
Environmental Co-Exposure to Potassium Perchlorate and Cd Caused Toxicity and Thyroid Endocrine Disruption in Zebrafish Embryos and Larvae (Danio rerio)

 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Zebrafish Maintenance and Embryo Collection
2.2. Survival Rate, Hatching Rate, and Morphology Score
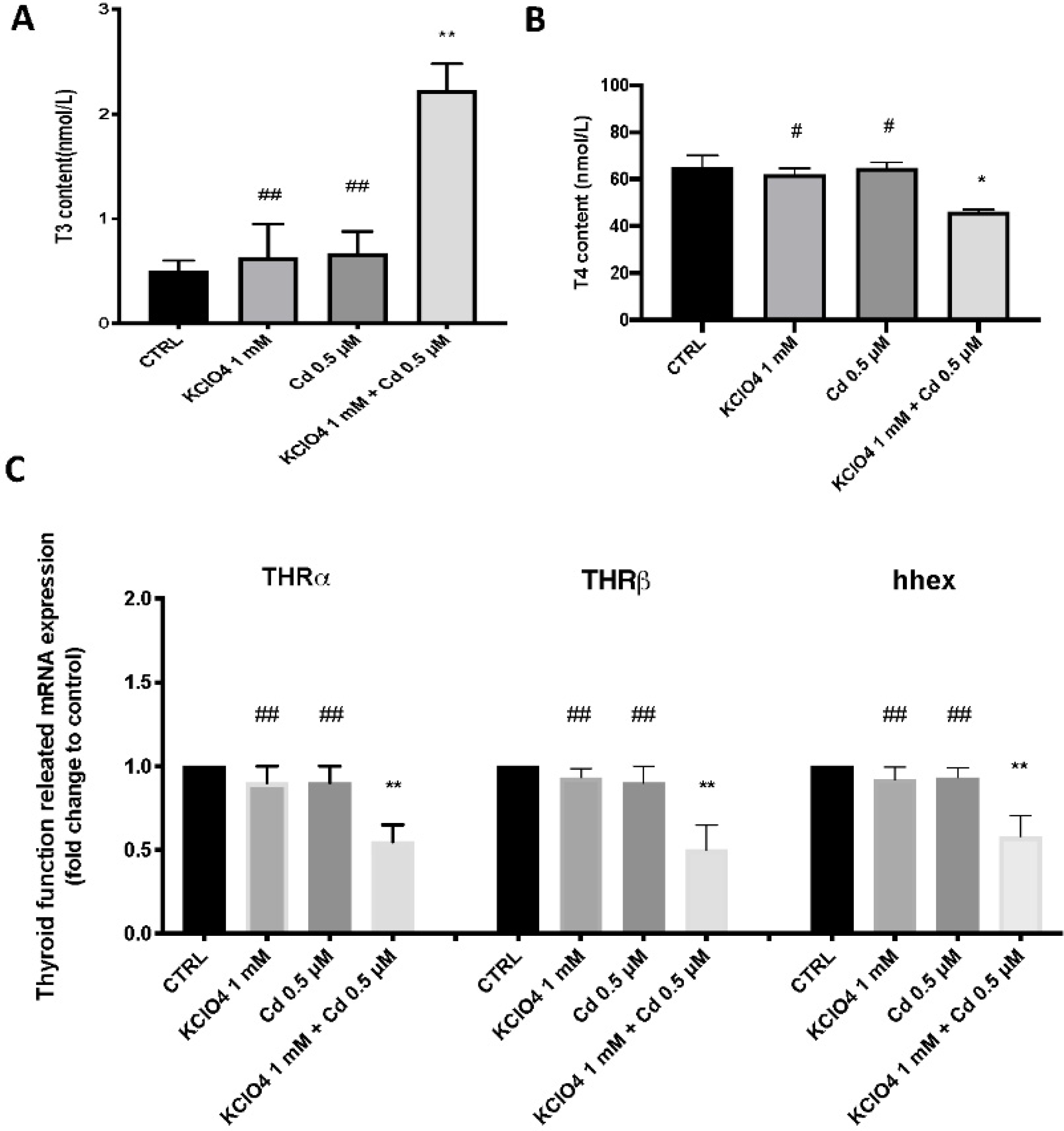
2.3. Thyroid Hormone Measurement
2.4. Gene Expression Analysis
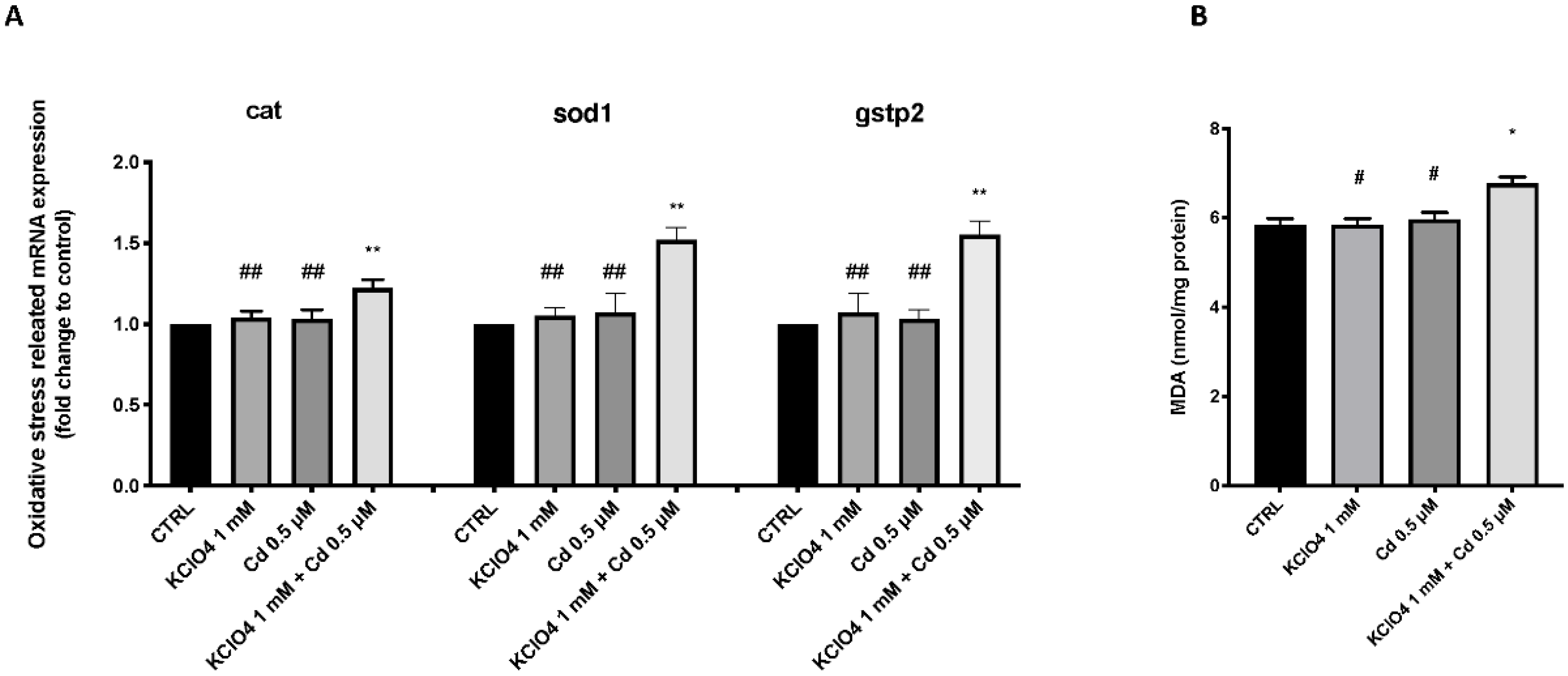
2.5. Lipid Peroxidation
2.6. Statistical Evaluation
3. Results
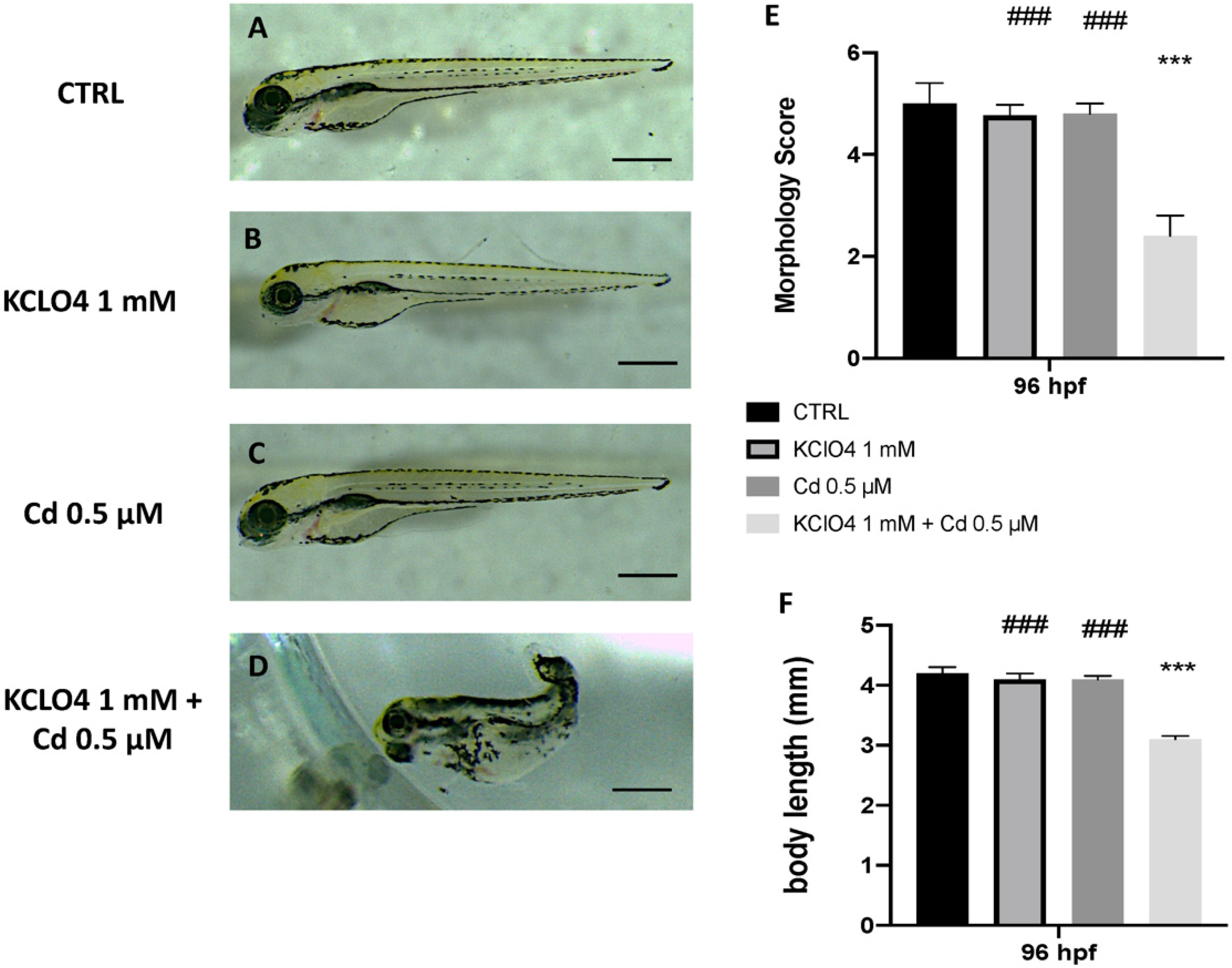
3.1. Survival, Hatching Rate, Morphology, and Thyroid Hormone Levels
3.2. Toxic Effect of Combined Exposure to KClO4 and Cd on Malformation and Body Lenght
3.3. Thyroid Function after Single Exposure and Co-Exposure to KClO4 and Cd
3.4. Effect of KClO4 and Cd Antioxidant Pathway and Lipid Peroxidation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Brown, D.D. The role of thyroid hormone in zebrafish and axolotl development. Proc. Natl. Acad. Sci. USA 1997, 94, 13011–13016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Einarsdóttir, I.E.; Silva, N.; Power, D.M.; Smaradottir, H.; Björnsson, B.T. Thyroid and pituitary gland development from hatching through metamorphosis of a teleost flatfish, the Atlantic halibut. Anat. Embryol. 2006, 211, 47–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miwa, S.; Inui, Y. Effects of various doses of thyroxine and triiodothyronine on the metamorphosis of flounder (Paralichthys olivaceus). General Comp. Endocrinol. 1987, 67, 356–363. [Google Scholar] [CrossRef]
- Schmidt, F.; Schnurr, S.; Wolf, R.; Braunbeck, T. Effects of the anti-thyroidal compound potassium-perchlorate on the thyroid system of the zebrafish. Aquat. Toxicol. 2012, 109, 47–58. [Google Scholar] [CrossRef]
- Elsalini, O.A.; Rohr, K.B. Phenylthiourea disrupts thyroid function in developing zebrafish. Dev. Genes Evol. 2003, 212, 593–598. [Google Scholar] [CrossRef] [PubMed]
- Alt, B.; Reibe, S.; Feitosa, N.M.; Elsalini, O.A.; Wendl, T.; Rohr, K.B. Analysis of origin and growth of the thyroid gland in zebrafish. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 2006, 235, 1872–1883. [Google Scholar] [CrossRef] [PubMed]
- Shi, G.; Wang, J.; Guo, H.; Sheng, N.; Cui, Q.; Pan, Y.; Guo, Y.; Sun, Y.; Dai, J. Parental exposure to 6: 2 chlorinated polyfluorinated ether sulfonate (F-53B) induced transgenerational thyroid hormone disruption in zebrafish. Sci. Total Environ. 2019, 665, 855–863. [Google Scholar] [CrossRef]
- Brown, S.B.; Adams, B.A.; Cyr, D.G.; Eales, J.G. Contaminant effects on the teleost fish thyroid. Environ. Toxicol. Chem. Int. J. 2004, 23, 1680–1701. [Google Scholar] [CrossRef]
- Urbansky, E.T. Perchlorate chemistry: Implications for analysis and remediation. Bioremediat. J. 1998, 2, 81–95. [Google Scholar] [CrossRef] [Green Version]
- Stephenson, J.B. Perchlorate: Occurrence is Widespread but at Varying Levels; Federal Agencies Have Taken Some Actions to Respond to and Lessen Releases; DIANE Publishing: Darby, PA, USA, 2011. [Google Scholar]
- Norris, D.O.; Carr, J.A. Endocrine Disruption: Biological Bases for Health Effects in Wildlife and Humans; Oxford University Press: Oxford, UK, 2005. [Google Scholar]
- Wolff, J. Perchlorate and the thyroid gland. Pharmacol. Rev. 1998, 50, 89–106. [Google Scholar]
- Varghese, S.; Shameena, B.; Oommen, O.V. Thyroid hormones regulate lipid peroxidation and antioxidant enzyme activities in Anabas testudineus (Bloch). Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2001, 128, 165–171. [Google Scholar] [CrossRef]
- Nriagu, J.O. A global assessment of natural sources of atmospheric trace metals. Nature 1989, 338, 47–49. [Google Scholar] [CrossRef]
- Woo, S.; Yum, S.; Park, H.-S.; Lee, T.-K.; Ryu, J.-C. Effects of heavy metals on antioxidants and stress-responsive gene expression in Javanese medaka (Oryzias javanicus). Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2009, 149, 289–299. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, S.; Yadav, P.; Bhatnagar, D. Cadmium-induced lipid peroxidation and the antioxidant system in rat erythrocytes: The role of antioxidants. J. Trace Elem. Med. Biol. 1997, 11, 8–13. [Google Scholar] [CrossRef]
- Cordaro, M.; Siracusa, R.; Fusco, R.; D’Amico, R.; Peritore, A.F.; Gugliandolo, E.; Genovese, T.; Scuto, M.; Crupi, R.; Mandalari, G. Cashew (Anacardium occidentale L.) nuts counteract oxidative stress and inflammation in an acute experimental model of Carrageenan-induced Paw edema. Antioxidants 2020, 9, 660. [Google Scholar] [CrossRef] [PubMed]
- Järup, L.; Åkesson, A. Current status of cadmium as an environmental health problem. Toxicol. Appl. Pharmacol. 2009, 238, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Capriello, T.; Grimaldi, M.C.; Cofone, R.; D’Aniello, S.; Ferrandino, I. Effects of aluminium and cadmium on hatching and swimming ability in developing zebrafish. Chemosphere 2019, 222, 243–249. [Google Scholar] [CrossRef]
- Yang, M.; Ren, B.; Qiao, L.; Ren, B.; Hu, Y.; Zhao, R.; Ren, Z.; Du, J. Behavior responses of zebrafish (Danio rerio) to aquatic environmental stresses in the characteristic of circadian rhythms. Chemosphere 2018, 210, 129–138. [Google Scholar] [CrossRef]
- Buschmann, J. The OECD guidelines for the testing of chemicals and pesticides. Methods Mol. Biol. 2013, 947, 37–56. [Google Scholar] [CrossRef]
- Parenti, C.C.; Ghilardi, A.; Della Torre, C.; Magni, S.; Del Giacco, L.; Binelli, A. Evaluation of the infiltration of polystyrene nanobeads in zebrafish embryo tissues after short-term exposure and the related biochemical and behavioural effects. Environ. Pollut. 2019, 254, 112947. [Google Scholar] [CrossRef]
- Kuder, R.S.; Gundala, H.P. Developmental toxicity of deltamethrin and 3-phenoxybenzoic acid in embryo-larval stages of zebrafish (Danio rerio). Toxicol. Mech. Methods 2018, 28, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, Y.; Liu, K.; He, Q.; Sun, C.; Han, J.; Han, L.; Tian, Q. Xiaoaiping induces developmental toxicity in zebrafish embryos through activation of ER stress, apoptosis and the Wnt pathway. Front. Pharmacol. 2018, 9, 1250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, L.; Deng, J.; Shi, X.; Liu, C.; Yu, K.; Zhou, B. Exposure to DE-71 alters thyroid hormone levels and gene transcription in the hypothalamic–pituitary–thyroid axis of zebrafish larvae. Aquat. Toxicol. 2010, 97, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Di Paola, D.; Natale, S.; Gugliandolo, E.; Cordaro, M.; Crupi, R.; Siracusa, R.; D’Amico, R.; Fusco, R.; Impellizzeri, D.; Cuzzocrea, S.; et al. Assessment of 2-Pentadecyl-2-oxazoline Role on Lipopolysaccharide-Induced Inflammation on Early Stage Development of Zebrafish (Danio rerio). Life 2022, 12, 128. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Chen, R.; Liu, W.; Fu, Z. Effect of endocrine disrupting chemicals on the transcription of genes related to the innate immune system in the early developmental stage of zebrafish (Danio rerio). Fish Shellfish. Immun. 2010, 28, 854–861. [Google Scholar] [CrossRef] [PubMed]
- Varela, M.; Dios, S.; Novoa, B.; Figueras, A. Characterisation, expression and ontogeny of interleukin-6 and its receptors in zebrafish (Danio rerio). Dev. Comp. Immunol. 2012, 37, 97–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Takagi, N.; Yuan, B.; Zhou, Y.; Si, N.; Wang, H.; Yang, J.; Wei, X.; Zhao, H.; Bian, B. The protection of indolealkylamines from LPS-induced inflammation in zebrafish. J. Ethnopharmacol. 2019, 243, 112122. [Google Scholar] [CrossRef]
- Hunt, R.F.; Hortopan, G.A.; Gillespie, A.; Baraban, S.C. A novel zebrafish model of hyperthermia-induced seizures reveals a role for TRPV4 channels and NMDA-type glutamate receptors. Exp. Neurol. 2012, 237, 199–206. [Google Scholar] [CrossRef] [Green Version]
- Steenbergen, P.J.; Bardine, N. Antinociceptive effects of buprenorphine in zebrafish larvae: An alternative for rodent models to study pain and nociception? Appl. Anim. Behav. Sci. 2014, 152, 92–99. [Google Scholar] [CrossRef]
- Kim, S.; Ji, K.; Lee, S.; Lee, J.; Kim, J.; Kim, S.; Kho, Y.; Choi, K. Perfluorooctane sulfonic acid exposure increases cadmium toxicity in early life stage of zebrafish, Danio rerio. Environ. Toxicol. Chem. 2011, 30, 870–877. [Google Scholar] [CrossRef]
- Di Paola, D.; Capparucci, F.; Lanteri, G.; Cordaro, M.; Crupi, R.; Siracusa, R.; D’Amico, R.; Fusco, R.; Impellizzeri, D.; Cuzzocrea, S. Combined Toxicity of Xenobiotics Bisphenol A and Heavy Metals on Zebrafish Embryos (Danio rerio). Toxics 2021, 9, 344. [Google Scholar] [CrossRef] [PubMed]
- Samaee, S.M.; Rabbani, S.; Jovanovic, B.; Mohajeri-Tehrani, M.R.; Haghpanah, V. Efficacy of the hatching event in assessing the embryo toxicity of the nano-sized TiO(2) particles in zebrafish: A comparison between two different classes of hatching-derived variables. Ecotoxicol. Environ. Saf. 2015, 116, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Li, Y.; Coelhan, M.; Chan, H.M.; Ma, W.; Liu, L. Relative developmental toxicity of short-chain chlorinated paraffins in Zebrafish (Danio rerio) embryos. Environ. Pollut. 2016, 219, 1122–1130. [Google Scholar] [CrossRef] [PubMed]
- Ismail, A.; Yusof, S. Effect of mercury and cadmium on early life stages of Java medaka (Oryzias javanicus): A potential tropical test fish. Mar. Pollut. Bull. 2011, 63, 347–349. [Google Scholar] [CrossRef]
- Papiya, S.; Kanamadi, R. Effect of mercurial fungicide Emisan®-6 on the embryonic developmental stages of zebrafish, Brachydanio (Danio) rerio. J. Adv. Zool. 2000, 21, 12–18. [Google Scholar]
- Morin, S.; Duong, T.; Dabrin, A.; Coynel, A.; Herlory, O.; Baudrimont, M.; Delmas, F.; Durrieu, G.; Schäfer, J.; Winterton, P. Long-term survey of heavy-metal pollution, biofilm contamination and diatom community structure in the Riou Mort watershed, South-West France. Environ. Pollut. 2008, 151, 532–542. [Google Scholar] [CrossRef]
- Kusch, R.C.; Krone, P.H.; Chivers, D.P. Chronic exposure to low concentrations of waterborne cadmium during embryonic and larval development results in the long-term hindrance of antipredator behavior in zebrafish. Environ. Toxicol. Chem. Int. J. 2008, 27, 705–710. [Google Scholar] [CrossRef]
- Yang, X.; Wang, X.; Gao, D.; Zhang, Y.; Chen, X.; Xia, Q.; Jin, M.; Sun, C.; He, Q.; Wang, R. Developmental toxicity caused by sanguinarine in zebrafish embryos via regulating oxidative stress, apoptosis and wnt pathways. Toxicol. Lett. 2021, 350, 71–80. [Google Scholar] [CrossRef]
- Li, H.; Zhang, Q.; Li, W.; Li, H.; Bao, J.; Yang, C.; Wang, A.; Wei, J.; Chen, S.; Jin, H. Role of Nrf2 in the antioxidation and oxidative stress induced developmental toxicity of honokiol in zebrafish. Toxicol. Appl. Pharmacol. 2019, 373, 48–61. [Google Scholar] [CrossRef]
- Baillie, T.A.; Rettie, A.E. Role of biotransformation in drug-induced toxicity: Influence of intra- and inter-species differences in drug metabolism. Drug Metab. Pharmacokinet. 2011, 26, 15–29. [Google Scholar] [CrossRef] [Green Version]
- Leung, L.; Kalgutkar, A.S.; Obach, R.S. Metabolic activation in drug-induced liver injury. Drug Metab. Rev. 2012, 44, 18–33. [Google Scholar] [CrossRef] [PubMed]
- Wijesinghe, W.A.; Kim, E.A.; Kang, M.C.; Lee, W.W.; Lee, H.S.; Vairappan, C.S.; Jeon, Y.J. Assessment of anti-inflammatory effect of 5beta-hydroxypalisadin B isolated from red seaweed Laurencia snackeyi in zebrafish embryo in vivo model. Environ. Toxicol. Pharmacol. 2014, 37, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Fusco, R.; Gugliandolo, E.; Siracusa, R.; Scuto, M.; Cordaro, M.; D’Amico, R.; Evangelista, M.; Peli, A.; Peritore, A.F.; Impellizzeri, D. Formyl peptide receptor 1 signaling in acute inflammation and neural differentiation induced by traumatic brain injury. Biology 2020, 9, 238. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Liu, Z.; Liu, F.; Ye, Y.; Peng, T.; Fu, Z. Embryonic exposure to cadmium (II) and chromium (VI) induce behavioral alterations, oxidative stress and immunotoxicity in zebrafish (Danio rerio). Neurotoxicol. Teratol. 2015, 48, 9–17. [Google Scholar] [CrossRef]
- Esposito, E.; Campolo, M.; Casili, G.; Lanza, M.; Franco, D.; Filippone, A.; Peritore, A.F.; Cuzzocrea, S. Protective effects of xyloglucan in association with the polysaccharide gelose in an experimental model of gastroenteritis and urinary tract infections. International Journal of Molecular Sciences 2018, 19, 1844. [Google Scholar] [CrossRef] [Green Version]
- Dong, M.; Zhu, L.; Shao, B.; Zhu, S.; Wang, J.; Xie, H.; Wang, J.; Wang, F. The effects of endosulfan on cytochrome P450 enzymes and glutathione S-transferases in zebrafish (Danio rerio) livers. Ecotoxicol. Environ. Saf. 2013, 92, 1–9. [Google Scholar] [CrossRef]
- Yim, M.; Chock, P.B.; Stadtman, E. Enzyme function of copper, zinc superoxide dismutase as a free radical generator. J. Biol. Chem. 1993, 268, 4099–4105. [Google Scholar] [CrossRef]
- Shi, X.; Liu, C.; Wu, G.; Zhou, B. Waterborne exposure to PFOS causes disruption of the hypothalamus–pituitary–thyroid axis in zebrafish larvae. Chemosphere 2009, 77, 1010–1018. [Google Scholar] [CrossRef]
- Chan, W.K.; Chan, K.M. Disruption of the hypothalamic-pituitary-thyroid axis in zebrafish embryo–larvae following waterborne exposure to BDE-47, TBBPA and BPA. Aquat. Toxicol. 2012, 108, 106–111. [Google Scholar] [CrossRef]
- Tian, J.; Hu, J.; He, W.; Zhou, L.; Huang, Y. Parental exposure to cadmium chloride causes developmental toxicity and thyroid endocrine disruption in zebrafish offspring. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2020, 234, 108782. [Google Scholar] [CrossRef]
- Chanoine, J.-P.; Braverman, L.E.; Farwell, A.P.; Safran, M.; Alex, S.; Dubord, S.; Leonard, J.L. The thyroid gland is a major source of circulating T3 in the rat. J. Clin. Investig. 1993, 91, 2709–2713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.-W.; Chan, W.-K. Thyroid hormones are important for embryonic to larval transitory phase in zebrafish. Differentiation 2002, 70, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Sandler, B.; Webb, P.; Apriletti, J.W.; Huber, B.R.; Togashi, M.; Lima, S.T.C.; Juric, S.; Nilsson, S.; Wagner, R.; Fletterick, R.J. Thyroxine-thyroid hormone receptor interactions. J. Biol. Chem. 2004, 279, 55801–55808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elsalini, O.A.; Von Gartzen, J.; Cramer, M.; Rohr, K.B. Zebrafish hhex, nk2. 1a, and pax2. 1 regulate thyroid growth and differentiation downstream of Nodal-dependent transcription factors. Dev. Biol. 2003, 263, 67–80. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Gene | Accession Number | Primer Orientation | Nucleotide Sequence |
|---|---|---|---|
| b-actin | NM_131031 | forward | 5′-CTTCCAGCAGATGTGGATCA-3′ |
| reverse | 5′-GCCATTTAAGGTGGCAACA-3′ | ||
| hhex | NM_130934.1 | forward | 5′-TGTGGTCTCCGTTCATCCAG-3′ |
| reverse | 5′-TTTGACCTGTCTCTCGCTGA-3′ | ||
| THRa | NM_131396.1 | forward | 5′-CAATGTACCATTTCGCGTTG-3′ |
| reverse | 5′-GCTCCTGC TCTGTGTTTTCC-3′ | ||
| THRb | NM_131340.1 | forward | 5′-TGGGAGATGATACGGGTTGT-3′ |
| reverse | 5′-ATAGGTGCCGATCCAATGTC-3′ | ||
| sod1 | NM_131294.1 | forward | 5′-GGCCAACCGATAGTGTTAGA-3′ |
| reverse | 5′-CCAGCGTTGCCAGTTTTTAG-3′ | ||
| cat | NM_130912.2 | forward | 5′-AGGGCAACTGGGATCTTACA-3′ |
| reverse | 5′-TTTATGGGACCAGACCTTGG-3′ | ||
| gstp2 | NM_001020513 | forward | 5′-CACAGACCTCGCTTTTCACAC-3′ |
| reverse | 5′-GAGAGAAGCCTCACAGTCGT-3′ |
| Survival | Hatching | Morphology | Thyroid Hormone | ||||
|---|---|---|---|---|---|---|---|
| 72 h | 96 h | 72 h | 96 h | 96 h | T3 | T4 | |
| CTRL | 100 ± 0 | 100 ± 0 | 100 ± 0 | 100 ± 0 | ND | 1.49 ± 0.005 | 78.33 ± 4.40 |
| KClO4 0.5 mM | 100 ± 0.57 | 98 ± 2 | 99.22 ± 0.57 | 100 ± 0.57 | ND | 1.51 ± 0.008 | 76 ± 2.64 |
| KClO4 1 mM | 100 ± 0.57 | 97.33 ± 3.78 | 99 ± 0.10 | 100 ± 0.57 | ND | 1.53 ± 0.006 | 75.67± 2.40 |
| KClO4 1.5 mM | 71 ± 1 *** | 51.67 ± 3.05 *** | 80.67 ± 2.08 *** | 93 ± 3 *** | Scoliosis | 1.98 ± 0.07 *** | 61 ± 3.05 ** |
| KClO4 2 mM | 0 | 0 | 0 | 0 | ND | // | // |
| KClO4 2.5 mM | 0 | 0 | 0 | 0 | ND | // | // |
| KClO4 5 mM | 0 | 0 | 0 | 0 | ND | // | // |
| Cd 0.05 μM | 100 ± 0 | 99 ± 0.57 | 100 ± 0 | 100 ± 0 | ND | 1.51 ± 0.015 | 77 ± 4.16 |
| Cd 0.5 μM | 99.67 ± 0.33 | 98.67 ± 0.66 | 98.67 ± 2.02 | 100 ± 0 | ND | 1.55 ± 0.003 | 76.67 ± 3.33 |
| Cd 1 μM | 99.33 ± 0.33 | 91.67 ± 0.88 * | 96.33 ± 2.02 * | 96.33 ± 2.02 * | ND | 1.63 ± 0.038 * | 64.33 ± 3.92 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Paola, D.; Natale, S.; Iaria, C.; Crupi, R.; Cuzzocrea, S.; Spanò, N.; Gugliandolo, E.; Peritore, A.F. Environmental Co-Exposure to Potassium Perchlorate and Cd Caused Toxicity and Thyroid Endocrine Disruption in Zebrafish Embryos and Larvae (Danio rerio). Toxics 2022, 10, 198. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics10040198
Di Paola D, Natale S, Iaria C, Crupi R, Cuzzocrea S, Spanò N, Gugliandolo E, Peritore AF. Environmental Co-Exposure to Potassium Perchlorate and Cd Caused Toxicity and Thyroid Endocrine Disruption in Zebrafish Embryos and Larvae (Danio rerio). Toxics. 2022; 10(4):198. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics10040198
Chicago/Turabian StyleDi Paola, Davide, Sabrina Natale, Carmelo Iaria, Rosalia Crupi, Salvatore Cuzzocrea, Nunziacarla Spanò, Enrico Gugliandolo, and Alessio Filippo Peritore. 2022. "Environmental Co-Exposure to Potassium Perchlorate and Cd Caused Toxicity and Thyroid Endocrine Disruption in Zebrafish Embryos and Larvae (Danio rerio)" Toxics 10, no. 4: 198. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics10040198