
Foliar Application of Wood Distillate Alleviates Ozone-Induced Damage in Lettuce (Lactuca sativa L.)



1
Department of Chemistry, Life Sciences, and Environmental Sustainability, University of Parma, Parco Area delle Scienze 11/a, 43124 Parma, Italy
2
Department of Life Sciences, University of Siena, Via PA Mattioli 4, I-53100 Siena, Italy
3
BAT Center-Interuniversity Center for Studies on Bioinspired Agro-Environmental Technology, University of Naples ‘Federico II’, 80138 Napoli, Italy
*
Author to whom correspondence should be addressed.
Toxics 2022, 10(4), 178; https://0-doi-org.brum.beds.ac.uk/10.3390/toxics10040178
Submission received: 15 March 2022
/
Revised: 2 April 2022
/
Accepted: 3 April 2022
/
Published: 5 April 2022
(This article belongs to the Topic Sustainable Environmental Technologies)
Abstract
:This study examined whether foliar applications of wood distillate (WD) have a protective effect on photosynthesis and the antioxidant power of lettuce when exposed to an ecologically relevant O3 concentration. Seedlings of lettuce (Lactuca sativa L.) were fumigated daily with 60 ppb of O3 for 30 days, five hours per day. Once per week, 50% of the fumigated plants were treated with foliar applications of 0.2% WD, while control plants were treated with water. The results clearly showed the ability of WD to protect lettuce plants from ozone-induced damage. Specifically, WD-treated plants exhibited lower damage to the photosynthetic machinery, assessed through a series of chlorophyll fluorescence parameters, a higher chlorophyll content, higher antioxidant power, as well as antioxidant molecules, i.e., caffeic acid and quercetin, and higher biomass. Counteracting the overproduction of ozone-generated reactive oxygen species (ROS) is speculated to be the main mechanism by which WD protects the plant from ozone-induced damage.
1. Introduction
Ozone (O3) is a global strongly oxidizing pollutant, present at ground level following the interaction of UV light with anthropogenic gases, such as carbon monoxide (CO), nitrogen oxides (NOX), methane (CH4), and non-methane volatile organic compounds (NMVOCS) (EPA 2021). Since the mid-20th century, global background ozone (O3) concentrations have rapidly increased [1] with current average concentrations for the mid-northern latitude of ca. 30–50 ppb [2].
Plants can tolerate and adapt to O3 when chronically exposed to concentrations below 20 nL/L (ca. 21 ppb; [3]). However, when ground-level exceeds the tolerance threshold generally estimated at ca. 40 ppb [4,5], O3 becomes phytotoxic and capable of damaging agricultural plants and forest vegetation, and even the biodiversity of a whole ecosystem [2,6]. Following the global increase in O3 background concentrations that occurred in the last 20–30 years, 2–15% reductions in the global crop yield were estimated, with a remarkable economic loss [7].
In recent years, the search for solutions to protect crop plants from O3 injury has become of global interest, and among the tested methodologies, i.e., the development of O3-resistant crops, the selection of resistant germplasm, the use of antiozonants and nanomaterials [8], the use of antioxidants (vitamins, phytohormones, flavonoids, and polyamines), is a very promising strategy to protect crop plants from O3 phytotoxicity [9,10,11,12].
Wood distillate (WD), also known as pyroligneous acid or wood vinegar, is a bio-based liquid product obtained from the distillation of the gases produced during the pyrolysis of woody biomass for the production of green energy [13]. Such a product, still unexploited in many agricultural sectors (especially those not involved in organic farming), is now having great success in agriculture as a phytostimulant for crop plants since its use can enhance both plant productivity and endogenous defenses against pathogens [14]. WD can be produced by different feedstocks but the common feature of all WDs is the presence, to a greater or lesser extent, of antioxidant molecules such as phenols [15]. Results from a study conducted on the horticultural plant Lactuca sativa L. revealed that foliar application of 0.2% WD deriving from sweet chestnut (Castanea sativa Mill.) enhances the content of chlorophyll and biomass of this model species [16], probably owing to its high content of antioxidant compounds, such as polyphenols [17]. Foliar application of WD is also used to increase the defenses of olive (Olea europaea L.) trees and grapevine (Vitis vinifera L.) plants against pathogens [18,19], but the effectiveness of WD in contrasting the phytotoxic action of O3 on crop plant metabolism is still unexplored.
For this reason, this study examines whether foliar applications of WD have a protective effect on lettuce exposed to an ecologically relevant O3 concentration.
In detail, we tested if lettuce plants exposed to O3 but not treated with WD have a similar response, in terms of photosynthetic system, antioxidants, and yield, as plants treated with WD.
2. Materials and Methods
2.1. Experimental
Seedlings of lettuce (Lactuca sativa L.) cv. Cappuccio bionda were bought from a local nursery. In the laboratory, plants were carefully removed from their growth phytocells and transplanted inside plastic pots (10 × 10 × 12 cm) using a commercial potting soil as the substrate. Seedlings were then acclimatized for one week in a climatic chamber at 20 °C, 70% RH, and 350 μM s−1 m−2 PAR with 15 h of photoperiod (from 5:00 a.m. to 20:00 p.m.). The whole pool of seedlings (24 plants) was then fumigated daily with 60 ppb of O3 for 5 h per day (from 12:00 to 17:00). Once per week, 50% of the fumigated plants (12 plants) were treated with foliar applications of 0.2% sweet chestnut (Castanea sativa Mill.) WD (Biodea®), while the remaining 50% of plants were sprayed with water and used as a positive control. Foliar applications were run after the light cycle, following the method described by Vannini et al. [16]. All plants were randomly rotated every three days to minimize any possible influence of microclimatic conditions inside the climatic chamber. The experiment lasted four weeks and was replicated three times. The sweet chestnut WD was selected because it had previously been investigated for its safety, for both the environment and humans [17,20,21].
2.2. Photosynthetic Parameters
Since photosynthesis is considered the main target of O3 phytotoxicity [22], its functionality was assessed by means of selected photosynthetic indicators: the chlorophyll fluorescence, the analysis of the fluorescence transients plotted on a logarithmic scale (OJIP transients), and the chlorophyll content.
2.2.1. Chlorophyll Fluorescence Analyses
The analysis of the chlorophyll fluorescence and the analysis of the OJIP transients are key methodologies to assess the functionality of the photosynthetic machinery following exposure to O3 [5,22]. The former was assessed through a number of the OJIP step fluorescence parameters, summarized in Table 1, which describes the ability of the photosynthetic system to absorb, trap, transmit, and convert the absorbed light into energy for CO2 fixation [23], while the latter analyzed the ΔVOP profile, calculated by the difference between the VOP profile of treated (Ozone + WD) and control (Ozone) samples (ΔVOP = VOPtreated (ozone + WD) − VOPcontrol(ozone); [24]). Prior to analysis, the seedlings were dark-adapted for 30 min under a dim green light (10 μmol photons m−2 s−1) and then lighted with an actinic light (3000 μmol photons m−2 s−1) for one second. Fluorescence analysis was run using a plant efficiency analyzer (Handy PEA, Hansatech Ltd., Norfolk, UK). Fifteen measurements were taken for each replicate.
2.2.2. Analysis of the Chlorophyll Content
The content of chlorophyll was measured by means of a chlorophyll content meter (CCM 300, Opti-Science Inc., Hudson, NH, USA), a non-destructive instrument that quantifies the amount of chlorophyll on a surface basis (mg/m2) [25]. Fifteen measurements were taken for each replicate.
2.3. Expression of Antioxidants
2.3.1. Total Antioxidant Power
Fresh samples (ca. 200 mg) were homogenized with 4 mL of 80% ethanol and then centrifuged at 15,000 rpm for five minutes. The supernatant (100 µL) was then added to 1 mL of a DPPH solution prepared following the protocol reported by Vannini et al. [26]. After the reaction (ca. 1 h) samples were read at 517 nm and the results were expressed as % Antiradical Activity (ARA%) following the Formulae:
where control absorbance is the absorbance of the reagents only.
2.3.2. Content of Caffeic Acid and Quercetin
Given the high number of antioxidant compounds in plant leaves, caffeic acid and quercetin were selected as indicators of polyphenols and flavonoids expression [27,28], respectively. The extraction of caffeic acid and quercetin from lettuce leaves was carried out according to Tokusoglu et al. [29], with modifications. The upper part of each of the major fresh leaves (ca. 1 g) was extracted with 3 mL of 70% acetone containing 1% HCl (v/v). After the homogenization, 0.6 mL of pure HCl was added and the final mixture was first vigorously shaken and then left at 90 °C for two hours. Subsequently, samples were shaken and then filtered at 0.45 µm. The extracts were directly analyzed by HPLC (Perkin-Elmer series 200) coupled with a Diode Array Detector (DAD). The analysis of both caffeic acid and quercetin was run according to the method used by Kumar et al. [30], combining water (solvent A) and acetonitrile (solvent B) eluted for 21 min as a mobile phase following the gradient: 0–5 min (80% A), 5–8 min (60% A), 8–12 min (50% A), 12–17 min (40% A), 17–21 min (20% A); an Agilent C18 column (4.6 x 250 mm; particle size 5µm) was used. Runs were monitored at 280 and 325 nm for caffeic acid and quercetin, respectively. Quantifications were carried out using calibration curves of caffeic acid and quercetin (from 5–100 µg/mL), prepared by dissolving the two pure reagents (Sigma, Sant Louis, MI, USA) in the same solvent used to extract both molecules from the samples.
2.4. Edible Fresh Biomass
From each plant, visibly undamaged leaves (marketable, i.e., those without necrotic and senescent areas) were removed from shoots and then weighed on a precision balance. Results were expressed as grams on a fresh weight basis (g FW).
2.5. Statistical Analysis
To disentangle differences between treated (Ozone + WD) and untreated (Ozone) plants in terms of chlorophyll fluorescence and chlorophyll content, a linear mixed-effect model (LMEM) was fitted for each variable, with treatment as a fixed effect and plant as a random effect [31]. For model validation, scatterplots of the residual and fitted values were used to check for homoscedasticity, and normal probability (qqnorm) plots as well as the Shapiro–Wilk test to check for normality. Models were fitted using the restricted maximum likelihood (REML) estimation, and the significance of the models was checked with type III Anova using the Satterthwaite method [32]. For all the other parameters analyzed, a permutation t-test was used to check for differences between treated (Ozone + WD) and untreated (Ozone) plants. All calculations were run using the free R software [33]; the packages ‘lme4’ and ‘RVAideMemoire’ were used.”
3. Results
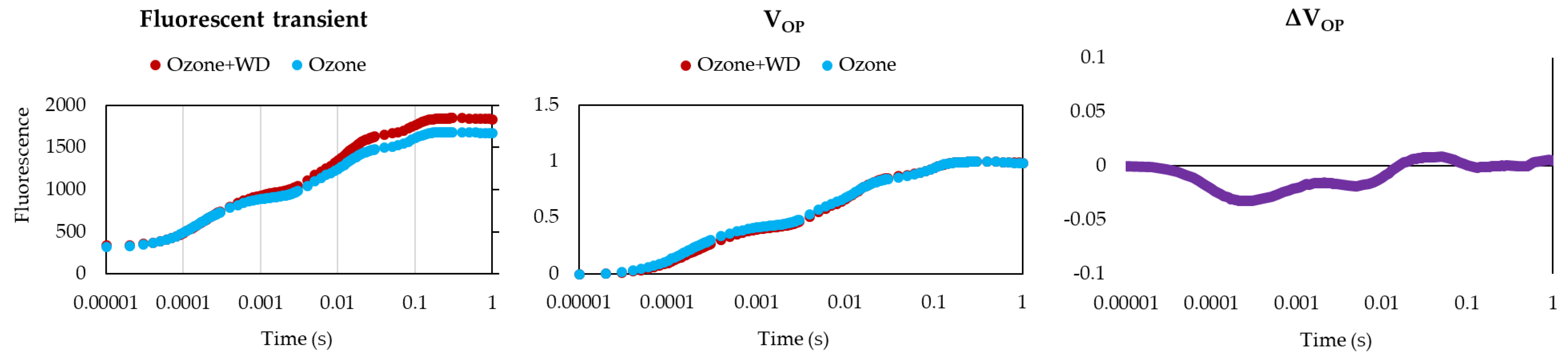
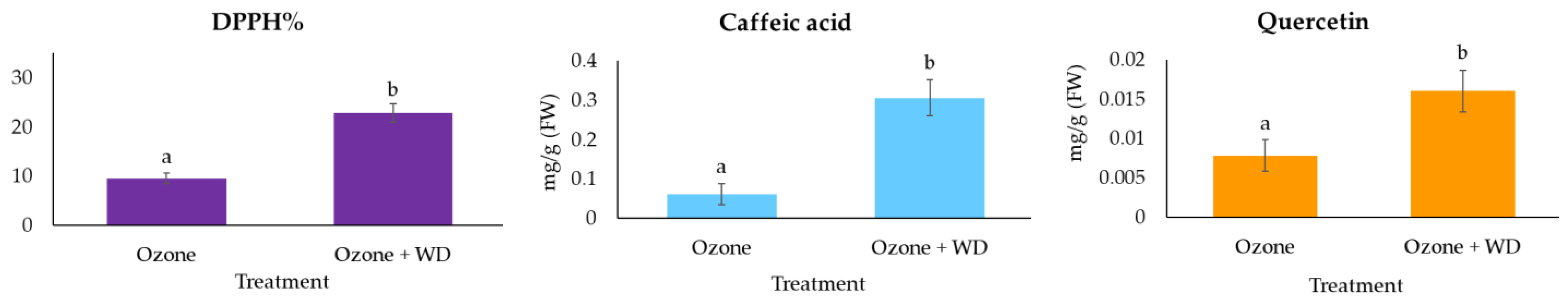
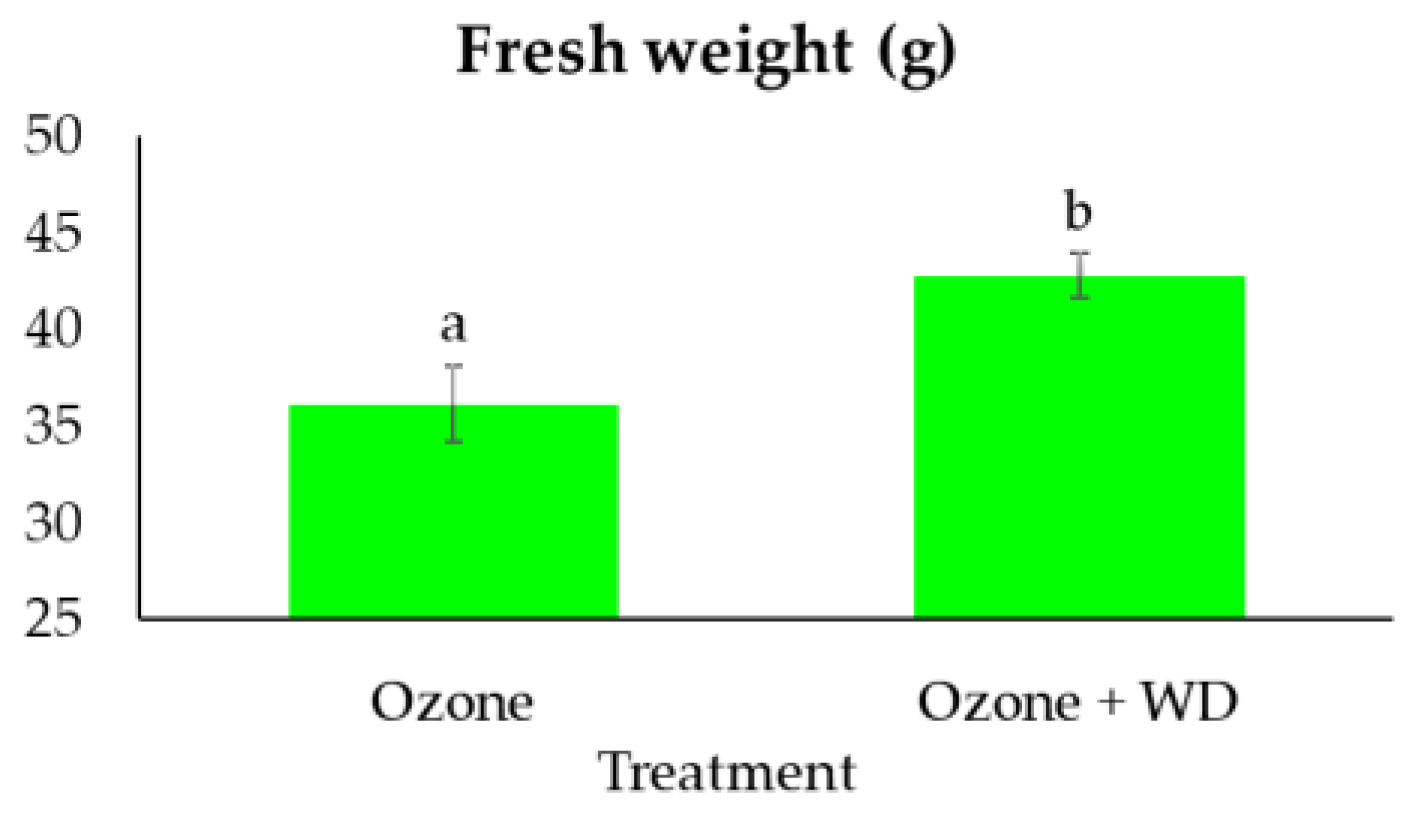
WD-treated samples experienced better photosynthetic performances than those only fumigated with O3 (Table 2). Specifically, WD-treated samples showed a higher absorbance for excited cross-section (ABS/CS0; ca. 3%), energy transmission (TR0/CS0; ca. 4%), electron transport (ET0/CS0; ca. 8%), number of reaction centers (RC/CS0; ca. 11%), as well as a lower energy dissipation (DI0/CS0; ca. 4%). Additionally, WD-treated plants showed a higher (p < 0.05) expression of the photosynthetic efficiency (FV/FM; ca. 1%), of the performance index (PIAbs; ca. 25%), and a higher content of chlorophyll (ca. 13%). Analysis of the two fluorescence profiles confirmed the results obtained by FV/FM, while those of the VOP test indicated only negligible differences between treatments (Figure 1). Moreover, samples treated with WD presented a higher (p < 0.05) expression of the total antioxidant power (ca. 140%), as well as of both caffeic acid (ca. 400%) and quercetin (ca. 105%) (Figure 2). A higher (p < 0.05) biomass (+18%) was also noted (Figure 3).
4. Discussion
Lettuce is an ozone-sensitive horticultural plant [34] and O3 fumigations caused a decrease in photosynthesis, chlorophyll content, and biomass of this species, consistently with other studies [35,36]. However, when 0.2% WD was sprayed on the leaves, plants showed an improvement in the photosynthetic system, a higher content of chlorophyll and antioxidant molecules, as well as higher biomass, thus suggesting the ability of WD to significantly alleviate the O3-induced damage.
WD is rich in antioxidants, such as polyphenols [15,17,37], and is effectively used to increase both the chlorophyll content and biomass of crop plants, as found in some recent studies [16,38,39,40,41]. Although the mechanisms behind its effectiveness are still in need of investigation, it has been suggested that this stimulant effect on plant productivity may be due to the action of antioxidant molecules on cell division [16], in response to the activation of specific transcription genes, as previously observed by Tanase et al. [42,43].
To the best of our knowledge, information on the exact mechanism of action of WD against the O3-induced damage is unknown but it is possible that antioxidant molecules play a very important role. We speculate that the pool of antioxidants could have acted in two ways: buffering the interaction between O3 and the apoplast, thus reducing the overproduction of ROS, i.e., the main factor responsible for the O3-induced toxicity in plants [44], and/or stimulating the synthesis of ROS scavenger molecules, such as superoxide dismutase, peroxidase, and catalase, suggested by Wang et al. [45]. In the first case, WD may have acted as a strengthener of the plant’s antioxidant defenses, while in the second case it could have acted as a stimulator. However, information on which of these two processes plays the greater role has not yet been investigated.
Plants can naturally counteract the oxidative action of O3 through the synthesis of antioxidants, such as reduced glutathione [46], but when insufficient or when the oxidative stress is too high, plants may experience reductions in O3-sensitive antioxidant molecules, such as caffeic acid and quercetin [47,48,49]. However, following the application of 0.2% WD, our lettuce plants showed a higher content of these molecules compared to the untreated ones, thus suggesting a protective role of WD in preventing the oxidation of antioxidant molecules devoted to counteracting the oxidative pressure induced by O3. Consistently with this assumption, WD-treated plants experienced higher antiradical scavenging activity (DPPH) than O3-fumigated plants. From a nutritional point of view, both caffeic acid and quercetin have antioxidant, anti-inflammatory, and anticarcinogenic activities [50,51] and in view of the protective action on these molecules from oxidative degradation, WD can be considered not only as a tool to increase plant productivity but also to defend its nutritional content from the oxidative stress. However, caffeic acid showed higher reductions than quercetin, probably because this molecule is involved in several antioxidative roles. It is reported that this compound is also deputed to shield the photosynthetic system from oxidative stresses [52], thus explaining its higher consumption in O3-fumigated plants.
Photosynthesis is the main target of O3 phytotoxicity [22] and following its damage, both energy transmission mechanisms, reallocation of nutrients, and plant growth can be significantly impaired [44]. Although the photosynthetic system is naturally provided with antioxidant molecules—i.e., caffeic acid—the concentration of ROS above the threshold of the system’s antioxidant defenses may damage it, and programmed cell death processes can be activated [53,54].
Ozone-induced ROS can affect photosynthesis by either damaging structurally the chloroplast, as evident by ultrastructural changes [26,55], and/or activating cascade signals which lead to stomatal closure and, in turn, generate further reductions in net photosynthesis, CO2 assimilation, and plant growth [2,44]. As a result, however, reductions in carbon assimilation may induce increases in the electron transfer with the consequent increase in unwanted ROS overproduction [56]. Hence, the photosynthetic system starts a self-regulatory process to achieve protection from photo-oxidation, leading to an immediate reduction in both the content of chlorophyll and the number of reaction centers [22,57,58], i.e., the structures dedicated to intercepting light to energetically supply the photosynthetic system [59]. With this process, dissipation (DI0) at the expense of absorption and transmission (TR0 and ET0) is favored [22] and reductions in the activity of both PSI and PSII (measured as FV/FM and PI) can be measured, as observed for Canola (Brassica napus L.) and tomato (Solanum lycopersicum L.) plants in open top-chamber fumigations [5,60]. Nevertheless, following WD foliar applications, all the above-mentioned negative effects on the electron transport chain of the photosynthetic system were significantly alleviated. In fact, WD-treated plants experienced a higher number of active reaction centers (RC), positive values for the parameters involved in the energy absorption, trapping, and transmission from the PSII to PSI (ABS, TR, ET), as well as lower heat dissipation energy (DI). Additionally, WD-treated plants showed also a higher expression of the photosynthetic efficiency FV/FM, the maximum quantum yield of primary photochemistry, the performance index PI, the overall indicator of the PSI and PSII functionality [61], and higher content of chlorophyll. Since RCs are naturally subjected to oxidative stress forcing them to be rebuilt very quickly [53], we speculate that WD, thanks to its content of antioxidants, may have greatly reduced this turnover or increased its frequency. Higher photosynthetic efficiency also means more biomass, with a 10% increase in the ratio for every 30% increase in photosynthesis [62]. Ozone can effectively reduce photosynthesis with inevitable consequences for plant growth [44], but by shielding the photosynthetic system from oxidation, the plant’s growth and biomass can also be protected.
5. Conclusions
The results of the present study clearly showed the ability of WD to protect lettuce plants from ozone-induced damage. Specifically, WD-treated plants exhibited lower damage to the photosynthetic machinery, a higher content of chlorophyll (ca. 13%), a higher antioxidant power (ca. 140%), as well as antioxidant molecules (i.e., caffeic acid and quercetin, ca. 400 and 105%, respectively), and higher biomass (ca. 18%). Counteracting the oxidative stress that occurred at the level of the photosynthetic system is speculated to be the main mechanism by which WD protects the plant from ozone-induced damage. Moving forward, WD can be seen not only as a natural product for stimulating plant productivity but also as a means of protecting crop plants against oxidative stress.
Author Contributions
Conceptualization, A.V., S.L. and R.F; methodology, A.V. and R.F.; formal analysis, A.V., R.F. and M.G.; data curation, A.V.; writing—original draft preparation, A.V. and R.F.; writing—review and editing, A.V. and S.L.; supervision, A.V. and S.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The raw data presented in this study are available on request from the corresponding author.
Acknowledgments
Thanks are due to Francesco Barbagli (BioDea and BioEsperia srl) for kindly providing Wood Distillate®.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Cooper, O.R.; Parrish, D.D.; Ziemke, J.R.; Balashov, N.V.; Cupeiro, M.; Galbally, I.; Gilge, S.; Horowitz, L.W.; Jensen, N.R.; Lamarque, J.-F.; et al. Global distribution and trends of tropospheric ozone: An observation-based review. Elementa Sci. Anthr. 2014, 2, 000029. [Google Scholar] [CrossRef]
- Emberson, L. Effects of ozone on agriculture, forests and grasslands. Philos. Trans. R. Soc. A 2020, 378, 20190327. [Google Scholar] [CrossRef]
- Grulke, N.E.; Heath, R.L. Ozone effects on plants in natural ecosystems. Plant Biol. 2019, 22, 12–37. [Google Scholar] [CrossRef] [PubMed]
- Allan, A.C.; Fluhr, R. Ozone and Reactive Oxygen Species. eLS 2001. [Google Scholar] [CrossRef]
- Maliba, B.G.; Inbaraj, P.M.; Berner, J. The Use of OJIP Fluorescence Transients to Monitor the Effect of Elevated Ozone on Biomass of Canola Plants. Water Air Soil Pollut. 2019, 230, 75. [Google Scholar] [CrossRef] [Green Version]
- Feng, Z.; De Marco, A.; Anav, A.; Gualtieri, M.; Sicard, P.; Tian, H.; Fornasier, F.; Tao, F.; Guo, A.; Paoletti, E. Economic losses due to ozone impacts on human health, forest productivity and crop yield across China. Environ. Int. 2019, 131, 104966. [Google Scholar] [CrossRef] [PubMed]
- Ainsworth, E.A. Understanding and improving global crop response to ozone pollution. Plant J. 2017, 90, 886–897. [Google Scholar] [CrossRef] [PubMed]
- Saitanis, C.J.; Agathokleous, E. Exogenous application of chemicals for protecting plants against ambient ozone pollution: What should come next? Curr. Opin. Environ. Sci. Health 2021, 19, 100215. [Google Scholar] [CrossRef] [PubMed]
- Didyk, N.P.; Blum, O.B. Natural antioxidants of plant origin against ozone damage of sensitive crops. Acta Physiol. Plant. 2010, 33, 25–34. [Google Scholar] [CrossRef]
- Bellini, E.; De Tullio, M.C. Ascorbic Acid and Ozone: Novel Perspectives to Explain an Elusive Relationship. Plants 2019, 8, 122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kittipornkul, P.; Treesubsuntorn, C.; Thiravetyan, P. Effect of exogenous catechin and salicylic acid on rice productivity under ozone stress: The role of chlorophyll contents, lipid peroxidation, and antioxidant enzymes. Environ. Sci. Pollut. Res. 2020, 27, 25774–25784. [Google Scholar] [CrossRef] [PubMed]
- Macias-Benitez, S.; Navarro-Torre, S.; Caballero, P.; Martín, L.; Revilla, E.; Castaño, A.; Parrado, J. Biostimulant Capacity of an Enzymatic Extract From Rice Bran Against Ozone-Induced Damage in Capsicum annum. Front. Plant Sci. 2021, 12, 749422. [Google Scholar] [CrossRef] [PubMed]
- Mathew, S.; Zakaria, Z.A. Pyroligneous acid—the smoky acidic liquid from plant biomass. Appl. Microbiol. Biotechnol. 2015, 99, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Grewal, A.; Abbey, L.; Gunupuru, L.R. Production, prospects and potential application of pyroligneous acid in agriculture. J. Anal. Appl. Pyrolysis 2018, 135, 152–159. [Google Scholar] [CrossRef]
- Mahmud, K.N.; Hashim, N.M.; Ani, F.N.; Zakaria, Z.A. Antioxidants, Toxicity, and Nitric Oxide Inhibition Properties of Pyroligneous Acid from Palm Kernel Shell Biomass. Waste Biomass Valorization 2020, 11, 6307–6319. [Google Scholar] [CrossRef]
- Vannini, A.; Moratelli, F.; Monaci, F.; Loppi, S. Effects of wood distillate and soy lecithin on the photosynthetic performance and growth of lettuce (Lactuca sativa L.). SN Appl. Sci. 2021, 3, 1–6. [Google Scholar] [CrossRef]
- Fačkovcová, Z.; Vannini, A.; Monaci, F.; Grattacaso, M.; Paoli, L.; Loppi, S. Effects of wood distillate (pyroligneous acid) on sensitive bioindicators (lichen and moss). Ecotoxicol. Environ. Saf. 2020, 204, 111117. [Google Scholar] [CrossRef] [PubMed]
- Biodea. Available online: https://biodea.bio/scheda-colturale/olive-trees/?lang=en (accessed on 15 January 2022).
- Chen, Y.-H.; Li, Y.-F.; Wei, H.; Li, X.-X.; Zheng, H.-T.; Dong, X.-Y.; Xu, T.-F.; Meng, J.-F. Inhibition efficiency of wood vinegar on grey mould of table grapes. Food Biosci. 2020, 38, 100755. [Google Scholar] [CrossRef]
- Fačkovcová, Z.; Vannini, A.; Monaci, F.; Grattacaso, M.; Paoli, L.; Loppi, S. Uptake of Trace Elements in the Water Fern Azolla filiculoides after Short-Term Application of Chestnut Wood Distillate (Pyroligneous Acid). Plants 2020, 9, 1179. [Google Scholar] [CrossRef]
- Filippelli, A.; Ciccone, V.; Loppi, S.; Morbidelli, L. Characterization of the Safety Profile of Sweet Chestnut Wood Distillate Employed in Agriculture. Safety 2021, 7, 79. [Google Scholar] [CrossRef]
- Bussotti, F.; Strasser, R.J.; Schaub, M. Photosynthetic behavior of woody species under high ozone exposure probed with the JIP-test: A review. Environ. Pollut. 2007, 147, 430–437. [Google Scholar] [CrossRef] [PubMed]
- Strasser, R.J.; Srivastava, A.; Tsimilli-Michael, M. The fluorescence transient as a tool to characterize and screen photosynthetic samples. In Probing Photosynthesis Mechanism, Regulation & Adaptation, 1st ed.; Mohammad, Y., Uday, P., Eds.; CRC Press: New York, NY, USA, 2000; pp. 445–483. [Google Scholar]
- Gomes, M.T.G.; da Luz, A.C.; dos Santos, M.R.; Batitucci, M.d.C.P.; Silva, D.M.; Falqueto, A.R. Drought tolerance of passion fruit plants assessed by the OJIP chlorophyll a fluorescence transient. Sci. Hortic. 2012, 142, 49–56. [Google Scholar] [CrossRef]
- A Gitelson, A.; Buschmann, C.; Lichtenthaler, H.K. The Chlorophyll Fluorescence Ratio F735/F700 as an Accurate Measure of the Chlorophyll Content in Plants. Remote Sens. Environ. 1999, 69, 296–302. [Google Scholar] [CrossRef]
- Vannini, A.; Canali, G.; Pica, M.; Nali, C.; Loppi, S. The Water Content Drives the Susceptibility of the Lichen Evernia prunastri and the Moss Brachythecium sp. to High Ozone Concentrations. Biology 2020, 9, 90. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Arif, Y.; Bajguz, A.; Hayat, S. The role of quercetin in plants. Plant Physiol. Biochem. 2021, 166, 10–19. [Google Scholar] [CrossRef]
- Riaz, U.; Kharal, M.A.; Murtaza, G.; Zaman, Q.U.; Javaid, S.; Malik, H.A.; Aziz, H.; Abbas, Z. Prospective Roles and Mechanisms of Caffeic Acid in Counter Plant Stress: A Mini Review. Pak. J. Agric. Res. 2018, 32, 32. [Google Scholar] [CrossRef]
- Tokuşoğlu, Ö.; Ünal, M.K.; Yıldırım, Z. HPLC-UV and GC-MS characterization of the fla-vonol aglycons quercetin, kaempferol, and myricetin in tomato pastes and other tomato-based products. Acta Chromatogr. 2003, 13, 196–207. [Google Scholar]
- Kumar, N.; Bhandari, P.; Singh, B.; Gupta, A.; Kaul, V.K. Reversed phase-HPLC for rapid determination of polyphenols in flowers of rose species. J. Sep. Sci. 2008, 31, 262–267. [Google Scholar] [CrossRef]
- Brauer, M.; Curtin, J.J. Linear mixed-effects models and the analysis of nonindependent data: A unified framework to analyze categorical and continuous independent variables that vary within-subjects and/or within-items. Psychol. Methods 2018, 23, 389–411. [Google Scholar] [CrossRef]
- Luke, S.G. Evaluating significance in linear mixed-effects models in R. Behav. Res. Methods 2016, 49, 1494–1502. [Google Scholar] [CrossRef] [PubMed]
- R Development Core Team. R: A language and environment for statistical computing; R Foundation for Statistical Computing: Vienna, Austria; Available online: https://www.R-project.org/ (accessed on 7 January 2022).
- Mills, G.; Buse, A.; Gimeno, B.; Bermejo, V.; Holland, M.; Emberson, L.; Pleijel, H. A synthesis of AOT40-based response functions and critical levels of ozone for agricultural and horticultural crops. Atmos. Environ. 2007, 41, 2630–2643. [Google Scholar] [CrossRef]
- Calatayud, A.; Ramirez, J.W.; Iglesias, D.J.; Barreno, E. Effects of ozone on photosynthetic CO2 exchange, chlorophyll a fluorescence and antioxidant systems in lettuce leaves. Physiol. Plant. 2002, 116, 308–316. [Google Scholar] [CrossRef]
- Calatayud, A.; Barreno, E. Response to ozone in two lettuce varieties on chlorophyll a fluorescence, photosynthetic pigments and lipid peroxidation. Plant Physiol. Biochem. 2004, 42, 549–555. [Google Scholar] [CrossRef] [PubMed]
- Gomez, J.P.; Velez, J.P.A.; Pinzon, M.A.; Arango, J.A.M.; Muriel, A.P. Chemical Characterization and Antiradical Properties of Pyroligneous Acid from a Preserved Bamboo, Guadua angustifolia Kunth. Braz. Arch. Biol. Technol. 2021, 64, 21190730. [Google Scholar] [CrossRef]
- Benzon, H.R.L.; Lee, S.C. Pyroligneous Acids Enhance Phytoremediation of Heavy Metal-Contaminated Soils Using Mustard. Commun. Soil Sci. Plant Anal. 2017, 48, 2061–2073. [Google Scholar] [CrossRef]
- Berahim, Z.; Panhwar, Q.A.; Ismail, M.R.; Saud, H.M.; Mondal, A.; Naher, U.A.; Islam, R. Rice yield improvement by foliar application of phytohormone. J. Food Agric. Environ. 2018, 12, 399–404. [Google Scholar]
- Fedeli, R.; Vannini, A.; Guarnieri, M.; Monaci, F.; Loppi, S. Bio-Based Solutions for Agriculture: Foliar Application of Wood Distillate Alone and in Combination with Other Plant-Derived Corroborants Results in Different Effects on Lettuce (Lactuca Sativa L.). Biology 2022, 11, 404. [Google Scholar] [CrossRef]
- Theerakulpisut, P.; Kanawapee, N.; Panwong, B. Seed Priming Alleviated Salt Stress Effects on Rice Seedlings by Improving Na+/K+ and Maintaining Membrane Integrity. Int. J. Plant Biol. 2016, 7, 6402. [Google Scholar] [CrossRef] [Green Version]
- Tanase, C.; Bara, C.I.; Popa, V.I. Cytogenetical effect of some polyphenol compounds separated from industrial by-products on. Cell. Chem. Technol. 2015, 49, 799–805. [Google Scholar]
- Tanase, C.; Bujor, O.-C.; Popa, V.I. Phenolic Natural Compounds and Their Influence on Physiological Processes in Plants. In Polyphenols in Plants; Elsevier: Amsterdam, The Netherlands, 2019; pp. 45–58. [Google Scholar] [CrossRef]
- Wilkinson, S.; Mills, G.; Illidge, R.; Davies, W.J. How is ozone pollution reducing our food supply? J. Exp. Bot. 2012, 63, 527–536. [Google Scholar] [CrossRef]
- Wang, Y.; Qiu, L.; Song, Q.; Wang, S.; Wang, Y.; Ge, Y. Root Proteomics Reveals the Effects of Wood Vinegar on Wheat Growth and Subsequent Tolerance to Drought Stress. Int. J. Mol. Sci. 2019, 20, 943. [Google Scholar] [CrossRef] [Green Version]
- Hassan, I.A.; Haiba, N.S.; Badr, R.H.; Basahi, J.M.; Almeelbi, T.; Ismail, I.M.; Taia, W.K. Effects of ambient ozone on reactive oxygen species and antioxidant metabolites in leaves of pea (Pisum sativum L.) plants. Pak. J. Bot. 2017, 49, 47–55. [Google Scholar]
- Diao, E.; Wang, J.; Li, X.; Wang, X.; Song, H.; Gao, D. Effects of ozone processing on patulin, phenolic compounds and organic acids in apple juice. J. Food Sci. Technol. 2019, 56, 957–965. [Google Scholar] [CrossRef] [PubMed]
- Jeon, J.; Baek, S.-A.; Kim, N.S.; Sathasivam, R.; Park, J.S.; Kim, J.K.; Park, S.U. Elevated Ozone Levels Affect Metabolites and Related Biosynthetic Genes in Tartary Buckwheat. J. Agric. Food Chem. 2020, 68, 14758–14767. [Google Scholar] [CrossRef]
- Kreczmer, B.; Dyba, B.; Barbasz, A.; Rudolphi-Szydło, E. Advantageous/Unfavorable Effect of Quercetin on the Membranes of SK-N-SH Neuroblastoma Cells. Molecules 2021, 26, 4945. [Google Scholar] [CrossRef] [PubMed]
- Espíndola, K.M.M.; Ferreira, R.G.; Narvaez, L.E.M.; Rosario, A.C.R.S.; Da Silva, A.H.M.; Silva, A.G.B.; Vieira, A.P.O.; Monteiro, M.C. Chemical and Pharmacological Aspects of Caffeic Acid and Its Activity in Hepatocarcinoma. Front. Oncol. 2019, 9, 541. [Google Scholar] [CrossRef] [Green Version]
- Salehi, B.; Machin, L.; Monzote, L.; Sharifi-Rad, J.; Ezzat, S.M.; Salem, M.A.; Merghany, R.M.; El Mahdy, N.M.; Kılıç, C.S.; Sytar, O.; et al. Therapeutic Potential of Quercetin: New Insights and Perspectives for Human Health. ACS Omega 2020, 5, 11849–11872. [Google Scholar] [CrossRef] [PubMed]
- Hura, T.; Hura, K.; Grzesiak, S. Possible contribution of cell-wall-bound ferulic acid in drought resistance and recovery in triticale seedlings. J. Plant Physiol. 2009, 166, 1720–1733. [Google Scholar] [CrossRef]
- Foyer, C.H. Reactive oxygen species, oxidative signaling and the regulation of photosynthesis. Environ. Exp. Bot. 2018, 154, 134–142. [Google Scholar] [CrossRef]
- Gandin, A.; Dizengremel, P.; Jolivet, Y. Integrative role of plant mitochondria facing oxidative stress: The case of ozone. Plant Physiol. Biochem. 2020, 159, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Yamane, K.; Kawasaki, M.; Taniguchi, M.; Miyake, H. Correlation between Chloroplast Ultrastructure and Chlorophyll Fluorescence Characteristics in the Leaves of Rice (Oryza sativa L.) Grown under Salinity. Plant Prod. Sci. 2008, 11, 139–145. [Google Scholar] [CrossRef]
- Lascano, H.; Melchiorre, M.N.; Luna, C.M.; Trippi, V.S. Effect of photooxidative stress induced by paraquat in two wheat cultivars with differential tolerance to water stress. Plant Sci. 2003, 164, 841–848. [Google Scholar] [CrossRef]
- Pellegrini, E.; Cotrozzi, L.; Neri, L.; Baraldi, R.; Carrari, E.; Nali, C.; Lorenzini, G.; Paoletti, E.; Hoshika, Y. Stress markers and physiochemical responses of the Mediterranean shrub Phillyrea angustifolia under current and future drought and ozone scenarios. Environ. Res. 2021, 201, 111615. [Google Scholar] [CrossRef] [PubMed]
- Pellegrini, E.; Francini, A.; Lorenzini, G.; Nali, C. Ecophysiological and antioxidant traits of Salvia officinalis under ozone stress. Environ. Sci. Pollut. Res. 2015, 22, 13083–13093. [Google Scholar] [CrossRef]
- Allen, J.; Williams, J. Photosynthetic reaction centers. FEBS Lett. 1998, 438, 5–9. [Google Scholar] [CrossRef] [Green Version]
- Thwe, A.A.; Kasemsap, P. Quantification of OJIP Fluorescence Transient in Tomato Plants Under Acute Ozone Stress. Agric. Nat. Resour. 2014, 48, 665–675. [Google Scholar]
- Gottardini, E.; Cristofori, A.; Cristofolini, F.; Nali, C.; Pellegrini, E.; Bussotti, F.; Ferretti, M. Chlorophyll-related indicators are linked to visible ozone symptoms: Evidence from a field study on native Viburnum lantana L. plants in northern Italy. Ecol. Indic. 2014, 39, 65–74. [Google Scholar] [CrossRef]
- Kirschbaum, M.U. Does Enhanced Photosynthesis Enhance Growth? Lessons Learned from CO2 Enrichment Studies. Plant Physiol. 2011, 155, 117–124. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Average log-probit OJIP fluorescence curves and VOP analysis of samples of Lactuca sativa after 30 days of fumigation with 60 ppb of O3 in combination with (Ozone + WD) or without (Ozone) weekly foliar applications of 0.2% chestnut wood distillate (WD).
Figure 1.
Average log-probit OJIP fluorescence curves and VOP analysis of samples of Lactuca sativa after 30 days of fumigation with 60 ppb of O3 in combination with (Ozone + WD) or without (Ozone) weekly foliar applications of 0.2% chestnut wood distillate (WD).

Figure 2.
Expression of the antioxidant power (DPPH%) and concentration (mean ± standard error) of both caffeic acid and quercetin in samples of Lactuca sativa after 30 days of fumigation with 60 ppb of O3 in combination with (Ozone + WD) or without (Ozone) weekly foliar applications of 0.2% chestnut wood distillate (WD). Different letters indicate statistically significant (p < 0.05) differences between treatments.
Figure 2.
Expression of the antioxidant power (DPPH%) and concentration (mean ± standard error) of both caffeic acid and quercetin in samples of Lactuca sativa after 30 days of fumigation with 60 ppb of O3 in combination with (Ozone + WD) or without (Ozone) weekly foliar applications of 0.2% chestnut wood distillate (WD). Different letters indicate statistically significant (p < 0.05) differences between treatments.

Figure 3.
Fresh weight (mean ± standard error) of samples of Lactuca sativa after 30 days of fumigation with 60 ppb of O3 in combination with (Ozone + WD) or without (Ozone) weekly foliar applications of 0.2% chestnut wood distillate. Different letters indicate statistically significant differences between treatments (p-value < 0.05).
Figure 3.
Fresh weight (mean ± standard error) of samples of Lactuca sativa after 30 days of fumigation with 60 ppb of O3 in combination with (Ozone + WD) or without (Ozone) weekly foliar applications of 0.2% chestnut wood distillate. Different letters indicate statistically significant differences between treatments (p-value < 0.05).


{kind=link}
{kind=link}
{kind=link}
Table 1.
Description of the measured photosynthetic parameters.
| Parameter | Description |
|---|---|
| ABS/CS0 | Absorbance for excited cross-section |
| TR0/CS0 | Trapping flux for excited cross-section |
| ET0/CS0 | Energy transmission for excited cross-section |
| RC/CS0 | Number of reaction centers for excited cross-section |
| DI0/CS0 | Heat dissipation for excited cross-section |
| FV/FM | Photosynthetic efficiency |
| PIABS | Performance index |
Table 2.
Expression of photosynthetic parameters (mean ± standard error) in samples of Lactuca sativa after 30 days of fumigation with 60 ppb of O3 in combination with (Ozone + WD) or without (Ozone) weekly foliar applications of 0.2% chestnut wood distillate (WD).
Table 2.
Expression of photosynthetic parameters (mean ± standard error) in samples of Lactuca sativa after 30 days of fumigation with 60 ppb of O3 in combination with (Ozone + WD) or without (Ozone) weekly foliar applications of 0.2% chestnut wood distillate (WD).
| Parameter | Ozone | Ozone + WD | p-Value |
|---|---|---|---|
| ABS/CS0 | 293 ± 2.5 a | 301 ± 2.6 b | p < 0.05 |
| DI0/CS0 | 51 ± 0.5 a | 49 ± 0.5 b | p < 0.01 |
| TR0/CS0 | 242 ± 2.0 a | 252 ± 2.1 b | p < 0.001 |
| ET0/CS0 | 131 ± 1.4 a | 141 ± 1.4 b | p < 0.001 |
| RC/CS0 | 105 ± 1.1 a | 117 ± 1.1 b | p < 0.001 |
| FV/FM | 0.826 ± 0.001 a | 0.837 ± 0.001 b | p < 0.001 |
| PIABS | 2.05 ± 0.03 a | 2.56 ± 0.04 b | p < 0.001 |
| Chlorophyll (mg/m2) | 200 ± 3.7 a | 225 ± 2.7 b | p < 0.001 |
Different letters indicate statistically significant (p < 0.05) differences between treatments. Abbreviations: absorbance for excited cross-section (ABS/CS0), heat dissipation for excited cross-section (DI0/CS0), trapping flux for excited cross-section (TR0/CS0), energy transmission for excited cross-section (ET0/CS0), number of reaction centers for excited cross-section (RC/CS0), photosynthetic efficiency (FV/FM), performance index (PIABS), chlorophyll content (Chlorophyll).
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Vannini, A.; Fedeli, R.; Guarnieri, M.; Loppi, S. Foliar Application of Wood Distillate Alleviates Ozone-Induced Damage in Lettuce (Lactuca sativa L.). Toxics 2022, 10, 178. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics10040178
AMA Style
Vannini A, Fedeli R, Guarnieri M, Loppi S. Foliar Application of Wood Distillate Alleviates Ozone-Induced Damage in Lettuce (Lactuca sativa L.). Toxics. 2022; 10(4):178. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics10040178
Chicago/Turabian StyleVannini, Andrea, Riccardo Fedeli, Massimo Guarnieri, and Stefano Loppi. 2022. "Foliar Application of Wood Distillate Alleviates Ozone-Induced Damage in Lettuce (Lactuca sativa L.)" Toxics 10, no. 4: 178. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics10040178
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.