
Enhanced Uptake of Arsenic Induces Increased Toxicity with Cadmium at Non-Toxic Concentrations on Caenorhabditis elegans

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Test Solutions
2.2. Test Organism
2.3. Toxicity Experiments
2.4. Cd and As Accumulation in C. elegans
2.5. Cd and As Distribution in C. elegans
2.6. As Species Alteration Corresponding to pH
2.7. Statistical Analysis
3. Results
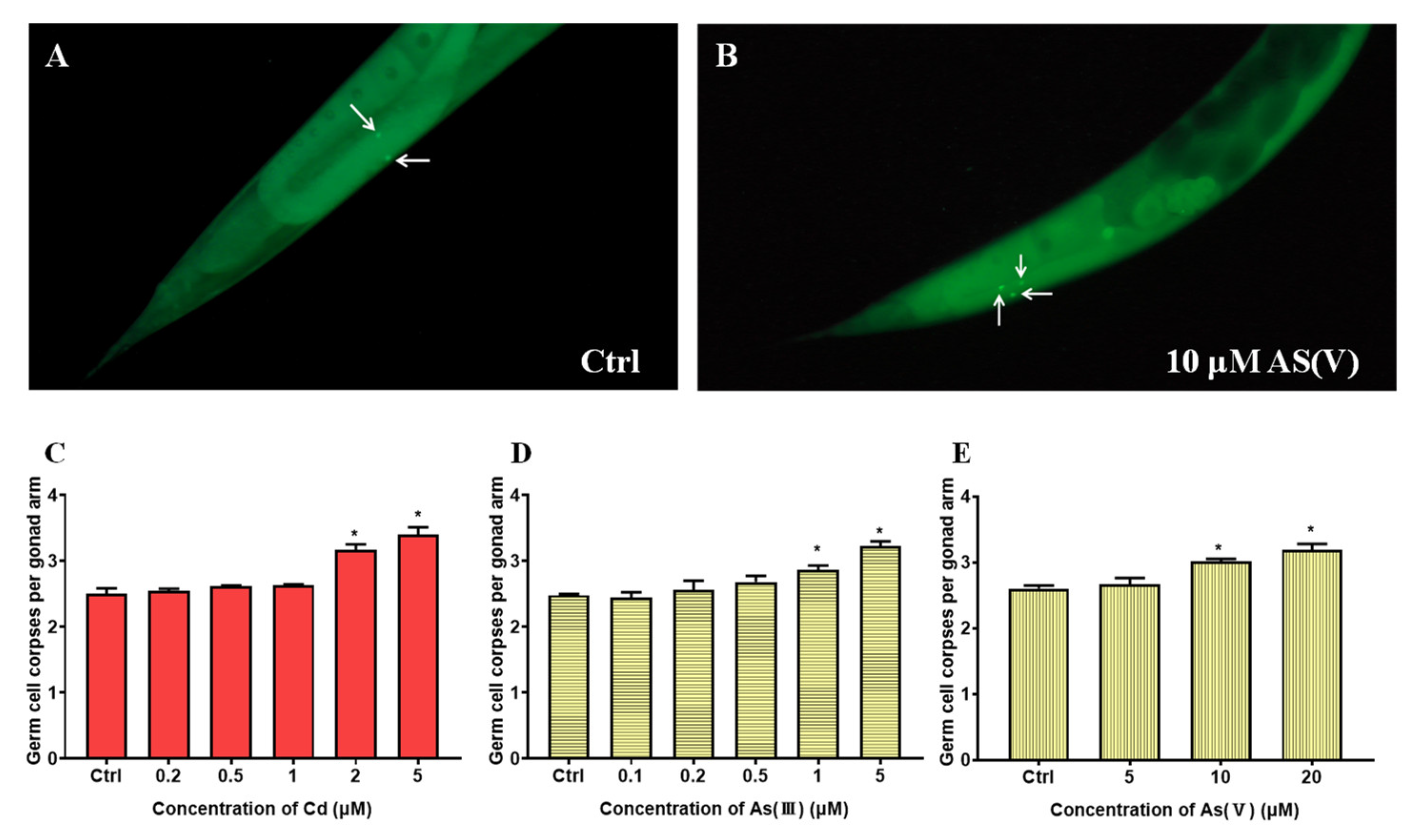
3.1. Joint Toxicity of Cd and As at Non-Toxic Concentrations on C. elegans
3.2. The Accumulation and Distribution of Cd and As in C. elegans
3.3. The Influence of Intestinal Microenvironment (pH) on As Species
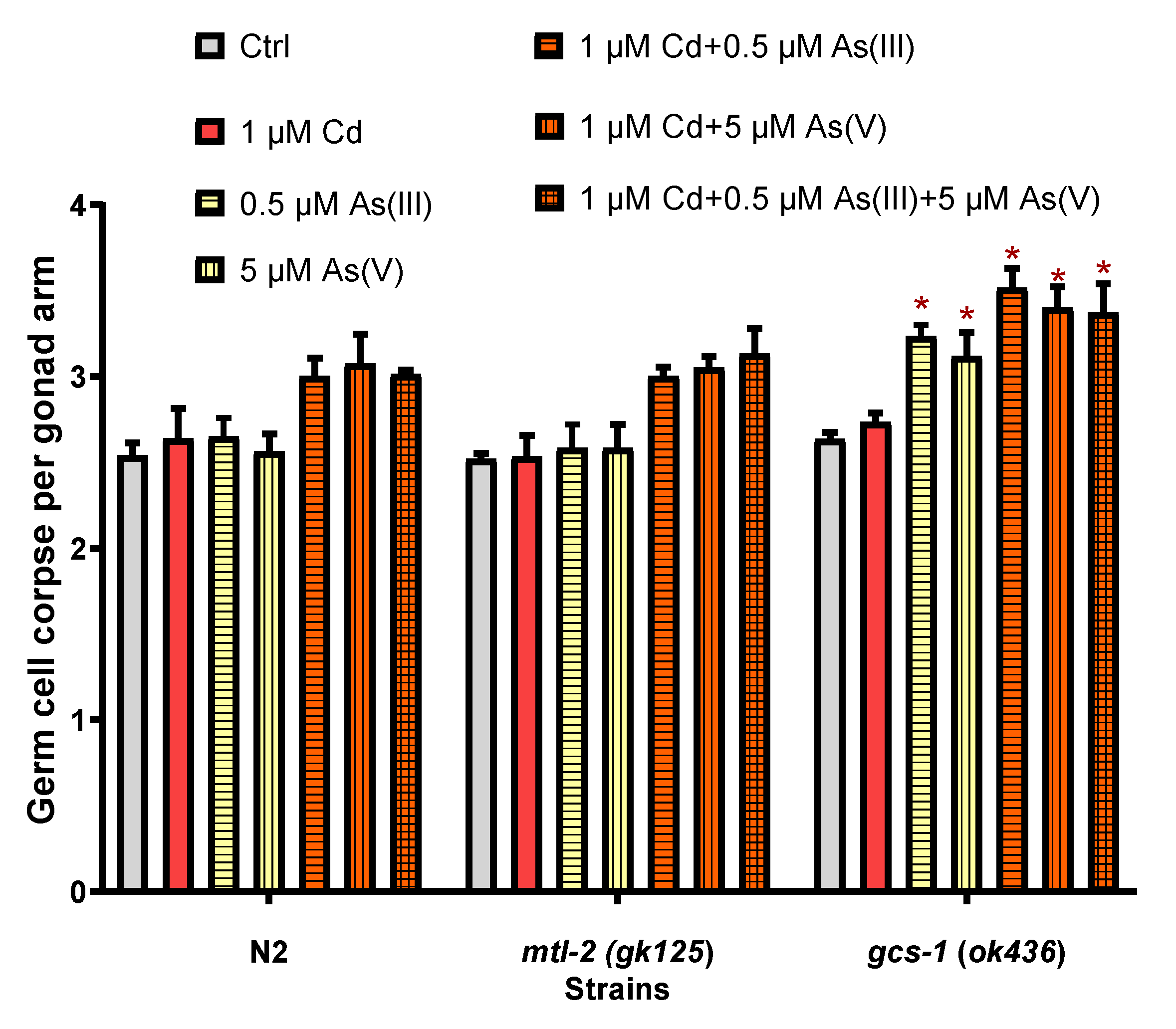
3.4. The Effects of mtl-2 and gcs-1 Gene on the Joint Toxicity of Cd and As
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hu, H.; Jin, Q.; Kavan, P. A study of heavy metal pollution in China: Current status, pollution-control policies and countermeasures. Sustainability 2014, 6, 5820–5838. [Google Scholar] [CrossRef] [Green Version]
- Fowler, B.A. Monitoring of human populations for early markers of cadmium toxicity: A review. Toxicol. Appl. Pharmacol. 2009, 238, 294–300. [Google Scholar] [CrossRef] [PubMed]
- Abdul, K.S.M.; Jayasinghe, S.S.; Chandana, E.P.; Jayasumana, C.; De Silva, P.M.C. Arsenic and human health effects: A review. Environ. Toxicol. Pharmacol. 2015, 40, 828–846. [Google Scholar] [CrossRef] [PubMed]
- Vardhan, K.H.; Kumar, P.S.; Panda, R.C. A review on heavy metal pollution, toxicity and remedial measures: Current trends and future perspectives. J. Mol. Liq. 2019, 290, 111197. [Google Scholar] [CrossRef]
- Genchi, G.; Carocci, A.; Lauria, G.; Sinicropi, M.S.; Catalano, A. Nickel: Human health and environmental toxicology. Int. J. Environ. Res. Public Health 2020, 17, 679. [Google Scholar] [CrossRef] [Green Version]
- Bernhoft, R.A. Cadmium toxicity and treatment. Sci. World J. 2013, 2013, 394652. [Google Scholar] [CrossRef]
- Cheng, C.-H.; Ma, H.-L.; Deng, Y.-Q.; Feng, J.; Jie, Y.-K.; Guo, Z.-X. Oxidative stress, cell cycle arrest, DNA damage and apoptosis in the mud crab (Scylla paramamosain) induced by cadmium exposure. Chemosphere 2021, 263, 128277. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.-K.; Thévenod, F. Cell organelles as targets of mammalian cadmium toxicity. Arch. Toxicol. 2020, 94, 1017–1049. [Google Scholar] [CrossRef] [Green Version]
- Ren, Y.; Shao, W.; Zuo, L.; Zhao, W.; Qin, H.; Hua, Y.; Lu, D.; Mi, C.; Zeng, S.; Zu, L. Mechanism of cadmium poisoning on testicular injury in mice. Oncol. Lett. 2019, 18, 1035–1042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Chu, Z.; Zhang, K.; Miao, G. Cadmium-induced serotonergic neuron and reproduction damages conferred lethality in the nematode Caenorhabditis elegans. Chemosphere 2018, 213, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Chen, J.; Wu, Y.; Wang, Q.; Li, H. Sublethal toxicity endpoints of heavy metals to the nematode Caenorhabditis elegans. PLoS ONE 2016, 11, e0148014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bovio, F.; Sciandrone, B.; Urani, C.; Fusi, P.; Forcella, M.; Regonesi, M.E. Superoxide dismutase 1 (SOD1) and cadmium: A three models approach to the comprehension of its neurotoxic effects. Neurotoxicology 2021, 84, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Roh, J.-Y.; Park, Y.-J.; Choi, J. A cadmium toxicity assay using stress responsive Caenorhabditis elegans mutant strains. Environ. Toxicol. Pharmacol. 2009, 28, 409–413. [Google Scholar] [CrossRef]
- Rahaman, M.S.; Rahman, M.M.; Mise, N.; Sikder, M.T.; Ichihara, G.; Uddin, M.K.; Kurasaki, M.; Ichihara, S. Environmental arsenic exposure and its contribution to human diseases, toxicity mechanism and management. Environ. Pollut. 2021, 289, 117940. [Google Scholar] [CrossRef] [PubMed]
- Moyson, S.; Vissenberg, K.; Fransen, E.; Blust, R.; Husson, S.J. Mixture effects of copper, cadmium, and zinc on mortality and behavior of Caenorhabditis elegans. Environ. Toxicol. Chem. 2018, 37, 145–159. [Google Scholar] [CrossRef] [PubMed]
- Sodhi, K.K.; Kumar, M.; Agrawal, P.K.; Singh, D.K. Perspectives on arsenic toxicity, carcinogenicity and its systemic remediation strategies. Environ. Technol. Innov. 2019, 16, 100462. [Google Scholar] [CrossRef]
- Jang, Y.; Somanna, Y.; Kim, H. Source, distribution, toxicity and remediation of arsenic in the environment—A review. Int. J. Appl. Environ. Sci. 2016, 11, 559–581. [Google Scholar]
- Rahman, M.M.; Chowdhury, U.K.; Mukherjee, S.C.; Mondal, B.K.; Paul, K.; Lodh, D.; Biswas, B.K.; Chanda, C.R.; Basu, G.K.; Saha, K.C. Chronic arsenic toxicity in Bangladesh and West Bengal, India review and commentary. J. Toxicol. Clin. Toxicol. 2001, 39, 683–700. [Google Scholar] [CrossRef]
- Khairul, I.; Wang, Q.Q.; Jiang, Y.H.; Wang, C.; Naranmandura, H. Metabolism, toxicity and anticancer activities of arsenic compounds. Oncotarget 2017, 8, 23905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, A.P.; Goel, R.K.; Kaur, T. Mechanisms pertaining to arsenic toxicity. Toxicol. Int. 2011, 18, 87. [Google Scholar] [PubMed] [Green Version]
- Rahman, Z.; Singh, V.P. The relative impact of toxic heavy metals (THMs) (arsenic (As), cadmium (Cd), chromium (Cr)(VI), mercury (Hg), and lead (Pb)) on the total environment: An overview. Environ. Monit. Assess. 2019, 191, 419. [Google Scholar] [CrossRef]
- Wang, S.; Teng, X.; Wang, Y.; Yu, H.Q.; Luo, X.; Xu, A.; Wu, L. Molecular control of arsenite-induced apoptosis in Caenorhabditis elegans: Roles of insulin-like growth factor-1 signaling pathway. Chemosphere 2014, 112, 248–255. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.-W.; How, C.M.; Liao, V.H.-C. Arsenite exposure accelerates aging process regulated by the transcription factor DAF-16/FOXO in Caenorhabditis elegans. Chemosphere 2016, 150, 632–638. [Google Scholar] [CrossRef]
- Yu, C.-W.; Liao, V.H.-C. Transgenerational reproductive effects of arsenite are associated with H3K4 dimethylation and SPR-5 downregulation in Caenorhabditis elegans. Environ. Sci. Technol. 2016, 50, 10673–10681. [Google Scholar] [CrossRef] [PubMed]
- Mikhailenko, A.V.; Ruban, D.A.; Ermolaev, V.A.; van Loon, A. Cadmium pollution in the tourism environment: A literature review. Geosci. J. 2020, 10, 242. [Google Scholar] [CrossRef]
- Hossain, S.; Latifa, G.A.; Prianqa, P.; Al Nayeem, A. Review of cadmium pollution in Bangladesh. J. Health Pollut. 2019, 9, 190913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, N.C.; Zheng, Y.J.; Xiao-Feng, H.E.; Xiao-Fei, L.I.; Zhang, X.X. Analysis of the Report on the national general survey of soil contamination. J. Agro Environ. Sci. 2017, 36, 1689–1692. [Google Scholar]
- Abbas, G.; Murtaza, B.; Bibi, I.; Shahid, M.; Niazi, N.K.; Khan, M.I.; Amjad, M.; Hussain, M.; Natasha. Arsenic uptake, toxicity, detoxification, and speciation in plants: Physiological, biochemical, and molecular aspects. Int. J. Environ. Res. Public Health 2018, 15, 59. [Google Scholar] [CrossRef] [Green Version]
- Carocci, A.; Catalano, A.; Lauria, G.; Sinicropi, M.S.; Genchi, G. Lead Toxicity, Antioxidant Defense and Environment. Rev. Environ. Contam. Toxicol. 2015, 238, 45–67. [Google Scholar]
- Mawia, A.M.; Hui, S.; Zhou, L.; Li, H.; Tabassum, J.; Lai, C.; Wang, J.; Shao, G.; Wei, X.; Tang, S.; et al. Inorganic Arsenic Toxicity and Alleviation Strategies in Rice. J. Hazard. Mater. 2020, 408, 124751. [Google Scholar] [CrossRef] [PubMed]
- Gu, Z.; de Silva, S.; Reichman, S.M. Arsenic concentrations and dietary exposure in rice-based infant food in Australia. Int. J. Environ. Res. Public Health 2020, 17, 415. [Google Scholar] [CrossRef] [Green Version]
- Medunić, G.; Fiket, Ž.; Ivanić, M. Arsenic contamination status in Europe, Australia, and other parts of the world. In Arsenic in Drinking Water and Food; Springer: Berlin/Heidelberg, Germany, 2020; pp. 183–233. [Google Scholar]
- Shi, Y.-L.; Chen, W.-Q.; Wu, S.-L.; Zhu, Y.-G. Anthropogenic cycles of arsenic in mainland China: 1990–2010. Environ. Sci. Technol. 2017, 51, 1670–1678. [Google Scholar] [CrossRef] [PubMed]
- Zimeng, L.; Xiaoqian, L.; ZHANG, W.; Yining, J.; Siyu, S.; Sujing, W.; Qirui, A.; Pengyang, L.; ZHENG, N. Research progress on the effects of heavy metal compound pollution on organisms. Environ. Chem. 2021, 40, 3331–3343. [Google Scholar]
- Martin, O.; Scholze, M.; Ermler, S.; McPhie, J.; Bopp, S.K.; Kienzler, A.; Parissis, N.; Kortenkamp, A. Ten years of research on synergisms and antagonisms in chemical mixtures: A systematic review and quantitative reappraisal of mixture studies. Environ. Int. 2021, 146, 106206. [Google Scholar] [CrossRef] [PubMed]
- Rai, A.; Maurya, S.K.; Khare, P.; Srivastava, A.; Bandyopadhyay, S. Characterization of developmental neurotoxicity of As, Cd, and Pb mixture: Synergistic action of metal mixture in glial and neuronal functions. Toxicol. Sci. 2010, 118, 586–601. [Google Scholar] [CrossRef] [PubMed]
- Adedara, I.A.; Adegbosin, A.N.; Abiola, M.A.; Odunewu, A.A.; Owoeye, O.; Owumi, S.E.; Farombi, E.O. Neurobehavioural and biochemical responses associated with exposure to binary waterborne mixtures of zinc and nickel in rats. Environ. Toxicol. Pharmacol. 2020, 73, 103294. [Google Scholar] [CrossRef]
- Bloom, M.S.; Parsons, P.J.; Steuerwald, A.J.; Schisterman, E.F.; Browne, R.W.; Kim, K.; Coccaro, G.A.; Conti, G.C.; Narayan, N.; Fujimoto, V.Y. Toxic trace metals and human oocytes during in vitro fertilization (IVF). Reprod. Toxicol. 2010, 29, 298–305. [Google Scholar] [CrossRef] [Green Version]
- Lanier, C.; Bernard, F.; Dumez, S.; Leclercq-Dransart, J.; Lemiere, S.; Vandenbulcke, F.; Nesslany, F.; Platel, A.; Devred, I.; Hayet, A. Combined toxic effects and DNA damage to two plant species exposed to binary metal mixtures (Cd/Pb). Ecotoxicol. Environ. Saf. 2019, 167, 278–287. [Google Scholar] [CrossRef]
- Meng, Q.J. Toxic Effects of Compound Pollution with Heavy Metals on the Growth of Wheat Seedlings. J. Anhui Agric. Sci. 2008, 36, 122. [Google Scholar]
- Corsi, A.K.; Wightman, B.; Chalfie, M. A transparent window into biology: A primer on Caenorhabditis elegans. Genetics 2015, 200, 387–407. [Google Scholar] [CrossRef] [Green Version]
- Hunt, P.R. The C. elegans model in toxicity testing. J. Appl. Toxicol. 2017, 37, 50–59. [Google Scholar] [CrossRef]
- Shen, P.; Yue, Y.; Park, Y. A living model for obesity and aging research: Caenorhabditis elegans. Crit. Rev. Food Sci. Nutr. 2018, 58, 741–754. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, J.; Gorishek, E.; Mach, P.; Sturtevant, D.; Ladage, M.; Suzuki, N.; Padilla, P.; Mittler, R.; Chapman, K.; Verbeck, G. Evaluation of a custom single Peltier-cooled ablation cell for elemental imaging of biological samples in laser ablation-inductively coupled plasma-mass spectrometry (LA-ICP-MS). J. Anal. At. Spectrom. 2016, 31, 1030–1033. [Google Scholar] [CrossRef]
- Crone, B.; Aschner, M.; Schwerdtle, T.; Karst, U.; Bornhorst, J. Elemental bioimaging of Cisplatin in Caenorhabditis elegans by LA-ICP-MS. Metallomics 2015, 7, 1189–1195. [Google Scholar] [CrossRef] [Green Version]
- Brenner, S.J.G. The genetics of Caenorhabditis elegans. Genetics 1974, 77, 71–94. [Google Scholar] [CrossRef] [PubMed]
- Gartner, A.; MacQueen, A.J.; Villeneuve, A.M. Methods for analyzing checkpoint responses in Caenorhabditis elegans. In Checkpoint Controls and Cancer; Springer: Berlin, Germany, 2004; pp. 257–274. [Google Scholar]
- Ruan, Q.L.; Ju, J.J.; Li, Y.H.; Li, X.B.; Liu, R.; Liang, G.Y.; Zhang, J.; Pu, Y.P.; Wang, D.Y.; Yin, L.H. Chlorpyrifos exposure reduces reproductive capacity owing to a damaging effect on gametogenesis in the nematode Caenorhabditis elegans. J. Appl. Toxicol. 2012, 32, 527–535. [Google Scholar] [CrossRef] [PubMed]
- Du, H.; Wang, M.; Wang, L.; Dai, H.; Wang, M.; Hong, W.; Nie, X.; Wu, L.; Xu, A. Reproductive Toxicity of Endosulfan: Implication from Germ Cell Apoptosis Modulated by Mitochondrial Dysfunction and Genotoxic Response Genes in Caenorhabditis elegans. Toxicol. Sci. Off. J. Soc. Toxicol. 2015, 145, 118–127. [Google Scholar] [CrossRef] [Green Version]
- Dai, H.; Liu, Y.; Wang, J.; Nie, Y.; Sun, Y.; Wang, M.; Wang, D.; Yang, Z.; Cheng, L.; Wang, J. Graphene oxide antagonizes the toxic response to arsenic via activation of protective autophagy and suppression of the arsenic-binding protein LEC-1 in Caenorhabditis elegans. Environ. Sci. Nano 2018, 5, 1711–1728. [Google Scholar] [CrossRef]
- Xin, X.W.; Zhang, D.K. Comparative Analysis of Heavy Metal Residue Indicator in Domestic and Foreign Food Standards. China Stand. 2018, 8, 124–128. [Google Scholar]
- Sinha, K.A.; Keen, J.K.; Ogun, S.A.; Holder, A.A. Comparison of two members of a multigene family coding for high-molecular mass rhoptry proteins of Plasmodium yoelii. Mol. Biochem. Parasitol. 1996, 76, 329. [Google Scholar] [CrossRef]
- Singh, N.; Kumar, D.; Sahu, A.P. Arsenic in the environment: Effects on human health and possible prevention. J. Environ. Biol. 2007, 28, 359. [Google Scholar]
- Berman, J.R.; Kenyon, C. Germ-cell loss extends C. elegans life span through regulation of DAF-16 by kri-1 and lipophilic-hormone signaling. Cell 2006, 124, 1055–1068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eisenmann, D. WormBook: The Online Review of C. Elegans Biology. Research Community, Wormbook. Available online: http://www.wormbook.org (accessed on 21 March 2007).
- Chauhan, V.M.; Orsi, G.; Brown, A.; Pritchard, D.I.; Aylott, J.W. Mapping the Pharyngeal and Intestinal pH of Caenorhabditis elegans and Real-Time Luminal pH Oscillations Using Extended Dynamic Range pH-Sensitive Nanosensors. ACS Nano 2013, 7, 5577–5587. [Google Scholar] [CrossRef] [PubMed]
- Pei, B.; Wang, S.; Guo, X.; Wang, J.; Yang, G.; Hang, H.; Wu, L. Arsenite-induced germline apoptosis through a MAPK-dependent, p53-independent pathway in Caenorhabditis elegans. Chem. Res. Toxicol. 2008, 21, 1530–1535. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Luo, X.; Wu, L. Assessment of Caenorhabditis Elegans Germline Cell Cycle Arrest and Apoptosis for Heavy Metals Toxicity Testing. In Proceedings of the 2009 3rd International Conference on Bioinformatics and Biomedical Engineering, Beijing, China, 11–13 June 2009; pp. 1–4. [Google Scholar]
- Wang, S.; Tang, M.; Pei, B.; Xiao, X.; Wang, J.; Hang, H.; Wu, L. Cadmium-induced germline apoptosis in Caenorhabditis elegans: The roles of HUS1, p53, and MAPK signaling pathways. Toxicol. Sci. 2008, 102, 345–351. [Google Scholar] [CrossRef]
- Sun, Y.; Li, Z.; Guo, B.; Chu, G.; Wei, C.; Liang, Y. Arsenic mitigates cadmium toxicity in rice seedlings. Environ. Exp. Bot. 2008, 64, 264–270. [Google Scholar] [CrossRef]
- Cao, Q.; Hu, Q.H.; Khan, S.; Wang, Z.J.; Lin, A.J. Wheat phytotoxicity from arsenic and cadmium separately and together in solution culture and in a calcareous soil—ScienceDirect. J. Hazard. Mater. 2007, 148, 377–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Driessnack, M.K.; Jamwal, A.; Niyogi, S. Effects of chronic exposure to waterborne copper and nickel in binary mixture on tissue-specific metal accumulation and reproduction in fathead minnow (Pimephales promelas). Chemosphere 2017, 185, 964–974. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, S.; Shan, X.Q.; Christie, P. Combined toxicity of cadmium and arsenate to wheat seedlings and plant uptake and antioxidative enzyme responses to cadmium and arsenate co-contamination. Ecotoxicol. Environ. Saf. 2007, 68, 305–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tremaroli, V.; Bäckhed, F. Keep It in the Family Professionals Are Handing Back Power to Parents over the Futureof Their Children, However Traumatised Relationships May Be. Glenda Cooper Reports. Nature 2012, 489, 242–249. [Google Scholar] [CrossRef]
- Wang, X.; Wang, Q.; Yu, L. Heavy Metal Contamination and Its Effect on Organism in Soil: A Review. Chin. Agric. Sci. Bull. 2017, 33, 86–92. [Google Scholar]
- Száková, J.; Zídek, V.; Miholová, D. Influence of elevated content of cadmium and arsenic in diet containing feeding yeast on organisms of rats. Czech J. Anim. Sci. 2009, 54, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Ali, A.S.; Bharwana, S.A.; Ahmad, R. Effect of different heavy metal pollution on fish. Res. J. Chem. Environ. Sci. 2014, 2, 74–79. [Google Scholar]
- Große Brinkhaus, S.; Bornhorst, J.; Chakraborty, S.; Wehe, C.A.; Niehaus, R.; Reifschneider, O.; Aschner, M.; Karst, U. Elemental bioimaging of manganese uptake in C. elegans. Metallomics 2014, 6, 617–621. [Google Scholar] [CrossRef]
- Nutt, L.K.; Gogvadze, V.; Uthaisang, W.; Mirnikjoo, B.; McConkey, D.J.; Orrenius, S. Research paper indirect effects of Bax and Bak initiate the mitochondrial alterations that lead to cytochrome c release during arsenic trioxide-induced apoptosis. Cancer Biol. 2005, 4, 459–467. [Google Scholar]
- Wang, S.; Zhao, Y.; Wu, L.; Tang, M.; Su, C.; Hei, T.K.; Yu, Z. Induction of germline cell cycle arrest and apoptosis by sodium arsenite in Caenorhabditis elegans. Chem. Res. Toxicol. 2007, 20, 181–186. [Google Scholar] [CrossRef]
- Arun, K.; Madhavan, A.; Sindhu, R.; Emmanual, S.; Binod, P.; Pugazhendhi, A.; Sirohi, R.; Reshmy, R.; Awasthi, M.K.; Gnansounou, E. Probiotics and gut microbiome—Prospects and challenges in remediating heavy metal toxicity. J. Hazard. Mater. 2021, 420, 126676. [Google Scholar] [CrossRef] [PubMed]
- Breton, J.; Massart, S.; Vandamme, P.; De Brandt, E.; Pot, B.; Foligné, B. Ecotoxicology inside the gut: Impact of heavy metals on the mouse microbiome. BMC Pharmacol. 2013, 14, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buchet, J.-P.; Lauwerys, R.; Roels, H. Comparison of the urinary excretion of arsenic metabolites after a single oral dose of sodium arsenite, monomethylarsonate, or dimethylarsinate in man. Int. Arch. Occup. Environ. Health 1981, 48, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Meng, Z.Y.; Lin, H.; Wang, A.Q. Comparative Study on the Inhibition of Arsenic and Arsenate to Brain Acetylcholinesterase from Different Fish Species. J. Ocean Univ. Qingdao 2005, 35, 471–474. [Google Scholar]
- Suhendrayatna; Ohki, A.; Maeda, S. Arsenic accumulation, transformation and tolerance on freshwater Daphnia magna. Toxicol. Environ. Chem. 1999, 72, 1–11. [Google Scholar] [CrossRef]
- Swain, S.; Keusekotten, K.; Baumeister, R.; Stürzenbaum, S.C. elegans metallothioneins: New insights into the phenotypic effects of cadmium toxicosis. J. Mol. Biol. 2004, 341, 951–959. [Google Scholar] [CrossRef] [PubMed]
- Tinkov, A.A.; Gritsenko, V.A.; Skalnaya, M.G.; Cherkasov, S.V.; Aaseth, J.; Skalny, A.V. Gut as a target for cadmium toxicity. Environ. Pollut. 2018, 235, 429–434. [Google Scholar] [CrossRef]
- Wang, J.; Nie, Y.; Dai, H.; Wang, M.; Cheng, L.; Yang, Z.; Chen, S.; Zhao, G.; Wu, L.; Guang, S. Parental exposure to TiO 2 NPs promotes the multigenerational reproductive toxicity of Cd in Caenorhabditis elegans via bioaccumulation of Cd in germ cells. Environ. Sci. Nano 2019, 6, 1332–1342. [Google Scholar] [CrossRef]
- Scott, N.; Hatlelid, K.M.; Mackenzie, N.E.; Carter, D.E. Reactions of arsenic(III) and arsenic(V) species with glutathione. Chem. Res. Toxicol. 1993, 6, 102–106. [Google Scholar] [CrossRef] [PubMed]
- Ochi, T. Arsenic compound-induced increases in glutathione levels in cultured Chinese hamster V79 cells and mechanisms associated with changes in γ-glutamylcysteine synthetase activity, cystine uptake and utilization of cysteine. Arch. Toxicol. 1997, 71, 730–740. [Google Scholar] [CrossRef] [PubMed]
- Urban, N.; Tsitsipatis, D.; Hausig, F.; Kreuzer, K.; Erler, K.; Stein, V.; Ristow, M.; Steinbrenner, H.; Klotz, L.-O. Non-linear impact of glutathione depletion on C. elegans life span and stress resistance. Redox Biol. 2017, 11, 502–515. [Google Scholar] [CrossRef]
- Huang, H.; Huang, C.; Wu, D.; Jinn, C.; Jan, K. Glutathione as a cellular defence against arsenite toxicity in cultured Chinese hamster ovary cells. Toxicology 1993, 79, 195–204. [Google Scholar] [CrossRef]
- Liao, V.H.-C.; Yu, C.-W. Caenorhabditis elegans gcs-1 confers resistance to arsenic-induced oxidative stress. Biometals 2005, 18, 519–528. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pei, C.; Sun, L.; Zhao, Y.; Ni, S.; Nie, Y.; Wu, L.; Xu, A. Enhanced Uptake of Arsenic Induces Increased Toxicity with Cadmium at Non-Toxic Concentrations on Caenorhabditis elegans. Toxics 2022, 10, 133. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics10030133
Pei C, Sun L, Zhao Y, Ni S, Nie Y, Wu L, Xu A. Enhanced Uptake of Arsenic Induces Increased Toxicity with Cadmium at Non-Toxic Concentrations on Caenorhabditis elegans. Toxics. 2022; 10(3):133. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics10030133
Chicago/Turabian StylePei, Chengcheng, Lingyan Sun, Yanan Zhao, Shenyao Ni, Yaguang Nie, Lijun Wu, and An Xu. 2022. "Enhanced Uptake of Arsenic Induces Increased Toxicity with Cadmium at Non-Toxic Concentrations on Caenorhabditis elegans" Toxics 10, no. 3: 133. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics10030133