
Insights into the Endocrine Disrupting Activity of Emerging Non-Phthalate Alternate Plasticizers against Thyroid Hormone Receptor: A Structural Perspective


Abstract
:1. Introduction
2. Materials and Methods
2.1. Protein Preparation
2.2. Ligand Preparation
2.3. Induced Fit Docking
2.4. Binding Affinity Calculations
3. Results
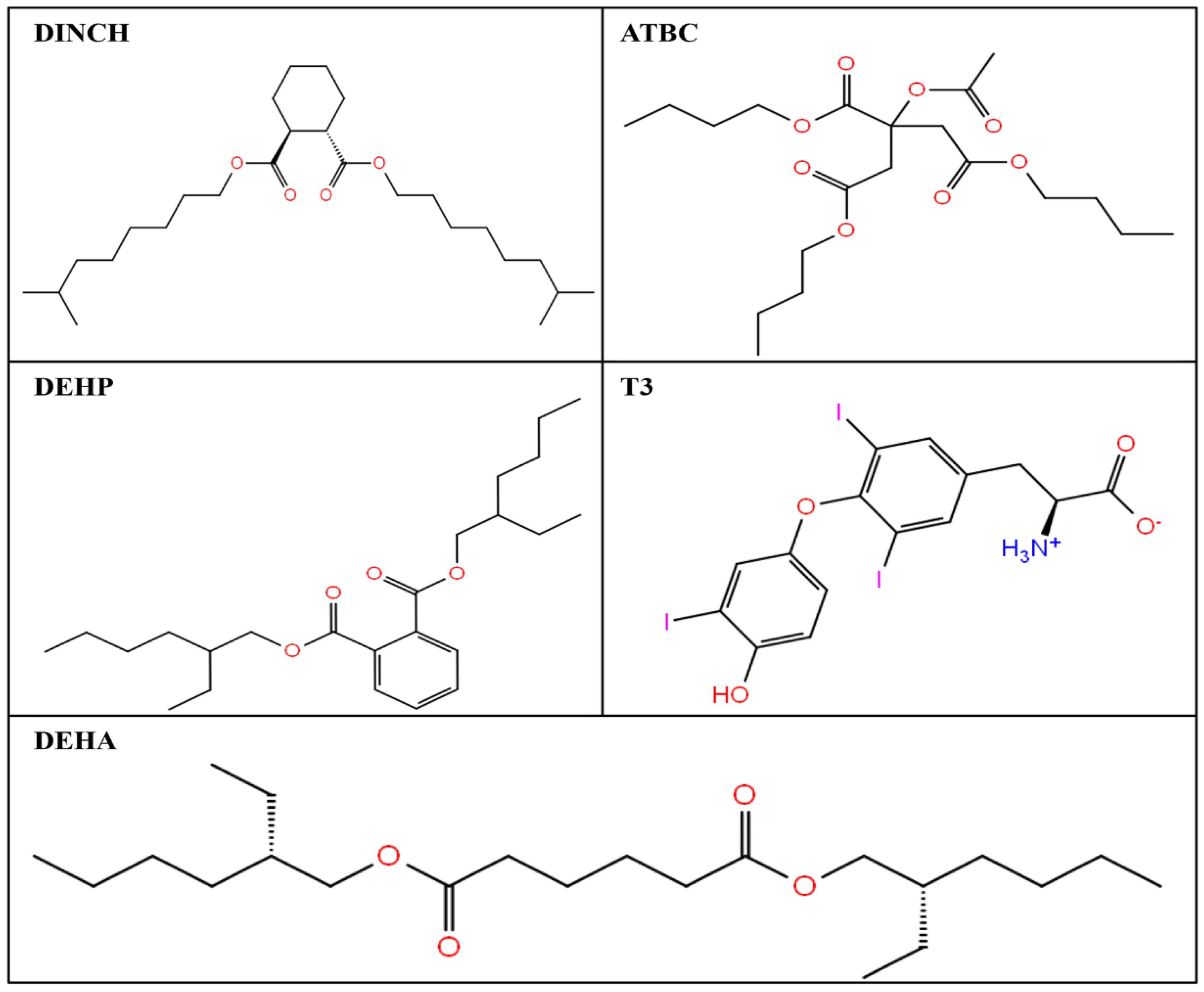
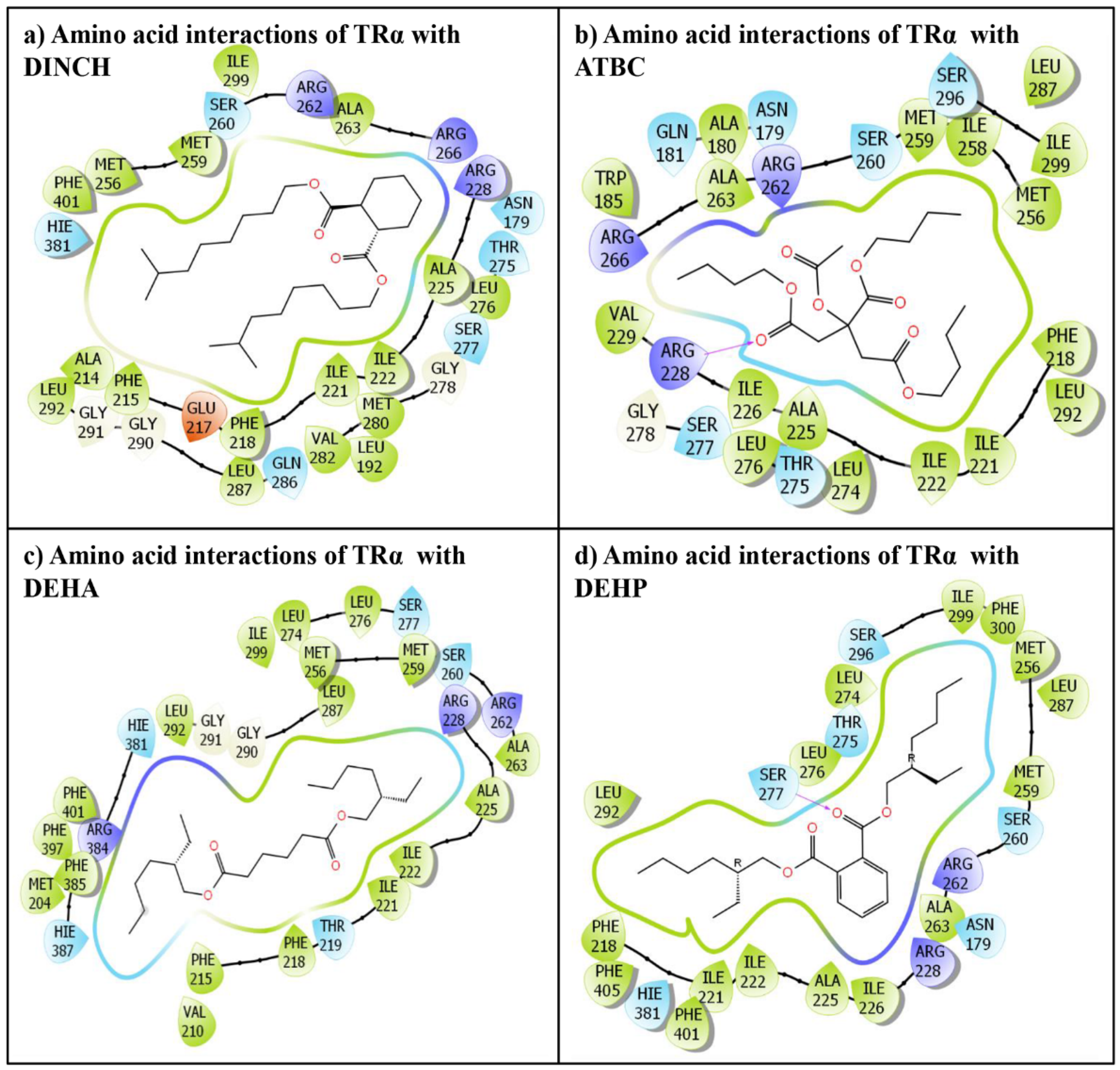
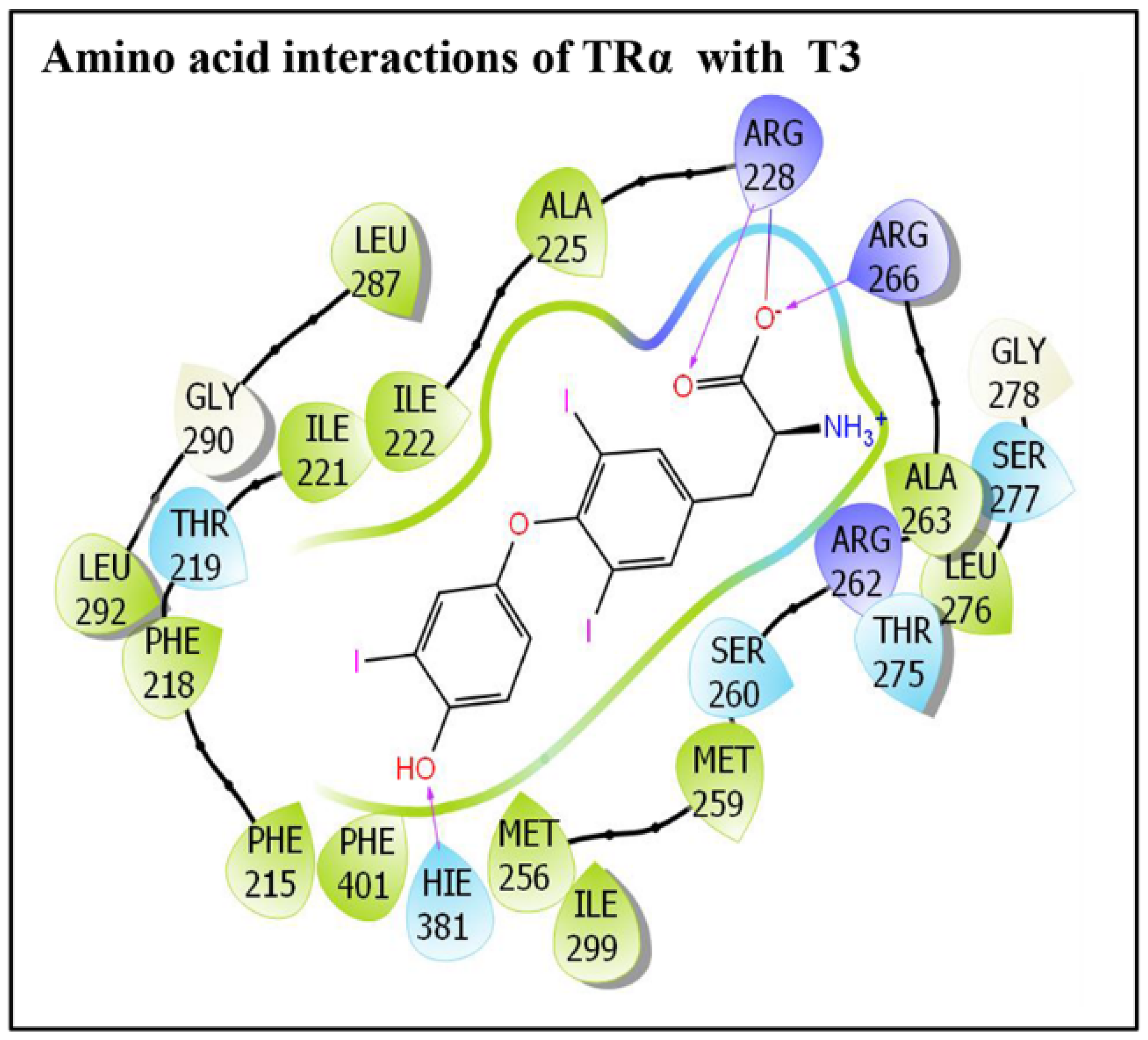
3.1. IFD of Phthalate Plasticizer Ligand, DEHP, with TRα
3.2. IFD of Non-Phthalate Alternate Plasticizers Plasticizers, DINCH, ATBC, and DEHA, with TRα
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- MAM 2022. Plasticizers Market. Plasticizers Market by Type (Low Plasticizers Market by Type (Phthalates, Non-Phthalates), Application (Flooring & Wall Covering, Wire & Cable, Coated Fabric, Consumer Goods, Film & Sheet), and Region (Asia Pacific, Europe, North America)—Global Forecast to 2022; Markets and Markets Research Private Ltd.: Pune, India, 2018; Available online: https://www.marketsandmarkets.com/Market-Reports/plasticizers-market-688.html#:~:text=Phthalates%20plasticizers%20account%20for%20more,easy%20availability%20at%20low%20cost (accessed on 30 March 2022).
- Qadeer, A.; Kirsten, K.L.; Ajmal, Z.; Jiang, X.; Zhao, X. Alternative plasticizers as emerging global environmental and health threat: Another regrettable substitution. Environ. Sci. Technol. 2022, 56, 1482–1488. [Google Scholar] [CrossRef]
- Kim, W.; Gye, M.C. Maleficent effects of phthalates and current states of their alternatives: A review. Korean J. Environ. Biol. 2017, 35, 21–36. [Google Scholar] [CrossRef]
- Rajkumar, A.; Luu, T.; Beal, M.A.; Barton-Maclaren, T.S.; Hales, B.F.; Robaire, B. Phthalates and alternative plasticizers differentially affect phenotypic parameters in gonadal somatic and germ cell lines. Biol. Reprod. 2022, 106, 613–627. [Google Scholar] [CrossRef]
- SCENIHR. Opinion on the Safety of Medical Devices Containing DEHP-Plasticized PVC or Other Plasticizers on Neonates and Other Groups Possibly at Risk. In Scientific Committee on Emerging and Newly-Identified Health Risks; Available online: https://ec.europa/health/scientific_committees/emerging/docs/scenihr_o_047.pdf (accessed on 25 March 2022).
- Guo, Y.; Kannan, K. Comparative assessment of human exposure to phthalate ethers from house dust in China and the United States. Environ. Sci. Technol. 2011, 45, 3788–3794. [Google Scholar] [CrossRef]
- Schecter, A.; Lorber, M.; Guo, Y.; Wu, Q.; Yun, S.; Kannan, K.; Miller, J.; Hynan, L.S.; Cheng, D.; Colacino, J.; et al. Phthalate concentrations and dietary exposure from food purchased in New York State. Environ. Health Perspect. 2013, 121, 473–479. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Kannan, K. A survey of phthalates and parabens in personal care products from the United States and its implications for human exposure. Environ. Sci. Technol. 2013, 47, 14442–14449. [Google Scholar] [CrossRef]
- Bernard, L.; Cueff, R.; Breysse, C.; Décaudin, B.; Sautou, V.; Armed Study Group. Migrability of PVC plasticizers from medical devices into a simulant of infused solutions. Int. J. Pharm. 2015, 485, 341–347. [Google Scholar]
- Demir, A.P.T.; Ulutan, S. Migration of phthalate and non-phthalate plasticizers out of plasticized PVC films into air. J. Appl. Polym. Sci. 2013, 128, 1948–1961. [Google Scholar]
- Fromme, H.; Küchler, T.; Otto, T.; Pilz, K.; Müller, J.; Wenzel, A. Occurrence of phthalates and bisphenol A and F in the environment. Water Res. 2002, 36, 1429–1438. [Google Scholar] [CrossRef]
- Munksgaard, E.C. Leaching of plasticizers from temporary denture soft lining materials. Eur. J. Oral Sci. 2004, 112, 101–104. [Google Scholar] [CrossRef]
- Nara, K.; Nishiyama, K.; Natsugari, H.; Takeshita, A.; Takahashi, H. Leaching of the plasticizer, acetyl tributyl citrate: (ATBC) from plastic kitchen wrap. J. Health Sci. 2009, 55, 281–284. [Google Scholar] [CrossRef] [Green Version]
- Deng, M.; Liang, X.; Du, B.; Luo, D.; Chen, H.; Zhu, C.; Zeng, L. Beyond classic phthalates: Occurrence of multiple emerging phthalate alternatives and their metabolites in human milk and implications for combined exposure in infants. Environ. Sci. Technol. Lett. 2021, 8, 705–712. [Google Scholar] [CrossRef]
- Begum, T.F.; Carpenter, D. Health effects associated with phthalate activity on nuclear receptors. Rev. Environ. Health. 2021, 00010151520200162. [Google Scholar] [CrossRef] [PubMed]
- Replacing Phthalates; ChemSec International Chemical Secretariat: Goteborg, Sweden, 2019; Available online: https://chemsec.org/publication/endocrine-disruptors,substitution/replacing-phthalates/ (accessed on 15 March 2022).
- IARC. Some Chemicals Present in Industrial and Consumer Products, Food and Drinking-Water: Di(2-ethylhexyl)phthalate. In International Agency for Research on Cancer IARC Monographs on the Evaluation of Carcinogenic Risks to Humans; WHO: Lyon, France, 2013; Volume 101, pp. 149–284. Available online: https://monographs.iarc.who.int/wp-content/uploads/2018/06/mono101.pdf (accessed on 10 March 2022).
- Regulation (EU) 2017/745 of the European Parliament and of the Council of 5 April 2017 on Medical Devices, Amending Directive 2001/83/EC, Regulation (EC) No 178/2002 and Regulation (EC) No 1223/2009 and Repealing Council Directives 90/385/EEC and 93/42/EEC (Text with EEA Relevance). Available online: http://data.europa.eu/eli/reg/2017/745/oj/eng (accessed on 5 March 2022).
- Bui, T.T.; Giovanoulis, G.; Cousins, A.P.; Magner, J.; Cousins, I.T.; de Wit, C.A. Human exposure, hazard and risk of alternative plasticizers to phthalate esters. Sci. Total Environ. 2016, 541, 451–467. [Google Scholar] [CrossRef]
- Nehring, A.; Bury, D.; Kling, H.W.; Weiss, T.; Bruning, T.; Koch, H.M. Determination of human urinary metabolites of the plasticizer di(2-ethylhexyl) adipate (DEHA) by online-SPE-HPLC-MS/MS. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2019, 1124, 239–246. [Google Scholar] [CrossRef]
- Non-Phthalate Plasticizers Market by Type (Adipates, Trimellitates, Benzoates, Epoxies, and Others), Application (Flooring & Wall Coverings, Wires & Cables, Films & Sheets, Coated Fabrics, Consumer Goods), and Region-Global Forecast to 2025; Market and Markets: Pune, India, 2020.
- Nagorka, R.; Koschorreck, J. Trends for plasticizers in German freshwater environments-evidence for the substitution of DEHP with emerging phthalate and non-phthalate alternatives. Environ. Pollut. 2020, 262, 114237. [Google Scholar] [CrossRef]
- Edwards, L.; McCray, N.L.; VanNoy, B.N.; Yau, A.; Geller, R.J.; Adamkiewicz, G.; Zota, A.R. Phthalate and novel plasticizer concentrations in food items from U.S. fast food chains: Preliminary analysis. J. Expo. Sci. Environ. Epidemiol. 2021, 2021, 1–8. [Google Scholar] [CrossRef]
- Kim, S.; Kim, Y.; Moon, H.B. Contamination and historical trends of legacy and emerging plasticizers in sediment from highly industrialized bays of Korea. Sci. Total Environ. 2021, 765, 142751. [Google Scholar] [CrossRef]
- Kim, Y.; Kim, S.; Liao, C.; Moon, H.B. Severe contamination and time trend of legacy and alternative plasticizers in a highly industrialized lake associated with regulations and coastal development. Mar. Pollut. Bull. 2021, 171, 112787. [Google Scholar] [CrossRef]
- Jebara, A.; Albergamo, A.; Rando, R.; Potortì, A.G.; Lo Turco, V.; Mansour, H.B.; Di Bella, G. Phthalates and non-phthalate plasticizers in tunisian marine samples: Occurrence, spatial distribution and seasonal variation. Mar. Pollut. Bull. 2021, 163, 111967. [Google Scholar] [CrossRef]
- Zhu, Y.D.; Han, X.; Wang, X.Q.; Ge, T.X.; Liu, H.; Fan, L.; Li, L.; Su, L.Q.; Wang, X.L. Effect of the phthalates exposure on sex steroid hormones in the US population. Ecotoxicol. Environ. Saf. 2022, 231, 113203. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.; Pälmke, C.; Ringbeck, B.; Ihn, Y.; Gotthardt, A.; Lee, G.; Alakeel, R.; Alrashed, M.; Tosepu, R.; Jayadipraja, E.A.; et al. Urinary concentrations of major phthalate and alternative plasticizer metabolites in children of Thailand, Indonesia, and Saudi Arabia, and associated risks. Environ. Sci. Technol. 2021, 55, 16526–16537. [Google Scholar] [CrossRef] [PubMed]
- Sheikh, I.A.; Beg, M.A. Structural characterization of potential endocrine disrupting activity of alternate plasticizers di-(2-ethylhexyl) adipate (DEHA), acetyl tributyl citrate (ATBC) and 2,2,4-trimethyl 1,3-pentanediol diisobutyrate (TPIB) with human sex hormone-binding globulin. Reprod. Toxicol. 2019, 83, 46–53. [Google Scholar] [PubMed]
- Park, J.; Park, C.; Gye, M.C.; Lee, Y. Assessment of endocrine-disrupting activities of alternative chemicals for bis(2-ethylhexyl)phthalate. Environ. Res. 2019, 172, 10–17. [Google Scholar] [CrossRef]
- Moche, H.; Chentouf, A.; Neves, S.; Corpart, J.M.; Nesslany, F. Comparison of in vitro endocrine activity of phthalates and alternative plasticizers. J. Toxicol. 2021, 2021, 8815202. [Google Scholar] [CrossRef]
- Pacyga, D.C.; Gardiner, J.C.; Flaws, J.A.; Li, Z.; Calafat, A.M.; Korrick, S.A.; Schantz, S.L.; Strakovsky, R.S. Maternal phthalate and phthalate alternative metabolites and urinary biomarkers of estrogens and testosterones across pregnancy. Environ. Int. 2021, 155, 106676. [Google Scholar] [CrossRef]
- Van T Erve, T.J.; Rosen, E.M.; Barrett, E.S.; Nguyen, R.H.; Sathyanarayana, S.; Milne, G.L.; Calafat, A.M.; Swan, S.H.; Ferguson, K.K. Phthalates and phthalate alternatives have diverse associations with oxidative stress and inflammation in pregnant women. Environ. Sci. Technol. 2019, 53, 3258–3267. [Google Scholar] [CrossRef]
- Rasmussen, L.M.; Sen, N.; Liu, X.; Craig, Z.R. Effects of oral exposure to the phthalate substitute acetyl tributyl citrate on female reproduction in mice. J. Appl. Toxicol. 2017, 37, 668–675. [Google Scholar] [CrossRef]
- Eljezi, T.; Pinta, P.; Richard, D.; Pinguet, J.; Chezal, J.M.; Chagnon, M.C.; Sautou, V.; Grimandi, G.; Moreau, E. In vitro cytotoxic effects of DEHP-Alternative plasticizers and their primary metabolites on a L929 cell line. Chemosphere 2017, 173, 452–459. [Google Scholar] [CrossRef]
- Sheikh, I.A.; Beg, M.A. Structural binding perspectives of common plasticizers and a flame retardant BDE-153 against thyroxine binding globulin: Potential for endocrine disruption. J. Appl. Toxicol. 2021, 42, 841–851. [Google Scholar] [CrossRef]
- Madariaga, A.G.; Palacios, S.S.; Guillén-Grima, F.; Galofre, J.C. The incidence and prevalence of thyroid dysfunction in Europe: A meta-analysis. J. Clin. Endocrinol. Metab. 2014, 99, 923–931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Souter, I.; Bellavia, A.; Williams, P.L.; Korevaar, T.I.M.; Meeker, J.D.; Braun, J.M.; de Poortere, R.A.; Broeren, M.A.; Ford, J.B.; Calafat, A.M.; et al. Urinary concentrations of phthalate metabolite mixtures in relation to serum biomarkers of thyroid function and autoimmunity among women from a fertility center. Environ. Health Perspect. 2020, 128, 67007. [Google Scholar] [CrossRef] [PubMed]
- Duarte-Guterman, P.; Navarro-Martín, L.; Trudeau, V.L. Mechanisms of crosstalk between endocrine systems: Regulation of sexsteroid hormone synthesis and action by thyroid hormones. Gen. Comp. Endocrinol. 2014, 203, 69–85. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Zhang, W.; Zhang, Y.; Kang, Z.; Miao, X.; Na, X. Novel insights into di-(2-ethylhexyl)phthalate activation: Implications for the hypothalamus-pituitary-thyroid axis. Mol. Med. Rep. 2021, 23, 290. [Google Scholar] [CrossRef]
- Zhang, X.; Qi, W.; Xu, Q.; Li, X.; Zhou, L.; Ye, L. Di(2-ethylhexyl) phthalate (DEHP) and thyroid: Biological mechanisms of interference and possible clinical implications. Environ. Sci. Pollut. Res. Int. 2022, 29, 1634–1644. [Google Scholar] [CrossRef]
- Derakhshan, A.; Shu, H.; Broeren, M.A.C.; Lindh, C.H.; Peeters, R.P.; Kortenkamp, A.; Demeneix, B.; Bornehag, C.G.; Korevaar, T.I.M. Association of phthalate exposure with thyroid function during pregnancy. Environ. Int. 2021, 157, 106795. [Google Scholar] [CrossRef]
- FCA 2016. 1,2-Cyclohexanedicarboxylic Acid, Diisononyl Ester (DINCH). Analysis of the Most Appropriate Risk Management Option (RMOA). Prepared by the French Agency for Food, Environmental and Occupational Health and Safety (Anses) under French National Strategy on Endocrine Disruptors for French Competent Authority, France. Available online: https://echa.europa.eu/documents/10162/9f62e492-4028-5700-e5ec-019c396b13bf (accessed on 10 March 2022).
- Ghisari, M.; Bonefeld-Jorgensen, E.C. Effects of plasticizers and their mixtures on estrogen receptor and thyroid hormone functions. Toxicol. Lett. 2009, 189, 67–77. [Google Scholar] [CrossRef]
- Huang, H.-B.; Chuang, C.-J.; Su, P.-H.; Sun, C.-W.; Wang, C.-J.; Wu, M.-T.; Wang, S.-L. Prenatal and Childhood Exposure to Phthalate Diesters and Thyroid Function in a 9-Year Follow-up Birth Cohort Study: Taiwan Maternal and Infant Cohort Study. Epidemiol. Camb. Mass. 2017, 28, S10–S18. [Google Scholar] [CrossRef]
- Kambia, N.; Séverin, I.; Farce, A.; Dahbi, L.; Dine, T.; Moreau, E.; Sautou, V.; Chagnon, M.C. Comparative Effects of Di-(2-ethylhexyl)phthalate and Di-(2-ethylhexyl)terephthalate Metabolites on Thyroid Receptors: In Vitro and In Silico Studies. Metabolites 2021, 11, 94. [Google Scholar] [CrossRef]
- Sun, D.; Zhou, L.; Wang, S.; Liu, T.; Zhu, J.; Jia, Y.; Xu, J.; Chen, H.; Wang, Q.; Xu, F.; et al. Effect of Di-(2-ethylhexyl) phthalate on the hypothalamus-pituitary-thyroid axis in adolescent rat. Endocr. J. 2022, 69, 217–224. [Google Scholar] [CrossRef]
- Sugiyama, S.; Shimada, N.; Miyoshi, H.; Yamauchi, K. Detection of thyroid system-disrupting chemicals using in vitro and in vivo screening assays in Xenopus laevis. Toxicol. Sci. 2005, 88, 367–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, H.; Wu, W.; Xu, Y.; Jin, Z.; Bao, H.; Zhu, P.; Su, P.; Sheng, J.; Hao, J.; Tao, F. Effects of Prenatal Phthalate Exposure on Thyroid Hormone Concentrations Beginning at The Embryonic Stage. Sci. Rep. 2017, 7, 13106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.J.; Moon, S.; Oh, B.-C.; Jung, D.; Choi, K.; Park, Y.J. Association between Diethylhexyl Phthalate Exposure and Thyroid Function: A Meta-Analysis. Thyroid 2019, 29, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Johns, L.E.; Ferguson, K.K.; McElrath, T.F.; Mukherjee, B.; Meeker, J.D. Associations between Repeated Measures of Maternal Urinary Phthalate Metabolites and Thyroid Hormone Parameters during Pregnancy. Environ. Health Perspect. 2016, 124, 1808–1815. [Google Scholar] [CrossRef] [Green Version]
- Qian, X.; Li, J.; Xu, S.; Wan, Y.; Li, Y.; Jiang, Y.; Zhao, H.; Zhou, Y.; Liao, J.; Liu, H.; et al. Prenatal exposure to phthalates and neurocognitive development in children at two years of age. Environ. Int. 2019, 131, 105023. [Google Scholar] [CrossRef]
- Sherif, N.A.E.; El-Banna, A.; Abdel-Moneim, R.A.; Sobh, Z.K.; Balah, M.I.F. The possible thyroid disruptive effect of di-(2-ethyl hexyl) phthalate and the potential protective role of selenium and curcumin nanoparticles: A toxicological and histological study. Toxicol. Res. (Camb.) 2021, 11, 108–121. [Google Scholar] [CrossRef]
- Kong, D.; Li, J.; Li, N.; Zhang, S.; Xu, Y. Multiple bioanalytical methods reveal a thyroid-disrupting mechanism related to the membrane receptor integrin alpha(v)beta(3). Environ. Pollut. 2021, 279, 116933. [Google Scholar] [CrossRef]
- Josh, M.K.S.; Pradeep, S.; Adarsh, V.K.; Amma, K.S.V.; Devi, R.S.; Balachandran, S.; Sreejith, M.N.; Jaleel, U.C.A.; Benjamin, S. In silico evidences for the binding of phthalates onto human estrogen receptor α, β subtypes and human estrogen-related receptor γ. Mol. Simul. 2013, 40, 408–417. [Google Scholar] [CrossRef]
- Kambia, N.; Farce, A.; Belarbi, K.; Gressier, B.; Luyckx, M.; Chavatte, P.; Dine, T. Docking study: PPARs interaction with the selected alternative plasticizers to di(2-ethylhexyl) phthalate. J. Enzym. Inhib. Med. Chem. 2016, 31, 448–455. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Chen, J.A.; Qiu, Z.; Zhao, Q.; Luo, J.; Yang, L.; Zeng, H.; Huang, Y.; Zhang, L.; Cao, J.; et al. Ovotoxicity and PPAR-mediated aromatase downregulation in female Sprague-Dawley rats following combined oral exposure to benzo[a]pyrene and di-(2-ethylhexyl) phthalate. Toxicol. Lett. 2010, 199, 323–332. [Google Scholar] [CrossRef]
- Huang, R.; Xia, M.; Cho, M.H.; Sakamuru, S.; Shinn, P.; Houck, K.A.; Dix, D.J.; Judson, R.S.; Witt, K.L.; Kavlock, R.J.; et al. Chemical genomics profiling of environmental chemical modulation of human nuclear receptors. Env. Health Perspect. 2011, 119, 1142–1148. [Google Scholar] [CrossRef] [PubMed]
- Takeshita, A.; Igarashi-Migitaka, J.; Nishiyama, K.; Takahashi, H.; Takeuchi, Y.; Koibuchi, N. Acetyl tributyl citrate, the most widely used phthalate substitute plasticizer, induces cytochrome p450 3a through steroid and xenobiotic receptor. Toxicol. Sci. 2011, 123, 460–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engel, A.; Buhrke, T.; Kasper, S.; Behr, A.C.; Braeuning, A.; Jessel, S.; Seidel, A.; Völkel, W.; Lampen, A. The urinary metabolites of DINCH have an impact on the activities of the human nuclear receptors ER, ER, AR, PPAR and PPAR. Toxicol. Lett. 2018, 287, 83–91. [Google Scholar] [CrossRef] [PubMed]
- ToxServices. GreenScreen Assessment for Hexamoll DINCH 1 (Diisononyl cyclohexanedicarboxylate) (CAS #166412-78-8, 474919-59-0). Clean Production Action Content February 2012. 2012, pp. 1–17. Available online: https://www.greenchemistryandcommerce.org/documents/Hexamoll-DINCHGS5.28.13.pdf (accessed on 15 March 2022).
- Kwon, B.; Ji, K. Estrogenic and Androgenic Potential of Phthalates and Their Alternatives. J. Environ. Health Sci. 2016, 42, 169–188. [Google Scholar] [CrossRef] [Green Version]
- Campioli, E.; Lee, S.; Lau, M.; Marques, L.; Papadopoulos, V. Effect of prenatal DINCH plasticizer exposure on rat offspring testicular function and metabolism. Sci. Rep. 2017, 7, 11072. [Google Scholar] [CrossRef] [PubMed]
- Albert, O.; Nardelli, T.C.; Lalancette, C.; Hales, B.F.; Robaire, B. Effects of In Utero and Lactational Exposure to New Generation Green Plasticizers on Adult Male Rats: A Comparative Study with Di(2-Ethylhexyl) Phthalate. Toxicol. Sci. 2018, 164, 129–141. [Google Scholar] [CrossRef] [PubMed]
- Saad, N.; Bereketoglu, C.; Pradhan, A. Di(Isononyl) cyclohexane-1,2-dicarboxylate (DINCH) alters transcriptional profiles, lipid metabolism and behavior in zebrafish larvae. Heliyon 2021, 7, e07951. [Google Scholar] [CrossRef]
- Vasconcelos, A.L.; Silva, M.J.; Louro, H. In Vitro Exposure to the Next-Generation Plasticizer Diisononyl Cyclohexane-1,2-Dicarboxylate (DINCH): Cytotoxicity and Genotoxicity Assessment in Human Cells. J. Toxicol. Environ. Health Part A Curr. Issues 2019, 82, 526–536. [Google Scholar] [CrossRef]
- CIR. Safety Assessment of Acetyl Trialkyl Citrates as Used in Cosmetics. Cosmetic Ingredient Review Washington, DC. 2019. Available online: https://www.cir-safety.org/sites/default/files/Acetyl%20Trialkyl%20Citrates.pdf (accessed on 1 February 2022).
- Strajhar, P.; Tonoli, D.; Jeanneret, F.; Imhof, R.M.; Malagnino, V.; Patt, M.; Kratschmar, D.V.; Boccard, J.; Rudaz, S.; Odermatt, A. Steroid profiling in H295R cells to identify chemicals potentially disrupting the production of adrenal steroids. Toxicology 2017, 381, 51–63. [Google Scholar] [CrossRef]
- Stuer-Lauridsen, F.; Mikkelsen, S.; Havelund, M.; Birkved, M.; Hansen, L.P. Environmental and Health Assessment of Alternatives to Phthalates and to Flexible PVC. Danish Environmental Protection Agency, COWI Consulting Engineers and Planners AS. Lyngby, Denmark. 2001. Available online: http://www2.mst.dk/udgiv/publications/2001/87-7944-407-5/pdf/87-7944-408-3.pdf (accessed on 25 January 2022).
- Hirata-Koizumi, M.; Takahashi, M.; Matsumoto, M.; Kawamura, T.; Ono, A.; Hirose, A. Toxicity effects of phthalate substitute plasticizers used in toys. Kokuritsu Iyakuhin Shokuhin Eisei Kenkyusho Hokoku 2012, 130, 31–42. [Google Scholar]
- Dalgaard, M.; Hass, U.; Vinggaard, A.M.; Jarfelt, K.; Lam, H.R.; Sorensen, I.K.; Sommer, H.M.; Ladefoged, O. Di(2-ethylhexyl) adipate (DEHA) induced developmental toxicity but not antiandrogenic effects in pre- and postnatally exposed Wistar rats. Reprod. Toxicol. 2003, 17, 163–170. [Google Scholar] [CrossRef]
- Lee, H.C.; Yamanouchi, K.; Nishihara, M. Effects of perinatal exposure to phthalate/adipate esters on hypothalamic gene expression and sexual behavior in rats. J. Reprod. Dev. 2006, 52, 343–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CPSC. Toxicity Review for Bis(2-Ethylhexyl) Adipate (DEHA). Contract No. CPSC-D-17-0001 Task Order No. 003, Risk Science Center, Department of Environmental Health, University of Cincinnati, Cincinnati, OH. Prepared for U.S. Consumer Product Safety Commission. 2018. Available online: https://www.cpsc.gov/s3fspublic/Toxicity%20Review%20of%20DEHA.pdf (accessed on 10 February 2022).




{kind=link}
{kind=link}
{kind=link}
| Ligand | Number of Interacting TRα Residues | Percentage of Interacting Residues Common with Native Ligand (%) | IFD Score | Docking Score (Kcal/mol) | Glide Score (Kcal/mol) | MMGB-SA (Kcal/mol) |
|---|---|---|---|---|---|---|
| DINCH | 30 | 95.45 | −563.87 | −9.53 | −9.53 | −156.49 |
| ATBC | 27 | 78.26 | −564.56 | −8.89 | −8.89 | −140.29 |
| DEHA | 28 | 86.95 | −564.36 | −7.96 | −7.96 | −130.04 |
| DEHP | 24 | 78.26 | −562.08 | −8.64 | −8.64 | −131.67 |
| T3 | 23 | 100 | −564.42 | −9.44 | −9.44 | −133.53 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zughaibi, T.A.; Sheikh, I.A.; Beg, M.A. Insights into the Endocrine Disrupting Activity of Emerging Non-Phthalate Alternate Plasticizers against Thyroid Hormone Receptor: A Structural Perspective. Toxics 2022, 10, 263. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics10050263
Zughaibi TA, Sheikh IA, Beg MA. Insights into the Endocrine Disrupting Activity of Emerging Non-Phthalate Alternate Plasticizers against Thyroid Hormone Receptor: A Structural Perspective. Toxics. 2022; 10(5):263. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics10050263
Chicago/Turabian StyleZughaibi, Torki A., Ishfaq Ahmad Sheikh, and Mohd Amin Beg. 2022. "Insights into the Endocrine Disrupting Activity of Emerging Non-Phthalate Alternate Plasticizers against Thyroid Hormone Receptor: A Structural Perspective" Toxics 10, no. 5: 263. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics10050263