
Two Hits of EDCs Three Generations Apart: Effects on Social Behaviors in Rats, and Analysis by Machine Learning


Abstract
:1. Introduction
2. Materials and Methods
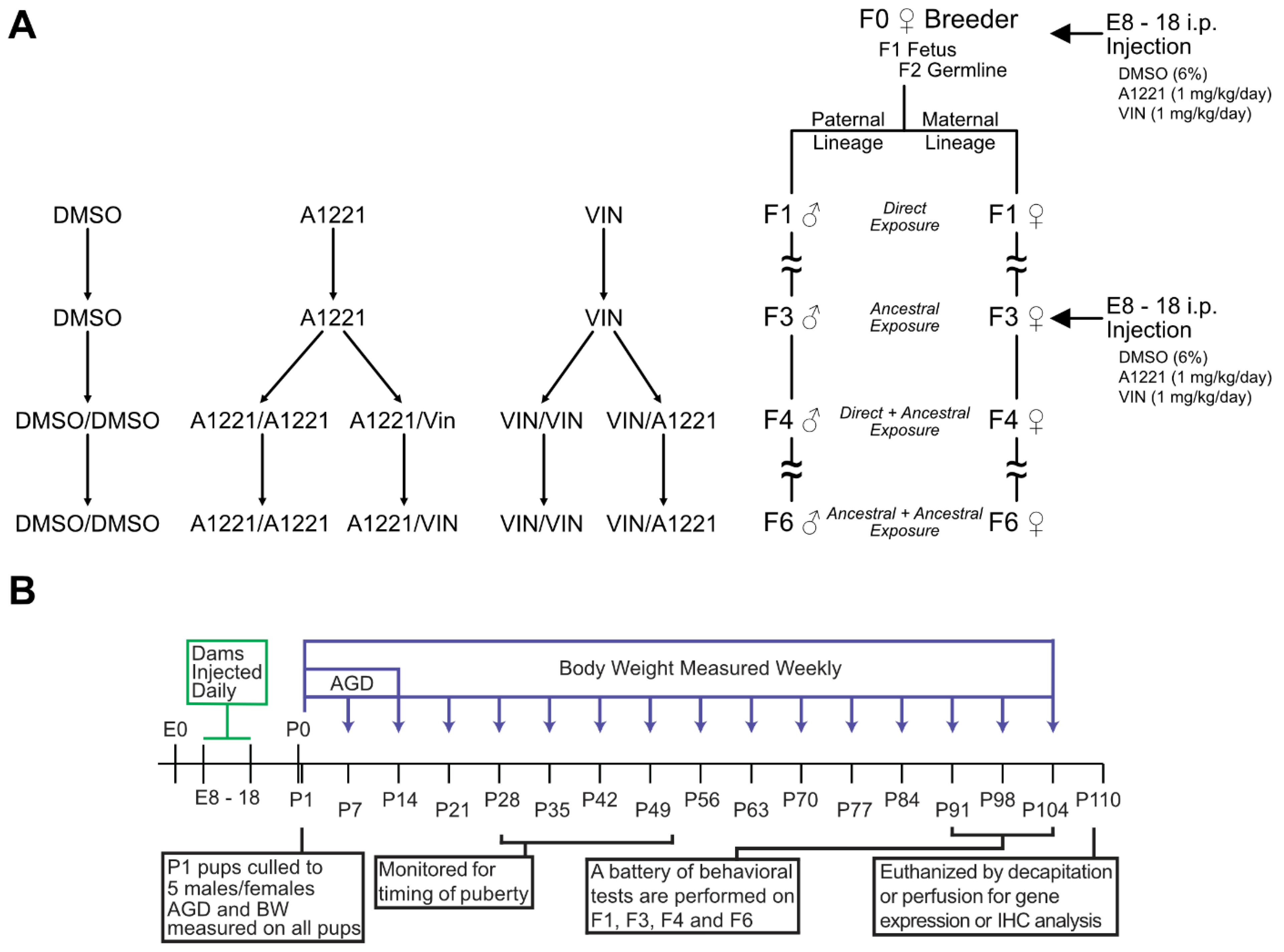
2.1. Animals and Treatment
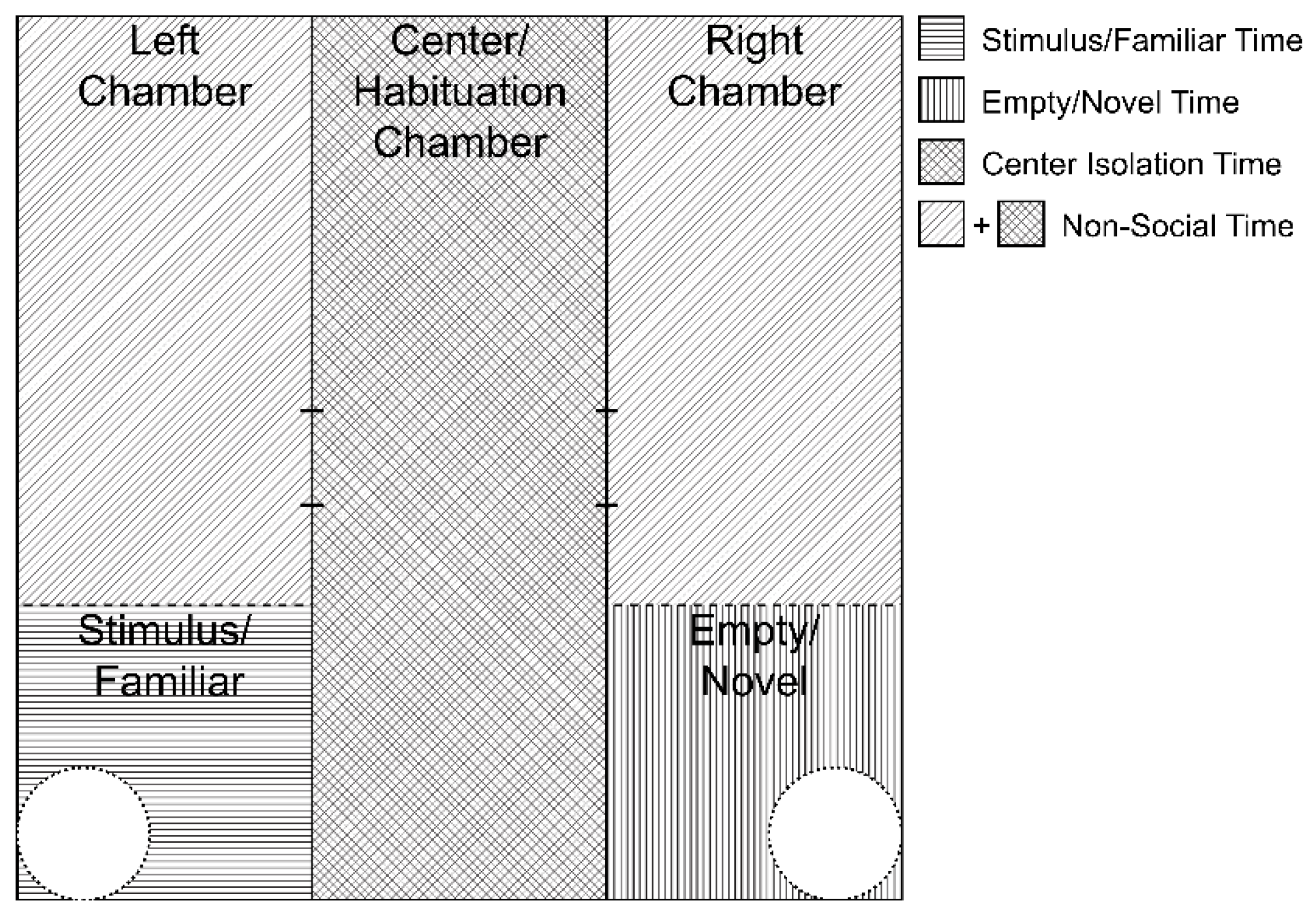
2.2. Sociability Apparatus and Behavioral Analysis
2.3. Data Exclusion Criteria
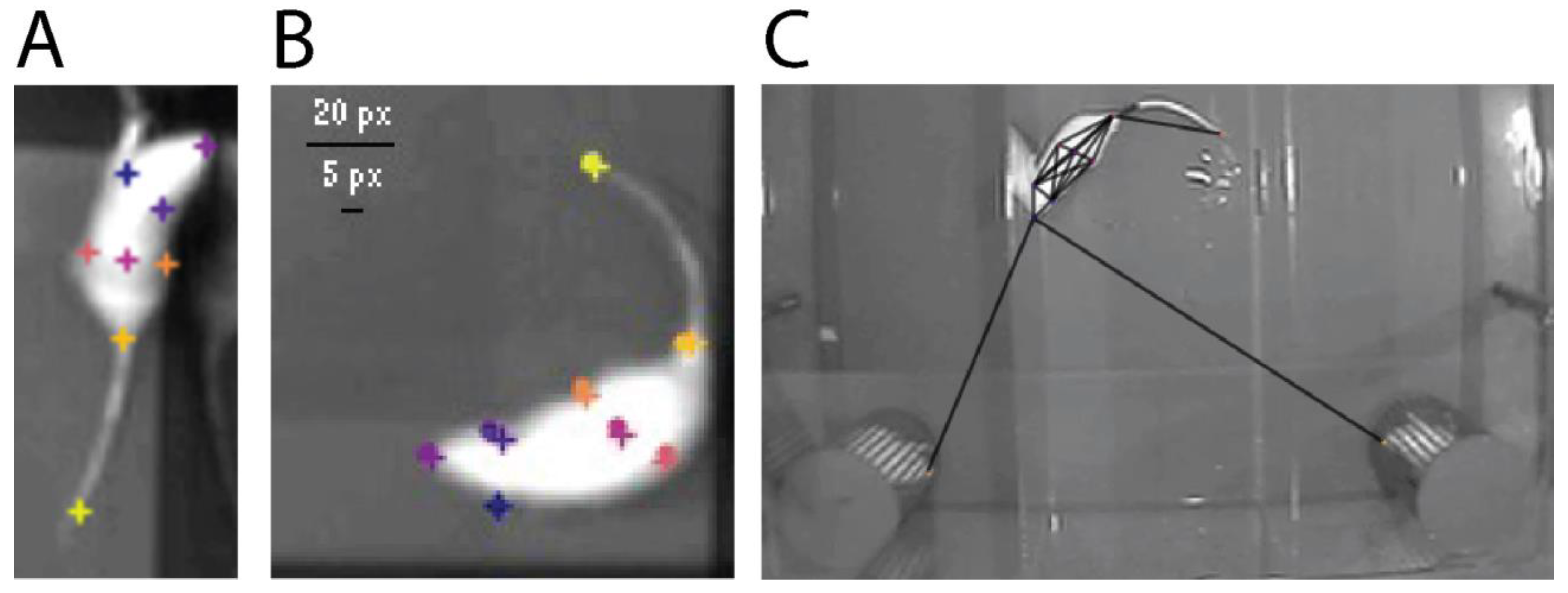
2.4. Nose Touch Detection with Machine Learning and DeepLabCut
2.5. Statistics
3. Results
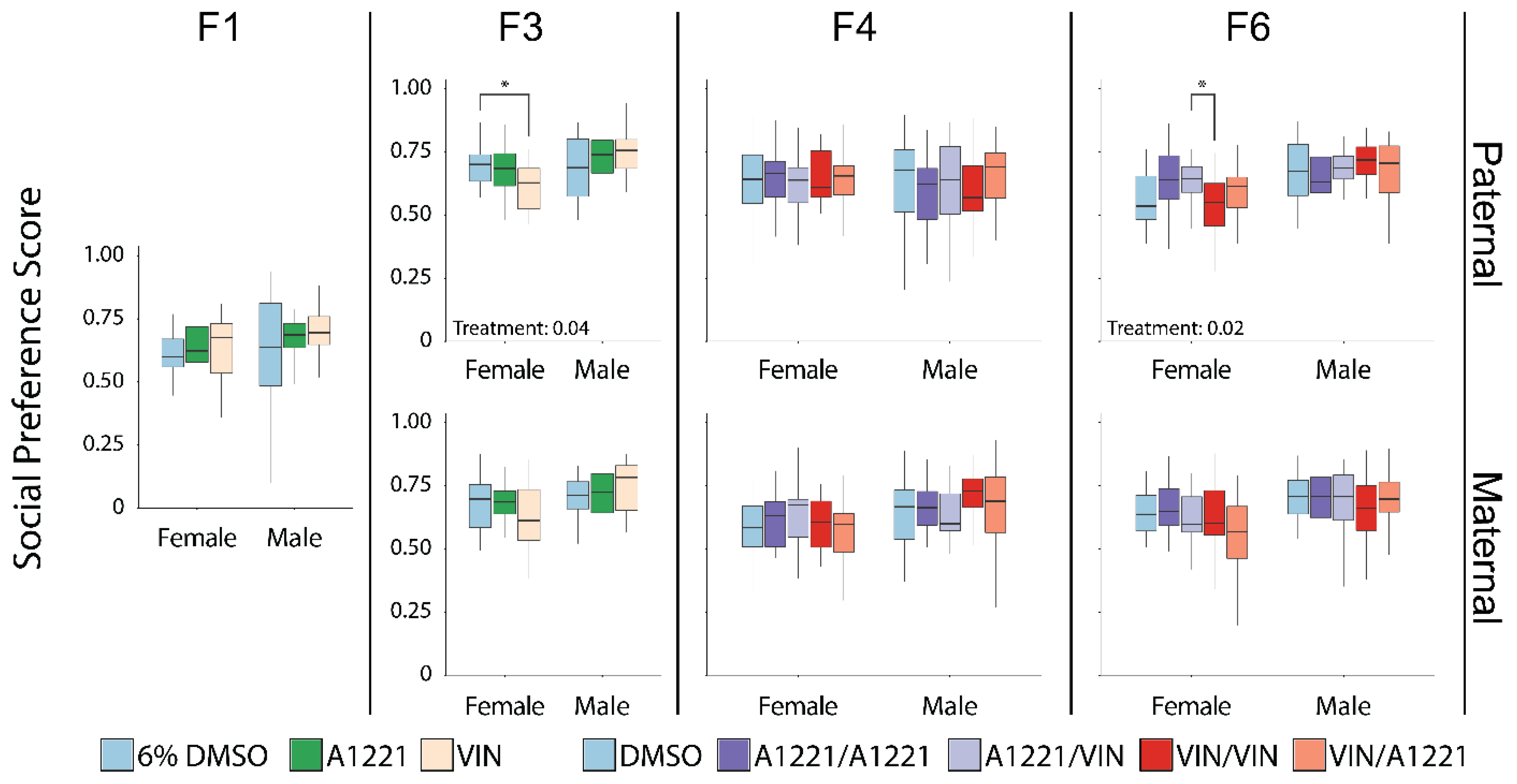
3.1. Sociability
3.1.1. Sex Differences
3.1.2. Effect of EDC Exposure
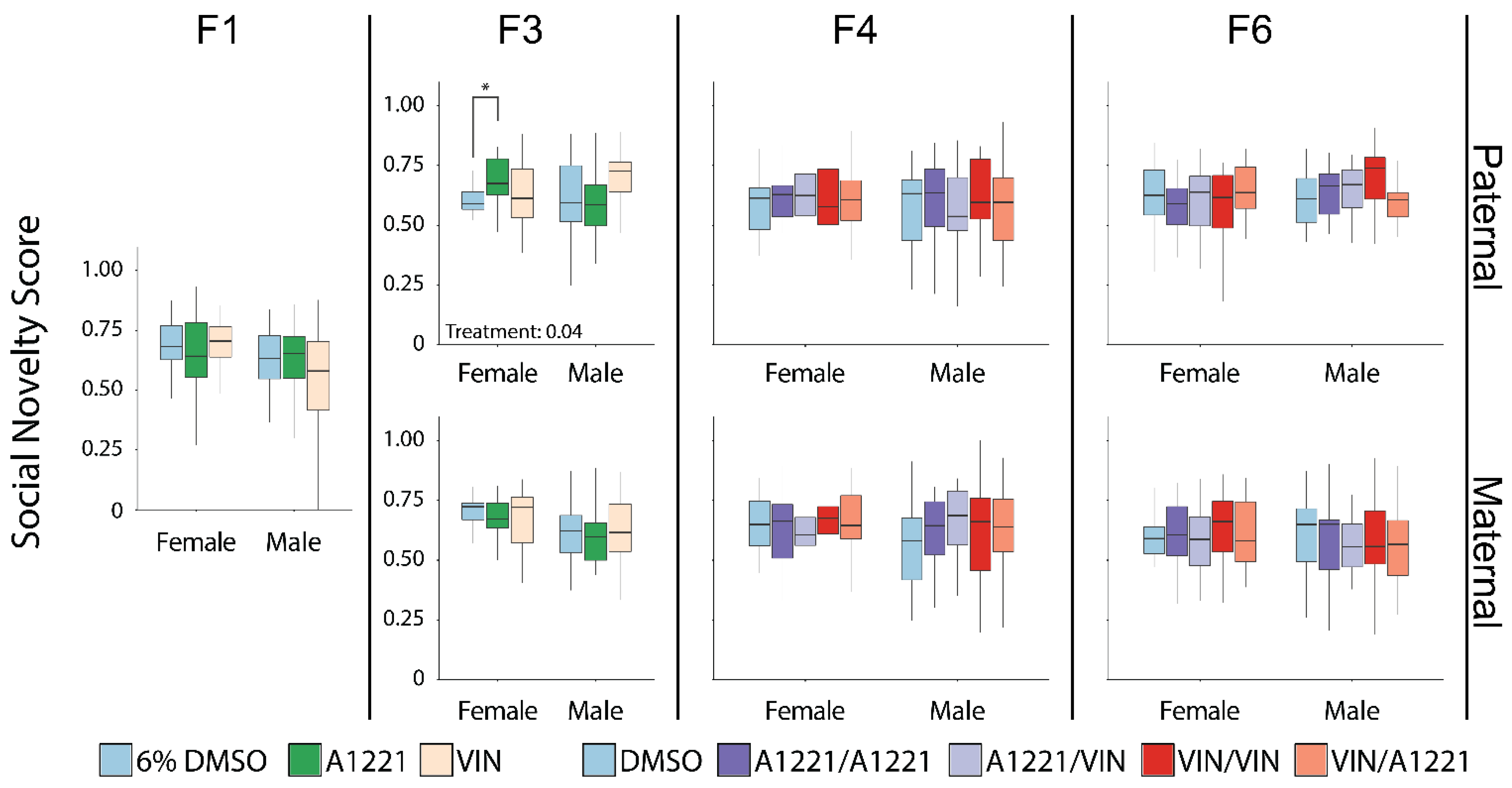
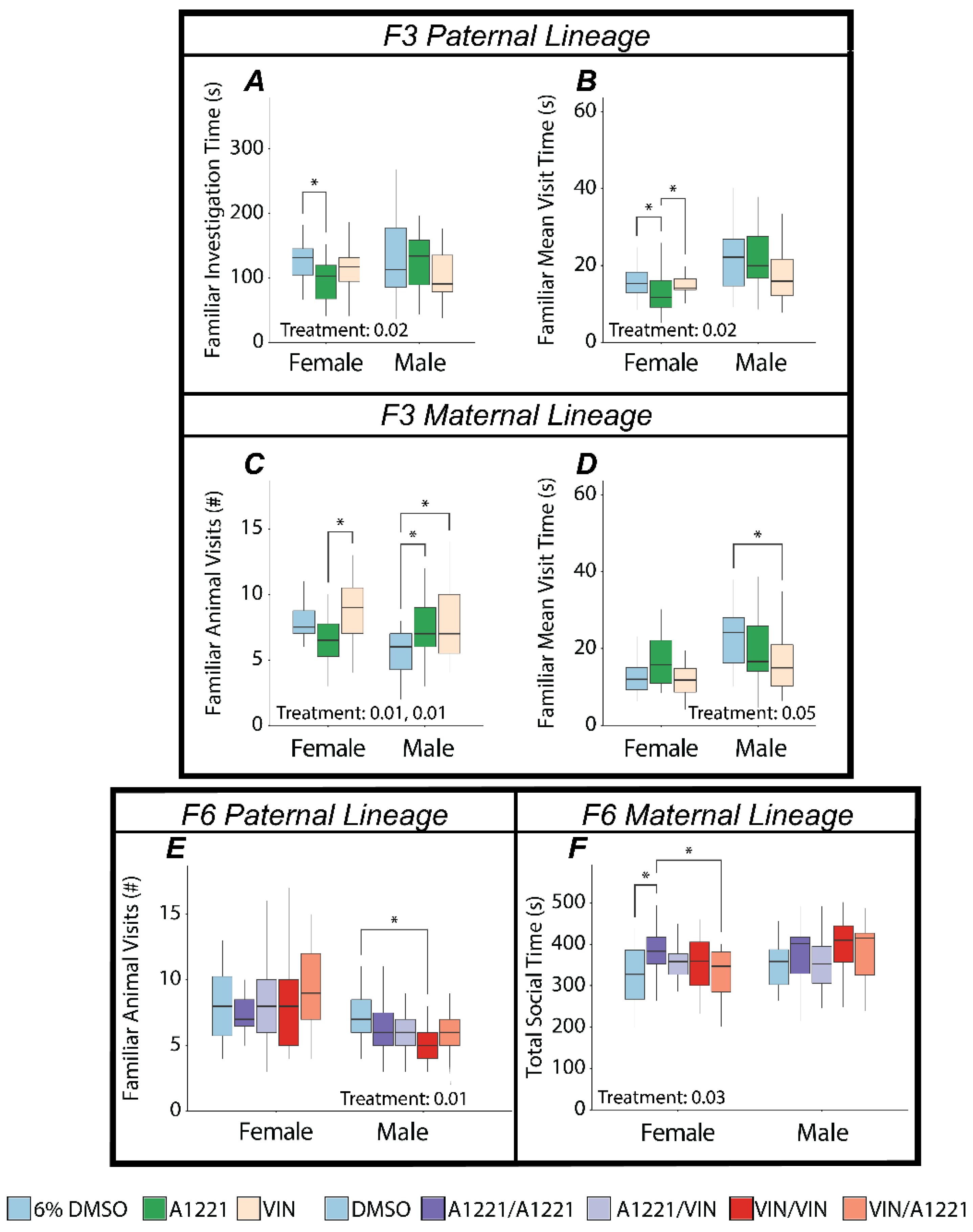
3.2. Social Novelty
3.2.1. Sex Differences
3.2.2. Effect of EDC Exposure
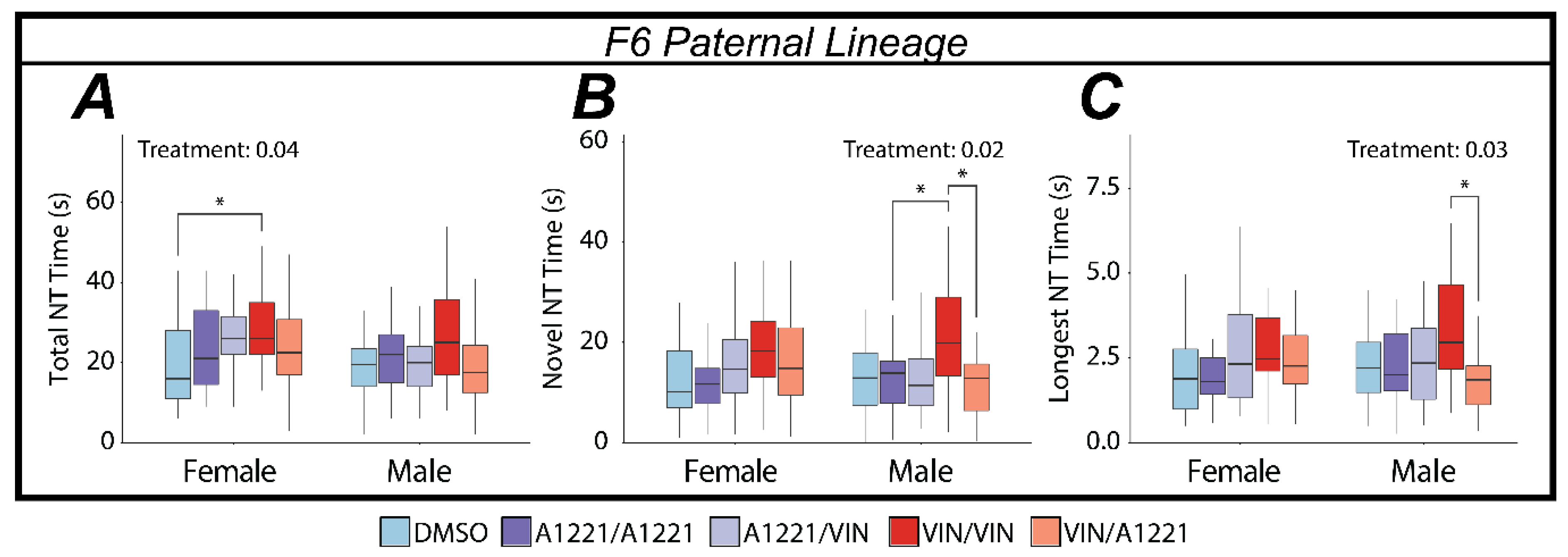
3.3. Nose-Touching Behaviors
3.3.1. Validation of Machine Learning Accuracy
3.3.2. Sex Differences in Nose Touching
3.3.3. EDC Effects on Nose Touching
4. Discussion
4.1. Direct Developmental Exposure to EDCs (F1 Generation) Have Few Effects on Social Behaviors
4.2. EDCs Affect Social Behavior in Ancestrally Exposed Individuals
4.2.1. The Paternal Lineage Females Are Most Vulnerable to Ancestral Exposure
4.2.2. PCBs Are an Underappreciated Agent of Transgenerational Perturbations
4.2.3. The Order of EDC Exposure Is Important
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gore, A.C.; Chappell, V.A.; Fenton, S.E.; Flaws, J.A.; Nadal, A.; Prins, G.S.; Toppari, J.; Zoeller, R.T. EDC-2: The Endocrine Society’s Second Scientific Statement on Endocrine-Disrupting Chemicals. Endocr. Rev. 2015, 36, E1–E150. [Google Scholar] [CrossRef]
- Ribeiro, E.; Ladeira, C.; Viegas, S. EDCs Mixtures: A Stealthy Hazard for Human Health? Toxics 2017, 5, 5. [Google Scholar] [CrossRef] [Green Version]
- Rissman, E.F.; Adli, M. Minireview: Transgenerational Epigenetic Inheritance: Focus on Endocrine Disrupting Compounds. Endocrinology 2014, 155, 2770–2780. [Google Scholar] [CrossRef]
- Van Cauwenbergh, O.; Di Serafino, A.; Tytgat, J.; Soubry, A. Transgenerational epigenetic effects from male exposure to endocrine-disrupting compounds: A systematic review on research in mammals. Clin. Epigenetics 2020, 12, 1–23. [Google Scholar] [CrossRef]
- Jolous-Jamshidi, B.; Cromwell, H.C.; McFarland, A.M.; Meserve, L.A. Perinatal exposure to polychlorinated biphenyls alters social behaviors in rats. Toxicol. Lett. 2010, 199, 136–143. [Google Scholar] [CrossRef] [Green Version]
- Reilly, M.P.; Weeks, C.D.; Topper, V.Y.; Thompson, L.M.; Crews, D.; Gore, A.C. The effects of prenatal PCBs on adult social behavior in rats. Horm. Behav. 2015, 73, 47–55. [Google Scholar] [CrossRef] [Green Version]
- Porrini, S.; Belloni, V.; Della Seta, D.; Farabollini, F.; Giannelli, G.; Dessì-Fulgheri, F. Early exposure to a low dose of bisphenol A affects socio-sexual behavior of juvenile female rats. Brain Res. Bull. 2005, 65, 261–266. [Google Scholar] [CrossRef]
- Wolstenholme, J.T.; Taylor, J.A.; Shetty, S.R.J.; Edwards, M.; Connelly, J.J.; Rissman, E.F. Gestational Exposure to Low Dose Bisphenol A Alters Social Behavior in Juvenile Mice. PLoS ONE 2011, 6, e25448. [Google Scholar] [CrossRef] [Green Version]
- Kougias, D.G.; Cortes, L.; Moody, L.; Rhoads, S.; Pan, Y.-X.; Juraska, J.M. Effects of Perinatal Exposure to Phthalates and a High-Fat Diet on Maternal Behavior and Pup Development and Social Play. Endocrinology 2018, 159, 1088–1105. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.-I.; Chiang, C.-W.; Lin, H.-C.; Zhao, J.-F.; Li, C.-T.; Shyue, S.-K.; Lee, T.-S. Maternal exposure to di-(2-ethylhexyl) phthalate exposure deregulates blood pressure, adiposity, cholesterol metabolism and social interaction in mouse offspring. Arch. Toxicol. 2016, 90, 1211–1224. [Google Scholar] [CrossRef]
- Venerosi, A.; Ricceri, L.; Tait, S.; Calamandrei, G. Sex dimorphic behaviors as markers of neuroendocrine disruption by environmental chemicals: The case of chlorpyrifos. Neuro Toxicol. 2012, 33, 1420–1426. [Google Scholar] [CrossRef]
- Colbert, N.K.W.; Pelletier, N.C.; Cote, J.M.; Concannon, J.B.; Jurdak, N.A.; Minott, S.B.; Markowski, V.P. Perinatal Exposure to Low Levels of the Environmental Antiandrogen Vinclozolin Alters Sex-Differentiated Social Play and Sexual Behaviors in the Rat. Environ. Health Perspect. 2005, 113, 700–707. [Google Scholar] [CrossRef] [Green Version]
- Gore, A.C.; Patisaul, H.B. Neuroendocrine disruption: Historical roots, current progress, questions for the future. Front. Neuroendocr. 2010, 31, 395–399. [Google Scholar] [CrossRef] [Green Version]
- Gore, A.C.; Krishnan, K.; Reilly, M.P. Endocrine-disrupting chemicals: Effects on neuroendocrine systems and the neurobiology of social behavior. Horm. Behav. 2019, 111, 7–22. [Google Scholar] [CrossRef]
- Wolstenholme, J.T.; Edwards, M.; Shetty, S.R.J.; Gatewood, J.D.; Taylor, J.A.; Rissman, E.F.; Connelly, J.J. Gestational Exposure to Bisphenol A Produces Transgenerational Changes in Behaviors and Gene Expression. Endocrinology 2012, 153, 3828–3838. [Google Scholar] [CrossRef] [Green Version]
- Wolstenholme, J.T.; Goldsby, J.A.; Rissman, E.F. Transgenerational effects of prenatal bisphenol a on social recognition. Horm. Behav. 2013, 64, 833–839. [Google Scholar] [CrossRef] [Green Version]
- Quinnies, K.M.; Harris, E.P.; Snyder, R.W.; Sumner, S.S.; Rissman, E.F. Direct and transgenerational effects of low doses of perinatal di-(2-ethylhexyl) phthalate (DEHP) on social behaviors in mice. PLoS ONE 2017, 12, e0171977. [Google Scholar] [CrossRef]
- Quinnies, K.M.; Doyle, T.J.; Kim, K.H.; Rissman, E.F. Transgenerational Effects of Di-(2-Ethylhexyl) Phthalate (DEHP) on Stress Hormones and Behavior. Endocrinology 2015, 156, 3077–3083. [Google Scholar] [CrossRef] [Green Version]
- Crews, D.; Gillette, R.; Scarpino, S.V.; Manikkam, M.; Savenkova, M.I.; Skinner, M.K. Epigenetic transgenerational inheritance of altered stress responses. Proc. Natl. Acad. Sci. USA 2012, 109, 9143–9148. [Google Scholar] [CrossRef] [Green Version]
- Robaire, B.; Delbes, G.; Head, J.A.; Marlatt, V.L.; Martyniuk, C.J.; Reynaud, S.; Trudeau, V.L.; Mennigen, J.A. A cross-species comparative approach to assessing multi- and transgenerational effects of endocrine disrupting chemicals. Environ. Res. 2022, 204, 112063. [Google Scholar] [CrossRef]
- Crews, D.; Gore, A.; Hsu, T.S.; Dangleben, N.L.; Spinetta, M.; Schallert, T.; Anway, M.D.; Skinner, M.K. Transgenerational epigenetic imprints on mate preference. Proc. Natl. Acad. Sci. USA 2007, 104, 5942–5946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krizhevsky, A.; Sutskever, I.; Hinton, G.E. ImageNet Classification with Deep Convolutional Neural Networks. Commun. Acm. 2017, 60, 84–90. [Google Scholar] [CrossRef]
- Luo, H.; Xiong, C.; Fang, W.; Love, P.; Zhang, B.; Ouyang, X. Convolutional neural networks: Computer vision-based workforce activity assessment in construction. Autom. Constr. 2018, 94, 282–289. [Google Scholar] [CrossRef]
- Connor, K.; Ramamoorthy, K.; Moore, M.; Mustain, M.; Chen, I.; Safe, S.; Zacharewski, T.; Gillesby, B.; Joyeux, A.; Balaguer, P. Hydroxylated Polychlorinated Biphenyls (PCBs) as Estrogens and Antiestrogens: Structure–Activity Relationships. Toxicol. Appl. Pharmacol. 1997, 145, 111–123. [Google Scholar] [CrossRef]
- Kelce, W.R.; Monosson, E.; Gamcsik, M.P.; Laws, S.C.; Gray, L.E. Environmental Hormone Disruptors: Evidence That Vinclozolin Developmental Toxicity Is Mediated by Antiandrogenic Metabolites. Toxicol. Appl. Pharmacol. 1994, 126, 276–285. [Google Scholar] [CrossRef]
- Haith, D.A.; Rossi, F.S. Risk Assessment of Pesticide Runoff from Turf. J. Environ. Qual. 2003, 32, 447–455. [Google Scholar] [CrossRef]
- Gray, L.E.; Ostby, J.; Furr, J.; Wolf, C.J.; Lambright, C.; Parks, L.; Veeramachaneni, D.N.; Wilson, V.; Price, M.; Hotchkiss, A.; et al. Effects of environmental antiandrogens on reproductive development in experimental animals. Hum. Reprod. Updat. 2001, 7, 248–264. [Google Scholar] [CrossRef]
- Park, H.-Y.; Hertz-Picciotto, I.; Sovcikova, E.; Kocan, A.; Drobna, B.; Trnovec, T. Neurodevelopmental toxicity of prenatal polychlorinated biphenyls (PCBs) by chemical structure and activity: A birth cohort study. Environ. Health 2010, 9, 51. [Google Scholar] [CrossRef] [Green Version]
- Streifer, M.; Gore, A.C. Epigenetics, Estrogenic Endocrine-Disrupting Chemicals (EDCs), and the Brain. Adv. Pharmacol. 2021, 92, 73–99. [Google Scholar] [CrossRef]
- Hernandez, M.E.; Gore, A.C. Chemical Contaminants—A Toxic Mixture for Neurodevelopment. Nat. Rev. Endocrinol. 2017, 13, 322–323. [Google Scholar] [CrossRef]
- Gillette, R.; Miller-Crews, I.; Nilsson, E.E.; Skinner, M.K.; Gore, A.C.; Crews, D. Sexually Dimorphic Effects of Ancestral Exposure to Vinclozolin on Stress Reactivity in Rats. Endocrinology 2014, 155, 3853–3866. [Google Scholar] [CrossRef] [Green Version]
- Topper, V.Y.; Reilly, M.P.; Wagner, L.M.; Thompson, L.M.; Gillette, R.; Crews, D.; Gore, A.C. Social and neuromolecular phenotypes are programmed by prenatal exposures to endocrine-disrupting chemicals. Mol. Cell. Endocrinol. 2019, 479, 133–146. [Google Scholar] [CrossRef]
- Dickerson, S.M.; Cunningham, S.L.; Patisaul, H.B.; Woller, M.J.; Gore, A.C. Endocrine Disruption of Brain Sexual Differentiation by Developmental PCB Exposure. Endocrinology 2011, 152, 581–594. [Google Scholar] [CrossRef] [Green Version]
- Fitzgerald, E.F.; Belanger, E.E.; Gomez, M.I.; Cayo, M.; McCaffrey, R.J.; Seegal, R.F.; Jansing, R.L.; Hwang, S.-A.; Hicks, H.E. Polychlorinated Biphenyl Exposure and Neuropsychological Status among Older Residents of Upper Hudson River Communities. Environ. Health Perspect. 2008, 116, 209–215. [Google Scholar] [CrossRef] [Green Version]
- Alyea, R.A.; Gollapudi, B.B.; Rasoulpour, R.J. Are we ready to consider transgenerational epigenetic effects in human health risk assessment? Environ. Mol. Mutagen. 2013, 55, 292–298. [Google Scholar] [CrossRef]
- Steinberg, R.M.; Juenger, T.; Gore, A.C. The effects of prenatal PCBs on adult female paced mating reproductive behaviors in rats. Horm. Behav. 2007, 51, 364–372. [Google Scholar] [CrossRef] [Green Version]
- Arnold, A.P.; Gorski, R.A. Gonadal Steroid Induction of Structural Sex Differences in the Central Nervous System. Annu. Rev. Neurosci. 1984, 7, 413–442. [Google Scholar] [CrossRef]
- Lee, H.J.; Hore, T.A.; Reik, W. Reprogramming the Methylome: Erasing Memory and Creating Diversity. Cell Stem Cell 2014, 14, 710–719. [Google Scholar] [CrossRef] [Green Version]
- Nadler, J.J.; Moy, S.S.; Dold, G.; Simmons, N.; Perez, A.; Young, N.B.; Barbaro, R.P.; Piven, J.; Magnuson, T.R.; Crawley, J.N. Automated apparatus for quantitation of social approach behaviors in mice. Genes Brain Behav. 2004, 3, 303–314. [Google Scholar] [CrossRef]
- Reilly, M.P.; Weeks, C.D.; Crews, D.; Gore, A.C. Application of a novel social choice paradigm to assess effects of prenatal endocrine-disrupting chemical exposure in rats (Rattus norvegicus). J. Comp. Psychol. 2018, 132, 253–267. [Google Scholar] [CrossRef]
- Mathis, A.; Mamidanna, P.; Cury, K.M.; Abe, T.; Murthy, V.N.; Mathis, M.W.; Bethge, M. DeepLabCut: Markerless pose estimation of user-defined body parts with deep learning. Nat. Neurosci. 2018, 21, 1281–1289. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. Available online: https://www.r-project.org/ (accessed on 10 November 2021).
- Royston, J.P. Algorithm AS 181: The W Test for Normality. J. R. Stat. Soc. Ser. C Appl. Stat. 1982, 31, 176. [Google Scholar] [CrossRef]
- Bartlett, M.S. Properties of sufficiency and statistical tests. Proc. R. Soc. Lond. Ser. A Math. Phys. Sci. 1937, 160, 268–282. [Google Scholar] [CrossRef]
- Kruskal, W.H.; Wallis, W.A. Use of Ranks in One-Criterion Variance Analysis. J. Am. Stat. Assoc. 1952, 47, 583–621. [Google Scholar] [CrossRef]
- Diener, M.J. Corsini Encyclopedia of Psychology; Wiley: Hoboken, NJ, USA, 2010. [Google Scholar] [CrossRef]
- Millard, S. EnvStats: An R Package for Environmental Statistics; Springer: New York, NY, USA, 2013; ISBN 978-1-4614-8455-4. [Google Scholar]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019. [Google Scholar]
- Navarro, D. Learning Statistics with R: A Tutorial for Psychology Students and other Beginners; University of Adelaide: Adelaide, Australia, 2015. [Google Scholar]
- Tukey, J.W. Exploratory Data Analysis; Addison-Wesley: Reading, MA, USA, 1977; Volume 2. [Google Scholar]
- Hernandez Scudder, M.E.; Weinberg, A.; Thompson, L.; Crews, D.; Gore, A.C. Prenatal EDCs Impair Mate and Odor Preference and Activation of the VMN in Male and Female Rats. Endocrinology 2020, 161, bqaa124. [Google Scholar] [CrossRef] [PubMed]
- Johnston, A.L.; File, S.E. Sex differences in animal tests of anxiety. Physiol. Behav. 1991, 49, 245–250. [Google Scholar] [CrossRef]
- Holmes, M.M.; Niel, L.; Anyan, J.J.; Griffith, A.T.; Monks, D.A.; Forger, N.G. Effects of Bax gene deletion on social behaviors and neural response to olfactory cues in mice. Eur. J. Neurosci. 2011, 34, 1492–1499. [Google Scholar] [CrossRef]
- Karlsson, S.A.; Haziri, K.; Hansson, E.; Kettunen, P.; Westberg, L. Effects of sex and gonadectomy on social investigation and social recognition in mice. BMC Neurosci. 2015, 16, 83. [Google Scholar] [CrossRef] [Green Version]
- Hotchkiss, A.; Ostby, J.; Vandenbergh, J.; Gray, L. An environmental antiandrogen, vinclozolin, alters the organization of play behavior. Physiol. Behav. 2003, 79, 151–156. [Google Scholar] [CrossRef]
- Vandenberg, L.N.; Colborn, T.; Hayes, T.B.; Heindel, J.J.; Jacobs, D.R., Jr.; Lee, D.-H.; Shioda, T.; Soto, A.M.; vom Saal, F.S.; Welshons, W.V.; et al. Hormones and Endocrine-Disrupting Chemicals: Low-Dose Effects and Nonmonotonic Dose Responses. Endocr. Rev. 2012, 33, 378–455. [Google Scholar] [CrossRef]
- Gillette, R.; Son, M.J.; Ton, L.; Gore, A.C.; Crews, D. Passing experiences on to future generations: Endocrine disruptors and transgenerational inheritance of epimutations in brain and sperm. Epigenetics 2018, 13, 1106–1126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soubry, A.; Hoyo, C.; Jirtle, R.L.; Murphy, S.K. A paternal environmental legacy: Evidence for epigenetic inheritance through the male germ line. BioEssays 2014, 36, 359–371. [Google Scholar] [CrossRef] [PubMed]
- Gore, A.C.; Thompson, L.M.; Bell, M.; A Mennigen, J. Transgenerational effects of polychlorinated biphenyls: Hypothalamic gene expression in rats. Biol. Reprod. 2021, 105, 690–704. [Google Scholar] [CrossRef] [PubMed]
- Mennigen, J.A.; Thompson, L.M.; Bell, M.; Santos, M.T.; Gore, A.C. Transgenerational effects of polychlorinated biphenyls: Development and physiology across 3 generations of rats. Environ. Health 2018, 17, 18. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| F1 | F3 | ||||||
| First Hit | Maternal | Paternal | |||||
| DMSO | 10/21/25 | 11/21/21 | 10/23/21 | ||||
| A1221 | 10/28/24 | 10/18/24 | 11/22/18 | ||||
| VIN | 10/28/24 | 12/19/23 | 11/18/21 | ||||
| F4 | F6 | ||||||
| First Hit/Second Hit | Maternal | Paternal | Maternal | Paternal | |||
| DMSO/DMSO | 13/22/21 | 12/25/22 | 12/22/23 | 12/21/24 | |||
| A1221/A1221 | 9/15/18 | 8/18/15 | 9/18/20 | 8/18/20 | |||
| A1221/VIN | 9/18/19 | 9/17/18 | 9/22/18 | 9/20/20 | |||
| VIN/A1221 | 12/21/22 | 11/21/21 | 11/19/23 | 11/22/21 | |||
| VIN/VIN | 13/20/22 | 12/19/20 | 12/22/22 | 10/19/22 | |||
| Sociability | ||||
| Females | Males | |||
| Measure | Mean | SEM | Mean | SEM |
| Distance Traveled (m) * | 52.63 | 0.36 | 38.58 | 0.3 |
| Social Preference Score * | 0.62 | 0.01 | 0.67 | 0.01 |
| Stimulus Investigation Time (s) * | 224.35 | 2.68 | 245.90 | 3.02 |
| Empty Investigation Time (s) * | 135.26 | 2.13 | 123.31 | 2.59 |
| Stimulus Animal # Visits * | 10.26 | 0.13 | 8.68 | 0.14 |
| Empty Chamber # Visits * | 8.42 | 0.11 | 6.25 | 0.09 |
| Stimulus Animal Mean Visit Time (s) * | 24.46 | 0.5 | 34.76 | 1.06 |
| Empty Chamber Mean Visit Time (s) * | 17.58 | 0.38 | 22.78 | 0.68 |
| Total Nonsocial Time (s) * | 240.01 | 2.79 | 229.86 | 2.86 |
| Center Isolation Time (s) * | 99.54 | 1.19 | 107.72 | 1.6 |
| Total Nose-Touch Time (s) | 20.21 | 0.40 | 19.51 | 0.39 |
| Average Nose-Touch Duration (s) * | 0.66 | 0.01 | 0.63 | 0.01 |
| Social Novelty | ||||
| Females | Males | |||
| Measure | Mean | SEM | Mean | SEM |
| Distance Traveled (m) * | 46.35 | 0.39 | 32.44 | 0.29 |
| Social Novelty Score * | 0.62 | 0.01 | 0.60 | 0.01 |
| Familiar Investigation Time (s) * | 124.31 | 2.16 | 142.47 | 2.84 |
| Novel Investigation Time (s) | 208.61 | 2.81 | 215.03 | 3.17 |
| Familiar Animal # Visits * | 8.01 | 0.11 | 6.32 | 0.09 |
| Novel Animal # Visits * | 10.09 | 0.13 | 7.74 | 0.11 |
| Familiar Animal Mean Visit Time (s) * | 16.89 | 0.55 | 24.67 | 0.6 |
| Novel Animal Mean Visit Time (s) * | 22.46 | 0.44 | 31.39 | 0.72 |
| Total Social Time (s) * | 332.92 | 2.96 | 357.50 | 3.16 |
| Total Nonsocial Time (s) * | 273.37 | 3.77 | 249.89 | 4.04 |
| Center Isolation Time (s) * | 125.54 | 1.78 | 116.37 | 1.92 |
| Nose-Touch Novelty Score | 0.65 | 0.01 | 0.64 | 0.01 |
| Familiar Animal Nose-Touch Time (s) * | 9.33 | 0.27 | 8.62 | 0.30 |
| Novel Animal Nose-Touch Time (s) * | 17.77 | 0.41 | 15.41 | 0.38 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gillette, R.; Dias, M.; Reilly, M.P.; Thompson, L.M.; Castillo, N.J.; Vasquez, E.L.; Crews, D.; Gore, A.C. Two Hits of EDCs Three Generations Apart: Effects on Social Behaviors in Rats, and Analysis by Machine Learning. Toxics 2022, 10, 30. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics10010030
Gillette R, Dias M, Reilly MP, Thompson LM, Castillo NJ, Vasquez EL, Crews D, Gore AC. Two Hits of EDCs Three Generations Apart: Effects on Social Behaviors in Rats, and Analysis by Machine Learning. Toxics. 2022; 10(1):30. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics10010030
Chicago/Turabian StyleGillette, Ross, Michelle Dias, Michael P. Reilly, Lindsay M. Thompson, Norma J. Castillo, Erin L. Vasquez, David Crews, and Andrea C. Gore. 2022. "Two Hits of EDCs Three Generations Apart: Effects on Social Behaviors in Rats, and Analysis by Machine Learning" Toxics 10, no. 1: 30. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics10010030