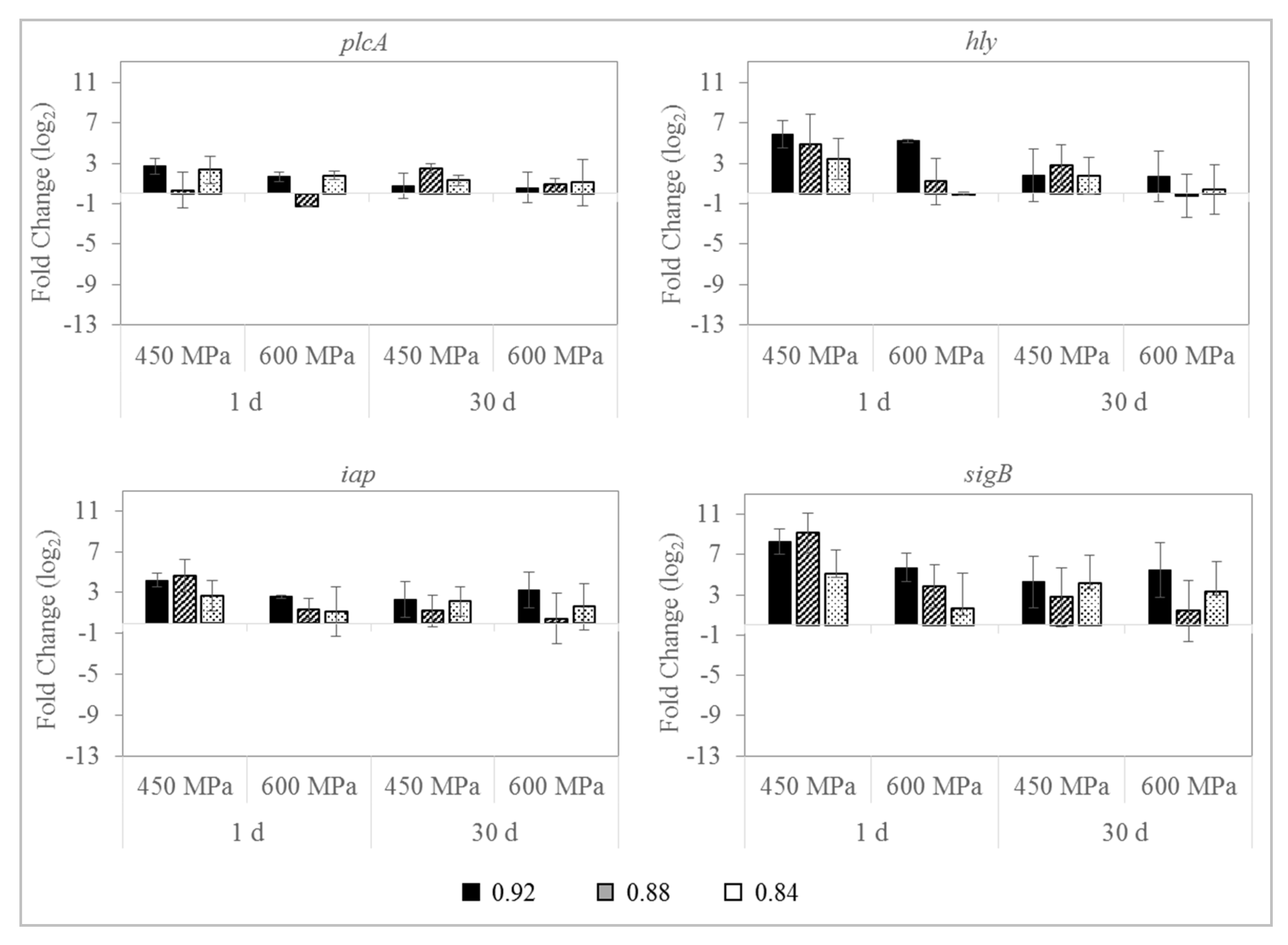
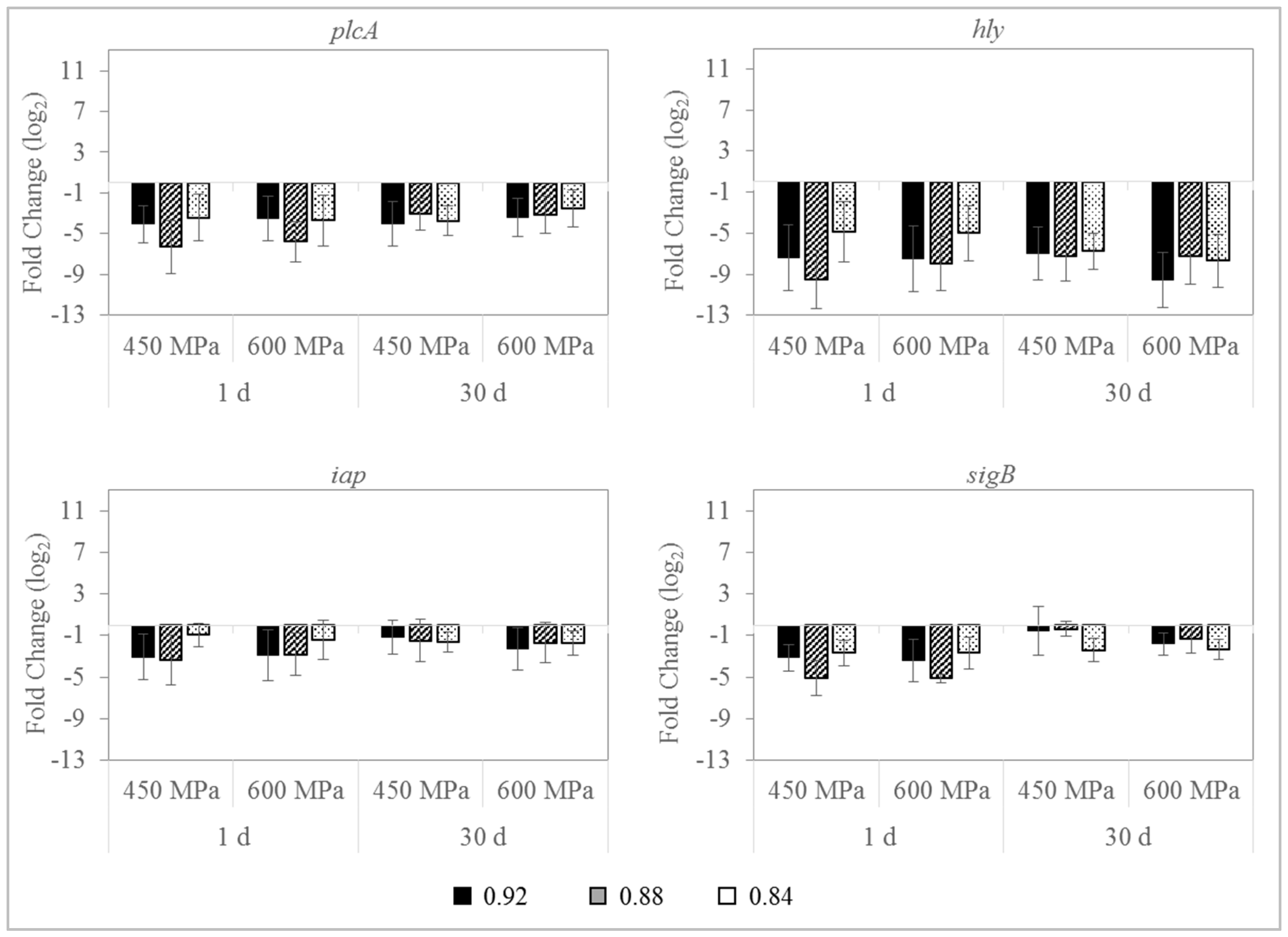
3.4. Effect on L. monocytogenes Gene Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- European Food Safety Authority (EFSA). Scientific report on the European Union One Health 2018 Zoonoses Report. EFSA J. 2019, 17, 1–276. [Google Scholar]
- Lado, B.; Yousef, A.E. Characteristics of Listeria monocytogenes important to food processors. In Listeria, Listeriosis and Food Safety, 3rd ed.; Ryser, E.T., Marth, E.H., Eds.; CRC Press Taylor & Francis Group: Boca Raton, FL, USA, 2007; pp. 157–213. [Google Scholar]
- European Commission (EC). Commission regulation (EC) No 2073/2005 of 15 November 2005 on microbiological criteria for foodstuffs. Off. J. Eur. 2005, L338, 1–29. [Google Scholar]
- Food Safety and Inspection Service (USDA-FSIS). FSIS Compliance Guideline: Controlling Listeria monocytogenes in Post-Lethality Exposed Ready-to-Eat Meat and Poultry Products. 2014. Available online: https://www.fsis.usda.gov/wps/wcm/connect/d3373299-50e6-47d6-a577-e74a1e549fde/Controlling-Lm-RTE-Guideline.pdf?MOD=AJPERES (accessed on 2 July 2020).
- Zhu, M.; Du, M.; Cordray, J.; Ahn, D.U. Control of Listeria monocytogenes contamination in ready-to-eat meat products. Compr. Rev. Food Sci. Food Saf. 2005, 4, 34–42. [Google Scholar] [CrossRef]
- Hugas, M.; Garriga, M.; Monfort, J.M. New mild technologies in meat processing: High pressure as a model technology. Meat Sci. 2002, 62, 359–371. [Google Scholar] [CrossRef]
- Campus, M. High Pressure Processing of Meat, Meat Products and Seafood. Food Eng. Rev. 2010, 2, 256–273. [Google Scholar] [CrossRef]
- Bover-Cid, S.; Belletti, N.; Aymerich, T.; Garriga, M. Modelling the impact of water activity and fat content of dry-cured ham on the reduction of Salmonella enterica by high pressure processing. Meat Sci. 2017, 123, 120–125. [Google Scholar] [CrossRef]
- Rubio, B.; Possas, A.; Rincón, F.; García-Gímeno, R.M.; Martínez, B. Model for Listeria monocytogenes inactivation by high hydrostatic pressure processing in Spanish chorizo sausage. Food Microbiol. 2018, 69, 18–24. [Google Scholar] [CrossRef]
- Ferreira, M.; Almeida, A.; Delgadillo, I.; Saraiva, J.; Cunha, A. Susceptibility of Listeria monocytogenes to high pressure processing: A review. Food Rev. Int. 2016, 32, 377–399. [Google Scholar] [CrossRef]
- Alía, A.; Rodríguez, A.; Andrade, M.J.; Gómez, F.M.; Córdoba, J.J. Combined effect of temperature, water activity and salt content on the growth and gene expression of Listeria monocytogenes in a dry-cured ham model system. Meat Sci. 2019, 155, 16–19. [Google Scholar] [CrossRef]
- Dorey, A.; Marinho, C.; Piveteau, P.; O’Byrne, C. Role and regulation of the stress activated sigma factor sigma B (σB) in the saprophytic and host-associated life stages of Listeria monocytogenes. Adv. Appl. Microbiol. 2019, 106, 1–48. [Google Scholar]
- Wemekamp-Kamphuis, H.H.; Wouters, J.A.; De Leeuw, P.P.L.A.; Hain, T.; Chakraborty, T.; Abee, T. Identification of sigma factor σB-controlled genes and their impact on acid stress, high hydrostatic pressure, and freeze survival in Listeria monocytogenes EGD-e. Appl. Environ. Microbiol. 2004, 70, 3457–3466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rantsiou, K.; Mataragas, M.; Alessandria, V.; Cocolin, L. Expression of virulence genes of Listeria monocytogenes in food. J. Food Saf. 2012, 32, 161–168. [Google Scholar] [CrossRef]
- Greppi, A.; Rantsiou, K. Methodological advancements in foodborne pathogen determination: From presence to behavior. Curr. Opin. Food Sci. 2016, 8, 80–88. [Google Scholar] [CrossRef]
- Ortiz, S.; López, V.; Villatoro, D.; López, P.; Dávila, J.C.; Martínez-Suárez, J.V. A 3-year surveillance of the genetic diversity and persistence of Listeria monocytogenes in an Iberian pig slaughterhouse and processing plant. Foodborne Pathog. Dis. 2010, 7, 1177–1184. [Google Scholar] [CrossRef]
- Andrade, M.J.; Thorsen, L.; Rodríguez, A.; Córdoba, J.J.; Jespersen, L. Inhibition of ochratoxigenic moulds by Debaryomyces hansenii strains for biopreservation of dry-cured meat products. Int. J. Food Microbiol. 2014, 170, 70–77. [Google Scholar] [CrossRef]
- International Organization for Standardization. Meat and Meat Products. Determination of Nitrite Content, ISO 2918; International Organization for Standardization: Geneva, Switzerland, 1975. [Google Scholar]
- Association of Official Analytical Chemists. Official Methods of Analysis, 19th ed.; AOAC 971:19; AOAC: Washington, DC, USA, 2012. [Google Scholar]
- Rodríguez-Lázaro, D.; Hernández, M.; Scortti, M.; Esteve, T.; Vázquez-Boland, J.A.; Pla, M. Quantitative detection of Listeria monocytogenes and Listeria innocua by Real-Time PCR: Assessment of hly, iap, and lin02483 targets and AmpliFluor technology. Appl. Environ. Microbiol. 2004, 70, 1366–1377. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Steel, R.G.D.; Torrie, J.H.; Dickey, D. Principles and Procedures of Statistics: A Biometrical Approach, 3rd ed.; McGraw-Hill: New York, NY, USA, 1996; pp. 1–666. [Google Scholar]
- Bover-Cid, S.; Belletti, N.; Aymerich, T.; Garriga, M. Modeling the protective effect of aw and fat content on the high pressure resistance of Listeria monocytogenes in dry-cured ham. Food Res. Int. 2015, 75, 194–199. [Google Scholar] [CrossRef]
- Hereu, A.; Bover-Cid, S.; Garriga, M.; Aymerich, T. High hydrostatic pressure and biopreservation of dry-cured ham to meet the Food Safety Objectives for Listeria monocytogenes. Int. J. Food Microbiol. 2012, 154, 107–112. [Google Scholar] [CrossRef]
- Patterson, M.F. Microbiology of pressure-treated foods. J. Appl. Microbiol. 2005, 98, 1400–1409. [Google Scholar] [CrossRef]
- Morales, P.; Calzada, J.; Nuñez, M. Effect of high-pressure treatment on the survival of Listeria monocytogenes Scott A in sliced vacuum-packaged Iberian and Serrano cured hams. J. Food Prot. 2006, 69, 2539–2543. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, M.; Núñez, F.; Córdoba, J.J.; Bermúdez, M.E.; Asensio, M.A. Evaluation of proteolytic activity of microorganisms isolated from dry cured ham. J. Appl. Microbiol. 1998, 85, 905–912. [Google Scholar] [CrossRef] [PubMed]
- Garriga, M.; Grèbol, N.; Aymerich, M.T.; Monfort, J.M.; Hugas, M. Microbial inactivation after high-pressure processing at 600 MPa in commercial meat products over its shelf life. Innov. Food Sci. Emerg. Technol. 2004, 5, 451–457. [Google Scholar] [CrossRef]
- Martínez-Onandi, N.; Castioni, A.; San Martín, E.; Rivas-Cañedo, A.; Nuñez, M.; Torriani, S.; Picon, A. Microbiota of high-pressure-processed Serrano ham investigated by culture-dependent and culture-independent methods. Int. J. Food Microbiol. 2017, 241, 298–307. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Baltar, A.; Serrano, A.; Bravo, D.; Montiel, R.; Medina, M. Combined effect of high pressure processing with enterocins or thymol on the inactivation of Listeria monocytogenes and the characteristics of sliced dry-cured ham. Food Bioprocess Technol. 2019, 12, 288–297. [Google Scholar] [CrossRef]
- Pérez-Baltar, A.; Serrano, A.; Montiel, R.; Medina, M. Listeria monocytogenes inactivation in deboned dry-cured hams by high pressure processing. Meat Sci. 2020, 160, 107960. [Google Scholar] [CrossRef]
- Montiel, R.; Quesille-Villalobos, A.; Alessandria, V.; Medina, M.; Cocolin, L.S.; Rantsiou, K. Antilisterial effect and influence on Listeria monocytogenes gene expression of enterocin or Enterococcus faecalis in sliced dry-cured ham stored at 7 °C. J. Food Prot. 2019, 82, 1598–1606. [Google Scholar] [CrossRef]
- Schrama, D.; Helliwell, N.; Neto, L.; Faleiro, M.L. Adaptation of Listeria monocytogenes in a simulated cheese medium: Effects on virulence using the Galleria mellonella infection model. Lett. Appl. Microbiol. 2013, 56, 421–427. [Google Scholar] [CrossRef]
- Chaturongakul, S.; Raengpradub, S.; Wiedmann, M.; Boor, K.J. Modulation of stress and virulence in Listeria monocytogenes. Trends Microbiol. 2008, 16, 388–396. [Google Scholar] [CrossRef] [Green Version]
- Sue, D.; Boor, K.J.; Wiedmann, M. σB-dependent expression patterns of compatible solute transporter genes opuCA and lmo1421 and the conjugated bile salt hydrolase gene bsh in Listeria monocytogenes. Microbiology 2003, 149, 3247–3256. [Google Scholar] [CrossRef] [Green Version]
- Mataragas, M.; Rovetto, F.; Bellio, A.; Alessandria, V.; Rantsiou, K.; Decastelli, L.; Cocolin, L. Differential gene expression profiling of Listeria monocytogenes in cacciatore and felino salami to reveal potential stress resistance biomarkers. Food Microbiol. 2015, 46, 408–417. [Google Scholar] [CrossRef] [PubMed]
- Olesen, I.; Thorsen, L.; Jespersen, L. Relative transcription of Listeria monocytogenes virulence genes in liver pâtés with varying NaCl content. Int. J. Food Microbiol. 2010, 141, S60–S68. [Google Scholar] [CrossRef]
- Bowman, J.P.; Bittencourt, C.R.; Ross, T. Differential gene expression of Listeria monocytogenes during high hydrostatic pressure processing. Microbiology 2008, 154, 462–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wuenscher, M.D.; Kohler, S.; Bubert, A.; Gerike, U.; Goebel, W. The iap gene of Listeria monocytogenes is essential for cell viability, and its gene product, p60, has bacteriolytic activity. J. Bacteriol. 1993, 175, 3491–3501. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Function and Scope of Use | Sequence (5′→3′) | Reference |
|---|---|---|---|
| IGS | Reference gene | F: GGCCTATAGCTCAGCTGGTTA | Rantsiou et al. 2012 [14] |
| R: GCTGAGCTAAGGCCCCATAAA | |||
| P: FAM-ATAAGAAATACAAATAATCATACCCTTTTAC-TAMRA | |||
| plcA | Escape from primary vacuoles | F: CTAGAAGCAGGAATACGGTACA | Rantsiou et al., 2012 [14] |
| R: ATTGAGTAATCGTTTCTAAT | |||
| P: HEX-AATTTATTTAAATGCATCACTTTCAGGT-TAMRA | |||
| hly | Lysis of vacuoles | F: CATGGCACCACCAGCATCT | Rodríguez-Lázaro et al., 2004 [20] |
| R: ATCCGCGTGTTTCTTTTCGA | |||
| P: HEX-CGCCTGCAAGTCCTAAGACGCCA-TAMRA | |||
| iap | Invasion associated secreted endopeptidase | F: AATCTTTAGCGCAACTTGGTTAA | Rodríguez-Lázaro et al., 2004 [20] |
| R: CACCTTTGATGGACGTAATAATACTGTT | |||
| P: HEX-CAACACCAGCGCCACTAGGACG-TAMRA | |||
| sigB | Transcription factor, regulation of virulence and stress-response genes | F: CCAAGAAAATGGCGATCAAGAC | Rantsiou et al., 2012 [14] |
| R: CGTTGCATCATATCTTCTAATAGCT | |||
| P: HEX-TGTTCATTACAAAAACCTAGTAGAGTCCAT-TAMRA |
| aw | NaCl (% wt/wt) | Nitrites (mg/kg) |
|---|---|---|
| 0.92 | 3.6 ± 0.3 a | 3.9 ± 0.3 a |
| 0.88 | 4.3 ± 1.3 a | 3.8 ± 0.0 a |
| 0.84 | 5.3 ± 0.0 a | 4.3 ± 0.2 a |
| aw | Treatment | Time (d) | ||
|---|---|---|---|---|
| 1 | 15 | 30 | ||
| 0.92 | NP | 5.82 ± 0.04 aA | 5.82 ± 0.08 aA | 5.89 ± 0.07 aA |
| 450 MPa/10 min | 5.88 ± 0.12 aA | 5.91 ± 0.08 aA | 5.87 ± 0.07 aA | |
| 600 MPa/5 min | 5.95 ± 0.08 aA | 6.06 ± 0.07 bB | 5.85 ± 0.02 aA | |
| 0.88 | NP | 5.88 ± 0.07 aB | 6.07 ± 0.06 bC | 5.78 ± 0.06 aA |
| 450 MPa/10 min | 5.86 ± 0.13 aAB | 5.98 ± 0.05 aB | 5.80 ± 0.05 aA | |
| 600 MPa/5 min | 5.92 ± 0.02 aB | 5.96 ± 0.04 aB | 5.86 ± 0.06 aA | |
| 0.84 | NP | 5.93 ± 0.08 aA | 6.12 ± 0.05 aB | 5.86 ± 0.03 abA |
| 450 MPa/10 min | 6.01 ± 0.12 aA | 6.12 ± 0.02 aB | 5.91 ± 0.03 bA | |
| 600 MPa/5 min | 5.97 ± 0.07 aB | 6.14 ± 0.12 aC | 5.83 ± 0.05 aA | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-Baltar, A.; Alía, A.; Rodríguez, A.; Córdoba, J.J.; Medina, M.; Montiel, R. Impact of Water Activity on the Inactivation and Gene Expression of Listeria monocytogenes during Refrigerated Storage of Pressurized Dry-Cured Ham. Foods 2020, 9, 1092. https://0-doi-org.brum.beds.ac.uk/10.3390/foods9081092
Pérez-Baltar A, Alía A, Rodríguez A, Córdoba JJ, Medina M, Montiel R. Impact of Water Activity on the Inactivation and Gene Expression of Listeria monocytogenes during Refrigerated Storage of Pressurized Dry-Cured Ham. Foods. 2020; 9(8):1092. https://0-doi-org.brum.beds.ac.uk/10.3390/foods9081092
Chicago/Turabian StylePérez-Baltar, Aida, Alberto Alía, Alicia Rodríguez, Juan José Córdoba, Margarita Medina, and Raquel Montiel. 2020. "Impact of Water Activity on the Inactivation and Gene Expression of Listeria monocytogenes during Refrigerated Storage of Pressurized Dry-Cured Ham" Foods 9, no. 8: 1092. https://0-doi-org.brum.beds.ac.uk/10.3390/foods9081092