
Study of Ultrastructural Abnormalities in the Renal Cells of Cyprinus carpio Induced by Toxicants

Abstract
:1. Introduction
2. Materials and Methods
2.1. Test Chemicals, Selection and Dosage of the Insecticides
2.2. Experimental Design
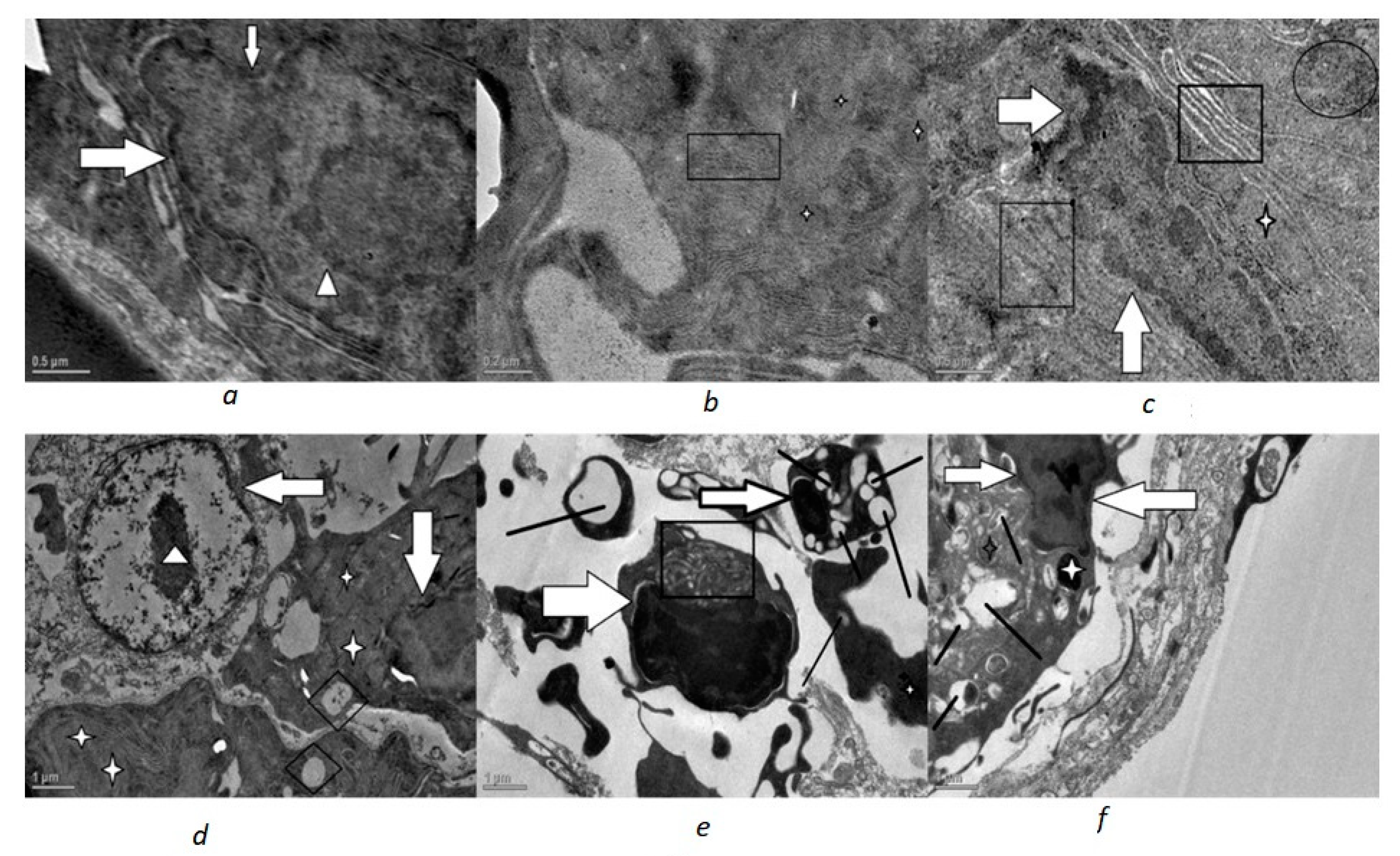
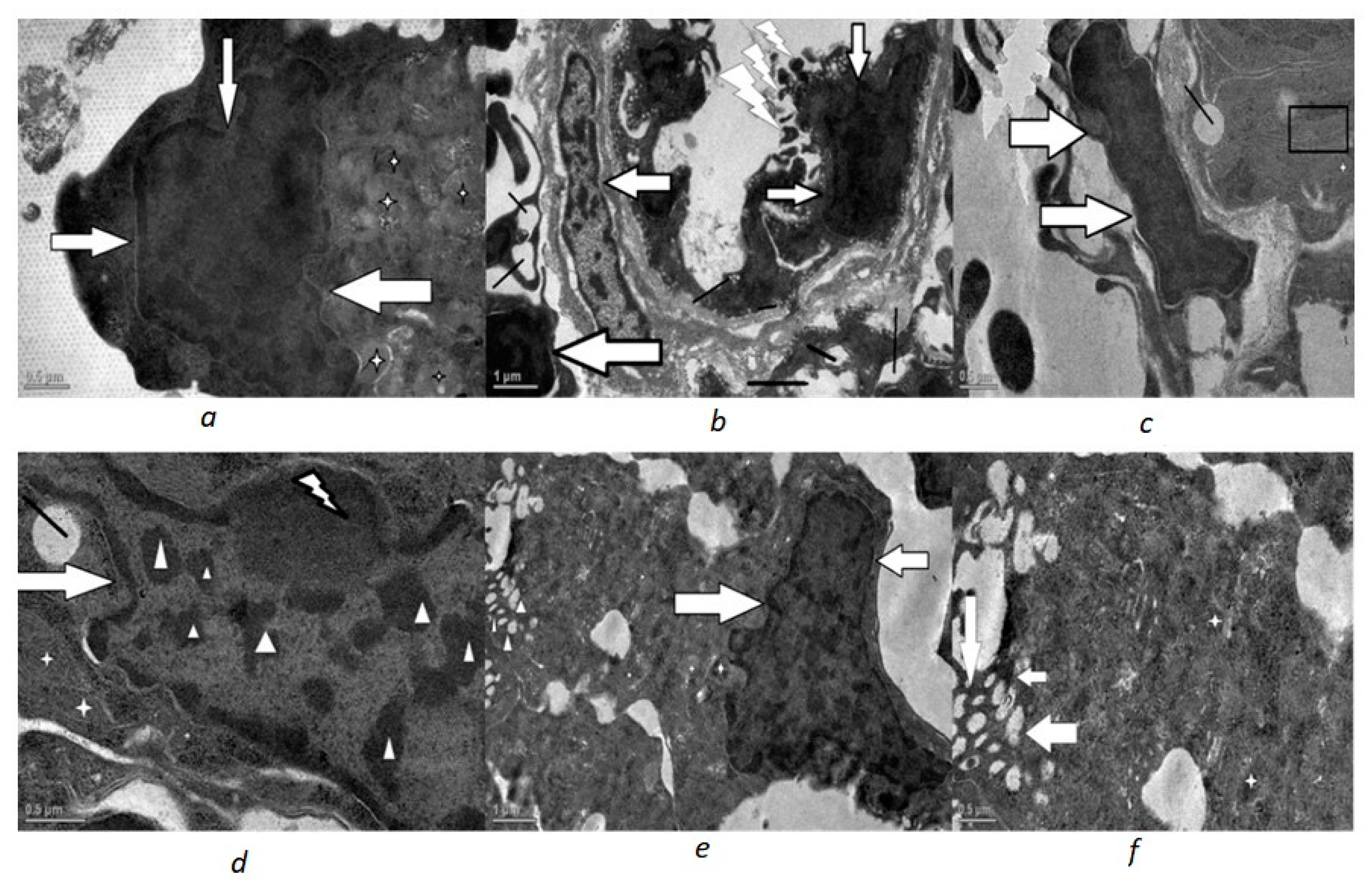
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kafula, Y.A.; Philippe, C.; Pinceel, T.; Munishi, L.K.; Moyo, F.; Vanschoenwinkel, B.; Brendonk, L.; Thore, E.S.J. Pesticide sensitivity of Nothobranchius neumanni, a temporary pond predator with a non generic life history. Chemosphere 2022, 291, 132823. [Google Scholar] [CrossRef] [PubMed]
- Insecticide Resistance Action Committee. 2017. Available online: http://www.irac-online.org/documents/moa-classification/ (accessed on 12 May 2019).
- Banae, M.; Mirvaghefei, A.R.; Rafei, G.R.; Majazi, A.B. Effect of sub lethal Diazinon concentrations on blood plasma biochemistry. Int. J. Environ. Res. 2008, 2, 189–198. [Google Scholar]
- Nwani, C.D.; Lakra, W.S.; Nagpureb, N.S.; Kumar, R.; Kushwaha, B.; Srivastava, S.K. Mutagenic and genotoxic effects of carbosulfan in freshwater fish Channa punctatus (Bloch) using micronucleus assay and alkaline single-cell gel electrophoresis. Food Chem. Toxicol. 2010, 48, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Stentiford, G.D.; Longshaw, M.; Lyons, B.P.; Jones, G.; Green, M.; Feist, S.W. Histopathological biomarkers in estuarine fish species for the assessment of biological effects of contaminants. Mar. Environ. Res. 2003, 55, 137–159. [Google Scholar] [CrossRef]
- Dar, S.A.; Yousuf, A.R.; Balkhi, M.H. An introduction about genotoxicology methods as tools for monitoring aquatic ecosystem: Present status and future perspectives. Fish Aquac. J. 2016, 7, 158. [Google Scholar]
- Saquib, Q.; Attia, S.M.; Siddiqui, M.A.; Aboul-Soud, M.A.M.; Al-Khedhairy, A.A.; Giesy, J.P.; Musarrat, J. Phorate-induced oxidative stress, DNA damage and transcriptional activation of p53and caspase genes in male Wistar rats. Toxicol. Appl. Pharmacol. 2012, 259, 54–65. [Google Scholar] [CrossRef]
- Lakshmaiah, G. Toxic impact of lethal and sublethal concentrations of Phorate on the total protein levels in different tissues of common carp Cyprinus carpio. Int. J. Pharma Bio Sci. 2014, 5, 422–428. [Google Scholar]
- Abhilash, P.C.; Singh, N. Pesticide use and application: An Indian scenario. J. Hazard. Mater. 2009, 165, 1–12. [Google Scholar] [CrossRef]
- Singh, R.N.; Pandey, R.K.; Singh, N.N.; Das, V.K. Acute toxicity and behavioral responses of common carp Cyprinus carpio (linn.) to an organophosphate (Dimethoate). World J. Zool. 2009, 4, 70–75. [Google Scholar]
- Singh, R.N.; Yadava, C.L.; Singh, K. Dimethoate induced alterations in carbohydrate metabolism of common carp, Cyprinus carpio (Linn.). Indian J. Sci. Res. 2017, 15, 175–180. [Google Scholar]
- Qayoom, I.; Shah, F.A.; Mukhtar, M.; Balkhi, M.H.; Bhat, F.A.; Bhat, B.A. Dimethoate induced behavioural changes in juveniles of Cyprinus carpio var. communis under temperate conditions of Kashmir, India. Sci. World J. 2016, 2016, 4726126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lakshmaiah, G.; Indira, P. Investigation of acute toxicity of phorate (an organophosphate) on fresh water fish Cyprinus carpio. Int. J. Pharma Bio Sci. 2014, 5, 104–110. [Google Scholar]
- Lakshmaiah, G. Effect of phorate on histopathological changes in the kidney of common carp Cyprinus carpio exposed to lethal concentrations. Int. J. Zool. Stud. 2016, 1, 25–28. [Google Scholar]
- Patel, S.K.M.; Indulkar, S.T.; Shaikh, A.L.A.H.; Pai, R. Lethal and sublethal toxicity of an organophosphate pesticide, Phorate 10G on fingerlings of Tilapia sp. Int. J. Pure App. Biosci. 2017, 5, 1153–1157. [Google Scholar]
- Lakshmaiah, G. Brain histopathology of the fish Cyprinus carpio exposed to lethal concentrations of an organophosphate insecticide phorate. Int. J. Adv. Res. Dev. 2017, 2, 668–672. [Google Scholar]
- Lakshmaiah, G. Effect of phorate lethal concentrations on the histological aspects of liver in common carp Cyprinus carpio (Linnaeus, 1758). Int. J. Chem. Stud. 2016, 4, 6–9. [Google Scholar]
- Pandey, R.K.; Singh, R.N.; Singh, S.; Singh, N.N.; Das, V.K. Acute toxicity bioassay of dimethoate on freshwater air breathing catfish, Heteropneustes fossilis (Bloch). J. Environ. Biol. 2009, 30, 437–440. [Google Scholar]
- Dogan, D.; Can, C. Hematological, biochemical, and behavioral responses of Oncorhynchus mykiss to dimethoate. Fish Physiol. Biochem. 2011, 37, 951–958. [Google Scholar] [CrossRef]
- Ganeshwade, R.M. Biochemical changes induced by dimethoate (Rogor 30% EC) in the gills of fresh water fish (Puntius ticto (Hamilton). J. Ecol. Nat. Environ. 2012, 4, 181–185. [Google Scholar] [CrossRef]
- Singh, R.N. Histopathological alterations in the kidney of Cyprinus carpio after exposure to dimethoate (EC 30%). Indian J. Sci. Res. 2012, 3, 127–131. [Google Scholar]
- Binukumari, S.; Vasanthi, J. The toxic effect of pesticide dimethoate 30% EC on the protein metabolism of the fresh water fish, Labeo rohita. Int. J. Curr. Microbiol. App. Sci. 2013, 2, 79–82. [Google Scholar]
- Singh, R.N. Effects of dimethoate (30%EC) an organophosphate pesticide on liver of common carp Cyprinus carpio. J. Environ. Biol. 2013, 34, 657–661. [Google Scholar] [PubMed]
- Singh, R.N. Effects of dimethoate (EC 30%) on gill morphology, oxygen consumption and serum electrolyte levels of common carp, Cyprinus carpio (Linn). Int. J. Sci. Res. Environ. Sci. 2014, 2, 192–198. [Google Scholar] [CrossRef]
- Singh, R.N. Testicular toxicity of dimethoate in common carp Cyprinus carpio (linn). Indian J. Sci. Res. 2017, 15, 152–156. [Google Scholar]
- Thore, E.S.J.; Philippe, C.; Brendonck, L.; Pinceel, T. Towards improved fish tests in ecotoxicology—Efficient chronic and multi-generational testing with the killifish Nothobranchius furzeri. Chemosphere 2021, 273, 129697. [Google Scholar] [CrossRef] [PubMed]
- Norton, W.N. Acute Exposure of Medaka to Carcinogens: An Ultrastructural, Cytochemical and Morphometric Analysis of Liver and Kidney; Southeastern Louisiana University Hammond: Hammond, LA, USA, 1992. [Google Scholar]
- Anvarifar, H.; Amirkolaie, A.K.; Jalali, A.M.; Miandare, H.K.; Sayed, A.H.; Ucuncu, S.I.; Ouraji, H.; Ceci, M.; Romano, N. Environmental pollution and toxic substances: Cellular apoptosis as a key parameter in a sensible model like fish. Aquat. Toxicol. 2018, 204, 144–159. [Google Scholar] [CrossRef]
- Rocha, C.A.M.D.; Santos, R.A.D.; Bahia, M.D.O.; Cunha, L.A.D.A.; Ribeiro, H.F.; Burbano, R.M.R. The micronucleus assay in fish species as an important tool for xenobiotic exposure risk assessment—A brief review and an example using Neotropical fish exposed to methylmercury. Rev. Fish. Sci. 2009, 17, 478–484. [Google Scholar] [CrossRef]
- Al-Sabti, K.; Metcalfe, C.D. Fish micronuclei for assessing genotoxicity in water. Mutat. Res. 1995, 343, 121–135. [Google Scholar] [CrossRef]
- APHA; AWWA; WPCF. Standard Method for Examination of Water and Wastewater, 20th ed.; APHA: Washington, DC, USA, 1998. [Google Scholar]
- Philpott, N.J.; Turner, A.J.C.; Scopes, J.; Westby, M.; Marsh, J.C.W.; Gordon-Smith, E.C.; Dalgleish, A.G.; Gibson, F.M. The use of 7-Amino actinomycin D in identifying apoptosis: Simplicity of use and broad spectrum of application compared with other techniques. Blood 1996, 87, 2244–2251. [Google Scholar] [CrossRef]
- Fontanetti, C.S.; Christofoletti, C.A.; Pinheiro, T.G.; Souza, T.S.; Pedro-Escher, J. Microscopy as a tool in toxicological evaluations. Microsc. Sci. Technol. Appl. Educ. 2010, 2, 1001–1007. [Google Scholar]
- Singla, N. An attempt to study ultrastructural changes in kidney of Ctenopharyngodon idellus (Cuvier and Valenciennes) induced by monocrotophos using TEM. Scitech J. 2015, 2, 28–33. [Google Scholar]
- Chavan, V.R. Lead induced morphological and biochemical fluctuations in kidney of Cirrhinus mrigala. Res. J. Life Sci. Bioinform. Pharm. Chem. Sci. 2016, 2, 1–11. [Google Scholar]
- Rajini, A.; Revathy, K.; Selvam, G. Histopathological changes in tissues of Danio rerio exposed to sub lethal concentration of combination pesticide. Indian J. Sci. Technol. 2015, 8, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Khatun, N.; Rahman, T.; Mahanta, R. Histopathological studies of chlorpyrifos toxicity in Catfish. Glob. J. Med. Res. 2016, 6, 48–54. [Google Scholar]
- Besirovic, H.; Alic, A.; Prasovic, S.; Drommer, W. Histopathological effects of chronic exposure to cadmium and zinc on kidneys and gills of brown trout (Salmo trutta m. fario). Turk. J. Fish. Aquat. Sci. 2010, 10, 255–262. [Google Scholar] [CrossRef]
- Hadi, A.A.; Alwan, S.F. Histopathological changes in gills, liver and kidney of fresh water fish, Tilapia zillii, exposed to aluminum. Int. J. Pharm. Life Sci. 2012, 3, 2071–2081. [Google Scholar]
- Samanta, P.; Pa, S.; Mukherjee, A.K.; Senapati, T.; Ghosh, A.R. Cytopathological effects of Almix herbicide on gill, liver and kidney of Oreochromis niloticus under field and laboratory conditions. Toxicol Open Access 2016, 2, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Massar, B.; Dey, S.; Dutta, K. Structural changes in kidneys of common carp (Cyprinus carpio L.) inhabiting a polluted reservoir, Umiam in Meghalaya, India. J. Adv. Microsc. Res. 2014, 9, 105–109. [Google Scholar] [CrossRef]
- Marlasca, M.J.; Valles, B.; Riva, M.C.; Crespo, S. Sublethal effects of synthetic dyes on rainbow trout Oncorhynchus mykiss: A light and electron microscope study. Dis. Aquat. Org. 1992, 12, 103–112. [Google Scholar] [CrossRef]
- Spazier, E.; Storch, V.; Braunbeck, T. Cytopathology of spleen in eel Anguilla anguilla exposed to a chemical spill in the Rhine River. Dis. Aquat. Org. 1992, 14, 1–22. [Google Scholar] [CrossRef]
- Godar, D.E.; Miller, S.A.; Thomas, D.P. Immediate and delayed apoptotic cell death mechanisms: UVA versus UVB and UVC radiation. Cell Death Differ. 1994, 1, 59–66. [Google Scholar] [PubMed]
- Anglade, P.; Vyas, S.; Hirsch, E.C.; Agid, Y. Apoptosis in dopaminergic neurons of the human substantia nigra during normal aging. Histol. Histopathol. 1997, 12, 603–610. [Google Scholar] [PubMed]
- Hagar, H.H.; Azza, H. A biochemical, histochemical, and ultrastructural evaluation of the effect of dimethoate intoxication on rat pancreas. Toxicol. Lett. 2002, 133, 161–170. [Google Scholar] [CrossRef]
- Bajruskovic, G.R.; Anoelija, B.S.; Slobodan, A.M.; Snezana, J.C.; Slavica, F.K. The ultrastructural investigation of mitochondria in B-CLL cells during apoptosis. Arch. Oncol. 2004, 12, 139–141. [Google Scholar] [CrossRef] [Green Version]
- Domingos, F.X.V.; Assis, H.C.S.; Silva, M.D.; Damian, R.C.; Almeida, A.I.M.; Cestari, M.M.; Randi, M.A.F.; Ribeiro, C.A.O. Anthropic impact evaluation of two Brazilian estuaries trough biomarkers in fish. J. Braz. Soc. Ecotoxicol. 2009, 4, 21–30. [Google Scholar] [CrossRef]
- Mossalam, H.H.; Abd-El Aty, O.A.; Morgan, E.N.; Youssaf, S.M.S.; Mackawy, A.M.H. Biochemical and ultra structural studies of the antioxidant effect of aqueous extract of Hibiscus sabdariffa on the nephrotoxicity induced by organophosphorous pesticide (malathion) on the adult albino rats. Life Sci. J. 2011, 8, 561–574. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S.NO | NAME | FORMULA | CAS | REG. NO | NATURE |
|---|---|---|---|---|---|
| 1 | Phorate | C7H17O2PS3 | O, O-diethyl s-[(ethyl thio)methyl] phosphorodithioate | 298-02-02 | Highly toxic systemic insecticide used as granules. It has fumigant action. |
| 2 | Dimethoate | C5H12O3PS2 | O, O-diethyl s-[2-(methylamino)-2-oxyethyl]phosphorodithioate | 60-51-5 | It is acutely toxic, has possible links to cancer. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nazir, S.; Ali, M.N.; Tantray, J.A.; Baba, I.A.; Jan, A.; Popescu, S.M.; Paray, B.A.; Gulnaz, A. Study of Ultrastructural Abnormalities in the Renal Cells of Cyprinus carpio Induced by Toxicants. Toxics 2022, 10, 177. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics10040177
Nazir S, Ali MN, Tantray JA, Baba IA, Jan A, Popescu SM, Paray BA, Gulnaz A. Study of Ultrastructural Abnormalities in the Renal Cells of Cyprinus carpio Induced by Toxicants. Toxics. 2022; 10(4):177. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics10040177
Chicago/Turabian StyleNazir, Sumayya, Md. Niamat Ali, Javeed Ahmad Tantray, Irfan Akram Baba, Arizo Jan, Simona Mariana Popescu, Bilal Ahamad Paray, and Aneela Gulnaz. 2022. "Study of Ultrastructural Abnormalities in the Renal Cells of Cyprinus carpio Induced by Toxicants" Toxics 10, no. 4: 177. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics10040177