
Effects of Soil Quality on the Microbial Community Structure of Poorly Evolved Mediterranean Soils

 , , , ,
, , , ,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Sampling
2.2. Soil Characteristic Determinations
2.3. Molecular Methods
2.4. Data Analyses
2.5. Community Interaction Networks
3. Results
3.1. Soil Use and Soil Properties
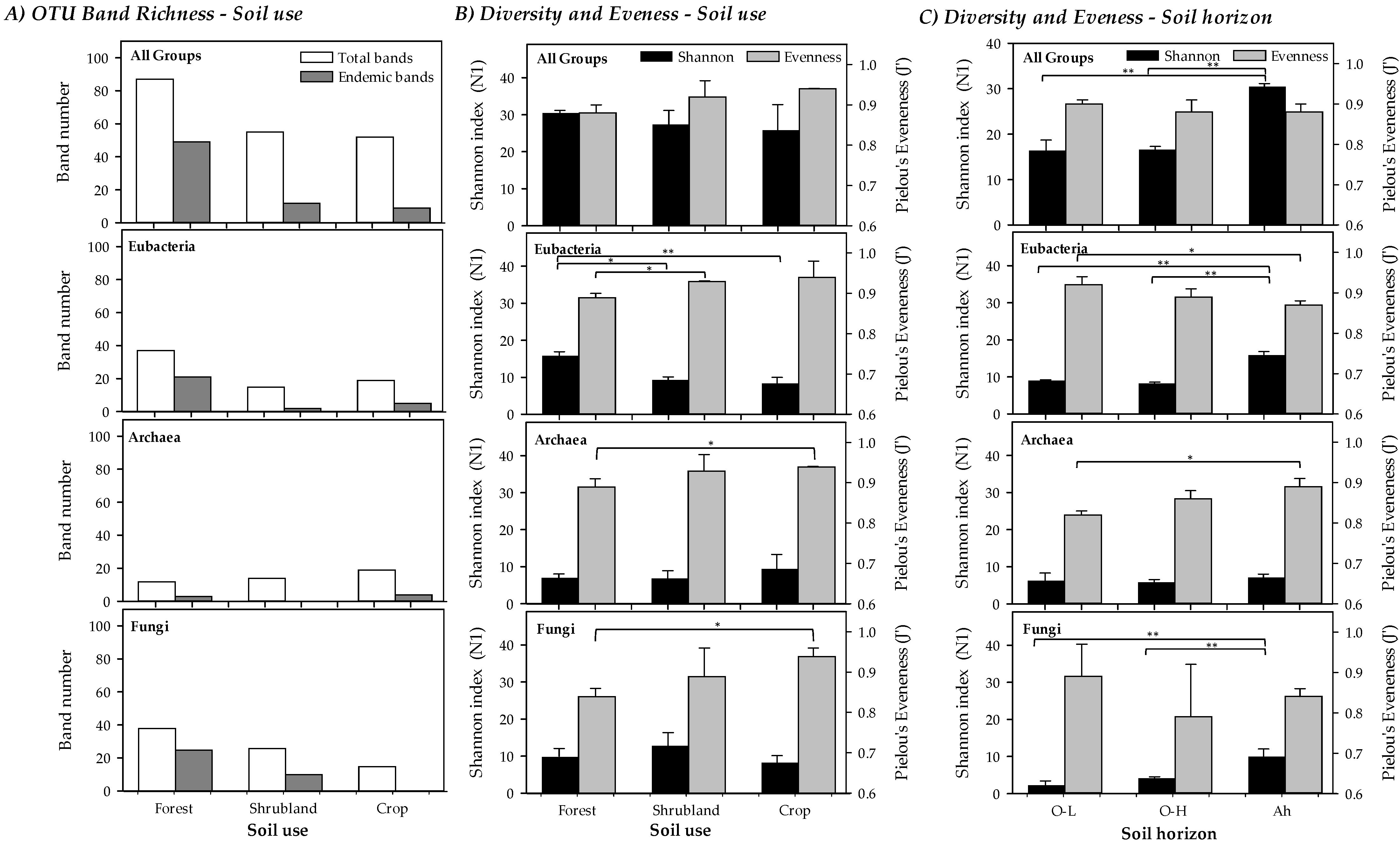
3.2. Soil Microbial Biomass and Diversity (Analysis of Soil Microbial Communities)
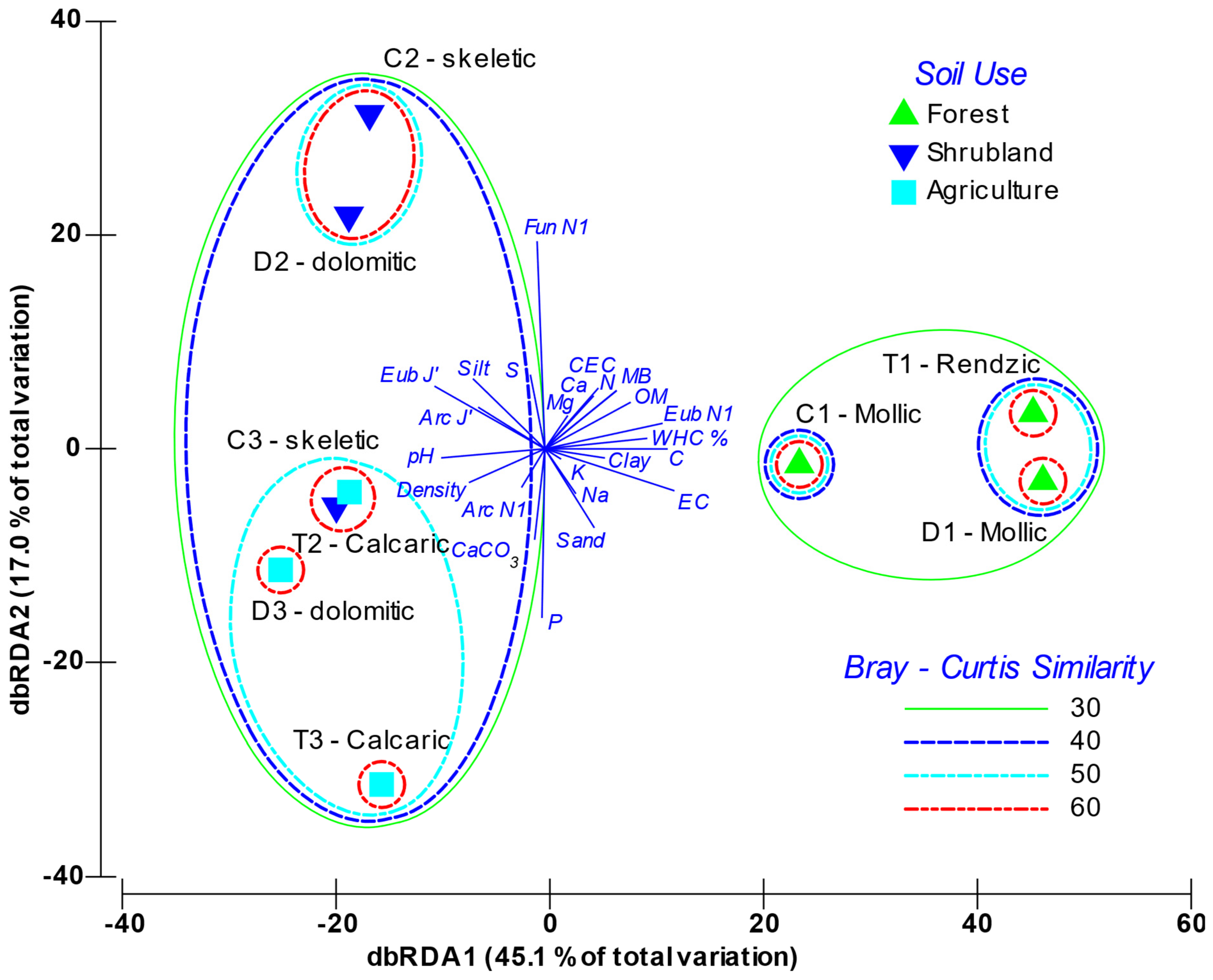
3.3. Relationship between Microbial Diversity and Soil Characteristics
3.4. Microbial Diversity of the Different Forest Soil Organic Horizons
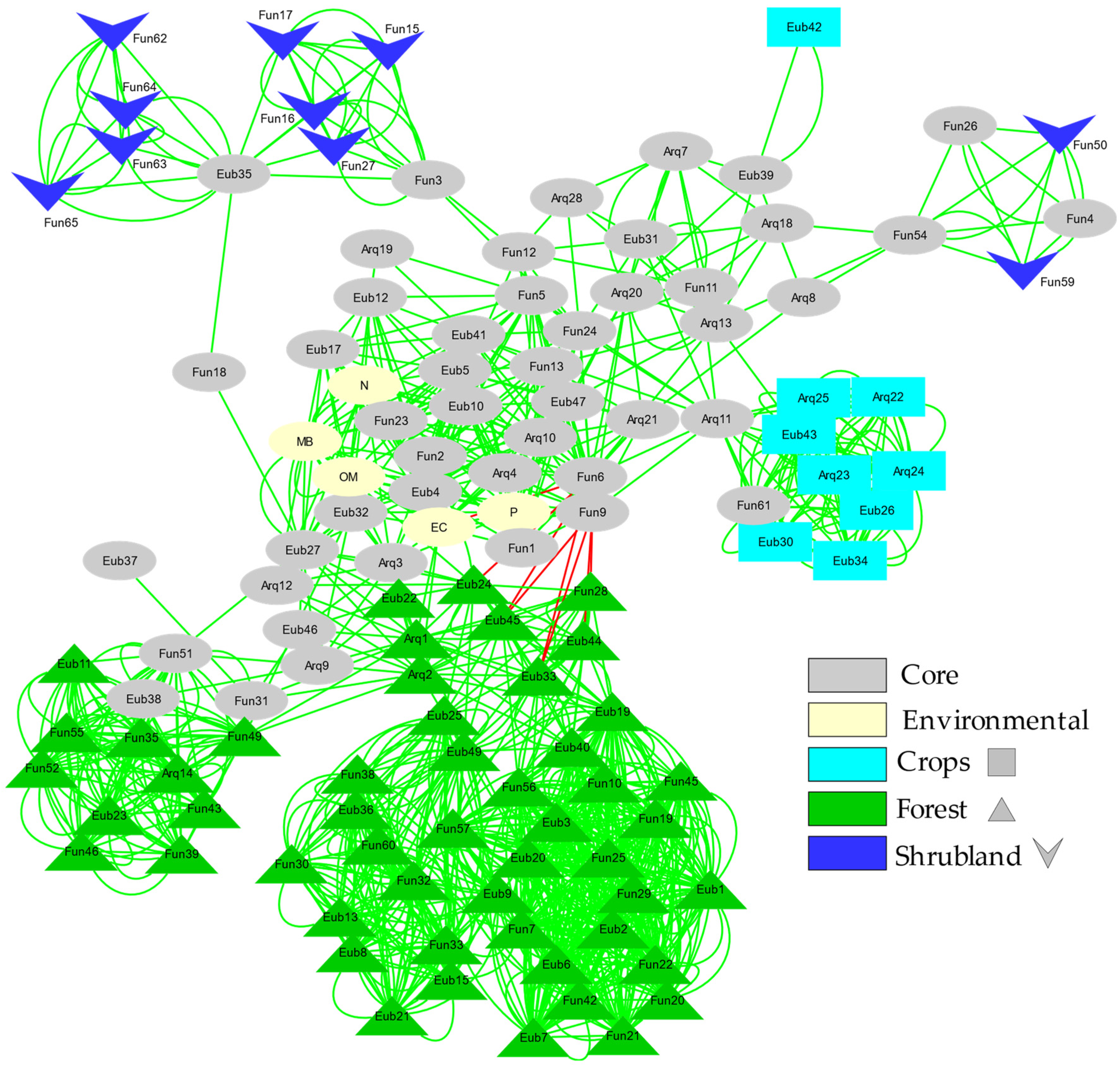
3.5. Microbial Network Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fitter, A.A.H.; Gilligan, C.A.; Hollinworth, K.; Kleczkowski, A.; Twyman, R.M.; Pitchford, J.W. Biodiversity and ecosystem functions in soil. Funct. Ecol. 2005, 19, 369–377. [Google Scholar] [CrossRef] [Green Version]
- Brussaard, L.; Kuyper, T.W.; Didden, W.A.M.; De Goede, R.G.M.; Bloem, J. Biological soil quality from biomass to biodiversity—Importance and resilience to management stress and disturbance. In Managing Soil Quality Challenges in Modern Agriculture; Schjønning, P., Elmholt, S., Christensen, B.T., Eds.; CAB International: Wallingford, CT, USA, 2004; pp. 139–161. [Google Scholar]
- Fischer, J.; Lindenmayer, D.B.; Manning, A.D. Biodiversity, ecosystem function, and resilience: Ten guiding principles for commodity production landscapes. Front. Ecol. Environ. 2006, 4, 80–86. [Google Scholar] [CrossRef] [Green Version]
- Pelfrêne, A.; Waterlot, C.; Douay, F. Influence of land use on human bioaccessibility of metals in smelter impacted soils. Environ. Pollut. 2013, 178, 80–88. [Google Scholar] [CrossRef]
- Santamans, A.C.; Boluda, R.; Picazo, A.; Gil, C.; Ramos-Miras, J.; Tejedo, P.; Pertierra, L.R.; Benayas, J.; Camacho, A. Soil features in rookeries of Antarctic penguins reveal sea to land biotransport of chemical pollutants. PLoS ONE 2017, 12, e0181901. [Google Scholar] [CrossRef] [Green Version]
- Gil, C.; Boluda, R.; Rodríguez Martín, J.A.; Guzmán, M.; del Moral, F.; Ramos-Miras, J.J. Assessing soil contamination and temporal trends of heavy metal contents in greenhouses on semiarid land. Land Degrad. Dev. 2018, 29, 3344–3354. [Google Scholar] [CrossRef]
- Ramos-Miras, J.J.; Gil, C.; Rodrıíguez Martın, J.A.; Bech, J.; Boluda, R. Ecological risk assessment of mercury and chromium in greenhouse soils. Environ. Geochem. Health 2020, 42, 313–324. [Google Scholar] [CrossRef]
- Iustman, L.J.R.; Amasqué, F.J.; Vullo, D.L. Microbiota Diversity Change as Quality Indicator of Soils Exposed to Intensive Periurban Agriculture. Curr. Microbiol. 2021, 78, 338–346. [Google Scholar] [CrossRef]
- Pizarro-Tobias, P.; Fernandez, M.; Niqui, J.L.; Solano, J.; Duque, E.; Ramos, J.L.; Roca, A. Restoration of a Mediterranean forest after a fire: Bioremediation and rhizoremediation field-scale trial. Microb. Biotechnol. 2015, 8, 77–92. [Google Scholar] [CrossRef]
- Rascio, I.; Curci, M.; Gattullo, C.E.; Lavecchia, A.; Khanghahi, M.Y.; Terzano, R.; Crecchio, C. Combined Effect of Laboratory-Simulated Fire and Chromium Pollution on Microbial Communities in an Agricultural. Soil Biol. 2021, 10, 587. [Google Scholar] [CrossRef]
- Hay, A. Chlorinated dioxins and the environment. Nature 1981, 289, 351–352. [Google Scholar] [CrossRef]
- Kirkok, S.K.; Kibet, J.K.; Kinyanjui, T.K.; Okanga, F.I. A review of persistent organic pollutants: Dioxins, furans, and their associated nitrogenated analogues. SN Appl. Sci. 2020, 2, 1729. [Google Scholar] [CrossRef]
- Mahfouz, S.; Mansour, G.; Murphy, D.J.; Hanano, A. Dioxin impacts on lipid metabolism of soil microbes: Towards effective detection and bioassessment strategies. Bioresour. Bioprocess. 2020, 7, 59. [Google Scholar] [CrossRef]
- Karlen, D.L.; Mausbach, M.J.; Doran, J.W.; Cline, R.G.; Harris, R.F.; Schuman, G.E. Soil quality: A concept, definition, and framework for evaluation. Soil Sci. Soc. Am. J. 1997, 61, 4–10. [Google Scholar] [CrossRef] [Green Version]
- Boluda, R.; Roca-Pérez, L.; Iranzo, M.; Gil, C.; Mormeneo, S. Determination of enzymatic activities using a miniaturized system as a rapid method to assess soil quality. Eur. J. Soil Sci. 2014, 65, 286–294. [Google Scholar] [CrossRef] [Green Version]
- Lehmann, J.; Bossio, D.A.; Kögel-Knabner, I.; Rillig, M.C. The concept and future prospects of soil health. Nat. Rev. Earth Environ. 2020, 1, 544–553. [Google Scholar] [CrossRef]
- Commision, E.U. The Implementation of the Soil Thematic Strategy and Ongoing Activities. 2012. Available online: https://ec.europa.eu/environment/soil/three_en.htm (accessed on 29 January 2021).
- Ekschmitt, K.; Griffiths, B.S. Soil biodiversity and its implications for ecosystems functioning in a heterogeneous and variable environment. Appl. Soil Ecol. 1998, 10, 201–215. [Google Scholar] [CrossRef]
- Breure, A.M. Soil biodiversity: Measurements, indicators, treats and soil functions. In Proceedings of the International Conference Soil and Compost Eco-Biology, Leon, Spain, 15–17 September 2004. [Google Scholar]
- Girvan, M.S.; Bullimore, J.; Pretty, J.N.; Osborn, A.M.; Ball, A.S. Soil type is the primary determinant of the composition of the total and active bacterial communities in arable soils. Appl. Environ. Microb. 2003, 69, 1800–1809. [Google Scholar] [CrossRef] [Green Version]
- Bossio, D.A.; Girvan, M.S.; Verchot, L.; Bullimore, J.; Borelli, T.; Albrecht, A.; Scow, K.M.; Ball, A.S.; Pretty, J.N.; Osborn, A.M. Soil microbial community response to land use change in an agricultural landscape of Western Kenya. Microb. Ecol. 2005, 49, 50–62. [Google Scholar] [CrossRef] [Green Version]
- Martin-Laurent, F.; Philippot, L.; Hallet, S.; Chaussod, R.; Germon, J.C.; Soulas, G.; Catroux, G. DNA extraction from soils: Old bias for new microbial diversity analysis methods. Appl. Environ. Microb. 2001, 67, 2354–2359. [Google Scholar] [CrossRef] [Green Version]
- Fierer, N.; Jackson, R.B. The diversity and biogeography of soil bacterial communities. Proc. Natl. Acad. Sci. USA 2006, 103, 626–631. [Google Scholar] [CrossRef] [Green Version]
- Christ, S.; Wubert, T.; Theuerl, S.; Herold, N.; Buscot, F. Fungal communities in bulk soil and stone compartments of different forest and soil types as revealed by a barcoding ITS rDNA and a functional laccase encoding gene marker. Soil Biol. Biochem. 2011, 43, 1292–1299. [Google Scholar] [CrossRef]
- Geml, J.; Timling, I.; Robinson, C.H.; Lennon, N.; Nusbaum, H.C.; Brochmann, C.; Noordeloos, M.E.; Taylor, D.L. An arctic community of symbiotic fungi assembled by long-distance dispersers: Phylogenetic diversity of ectomycorrhizal basidiomicetes in Svalbard based on soil and sporocarp DNA. J. Biogeogr. 2012, 39, 74–88. [Google Scholar] [CrossRef]
- De los Reyes, A.M.; Ocampo, E.T.; Manuel, M.C.; Mendoza, B.C. Analysis of the bacterial and fungal community profiles in bulk soil and rhizospheres of three mungbean [Vigna radiata (L.) R. Wilczek] genotypes through PCR-DGGE. Int Lett Nat. Sci. 2020, 77, 1–26. [Google Scholar] [CrossRef]
- Nakatsu, C.H. Soil microbial community analysis using denaturing gradient gel electrophoresis. Soil Sci. Soc. Am. J. 2007, 71, 562–571. [Google Scholar] [CrossRef] [Green Version]
- Gabriel, J. Development of soil microbiology methods: From respirometry to molecular approaches. J. Ind. Microbiol. Biot. 2010, 37, 1289–1297. [Google Scholar] [CrossRef]
- Seybold, C.A.; Mansbach, M.J.; Karlen, D.L.; Rogers, H.H. Quantification of soil quality. In Soil Processes and the Carbon Cycle; CRC Press: Boca Raton, FL, USA, 2018; pp. 387–404. [Google Scholar]
- Rubio, J.L.; Bochet, E. Desertification indicators as diagnosis criteria for desertification risk assessment in Europe. J. Arid Environ. 1998, 39, 113–120. [Google Scholar] [CrossRef]
- Recatalá, L.; Ive, J.R.; Baird, I.A.; Hamilton, N.; Sánchez, J. Land-use planning in the Valencian Mediterranean Region: Using LUPIS to generate issue relevant plans. J. Environ. Manag. 2000, 59, 169–184. [Google Scholar] [CrossRef]
- Gamón, M.; Sáez, E.; Gil, J.; Boluda, R. Direct and indirect exogenous contamination by pesticides of rice-farming soils in a Mediterranean wetland. Arch. Environ. Contam. Toxicol. 2003, 44, 0141–0151. [Google Scholar] [CrossRef]
- Gil, C.; Boluda, R.; Ramos, J. Determination and evaluation of cadmium, lead and nickel in greenhouse soils of Almería (Spain). Chemosphere 2004, 55, 1027–1034. [Google Scholar] [CrossRef]
- Gil, C.; Ramos-Miras, J.; Roca-Pérez, L.; Boluda, R. Determination and assessment of mercury content in calcareous soils. Chemosphere 2010, 78, 409–415. [Google Scholar] [CrossRef]
- Roca-Perez, L.; Gil, C.; Cervera, M.L.; Gonzálvez, A.; Ramos-Miras, J.; Pons, V.; Bech, L.; Boluda, R. Selenium and heavy metals content in some Mediterranean soils. J. Geochem. Explor. 2010, 107, 110–116. [Google Scholar] [CrossRef]
- Ramos-Miras, J.J.; Roca-Perez, L.; Guzmán-Palomino, M.; Boluda, R.; Gil, C. Background levels and baseline values of available heavy metals in Mediterranean greenhouse soils (Spain). J. Geochem. Explor. 2011, 110, 186–192. [Google Scholar] [CrossRef]
- Cañizares, R.; Moreno, B.; Benitez, E. Biochemical characterization with detection and expression of bacterial β-glucosidase encoding genes of a Mediterranean soil under different long-term management practices. Biol. Fert. Soils 2012, 48, 651–663. [Google Scholar] [CrossRef]
- Yang, T.; Lupwayi, N.; Marc, S.A.; Siddique, K.H.; Bainard, L.D. Anthropogenic drivers of soil microbial communities and impacts on soil biological functions in agroecosystems. Glob. Ecol. Conserv. 2021, 27, e01521. [Google Scholar] [CrossRef]
- Boluda, R.; Andreu, V.; Moraleda, M.; Sánchez, J. Factores ecológicos (geología vegetación y clima) de la comarca La Plana de Requena-Utiel (Valencia): I Relaciones suelo-material de origen. An. Edaf. Agrobiol. 1988, 47, 887–917. [Google Scholar]
- IUSS Working Group WRB. World Reference Base for Soil Resources 2014, update 2015 International Soil Classify-Cation System for Naming Soils and Creating Legends for Soil Maps; World Soil Resources Reports No. 106; FAO: Rome, Italy, 2015; Available online: https://www.fao.org/3/i3794en/I3794en.pdf (accessed on 29 January 2021).
- ISO 11464. Soil Quality—Pretreatment of Samples for Physic-Chemical Analysis; ISO: Geneva, Switzerland, 1994. [Google Scholar]
- ISO 10390. Soil Quality—Determination of PH; ISO: Geneva, Switzerland, 1994. [Google Scholar]
- ISO 11265. Soil Quality—Determination Specific Electrical Conductivity; ISO: Geneva, Switzerland, 1994. [Google Scholar]
- ISO 10693. Soil Quality—Determination of Carbonate Content. Volumetric Method; ISO: Geneva, Switzerland, 1995. [Google Scholar]
- ISO 11272. Soil Quality—Determination of Dry Bulk Density; ISO: Geneva, Switzerland, 1998. [Google Scholar]
- Richards, L.A. Diagnosis and Improvement of Saline and Alkaline Soils; USDA: Washington, DC, USA, 1954. [Google Scholar]
- Day, P.R. Particle fractionation and particle-size analysis. In Methods of Soil Analysis: Part 1 Physical and Mineralogical Properties, Including Statistics of Measurement and Sampling; Black, C.A., Ed.; American society of Agronomy Inc.: Madison, WI, USA, 1965; pp. 545–567. [Google Scholar]
- ISO 10694. Soil Quality—Determination of Organic and Total Carbon after Dry Combustion (Elementary Analysis); ISO: Geneva, Switzerland, 1995. [Google Scholar]
- ISO 13878. Soil Quality—Determination of Total Nitrogen Content by Dry Combustion (“Elemental Analysis”); ISO: Geneva, Switzerland, 1998. [Google Scholar]
- ISO 15178. Soil Quality—Determination of Total Sulfur by Dry Combustion; ISO: Geneva, Switzerland, 2000. [Google Scholar]
- ISO 11260. Soil Quality—Determination of Effective Cation Exchange Capacity and Base Saturation Level Using Barium Chloride Solution; ISO: Geneva, Switzerland, 1994. [Google Scholar]
- Walkley, A.; Black, I.A. An examination of Degtjareff method. For determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- ISO 14240. Soil Quality—Determination of Soil Microbial Biomass—Part 1: Substrate-Induced Respiration Method; ISO: Geneva, Switzerland, 1997. [Google Scholar]
- ISO 11263. Soil Quality—Determination of Phosphorus. Spectrometric Determination of Phosphorus Soluble in Sodium Hydrogen Carbonate Solution; ISO: Geneva, Switzerland, 1994. [Google Scholar]
- Dar, S.A.; Kuenen, J.G.; Muyzer, G. Nested PCR-denaturing gradient gel electrophoresis approach to determine the diversity of sulfate-reducing bacteria in complex microbial communities. Appl. Environ. Microbiol. 2005, 71, 2325–2330. [Google Scholar] [CrossRef] [Green Version]
- Benlloch, S.; López-López, A.; Casamayor, E.O.; Øvreås, L.; Goddard, V.; Daae, F.L.; Smerdon, G.; Massana, R.; Joint, I.; Thingstad, F.; et al. Prokaryotic genetic diversity throughout the salinity gradient of a coastal solar saltern. Environ. Microbiol. 2002, 4, 349–360. [Google Scholar] [CrossRef]
- Landeweert, R.; Leeflang, P.; Smit, E.; Kuyper, T. Diversity of an ectomycorrhizal fungal community studied by a root tip and total soil DNA approach. Mycorrhiza 2005, 15, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Chan, O.C.; Wolf, M.; Hepperle, D.; Casper, P. Methanogenic archaeal community in the sediment of an artificially partitioned acidic bog lake. FEMS Microbiol. Ecol. 2002, 42, 119–129. [Google Scholar] [CrossRef]
- Hill, M.O. Diversity and evenness: A unifying notation and its consequences. Ecology 1973, 54, 427–432. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.; Rodrigues, J.L.; Ng, J.P.; Gentry, T.J. Hill number as a bacterial diversity measure framework with high-throughput sequence data. Sci. Rep. 2016, 6, 38263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jost, L. The relation between evenness and diversity. Diversity 2010, 2, 207–232. [Google Scholar] [CrossRef]
- Legendre, P.; Anderson, M.J. Distance-based redundancy analysis: Testing multispecies responses in multifactorial ecological experiments. Ecol. Monogr. 1999, 69, 1–24. [Google Scholar] [CrossRef]
- The R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015. [Google Scholar]
- Faust, K.; Raes, J. CoNet app: Inference of biological association networks using Cytoscape. F1000Research 2016, 5, 1519. [Google Scholar] [CrossRef]
- Lopes, C.T.; Franz, M.; Kazi, F.; Donaldson, S.L.; Morris, Q.; Bader, G.D. Cytoscape Web: An interactive web-based network browser. Bioinformatics 2010, 26, 2347–2348. [Google Scholar] [CrossRef] [Green Version]
- Sahani, U.; Behera, N. Impact of deforestation on soil physicochemical characteristics, microbial biomass and microbial activity of tropical soil. Land Degrad. Dev. 2001, 12, 93–105. [Google Scholar] [CrossRef]
- Tinoco, P.; Piedra Buena, A.; Zancada, M.; Sanz, J.; Almendros, G. Biogeochemical proxies of anthropic impact in Mediterranean forest soils. Soil Use Manag. 2010, 26, 320–331. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, J.; González-Pérez, J.A.; Turmero, A.; Hernández, M.; Ball, A.S.; González-Vila, F.J.; Arias, M.E. Wildfire effects on the microbial activity and diversity in a Mediterranean forest soil. Catena 2017, 158, 82–88. [Google Scholar] [CrossRef] [Green Version]
- Certini, G.; Moya, D.; Lucas-Borja, M.E.; Mastrolonardo, G. The impact of fire on soil-dwelling biota: A review. For. Ecol. Manag. 2021, 488, 118989. [Google Scholar] [CrossRef]
- Prashar, P.; Shah, S. Impact of fertilizers and pesticides on soil microflora in agriculture. In Sustainable Agriculture Reviews; Springer: Cham, Switzerland, 2016; pp. 331–361. [Google Scholar] [CrossRef]
- García-Orenes, F.; Morugán-Coronado, A.; Zornoza, R.; Scow, K. Changes in soil microbial community structure influenced by agricultural management practices in a Mediterranean agro-ecosystem. PLoS ONE 2013, 8, e80522. [Google Scholar] [CrossRef]
- Lutzow, M.V.; Kogel-Knabner, I.; Ekschmitt, K.; Matzner, E.; Guggenberger, G.; Marschner, B.; Flessa, H. Stabilization of organic matter in temperate soils: Mechanisms and their relevance under different soil conditions—A review. Eur. J. Soil Sci. 2006, 57, 426–445. [Google Scholar] [CrossRef]
- Rovira, P.; Jorba, M.; Romanyà, J. Active and passive organic matter fractions in Mediterranean forest soils. Biol. Fert. Soils 2010, 46, 355–369. [Google Scholar] [CrossRef]
- Barrow, C.J. Land Degradation: Development and Breakdown of Terrestrial Environments; Cambridge University Press: New York, NY, USA, 1991. [Google Scholar]
- Lal, R.; Stewart, B.A. Land Degradation. Advances in Soil Science; Springer: New York, NY, USA, 1994; Volume 11. [Google Scholar]
- Clark, M.; Horwath, W.; Shennan, C. Changes in soil chemical properties resulting from organic and low-input farming practices. Agron. J. 1998, 90, 662–671. [Google Scholar] [CrossRef] [Green Version]
- Kalbitz, K.; Schwesig, D.; Schmerwitz, J.; Kaiser, K.; Haumaier, L.; Glaser, B.; Ellerbrock, R.; Leinwebere, P. Changes in properties of soil-derived dissolved organic matter induced by biodegradation. Soil Biol. Biochem. 2003, 35, 1129–1142. [Google Scholar] [CrossRef]
- Trasar-Cepeda, C.; Leirós, M.C.; Gil-Sotres, F. Hydrolytic enzyme activities in agricultural and forest soils. Some implications for their use as indicators of soil quality. Soil Biol. Biochem. 2008, 40, 2146–2155. [Google Scholar] [CrossRef] [Green Version]
- Kurganova, I.N.; Lopes de Gerenyu, V.O.; Gallardo-Lancho, J.F.; Oehm, C.T. Evaluation of the rates of soil organic matter mineralization in forest ecosystems of temperate continental, Mediterranean, and tropical monsoon climates. Eurasian. Soil Sci. 2012, 45, 68–79. [Google Scholar] [CrossRef]
- Casamayor, E.O.; Pedrós-Alió, C.; Muyzer, G.; Amann, R. Microheterogeneity in 16S ribosomal DNA-defined bacterial populations from a stratified planktonic environment is related to temporal changes and to ecological adaptations. Appl. Environ. Microb. 2002, 68, 1706–1714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenny, H. Factors of Soil Formation: A System of Quantitative Pedology; Dover Press: New York, NY, USA, 1994. [Google Scholar]
- Kennedy, A.C.; Gewin, V.L. Soil microbial diversity: Present and future considerations. Soil Sci. 1997, 162, 607–617. [Google Scholar] [CrossRef]
- Müller, A.K.; Westergaard, K.; Christensen, S.; Sørensen, S.J. The diversity and function of soil microbial communities exposed to different disturbances. Microb. Ecol. 2002, 44, 49–58. [Google Scholar] [CrossRef]
- Nannipieri, P.; Ascher, J.; Ceccherini, M.T.; Landi, L.; Pietramellara, G.; Renella, G. Microbial diversity and soil functions. Eur. J. Soil Sci. 2003, 54, 655–670. [Google Scholar] [CrossRef]
- VanInsberghe, D.; Hartmann, M.; Stewart, G.R.; Mohn, W.W. Isolation of a substantial proportion of forest soil bacterial communities detected via pyrotagsequencing. Appl. Environ. Microb. 2013, 79, 2096–2098. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Code | Soil Use | Bioclimatic Belt | Parent Material | Vegetation | Soil Horizon | Soil Type (FAO) |
|---|---|---|---|---|---|---|
| C1 | Forest | Supra-Mediterranean | Limestone | Holm Oakwood | Mollic (Ah) | mollic Leptosol |
| C2 | Shrubland | Supra-Mediterranean | Limestone | Shrubland | Ochric (Ah) | skeletic Leptosol |
| C3 | Crops | Meso-Mediterranean | Limestone | Cereals | Anthropic (Ap) | skeletic Leptosol |
| D1 | Forest | Supra-Mediterranean | Dolomites | Holm Oakwood | Mollic (Ah) | mollic Leptosol |
| D2 | Shrubland | Supra-Mediterranean | Dolomites | Shrubland | Ochric (Ah) | dolomitic Leptosol |
| D3 | Crops | Supra-Mediterranean | Dolomites | Cereals | Anthropic (Ap) | dolomitic Leptosol |
| T1 | Forest | Meso-Mediterranean | Marl and limestone | Holm Oakwood | Mollic (Ah) | rendzic Leptosol (*) |
| T2 | Shrubland | Meso-Mediterranean | Marl and limestone | Shrubland | Ochric (Ah) | calcaric Regosol |
| T3 | Crops | Meso-Mediterranean | Marl and limestone | Almonds and vineyards | Anthropic (Ap) | calcaric Regosol |
| C1 | C2 | C3 | D1 | D2 | D3 | T1 | T2 | T3 | |
|---|---|---|---|---|---|---|---|---|---|
| Sand % | 27 ± 7.16 | 17.75 ± 0.96 | 25.25 ± 2.06 | 24.75 ± 1.26 | 13.5 ± 3.51 | 19.75 ± 2.36 | 30.25 ± 2.36 | 39.25 ± 6.18 | 28.5 ± 0.58 |
| Silt % | 35.75 ± 2.36 | 41.5 ± 9.85 | 41.75 ± 3.77 | 32.5 ± 6.14 | 43.5 ± 5.26 | 42.25 ± 2.87 | 25.75 ± 6.65 | 25.25 ± 4.92 | 28.5 ± 3.51 |
| Clay % | 37.25 ± 5.56 | 40.75 ± 10.24 | 32.5 ± 1.29 | 43 ± 5.23 | 42.75 ± 3.10 | 37.75 ± 0.50 | 44 ± 4.69 | 35 ± 2.94 | 43 ± 3.56 |
| Bulk density (g cm−3) | 0.44 ± 0.03 | 0.81 ± 0.03 | 1.28 ± 0.11 | 0.47 ± 0.04 | 1.05 ± 0.05 | 1.23 ± 0.02 | 0.68 ± 0.06 | 0.94 ± 0.10 | 1.17 ± 0.16 |
| WHC % | 21.85 ± 1.43 | 12.63 ± 1.85 | 8.78 ± 1.10 | 27.6 ± 1.23 | 13.2 ± 1.16 | 8.98 ± 0.56 | 19.23 ± 2.42 | 10.78 ± 2.10 | 10.33 ± 0.76 |
| EC (dS m−1) | 0.25 ± 0.01 | 0.13 ± 0.02 | 0.15 ± 0.00 | 0.24 ± 0.02 | 0.11 ± 0.00 | 0.11 ± 0.00 | 0.24 ± 0.02 | 0.12 ± 0.01 | 0.14 ± 0.02 |
| pH | 7.54 ± 0.29 | 8.17 ± 0.10 | 8.41 ± 0.02 | 7.35 ± 0.21 | 8.07 ± 0.14 | 8.45 ± 0.04 | 7.70 ± 0.20 | 8.39 ± 0.12 | 8.34 ± 0.05 |
| CaCO3 (g kg−1) | 63.5 ± 5.76 | 42.25 ± 30.39 | 419.5 ± 33.51 | 20.75 ± 5.19 | 45.25 ± 15.9 | 446 ± 17.26 | 90 ± 9.63 | 191.75 ± 35.27 | 121 ± 57.95 |
| C % | 12.09 ± 1.6 | 5.91 ± 2.26 | 6.01 ± 0.14 | 13.27 ± 1.39 | 5.09 ± 0.58 | 6.49 ± 0.04 | 9.77 ± 2.45 | 3.71 ± 0.13 | 2.85 ± 0.41 |
| S % | 0.01 ± 0.00 | 0.02 ± 0.01 | 0.02 ± 0.00 | 0.03 ± 0.01 | 0.03 ± 0.00 | 0.02 ± 0.00 | 0.01 ± 0.00 | 0.01 ± 0.00 | 0.01 ± 0.00 |
| CEC (cmol kg−1) | 45.38 ± 1.36 | 34.25 ± 3.30 | 13.58 ± 1.61 | 39.68 ± 2.71 | 35.78 ± 1.31 | 23.8 ± 0.49 | 38.65 ± 1.39 | 23.88 ± 2.22 | 22.13 ± 1.68 |
| ExCa (cmol kg−1) | 37.13 ± 1.78 | 30.5 ± 3.47 | 12.18 ± 1.68 | 34.57 ± 3.06 | 32.95 ± 0.98 | 22.58 ± 0.4 | 35.71 ± 1.33 | 22.46 ± 2.15 | 19.5 ± 2.22 |
| ExMg (cmol kg−1) | 7.99 ± 0.52 | 2.65 ± 1.62 | 0.57 ± 0.05 | 3.83 ± 0.71 | 1.73 ± 0.68 | 0.48 ± 0.04 | 1.90 ± 0.36 | 0.64 ± 0.06 | 0.83 ± 0.17 |
| ExK (cmol kg−1) | 0.72 ± 0.02 | 1.04 ± 0.46 | 0.84 ± 0.05 | 1.18 ± 0.32 | 1.05 ± 0.36 | 0.64 ± 0.08 | 0.98 ± 0.12 | 0.69 ± 0.04 | 1.4 ± 0.14 |
| ExNa (cmol kg−1) | 0.08 ± 0.01 | 0.07 ± 0.02 | 0.02 ± 0.02 | 0.1 ± 0.03 | 0.05 ± 0.01 | 0.09 ± 0.04 | 0.07 ± 0.03 | 0.09 ± 0.01 | 0.06 ± 0.02 |
| Available P2O5 (mg 100g−1) | 2.16 ± 1.15 | 0.99 ± 0.21 | 1.73 ± 0.46 | 2.37 ± 1.10 | 0.74 ± 0.22 | 1.24 ± 0.22 | 0.94 ± 0.29 | 1.14 ± 0.27 | 3.77 ± 1.61 |
| N % | 0.7 ± 0.07 | 0.46 ± 0.12 | 0.18 ± 0.02 | 0.85 ± 0.13 | 0.44 ± 0.05 | 0.21 ± 0.01 | 0.49 ± 0.05 | 0.19 ± 0.03 | 0.19 ± 0.03 |
| SOM % | 16.9 ± 1.63 | 8.45 ± 1.53 | 2.03 ± 0.13 | 20.75 ± 0.70 | 7.13 ± 0.87 | 2.25 ± 0.13 | 12.35 ± 2.64 | 3.53 ± 0.78 | 2.83 ± 0.34 |
| MB (C, mg kg−1) | 2419 ± 311 | 1374 ± 234 | 534 ± 75 | 2080 ± 405 | 838 ± 194 | 446 ± 49 | 1407 ± 315 | 494 ± 80 | 399 ± 220 |
| Parameters | Soil Use ab | Soil Type (FAO) b | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sand % | Fo-Sh-Cr (25.11) | C-R (32.67) | > | R-M (27.33) | > | M-Hc (24.19) | > | Hc-Hs (19.06) | ||||
| Silt % | Cr-Sh (37.13) | > | Sh-Fo (34.04) | Hc-Hs-M (39.54) | > | M-R-C (29.55) | ||||||
| Clay % | Fo-Sh-Cr (39.56) | M-R-Hs-Hc-C (39.56) | ||||||||||
| Bulk density (g cm−3) | 3 (1.22) | > | Sh (0.93) | > | Fo (0.53) | Hc-C (1.15) | > | C-Hs (0.99) | > | R (0.68) | > | M (0.45) |
| WHC % | Fo (22.89) | > | Sh (12.20) | > | Cr (9.36) | M (24.73) | > | R (19.23) | > | Hs-C (11.73) | > | C-Hc (9.71) |
| EC (dS m−1) | Fo (0.24) | > | Sh-Cr (0.13) | M-R (0.24) | > | C-Hs-Hc (0.13) | ||||||
| pH | Cr (8.40) | > | Sh (8.21) | > | Fo (7.53) | Hc-C (8.39) | > | C-Hs (8.24) | > | R (7.70) | > | M (7.44) |
| CaCO3 (g kg−1) | Cr (328.83) | > | Fo-Sh (75.58) | Hc (432.75) | > | C (156.38) | > | R-M-Hs (52.35) | ||||
| C % | Fo (11.71) | > | Sh -3 (5.01) | M (12.68) | > | R (9.77) | > | Hs-Hc (5.87) | > | Hc-C (4.39) | ||
| S % | Fo-Sh-Cr (0.02) | M-R-Hs-Hc-C (0.02) | ||||||||||
| CEC (cmol kg−1) | Fo (41.23) | > | Sh (31.30) | > | Cr (19.83) | M-R (41.23) | > | R-Hs (36.23) | > | C-Hc (20.84) | ||
| ExCa (cmol kg−1) | Fo (35.80) | > | Sh (28.64) | > | Cr (18.09) | M-R-Hs (34.17) | > | C-Hc (19.18) | ||||
| ExMg (cmol kg−1) | Fo (4.57) | > | Sh (1.67) | > | Cr (0.63) | M (5.91) | > | R-Hs-Hc-C (1.26) | ||||
| ExK (cmol kg−1) | Fo-Sh-Cr (0.95) | M-R-Hs-Hc-C (0.95) | ||||||||||
| ExNa (cmol kg−1) | Fo-Sh-Cr (0.07) | M-R-Hs-Hc-C (0.07) | ||||||||||
| P2O5 (mg 100g−1) | Fo-Cr (2.03) | > | Sh (0.96) | C-M-Hc (2.07) | > | M-Hc-R (1.69) | > | R-Hc-Hs (1.13) | ||||
| N % | Fo (0.68) | > | Sh (0.36) | > | Cr (0.19) | M (0.77) | > | R-Hs (0.46) | > | C-Hc (0.19) | ||
| SOM % | Fo (16.66) | > | Sh (6.37) | > | Cr (2.37) | M (18.83) | > | R (12.35) | > | Hs (7.79) | > | C-Hc (2.66) |
| MB (C mg kg−1) | Fo (1968) | > | Sh (902) | > | Cr (460) | M (2250) | > | R-Hs (1206) | > | C-Hc (468) | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Camacho, A.; Mora, C.; Picazo, A.; Rochera, C.; Camacho-Santamans, A.; Morant, D.; Roca-Pérez, L.; Ramos-Miras, J.J.; Rodríguez-Martín, J.A.; Boluda, R. Effects of Soil Quality on the Microbial Community Structure of Poorly Evolved Mediterranean Soils. Toxics 2022, 10, 14. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics10010014
Camacho A, Mora C, Picazo A, Rochera C, Camacho-Santamans A, Morant D, Roca-Pérez L, Ramos-Miras JJ, Rodríguez-Martín JA, Boluda R. Effects of Soil Quality on the Microbial Community Structure of Poorly Evolved Mediterranean Soils. Toxics. 2022; 10(1):14. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics10010014
Chicago/Turabian StyleCamacho, Antonio, César Mora, Antonio Picazo, Carlos Rochera, Alba Camacho-Santamans, Daniel Morant, Luis Roca-Pérez, José Joaquín Ramos-Miras, José A. Rodríguez-Martín, and Rafael Boluda. 2022. "Effects of Soil Quality on the Microbial Community Structure of Poorly Evolved Mediterranean Soils" Toxics 10, no. 1: 14. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics10010014