
The Recolonization Concentration Concept: Using Avoidance Assays with Soil Organisms to Predict the Recolonization Potential of Contaminated Sites


Abstract
:1. Introduction
2. Material and Methods
2.1. Test Soil
2.2. Test Organisms and Culture Conditions
3. Experimental Procedure
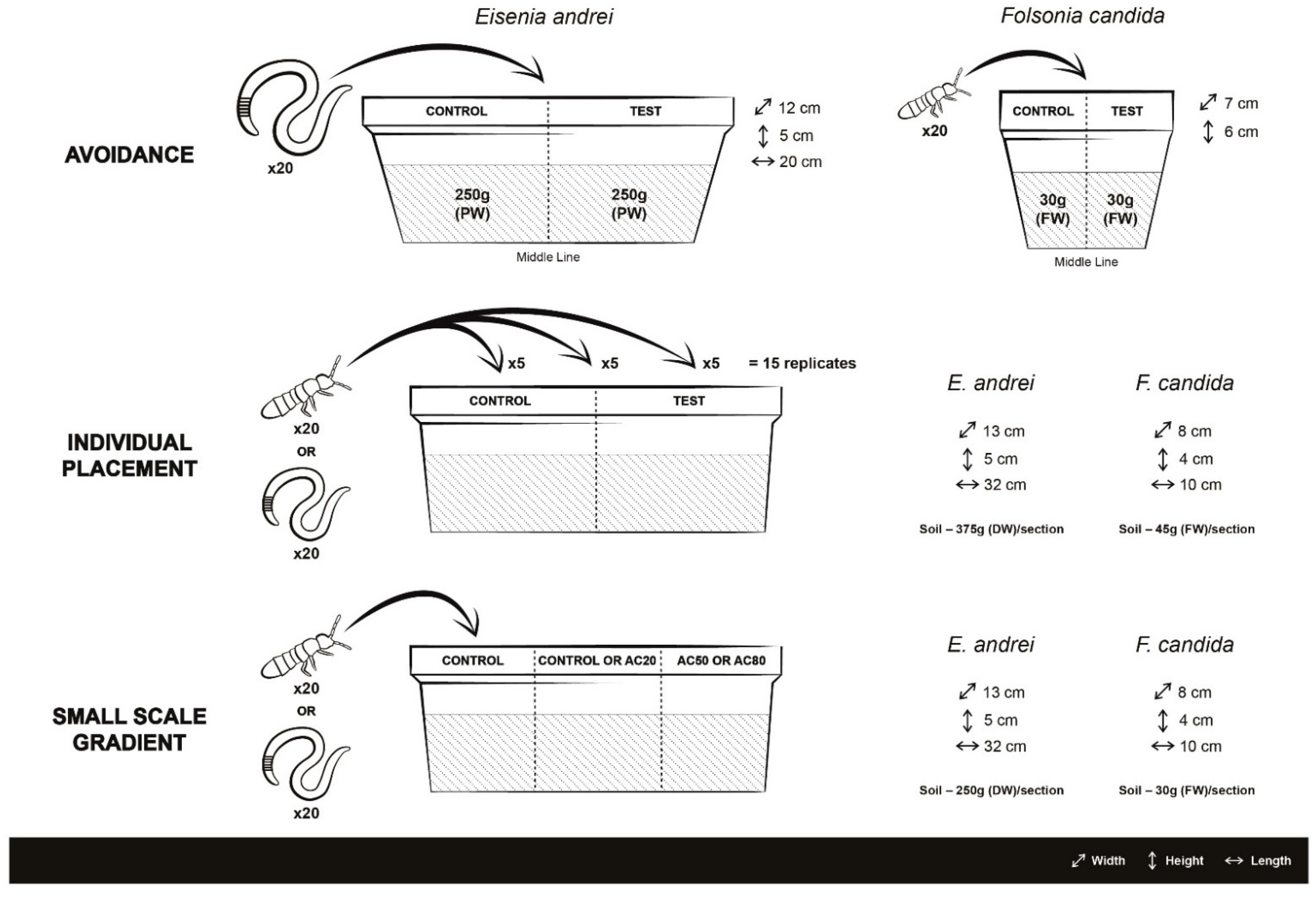
3.1. Avoidance Tests
3.2. Individual Placement Tests
3.3. Small-Scale Gradient Tests
4. Metal Analysis
5. Statistical Analysis
6. Results
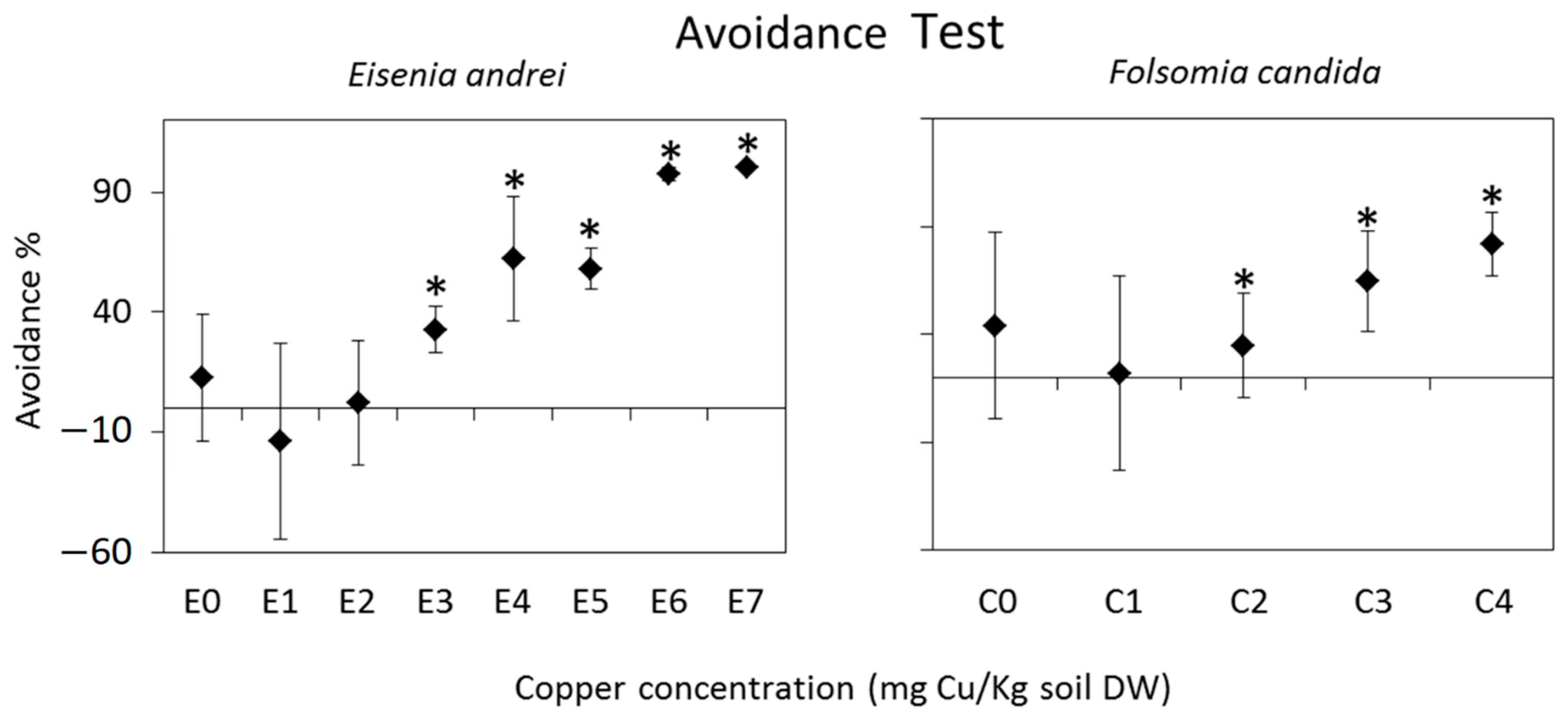
6.1. Avoidance Tests
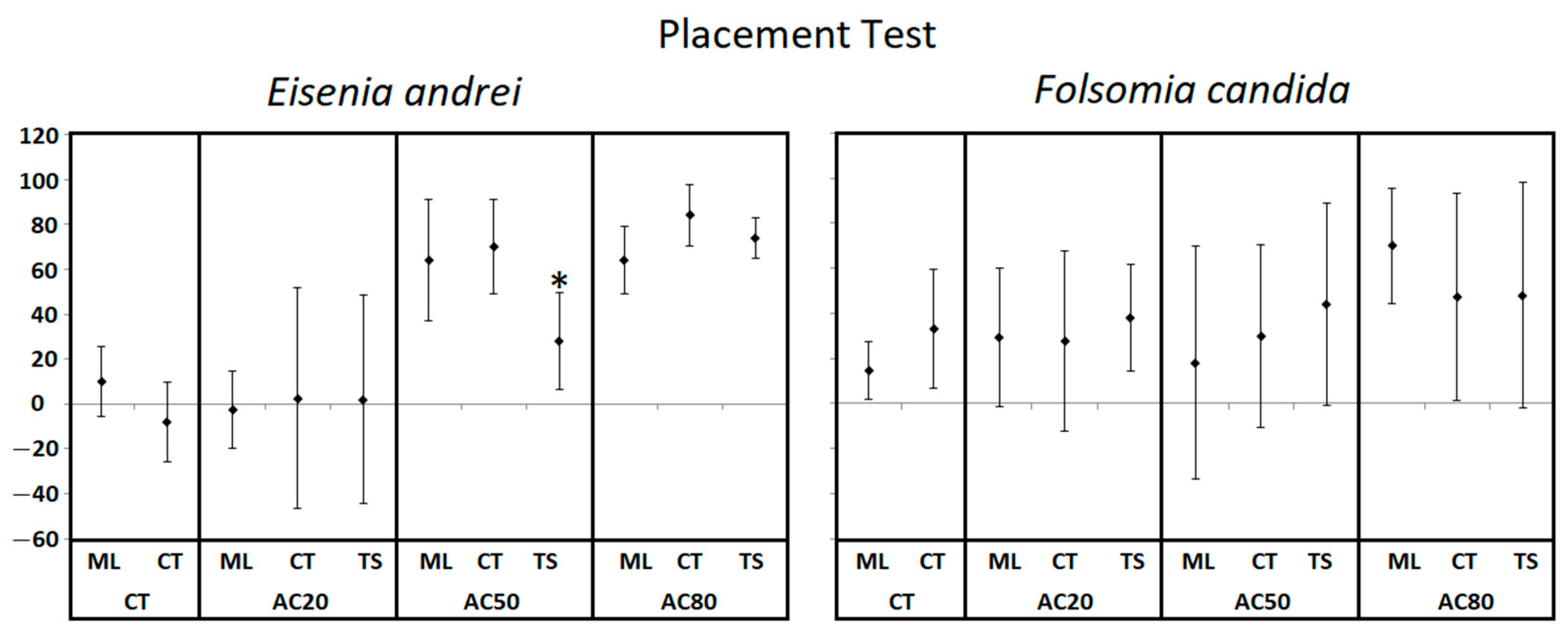
6.2. Individual Placement Test
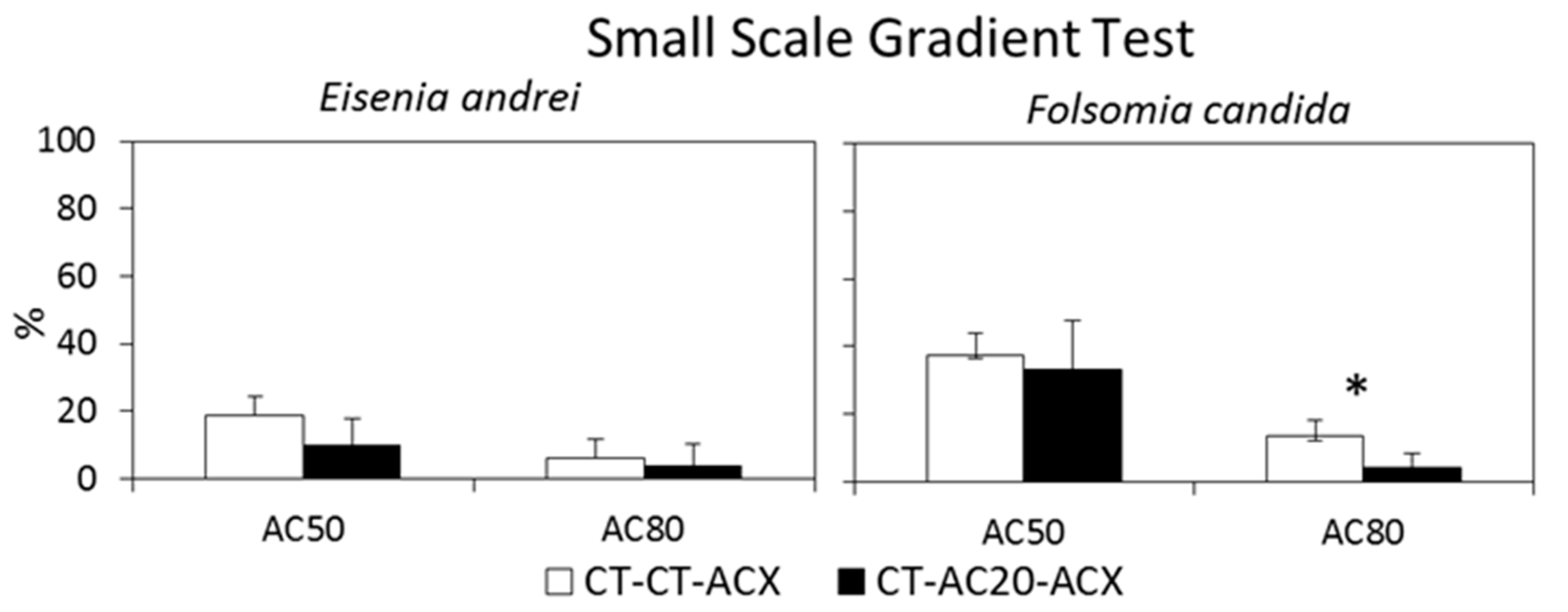
6.3. Small-Scale Gradient Tests
7. Discussion
8. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Brock, T.; Arts, G.; Matlby, L.; Van den Brink, P. Aquatic risks of pesticides, ecological protection goals, and common aims in European Union legislation. Integr. Environ. Assess. Manag. 2006, 2, 20–46. [Google Scholar] [CrossRef]
- Nienstedt, K.M.; Brock, T.C.M.; van Wensem, J.; Montforts, M.; Hart, A.; Aagaard, A.; Alix, A.; Boesten, J.; Bopp, S.K.; Brown, C.; et al. Development of a framework based on an ecosystem services approach for deriving specific protection goals for environmental risk assessment of pesticides. Sci. Total Environ. 2012, 415, 31–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Linden, A.M.A.; Boesten, J.J.T.; Brock, T.C.M.; Van Eekelen, G.M.A.; Horst, M.M.S.; Montforts, M.H.M.M.; Pol, J.W. Revised Proposal for the Risk Assessment of Persistence of Plant Protection Products in Soil; RIVM: Bilthoven, The Netherlands, 2008; ISBN 978-90-6960-198-4. [Google Scholar]
- Ockleford, C.; Adriaanse, P.; Berny, P.; Brock, T.; Duquesne, S.; Grilli, S.; Hernandez-Jerez, A.F.; Bennekou, S.H.; Klein, M.; Kuhl, T.; et al. Scientific Opinion addressing the state of the science on risk assessment of plant protection products for in-soil organisms. EFSA J. 2017, 15, e04690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lange, H.; Lahr, J.; Pol, V.; Wessels, Y.; Faber, J. Ecological vulnerability in wildlife: An expert judgment and multicriteria analysis tool using ecological traits to assess relative impact of pollutants. Environ. Toxicol. Chem. 2009, 28, 2233–2240. [Google Scholar] [CrossRef]
- Alvarez, T.; Frampton, G.; Goulson, D. The role of hedgerows in the recolonisation of arable fields by epigeal Collembola. Pedobiologia 2000, 44, 516–526. [Google Scholar] [CrossRef]
- Frampton GGould, P.; Van den Brink, P.; Hendy, E. Type ‘A’ and ‘B’ recovery revisited: The role of field-edge habitats for Collembola and macroarthropod community recovery after insecticide treatment. Environ. Pollut. 2006, 145, 874–883. [Google Scholar] [CrossRef]
- Marinissen, J.C.Y.; Van den Bosch, F. Colonization of new habitats by earthworms. Oecologia 1992, 91, 371–376. [Google Scholar] [CrossRef]
- Ojala, R.; Huhta, V. Dispersal of microarthropods in forest soil. Pedobiologia 2001, 45, 443–450. [Google Scholar] [CrossRef]
- Auclerc, A.; Ponge, J.F.; Barot, S.; Dubs, F. Experimental assessment of habitat preference and dispersal ability of soil springtails. Soil Biol. Biochem. 2009, 41, 1596–1604. [Google Scholar] [CrossRef] [Green Version]
- Caro, G.; Decaëns, T.; Mathieu, J. The dispersal of the earthworm Aporrectodea giardi responds faster to habitat quality than to cumulative use of habitat in experimental conditions. Appl. Soil Ecol. 2013, 71, 45–47. [Google Scholar] [CrossRef]
- Mathieu, J.; Barot, S.; Blouin, M.; Caro, G.; Decaëns, T.; Dubs, F.; Dupont, L.; Jouquet, P.; Nai, P. Habitat quality, conspecific density, and habitat pre-use affect the dispersal behaviour of two earthworm species, Aporrectodea icterica and Dendrobaena veneta, in a mesocosm experiment. Soil Biol. Biochem. 2010, 42, 203–209. [Google Scholar] [CrossRef]
- Fenoglio, S.; Cucco, M.; Bo, T.; Gay, P.; Malacarne, G. Colonisation by ground and edaphic invertebrates of soil patches with different pollution levels. Period. Biol. 2007, 109, 37–45. [Google Scholar]
- Holmstrup, M.; Hornum, H.D. Earthworm colonisation of abandoned arable soil polluted by copper. Pedobiologia 2012, 55, 63–65. [Google Scholar] [CrossRef]
- Forbes, V.E.; Calow, P.; Grimm, V.; Hayashi, T.; Jager, T.; Palmqvist, A.; Pastorok, R.; Salvito, D.; Sibly, R.; Spromberg, J.; et al. Integrating population modeling into ecological risk assessment. Integr. Environ. Assess. Manag. 2010, 6, 191–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forbes, V.E.; Calow, P.; Sibly, R.M. The extrapolation problem and how population modeling can help. Environ. Toxicol. Chem. 2008, 27, 1987–1994. [Google Scholar] [CrossRef] [Green Version]
- Grimm, V.; Thorbek, P. Population models for ecological risk assessment of chemicals: Short introduction and summary of a special issue. Ecol. Model. 2014, 280, 1–4. [Google Scholar] [CrossRef]
- Hamda, N.T.; Forbes, V.E.; Stark, J.D.; Laskowski, R. Stochastic density-dependent matrix model for extrapolating individual-level effects of chemicals to the population: Case study on effects of Cd on Folsomia candida. Ecol. Model. 2014, 280, 53–64. [Google Scholar] [CrossRef]
- Meli, M.; Auclerc, A.; Palmqvist, A.; Forbes, V.E.; Grimm, V. Population-level consequences of spatially heterogeneous exposure to heavy metals in soil: An individual-based model of springtails. Ecol. Model. 2013, 250, 338–351. [Google Scholar] [CrossRef]
- Meli, M.; Palmqvist, A.; Forbes, V.E.; Groeneveld, J.; Grimm, V. Two pairs of eyes are better than one: Combining individual-based and matrix models for ecological risk assessment of chemicals. Ecol. Model. 2014, 280, 40–52. [Google Scholar] [CrossRef]
- ISO 17512-1; Soil Quality—Avoidance Test for Testing the Quality of Soils and Effects of Chemicals on Behaviour—Test with Earthworms (Eisenia fetida and Eisenia andrei). International Organization for Standardization: Paris, France, 2007.
- ISO 17512-2; Soil quality—Avoidance Test for Determining the Quality of Soils and Effects of Chemicals on Behaviour—Part 2: Test with Collembolans (Folsomia candida). International Organization for Standardization: Geneva, Switzerland, 2008.
- Natal-da-Luz, T.; Ribeiro, R.; Sousa, J.P. Avoidance tests with Collembola and earthworms as early screening tools for site-specific assessment of polluted soils. Environ. Toxicol. Chem. 2004, 23, 2188–2193. [Google Scholar] [CrossRef]
- Natal-da-Luz, T.; Römbke, J.; Paulo Sousa, J. Avoidance tests in site-specific risk assessment—Influence of soil propertires on the avoidance response of collembola and earthworms. Environ. Toxicol. Chem. 2008, 27, 1112–1117. [Google Scholar] [CrossRef] [PubMed]
- Niemeyer, J.C.; Moreira-Santos, M.; Nogueira, M.A.; Carvalho, G.M.; Ribeiro, R.; Silva, E.M.; Sousa, J.P. Environmental risk assessment of a metal-contaminated area in the Tropics. Tier I: Screening phase. J. Soils Sediments 2010, 10, 1557–1571. [Google Scholar] [CrossRef]
- Salminen, J.; Sulkava, P. Distribution of soil animals in patchily contaminated soil. Soil Biol. Biochem. 1996, 28, 1349–1355. [Google Scholar] [CrossRef]
- Haimi, J.; Paavola, S. Responses of two earthworm populations with different exposure histories to chlorophenol contamination. Environ. Toxicol. Chem. 1998, 17, 1114–1117. [Google Scholar] [CrossRef]
- Araújo, C.V.M.; Moreira-Santos, M.; Ribeiro, R. Stressor-driven emigration and recolonisation patterns in disturbed habitats. Sci. Total Environ. 2018, 643, 884–889. [Google Scholar] [CrossRef]
- LNEC-E 239; Solos—Análise Granulométrica por Peneiração Húmida. Laboratório Nacional de Engenharia Civil (LNEC): Lisboa, Portugal, 1970.
- ISO 11274; Soil Quality—Determination of Water-Retention Characteristic—Laboratory Methods. International Organization for Standardization: Paris, France, 2007.
- Kjeldahl, J. Neue Methode zul Bestimmung des Stickstoffs in organischen Körpern. Z. Anal. Chem. 1883, 22, 366–382. [Google Scholar] [CrossRef] [Green Version]
- ISO 11260; Soil Quality—Determination of Effective Cation Exchange Capacity and Base Saturation Level Using Barium Chloride Solution. International Organization for Standardization: Geneva, Switzerland, 2007.
- Sakuma, M. Probit analysis of preference data. Appl. Entomol. Zool. 1998, 33, 339–347. [Google Scholar] [CrossRef] [Green Version]
- Loureiro, S.; Soares, A.M.V.M.; Nogueira, A.J.A. Terrestrial avoidance behaviour tests as screening tool to assess soil contamination. Environ. Pollut. 2005, 138, 121–131. [Google Scholar] [CrossRef]
- Xing, Y.; Luo, J.; Zhang, J.; Li, B.; Gong, X.; Liu, Z.; Liu, C. Effects of single and combined exposures to copper and benzotriazole on Eisenia fetida. Chemosphere 2017, 186, 108–115. [Google Scholar] [CrossRef]
- Greenslade, P.; Vaughan, G.T. A comparison of Collembola species for toxicity testing of Australian soils. Pedobiologia 2003, 47, 171–179. [Google Scholar] [CrossRef]
- Criel, P.; Lock, K.; Van Eeckhout, H.; Oorts, K.; Smolders, E.; Janssen, C.R. Influence of soil properties on copper toxicity for two soil invertebrates. Environ. Toxicol. Chem. 2008, 27, 1748–1755. [Google Scholar] [CrossRef] [PubMed]
- Smolders, E.; Oorts, K.; Sprang, P.V.; Schoeters, I.; Janssen, C.R.; McGrath, S.P. Toxicity of trace metals in soi as affected by soil type and aging after contamination: Using calibrated bioavailability models to set ecological soil standards. Environ. Toxicol. Chem. 2009, 28, 1633–1641. [Google Scholar] [CrossRef] [PubMed]
- Arnold, R.E.; Hodson, M.E.; Black, S.; Davies, N.A. The influence of mineral solubility and soil solution concentration on the toxicity of copper to Eisenia fetida Savigny. Pedobiologia 2003, 2, 622–632. [Google Scholar] [CrossRef]
- Boiteau, G.; Lynch, D.H.; Mackinley, P. Avoidance tests with Folsomia candida for the assessment of copper contamination in agricultural soils. Environ. Pollut. 2011, 159, 903–906. [Google Scholar] [CrossRef] [PubMed]
- Heupel, K. Avoidance response of different collembolan species to Betanal. Eur. J. Soil Biol. 2002, 38, 273–276. [Google Scholar] [CrossRef]
- Bengtsson, G.; Rundgren, S.; Sjögren, M. Modelling dispersal distances in a soil gradient: The influence of metal resistance, competition, and experience. Oikos 1994, 71, 13–23. [Google Scholar] [CrossRef]
- Pereira, C.M.S.; Novais, S.C.; Soares, A.M.V.M.; Amorim, M.J.B. Dimethoate Affects Cholinesterases in Folsomia candida and Their Locomotion—False Negative Results of an Avoidance Behaviour Test. Sci. Total Environ. 2013, 443, 821–827. [Google Scholar] [CrossRef]
- Szabó, B.; Seres, A.; Bakonyi, G. Folsomia candida (collembola) locomotor activity pattern is changed by a neurotoxicant pesticide. Acta Zool. Acad. Sci. Hung. 2018, 64, 355–368. [Google Scholar] [CrossRef]
- Natal-da-Luz, T.; Amorim, M.J.B.; Römbke, J.; Sousa, J.P. Avoidance tests with earthworms and springtails: Defining the minimum exposure time to observe a significant response. Ecotoxicol. Environ. Saf. 2008, 71, 545–551. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Value |
|---|---|
| Water-holding capacity | 36.2 ± 0.4% |
| Organic matter | 3.3 ± 0.1% |
| pH (KCl 1 M) | 6.9 |
| Total N | 0.83 mg/g |
| Cation-exchange capacity | 0.0125 cmol/g |
| Sand | 62.40% |
| Silt | 21.20% |
| Clay | 16.40% |
| Soil texture class | Sandy-loam |
| Soil type | Cambisol |
| Nominal (mg/kg) | n | Total Copper | Extractable Copper | ||
|---|---|---|---|---|---|
| (mg/kg) | (mg/kg) | ||||
| Eisenia andrei | E0/CT | 0 | 15 | 29.37 ± 2.64 | 0.06 ± 0.12 |
| E1 | 5 | 3 | 34.45 ± 3.77 | 0.05 ± 0.03 | |
| E2 | 10 | 3 | 48.25 ± 3.59 | 0.06 ± 0.1 | |
| E3 | 25 | 3 | 54.27 ± 7.49 | 0.58 ± 0.23 | |
| E4 | 50 | 3 | 89.12 ± 5.42 | 1.20 ± 0.07 | |
| E5 | 100 | 3 | 140.60 ± 3.71 | 5.42 ± 0.45 | |
| E6 | 200 | 3 | 239.99 ± 31.36 | 22.11 ± 2.60 | |
| E7 | 400 | 3 | 551.57 ± 30.6 | 73.2 ± 5.80 | |
| AC20 | 20 | 9 | 50.79 ± 4.48 | 0.42 ± 0.15 | |
| AC50 | 50 | 6 | 74.49 ± 9.59 | 1.90 ± 0.28 | |
| AC80 | 100 | 6 | 147.09 ± 15.43 | 5.66 ± 0.61 | |
| Folsomia candida | C0/CT | 0 | 9 | 30.21 ± 3.77 | 0 ± 0.16 |
| C1 | 100 | 3 | 185.29 ± 18.44 | 6.87 ± 0.44 | |
| C2 | 200 | 3 | 330.84 ± 21.46 | 26.15 ± 0.27 | |
| C3 | 800 | 3 | 1173.89 ± 22.30 | 159.25 ± 21.66 | |
| C4 | 1600 | 3 | 2257.49 ± 34.95 | 709.91 ± 77.53 | |
| AC20 | 300 | 6 | 382.36 ± 47.44 | 42.50 ± 9.80 | |
| AC50 | 1000 | 6 | 1061.51 ± 71.79 | 264.28 ± 46.70 | |
| AC80 | 3300 | 6 | 3765.78 ± 276.73 | 1466.79 ± 143.88 |
| ACx | Eisenia andrei | Folsomia candida |
|---|---|---|
| AC20 | 21.8 (3.9–39.7) | 301 (49.8–527) |
| AC50 | 49.5 (20.6–78.0) | 1008 (601–1890) |
| AC80 | 112 (70.9–237) | 3371 (1824–26,987) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Renaud, M.; Natal-da-Luz, T.; Ribeiro, R.; Sousa, J.P. The Recolonization Concentration Concept: Using Avoidance Assays with Soil Organisms to Predict the Recolonization Potential of Contaminated Sites. Toxics 2022, 10, 127. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics10030127
Renaud M, Natal-da-Luz T, Ribeiro R, Sousa JP. The Recolonization Concentration Concept: Using Avoidance Assays with Soil Organisms to Predict the Recolonization Potential of Contaminated Sites. Toxics. 2022; 10(3):127. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics10030127
Chicago/Turabian StyleRenaud, Mathieu, Tiago Natal-da-Luz, Rui Ribeiro, and José Paulo Sousa. 2022. "The Recolonization Concentration Concept: Using Avoidance Assays with Soil Organisms to Predict the Recolonization Potential of Contaminated Sites" Toxics 10, no. 3: 127. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics10030127