Astrocytes Are More Vulnerable than Neurons to Silicon Dioxide Nanoparticle Toxicity in Vitro

 , , ,
, , ,  ,
,  ,
,  , and
, and 
Abstract
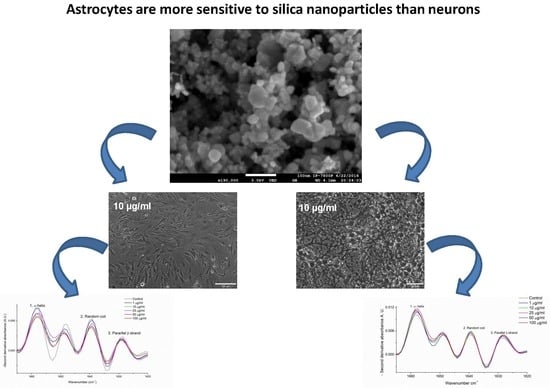
:
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Bioethics
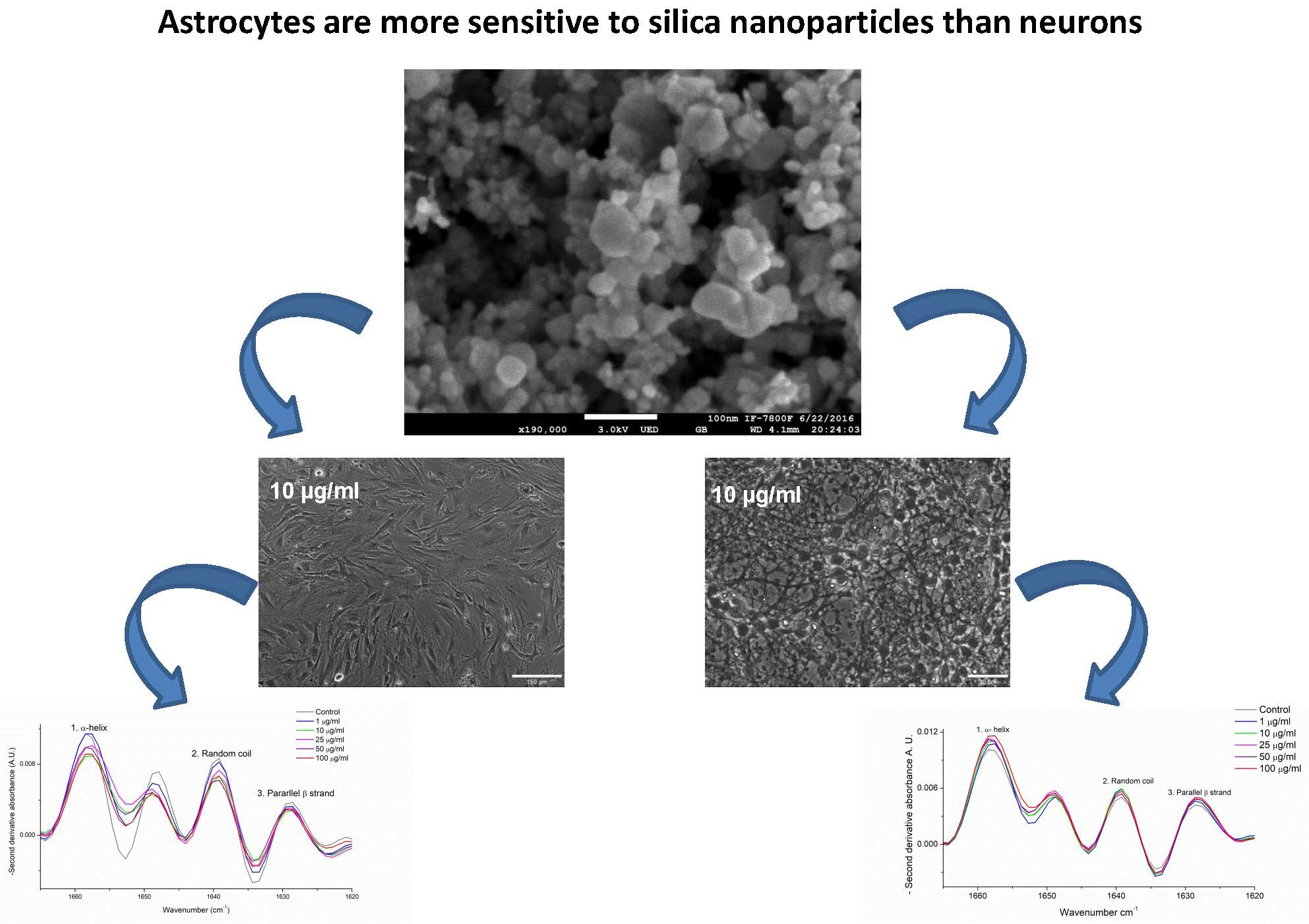
2.3. Scanning Electron Microscopy (SEM) and Size Distribution
2.4. SiO2-NPs Hydrodynamic Size, Polydispersion Index and Zeta (ζ) Potential
2.5. Secondary Culture of Rat Cerebellar Astrocytes
2.6. Primary Culture of Rat Cerebellar Granule Neurons
2.7. Concentration-Response Analysis to SiO2-NPs
2.8. ATR-FTIR Spectroscopy
2.9. FTIR Microspectroscopy Mapping (IQ Mapping)
2.10. Cell Viability Tests
2.11. Statistical Analysis
3. Results
3.1. FTIR and SEM Characterization of SiO2-NPs
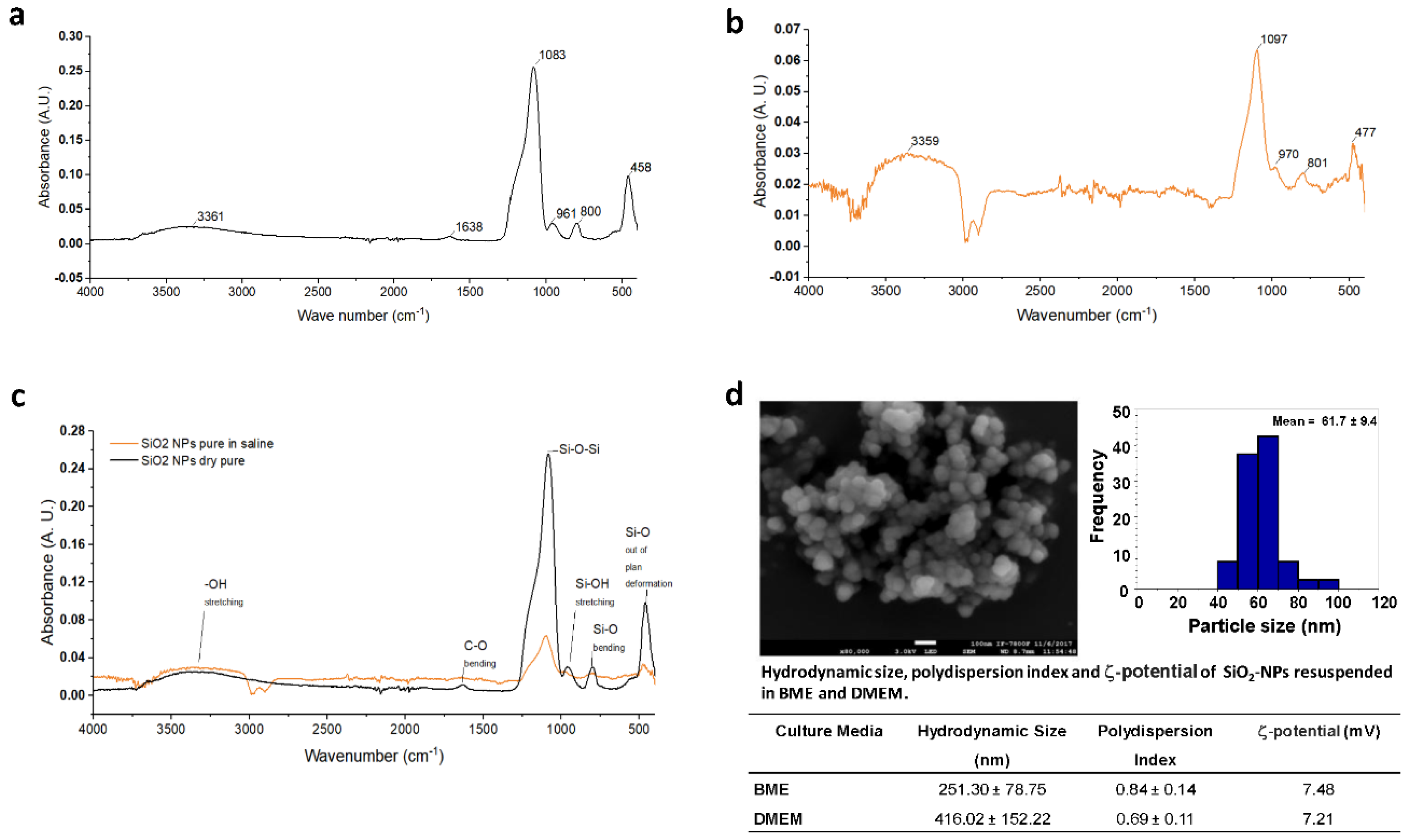
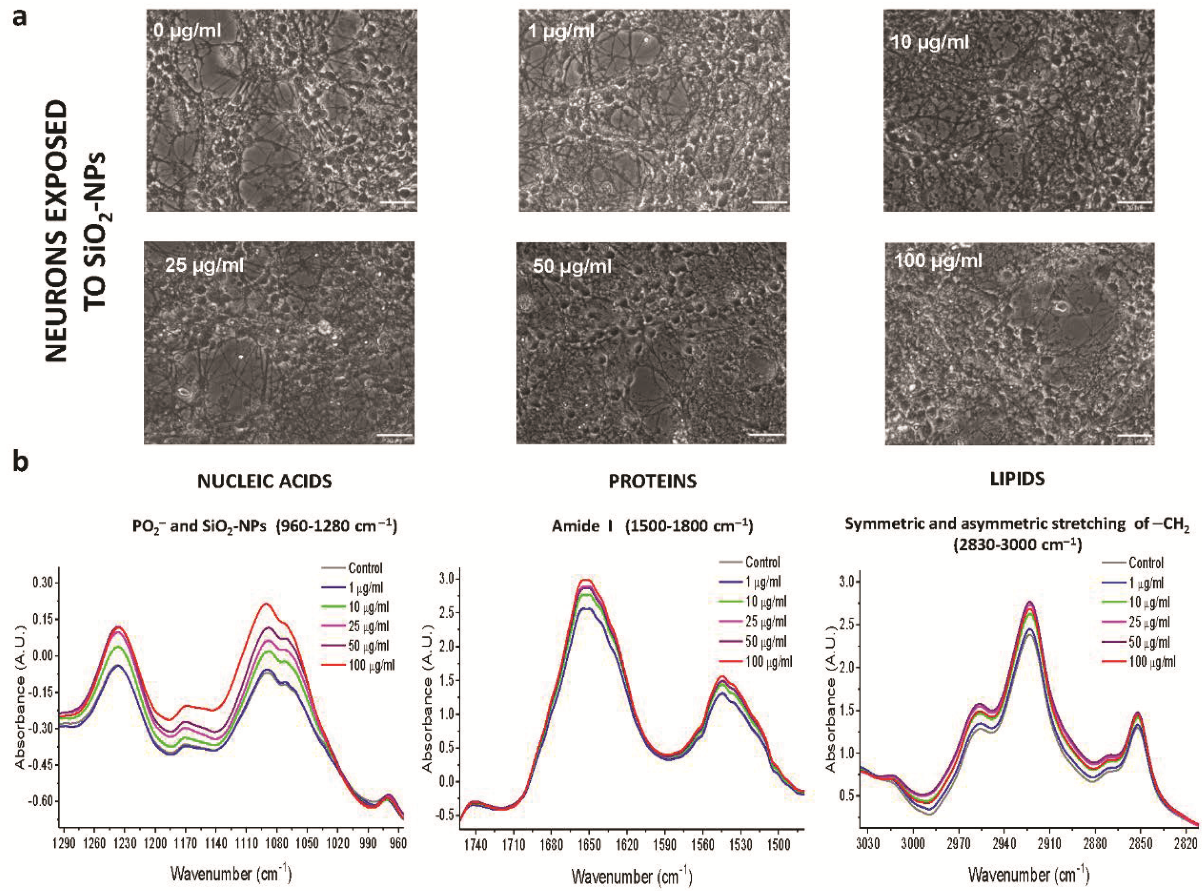
3.2. Morphological Changes of Rat Cerebellar Astrocytes and Neurons Exposed to SiO2-NPs
3.3. ATR-FTIR Spectra of Rat Cerebellar Astrocytes and Neurons Exposed to SiO2-NPs
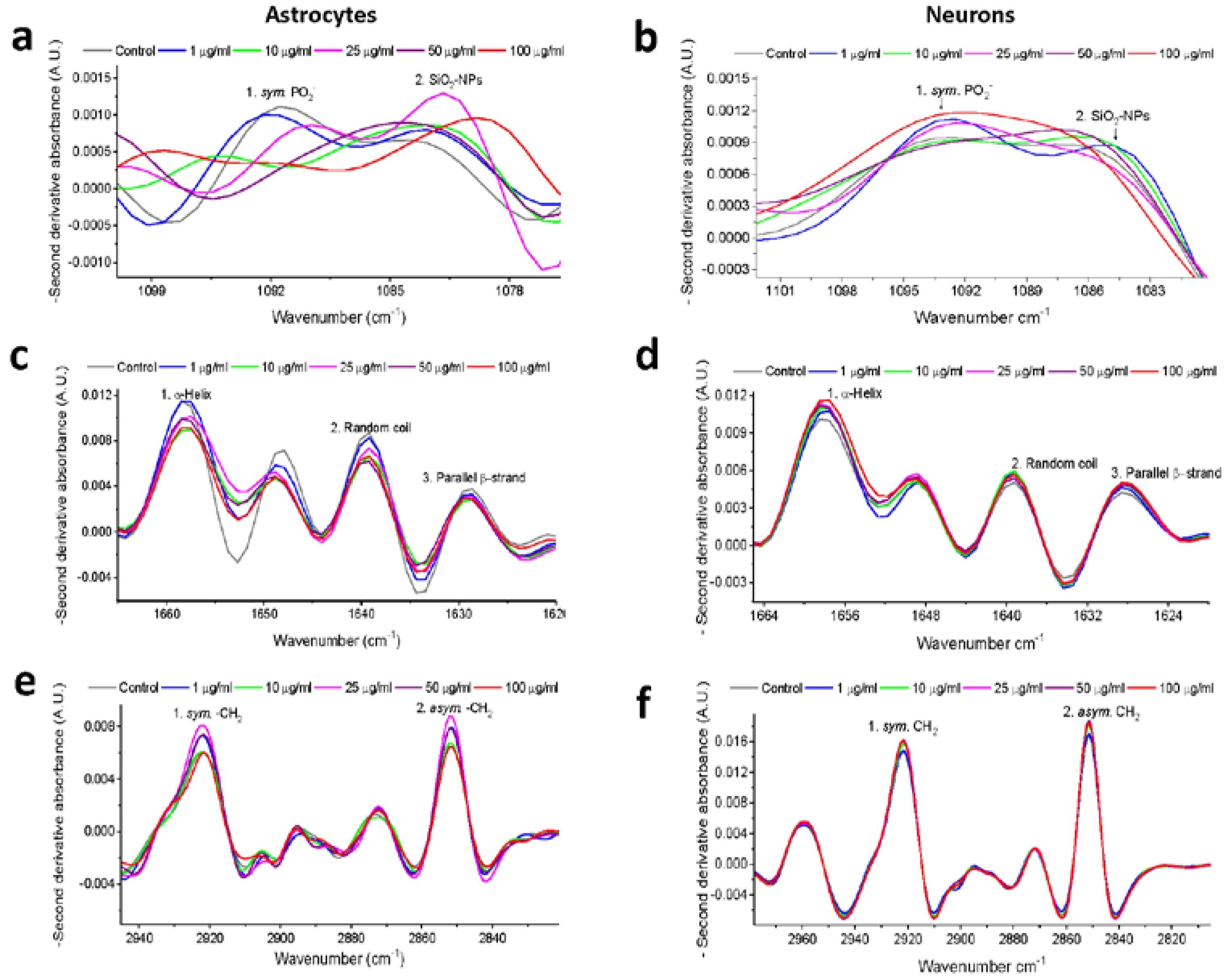
3.4. Second Derivative Analyses of Spectral Bands
3.5. Second Derivative Analyses of the Nucleic Acid Spectral Region
3.6. Second Derivative Analyses of the Protein Spectral Region
3.7. Second Derivative Analyses of the Lipid Spectral Region
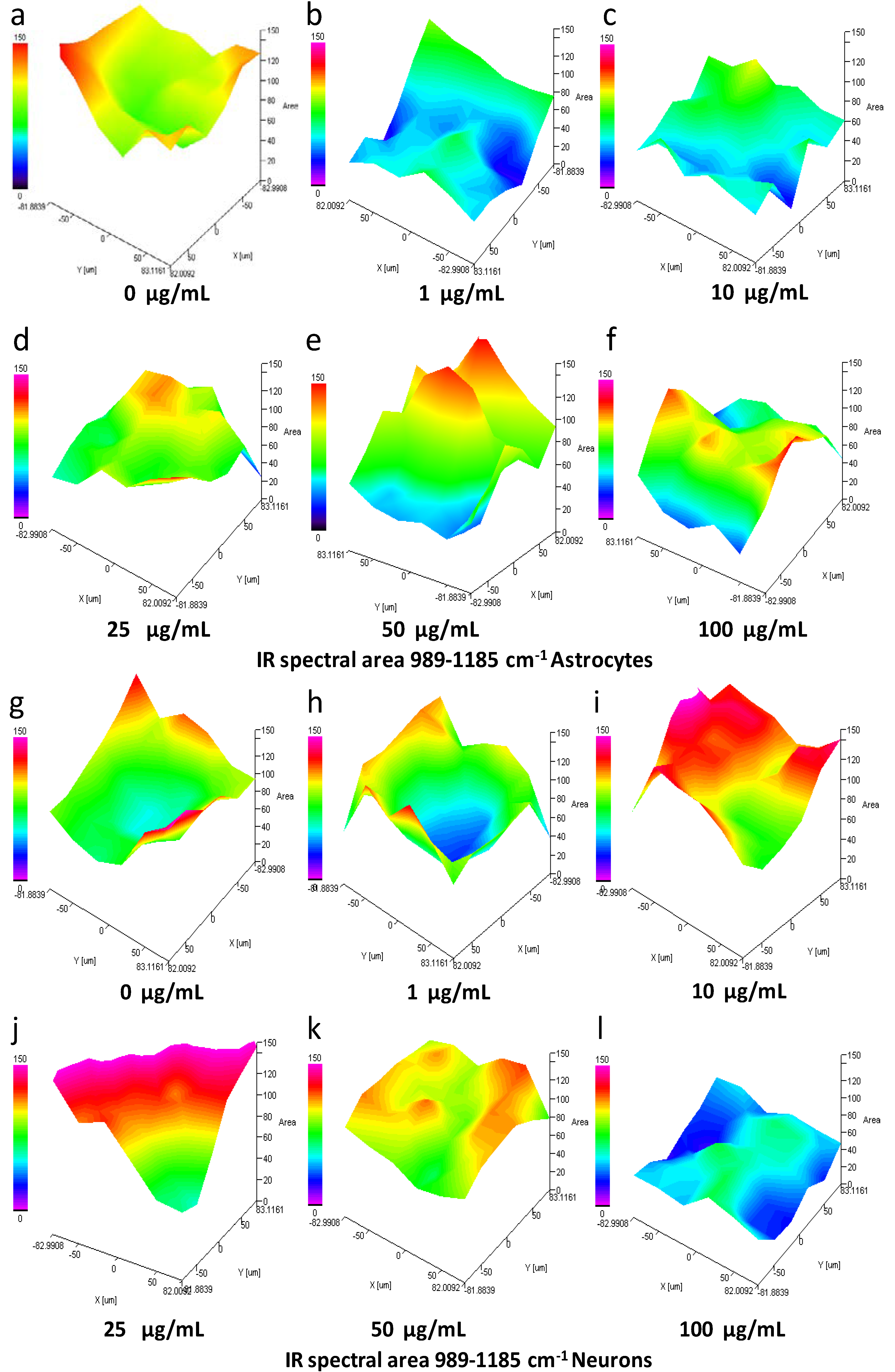
3.8. IQ mapping of Nucleic Acid Region 989–1185 cm−1 in Astrocytes and Neurons after Exposure to SiO2-NPs
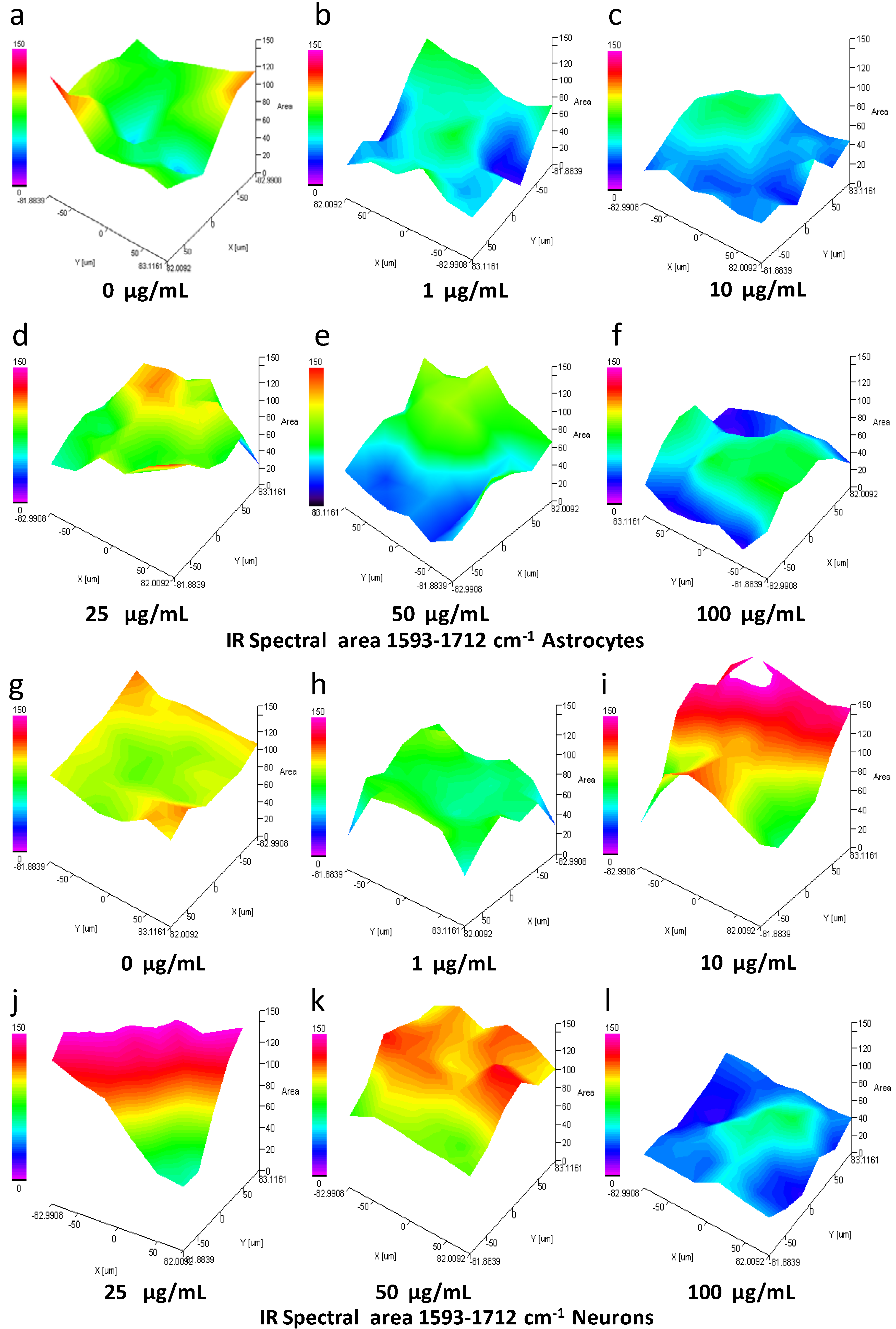
3.9. IQ Mapping of the Protein Amide I Region 1593–1712 cm−1 in Astrocytes and Neurons after Exposure to SiO2-NPs
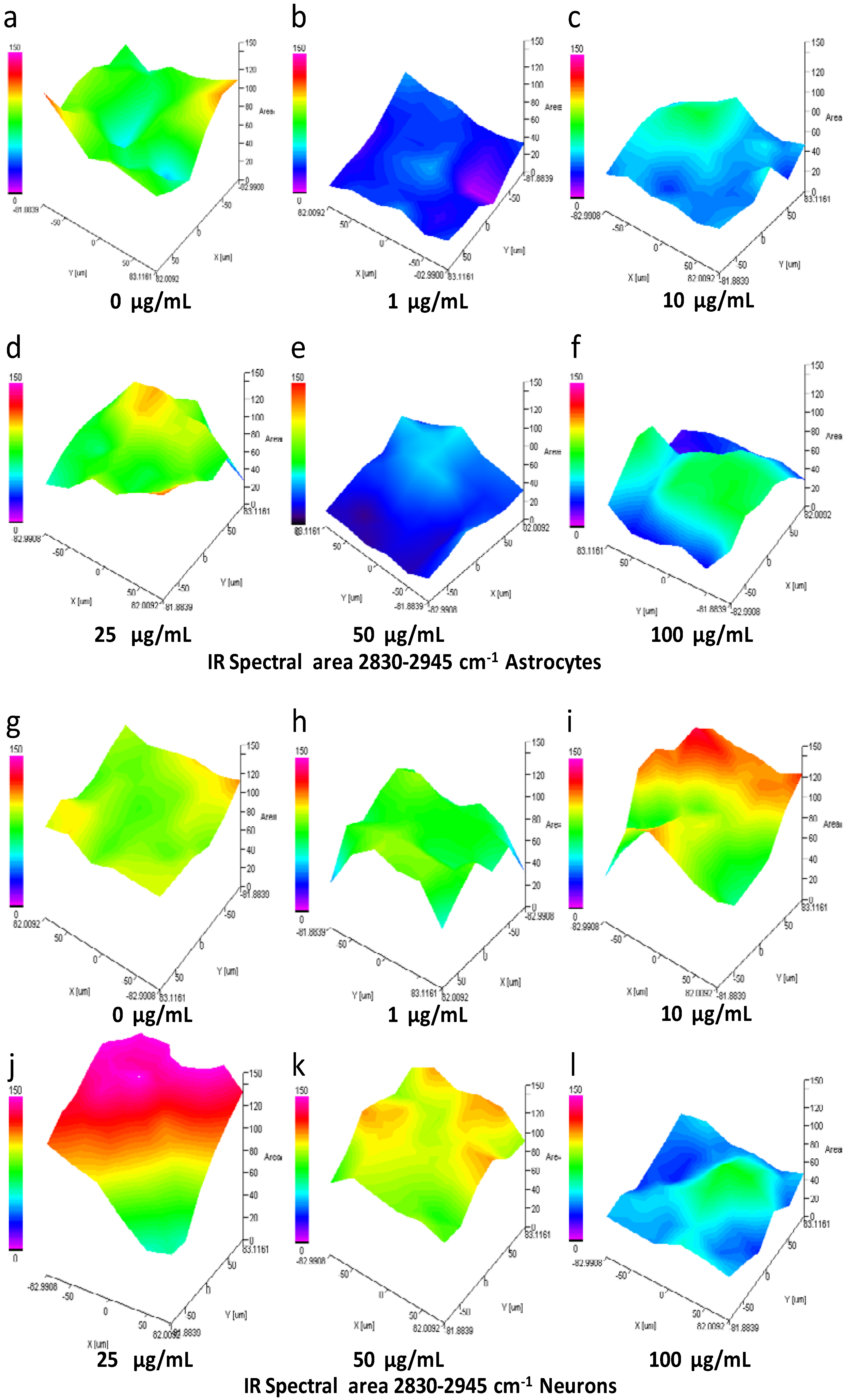
3.10. IQ Mapping of the Lipid Region 2830-2945 cm−1 in Astrocytes and Neurons after Exposure of SiO2-NPs
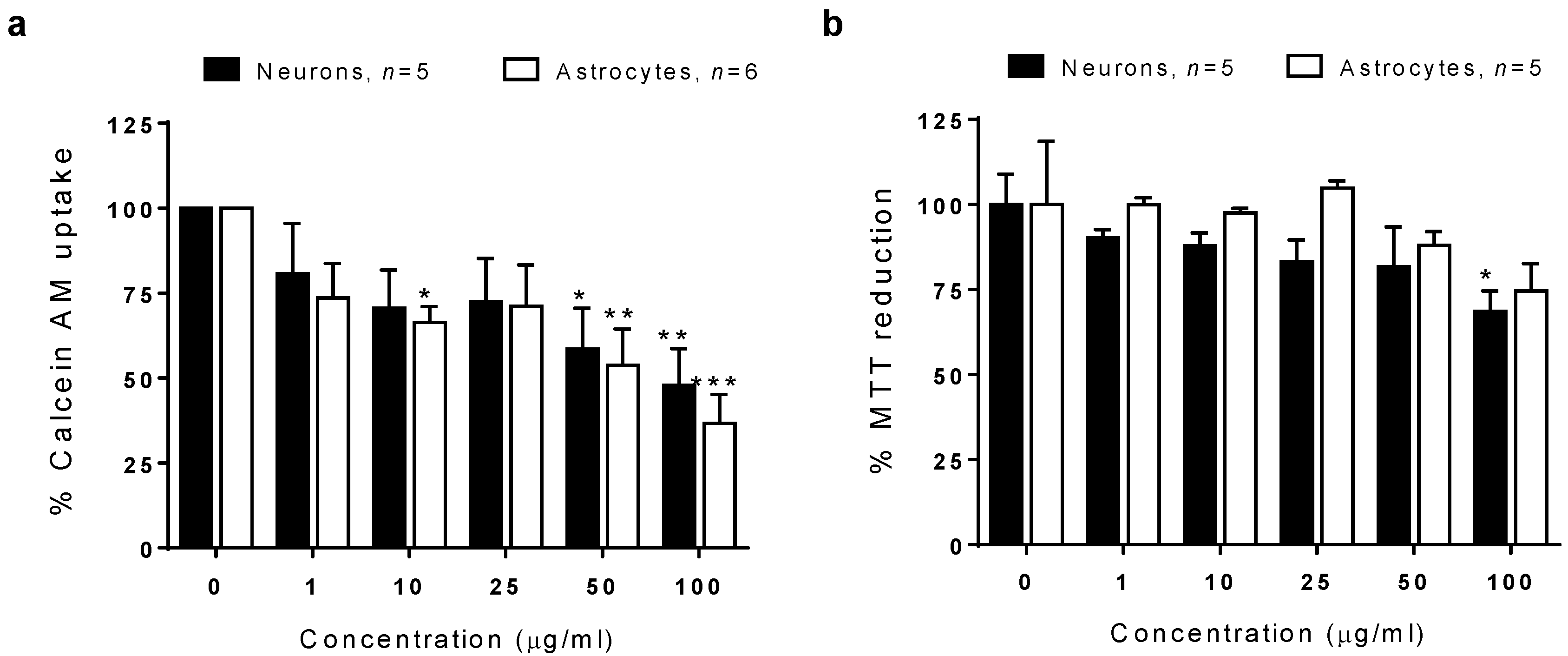
3.11. Cell Viability Determination
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dekkers, S.; Krystek, P.; Peters, R.J.; Lankveld, D.P.; Bokkers, B.G.; van Hoeven-Arentzen, P.H.; Bouwmeester, H.; Oomen, A.G. Presence and risks of nanosilica in food products. Nanotoxicology 2011, 5, 393–405. [Google Scholar] [CrossRef]
- Younes, M.; Aggett, P.; Aguilar, F.; Crebelli, R.; Dusemund, B.; Filipic, M. Re-evaluation of silicon dioxide (E 551) as a food additive. EFSA J. 2018, 16, e05088. [Google Scholar] [CrossRef] [Green Version]
- Napierska, D.; Thomassen, L.C.J.; Lison, D.; Martens, J.A.; Hoet, P.H. The nanosilica hazard: Another variable entity. Part. Fibre Toxicol. 2010, 7, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turci, F.; Pavan, C.; Leinardi, R.; Tomatis, M.; Pastero, L.; Garry, D. Revisiting the paradigm of silica pathogenicity with synthetic quartz crystals: The role of crystallinity and surface disorder. Part. Fibre Toxicol. 2016, 13, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murugadoss, S.; Lison, D.; Godderis, L.; Van den Brule, S.; Mast, J.; Brassinne, F. Toxicology of silica nanoparticles: An update. Arch Toxicol. 2017, 91, 2967–3010. [Google Scholar] [CrossRef] [PubMed]
- Du, Z.J.; Chen, S.Y.; Cui, G.Q.; Yang, Y.; Zhang, E.G.; Wang, Q. Silica nanoparticles induce cardiomyocyte apoptosis via the mitochondrial pathway in rats following intratracheal instillation. Int. J. Mol. Med. 2019, 43, 1229–1240. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, K.; Yoshioka, Y.; Higashisaka, K.; Mimura, K.; Morishita, Y.; Nozaki, M. Silica and titanium dioxide nanoparticles cause pregnancy complications in mice. Nat. Nanotechnol. 2011, 6, 321–328. [Google Scholar] [CrossRef]
- Wu, J.; Wang, C.; Sun, J.; Xue, Y. Neurotoxicity of Silica Nanoparticles: Brain Localization and Dopaminergic Neurons Damage Pathways. ACS Nano 2011, 5, 4476–4489. [Google Scholar] [CrossRef]
- Parveen, A.; Rizvi, S.H.M.; Mahdi, F.; Tripathi, S.; Ahmad, I.; Shukla, R.K. Silica nanoparticles mediated neuronal cell death in corpus striatum of rat brain: Implication of mitochondrial, endoplasmic reticulum and oxidative stress. J. Nanopart. Res. 2014, 16, 2664. [Google Scholar] [CrossRef]
- Parveen, A.; Rizvi, S.H.M.; Sushma; Mahdi, F.; Ahmad, I.; Singh, P.P. Intranasal exposure to silica nanoparticles induces alterations in pro-inflammatory environment of rat brain: Involvement of oxidative stress. Toxicol. Ind. Health 2017, 33, 119–132. [Google Scholar] [CrossRef]
- Liu, X.; Sui, B.; Sun, J. Blood-brain barrier dysfunction induced by silica NPs in vitro and in vivo: Involvement of oxidative stress and Rho-kinase/JNK signaling pathways. Biomaterials 2017, 121, 64–82. [Google Scholar] [CrossRef] [PubMed]
- Mourdikoudis, S.; Pallares, R.M.; Thanh, N.T.K. Characterization techniques for nanoparticles: Comparison and complementarity upon studying nanoparticle properties. Nanoscale 2018, 10, 12871–12934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.Y.; Yang, S.N.; Kong, J.L.; Dong, A.C.; Yu, S.N. Obtaining information about protein secondary structures in aqueous solution using Fourier transform IR spectroscopy. Nat. Protoc. 2015, 10, 382–396. [Google Scholar] [CrossRef] [PubMed]
- Baker, M.J.; Trevisan, J.; Bassan, P.; Bhargava, R.; Butler, H.J.; Dorling, K.M. Using Fourier transform IR spectroscopy to analyze biological materials. Nat. Protoc. 2014, 9, 1771–1791. [Google Scholar] [CrossRef] [Green Version]
- Escudero-Lourdes, C.; Uresti-Rivera, E.E.; Oliva-González, C.; Torres-Ramos, M.A.; Aguirre-Bañuelos, P.; Gandolfi, A.J. Cortical Astrocytes Acutely Exposed to the Monomethylarsonous Acid (MMAIII) Show Increased Pro-inflammatory Cytokines Gene Expression that is Consistent with APP and BACE-1: Over-expression. Neurochem. Res. 2016, 41, 2559–2572. [Google Scholar] [CrossRef] [Green Version]
- Orozco-Ibarra, M.; Estrada-Sanchez, A.M.; Massieu, L.; Pedraza-Chaverri, J. Heme oxygenase-1 induction prevents neuronal damage triggered during mitochondrial inhibition: Role of CO and bilirubin. Int. J. Biochem. Cell Biol. 2009, 41, 1304–1314. [Google Scholar] [CrossRef]
- Vazquez-Zapien, G.J.; Mata-Miranda, M.M.; Sanchez-Monroy, V.; Delgado-Macuil, R.J.; Perez-Ishiwara, D.G.; Rojas-Lopez, M. FTIR Spectroscopic and Molecular Analysis during Differentiation of Pluripotent Stem Cells to Pancreatic Cells. Stem Cells Int. 2016, 2016, 6709714. [Google Scholar] [CrossRef] [Green Version]
- Mata-Miranda, M.M.; Guerrero-Ruiz, M.; Gonzalez-Fuentes, J.R.; Hernandez-Toscano, C.M.; Garcia-Andino, J.R.; Sanchez-Brito, M.; Vazquez-Zapien, G.J. Characterization of the Biological Fingerprint and Identification of Associated Parameters in Stress Fractures by FTIR Spectroscopy. Biomed. Res. Int. 2019, 2019, 1241452. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Reyes, S.; Orozco-Ibarra, M.; Guzman-Beltran, S.; Molina-Jijon, E.; Massieu, L.; Pedraza-Chaverri, J. Neuroprotective role of heme-oxygenase 1 against iodoacetate-induced toxicity in rat cerebellar granule neurons: Role of bilirubin. Free Radic. Res. 2009, 3, 214–223. [Google Scholar] [CrossRef]
- Tejaswi, J.; Anirudh, K.V.S.; Majeti, L.R.; Kotagiri, D.; Shaik, K.B.; Chaitanya, K.V. Investigation of Biological Activity of Nanoparticles Using Cell Lines. In Model Organisms to Study Biological Activities and Toxicity of Nanoparticles; Siddhardha, B., Dyavaiah, M., Kasinathan, K., Eds.; Springer Nature: Berlin, Germany, 2020; pp. 117–138. [Google Scholar]
- Rieppo, L.; Saarakkala, S.; Narhi, T.; Helminen, H.J.; Jurvelin, J.S.; Rieppo, J. Application of second derivative spectroscopy for increasing molecular specificity of fourier transform infrared spectroscopic imaging of articular cartilage. Osteoarthr. Cartil. 2012, 20, 451–459. [Google Scholar] [CrossRef] [Green Version]
- Zelig, U.; Kapelushnik, J.; Moreh, R.; Mordechai, S.; Nathan, I. Diagnosis of Cell Death by Means of Infrared Spectroscopy. Biophys. J. 2009, 97, 2107–2114. [Google Scholar] [CrossRef] [Green Version]
- Petibois, C.; Deleris, G. Chemical mapping of tumor progression by FT-IR imaging: Towards molecular histopathology. Trends Biotechnol. 2006, 24, 455–462. [Google Scholar] [CrossRef] [PubMed]
- Kimber, J.A.; Kazarian, S.G. Spectroscopic imaging of biomaterials and biological systems with FTIR microscopy or with quantum cascade lasers. Anal. Bioanal. Chem. 2017, 409, 5813–5820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petibois, C.; Wehbe, K.; Belbachir, K.; Noreen, R.; Deleris, G. Current Trends in the Development of FTIR Imaging for the Quantitative Analysis of Biological Samples. Acta Phys. Pol. A 2009, 115, 507–512. [Google Scholar] [CrossRef]
- Fisichella, M.; Dabboue, H.; Bhattacharyya, S.; Saboungi, M.L.; Salvetat, J.P.; Hevor, T.; Guerin, M. Mesoporous silica nanoparticles enhance MTT formazan exocytosis in HeLa cells and astrocytes. Toxicol. In Vitro 2009, 23, 697–703. [Google Scholar] [CrossRef]
- Marfin, Y.S.; Vashurin, A.S.; Rumyantsev, E.V.; Puhovskaya, S.G. Sol–gel synthesis of highly effective catalyst based on cobalt tetrasulfophthalocyanine complex and silicon oxide. J. Solgel. Sci. Technol. 2013, 66, 306–311. [Google Scholar] [CrossRef]
- Mourant, J.R.; Yamada, Y.R.; Carpenter, S.; Dominique, L.R.; Freyer, J.P. FTIR spectroscopy demonstrates biochemical differences in mammalian cell cultures at different growth stages. Biophys. J. 2003, 85, 1938–1947. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Kim, C.S.; Ignacio, R.M.C.; Kim, D.H.; Sajo, M.E.J.; Maeng, E.H.; Qi, X.F.; Park, S.E.; Kim, Y.R.; Kim, M.K.; et al. Immunotoxicity of silicon dioxide nanoparticles with different sizes and electrostatic charge. Int. J. Nanomed. 2014, 9, 183–193. [Google Scholar] [CrossRef] [Green Version]
- Izak-Nau, E.; Kenesei, K.; Murali, K.; Voetz, M.; Eiden, S.; Puntes, V.F.; Duschl, A.; Madarász, E. Interaction of differently functionalized fluorescent silica nanoparticles with neural stem- and tissue-type cells. Nanotoxicology 2014, 8, 138–148. [Google Scholar] [CrossRef] [Green Version]
- Sun, D.; Gong, L.; Xie, J.; He, X.; Chen, S.; Luodan, A.; Li, Q.; Gu, Z.; Xu, H. Evaluating the toxicity of silicon dioxide nanoparticles on neural stem cells using RNA-Seq. RSC Adv. 2017, 7, 47552–47564. [Google Scholar] [CrossRef]
- Ye, Y.; Hui, L.; Lakpa, K.L.; Xing, Y.Q.; Wollenzien, H.; Chen, X.S. Effects of silica nanoparticles on endolysosome function in primary cultured neurons. Can. J. Physiol. Pharmacol. 2019, 97, 297–305. [Google Scholar] [CrossRef]
- Hashimoto, M.; Imazato, S. Cytotoxic and genotoxic characterization of aluminum and silicon oxide nanoparticles in macrophages. Dent. Mater. 2015, 31, 556–564. [Google Scholar] [CrossRef] [PubMed]
- McCracken, C.; Zane, A.; Knight, D.A.; Dutta, P.K.; Waldman, W.J. Minimal intestinal epithelial cell toxicity in response to short- and long-term food-relevant inorganic nanoparticle exposure. Chem. Res. Toxicol. 2013, 26, 1514–1525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nabeshi, H.; Yoshikawa, T.; Matsuyama, K.; Nakazato, Y.; Tochigi, S.; Kondoh, S.; Hirai, T.; Akase, T.; Nagano, K.; Abe, Y.; et al. Amorphous nanosilica induce endocytosis-dependent ROS generation and DNA damage in human keratinocytes. Part. Fibre Toxicol. 2011, 8, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nabeshi, H.; Yoshikawa, T.; Matsuyama, K.; Nakazato, Y.; Matsuo, K.; Arimori, A.; Isobe, M.; Tochigi, S.; Kondoh, S.; Hirai, T.; et al. Systemic distribution, nuclear entry and cytotoxicity of amorphous nanosilica following topical application. Biomaterials 2011, 32, 2713–2724. [Google Scholar] [CrossRef] [PubMed]
- Reemst, K.; Noctor, S.C.; Lucassen, P.J.; Hol, E.M. The Indispensable Roles of Microglia and Astrocytes during Brain Development. Front. Hum. Neurosci. 2016, 10, 566. [Google Scholar] [CrossRef] [Green Version]
- Schafer, D.P.; Stevens, B. Phagocytic glial cells: Sculpting synaptic circuits in the developing nervous system. Curr. Opin. Neurobiol. 2013, 23, 1034–1040. [Google Scholar] [CrossRef] [Green Version]
- Wakida, N.M.; Cruz, G.M.S.; Ro, C.C.; Moncada, E.G.; Khatibzadeh, N.; Flanagan, L.A. Phagocytic response of astrocytes to damaged neighboring cells. PLoS ONE 2018, 13, e0196153. [Google Scholar] [CrossRef]
- Fernández-Bertólez, N.; Costa, C.; Brandão, F.; Kiliç, G.; Duarte, J.A.; Teixeira, J.P. Toxicological assessment of silica-coated iron oxide nanoparticles in human astrocytes. Food Chem. Toxicol. 2018, 118, 13–23. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.; Zheng, Q.; Katz, H.E.; Guilarte, T.R. Silica-based nanoparticle uptake and cellular response by primary microglia. Environ. Health Perspect. 2010, 118, 589–595. [Google Scholar] [CrossRef] [Green Version]
- Megias, L.; Guerri, C.; Fornas, E.; Azorin, I.; Bendala, E.; Sancho-Tello, M. Endocytosis and transcytosis in growing astrocytes in primary culture. Possible implications in neural development. Int. J. Dev. Biol. 2000, 44, 209–221. [Google Scholar] [PubMed]
- Wilson, C.L.; Natarajan, V.; Hayward, S.L.; Khalimonchuk, O.; Kidambi, S. Mitochondrial dysfunction and loss of glutamate uptake in primary astrocytes exposed to titanium dioxide nanoparticles. Nanoscale 2015, 7, 18477–18488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, L.; Dan, M.; Shao, A.; Cheng, X.; Zhang, C.; Yokel, R.A.; Takemura, T.; Hanagata, N.; Niwa, M.; Watanabe, D. Silver nanoparticles induce tight junction disruption and astrocyte neurotoxicity in a rat blood-brain barrier primary triple coculture model. Int. J. Nanomed. 2015, 10, 105–118. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Du, X.J.; Wang, Q.; Liu, J.W.; Zhang, E.G.; Sai, L.L. Mechanism of cell death induced by silica nanoparticles in hepatocyte cells is by apoptosis. Int. J. Mol. Med. 2019, 44, 903–912. [Google Scholar] [CrossRef] [Green Version]
- Iavicoli, I.; Leso, V.; Fontana, L.; Calabrese, E.J. Nanoparticle Exposure and Hormetic Dose-Responses: An Update. Int. J. Mol. Sci. 2018, 19, 805. [Google Scholar] [CrossRef] [Green Version]
- Mytych, J.; Wnuk, M.; Rattan, S.I.S. Low doses of nanodiamonds and silica nanoparticles have beneficial hormetic effects in normal human skin fibroblasts in culture. Chemosphere 2016, 148, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Krętowski, R.; Kusaczuk, M.; Naumowicz, M.; Kotyńska, J.; Szynaka, B.; Cechowska-Pasko, M. The Effects of Silica Nanoparticles on Apoptosis and Autophagy of Glioblastoma Cell Lines. Nanomaterials (Basel) 2017, 7, 230. [Google Scholar] [CrossRef]
- Arnoldussen, Y.J.; Kringlen Ervik, T.; Baarnes Eriksen, M.; Kero, I.; Skaug, V.; Zienolddiny, S. Cellular Responses of Industrially Relevant Silica Dust on Human Glial Cells in Vitro. Int. J. Mol. Sci. 2019, 20, 358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, M.; Xie, L.L.; Fang, C.J.; Yang, H.; Wang, Y.J.; Zhen, X.Y. Implications for blood-brain-barrier permeability, in vitro oxidative stress and neurotoxicity potential induced by mesoporous silica nanoparticles: Effects of surface modification. RSC Adv. 2016, 6, 2800–2809. [Google Scholar] [CrossRef]
- Yang, X.F.; Liu, J.J.; He, H.W.; Zhou, L.; Gong, C.M.; Wang, X.M. SiO2 nanoparticles induce cytotoxicity and protein expression alteration in HaCaT cells. Part. Fibre Toxicol. 2010, 7, 1. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Fabuel, I.; Le Douce, J.; Logan, A.; James, A.M.; Bonvento, G.; Murphy, M.P. Complex I assembly into supercomplexes determines differential mitochondrial ROS production in neurons and astrocytes. Proc. Natl. Acad. Sci. USA 2016, 113, 13063–13068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.F.; He, C.E.; Li, J.; Chen, H.B.; Ma, Q.; Sui, X.J. Uptake of silica nanoparticles: Neurotoxicity and Alzheimer-like pathology in human SK-N-SH and mouse neuro2a neuroblastoma cells. Toxicol. Lett. 2014, 229, 240–249. [Google Scholar] [CrossRef] [PubMed]
- Gélinas, S.; Chapados, C.; Beauregard, M.; Gosselin, I.; Martinoli, M.G. Effect of oxidative stress on stability and structure of neurofilament proteins. Biochem. Cell Biol. 2000, 78, 667–674. [Google Scholar] [CrossRef] [PubMed]
- Xie, H.J.; Wu, J. Silica nanoparticles induce alpha-synuclein induction and aggregation in PC12-cells. Chem. Biol. Interact. 2016, 258, 197–204. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Type/Chemical Bond | Concentration of SiO2-NPs (µg/mL) | |||||||
|---|---|---|---|---|---|---|---|---|
| 0 | 1 | 10 | 25 | 50 | 100 | |||
| Astrocytes | ||||||||
| Vibration | Spectral Band | n= 8 | n= 8 | n= 8 | n= 8 | n= 8 | n= 8 | ANOVA |
| P | ||||||||
| SiO2-NPs | 1083 cm−1 | 0.023 ± 0.010 | 0.019 ± 0.008 | 0.014 ± 0.003 § | 0.017 ± 0.006 | 0.020 ± 0.006 | 0.028 ± 0.011 | 0.0177 * |
| Symmetric PO2− | 1090 cm−1 | 0.024 ± 0.011 | 0.019 ± 0.008 | 0.013 ± 0.003 § | 0.017 ± 0.006 | 0.020 ± 0.007 | 0.027 ± 0.012 | 0.0211 * |
| Parallel beta strand | 1627 cm−1 | 0.026 ± 0.010 | 0.021 ± 0.008 | 0.015 ± 0.003 § | 0.019 ± 0.005 | 0.022 ± 0.006 | 0.029 ± 0.011 | 0.0167 * |
| Random coil | 1639 cm−1 | 0.031 ± 0.012 | 0.026 ± 0.008 | 0.019 ± 0.003 § | 0.023 ± 0.005 | 0.025 ± 0.007 | 0.033 ± 0.011 | 0.0241 * |
| Alpha Helix | 1654 cm−1 | 0.034 ± 0.012 | 0.030 ± 0.009 | 0.022 ± 0.003 § | 0.026 ± 0.005 | 0.029 ± 0.008 | 0.036 ± 0.010 | 0.0246 * |
| Symmetric -CH2 | 2852 cm−1 | 0.029 ± 0.010 | 0.026 ± 0.009 | 0.019 ± 0.003 § | 0.025 ± 0.004 | 0.026 ± 0.008 | 0.033 ± 0.011 | 0.0452 * |
| Asymmetric -CH2 | 2923 cm−1 | 0.029 ± 0.010 | 0.025 ± 0.008 | 0.019 ± 0.003 § | 0.024 ± 0.004 | 0.026 ± 0.008 | 0.032 ± 0.011 | 0.0292 * |
| Neurons | ||||||||
| Vibration | Spectral Band | n= 11 | n= 11 | n= 11 | n= 11 | n= 11 | n= 11 | ANOVA |
| P | ||||||||
| SiO2-NPs | 1083 cm−1 | 0.012 ± 0.003 | 0.014 ± 0.006 | 0.014 ± 0.004 | 0.012 ± 0.003 | 0.013 ± 0.004 | 0.015 ± 0.007 | 0.5955 |
| Symmetric PO2− | 1090 cm−1 | 0.012 ± 0.003 | 0.014 ± 0.007 | 0.014 ± 0.004 | 0.013 ± 0.003 | 0.014 ± 0.004 | 0.016 ± 0.007 | 0.5404 |
| Parallel beta strand | 1627 cm−1 | 0.015 ± 0.003 | 0.018 ± 0.006 | 0.018 ± 0.004 | 0.016 ± 0.003 | 0.017 ± 0.003 | 0.019 ± 0.007 | 0.4314 |
| Random coil | 1639 cm−1 | 0.016 ± 0.003 | 0.019 ± 0.008 | 0.019 ± 0.005 | 0.017 ± 0.003 | 0.018 ± 0.00 | 0.020 ± 0.007 | 0.5086 |
| Alpha Helix | 1654 cm−1 | 0.021 ± 0.004 | 0.024 ± 0.008 | 0.024 ± 0.005 | 0.028 ± 0.003 | 0.024 ± 0.004 | 0.027 ± 0.008 | 0.3475 |
| Symmetric -CH2 | 2852 cm−1 | 0.028 ± 0.003 | 0.030 ± 0.007 | 0.031 ± 0.004 | 0.030 ± 0.003 | 0.031 ± 0.004 | 0.033 ± 0.008 | 0.4128 |
| Asymmetric -CH2 | 2923 cm−1 | 0.026 ± 0.003 | 0.028 ± 0.007 | 0.029 ± 0.004 | 0.028 ± 0.003 | 0.029 ± 0.004 | 0.030 ± 0.008 | 0.4027 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Limón-Pacheco, J.H.; Jiménez-Barrios, N.; Déciga-Alcaraz, A.; Martínez-Cuazitl, A.; Mata-Miranda, M.M.; Vázquez-Zapién, G.J.; Pedraza-Chaverri, J.; Chirino, Y.I.; Orozco-Ibarra, M. Astrocytes Are More Vulnerable than Neurons to Silicon Dioxide Nanoparticle Toxicity in Vitro. Toxics 2020, 8, 51. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics8030051
Limón-Pacheco JH, Jiménez-Barrios N, Déciga-Alcaraz A, Martínez-Cuazitl A, Mata-Miranda MM, Vázquez-Zapién GJ, Pedraza-Chaverri J, Chirino YI, Orozco-Ibarra M. Astrocytes Are More Vulnerable than Neurons to Silicon Dioxide Nanoparticle Toxicity in Vitro. Toxics. 2020; 8(3):51. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics8030051
Chicago/Turabian StyleLimón-Pacheco, Jorge Humberto, Natalie Jiménez-Barrios, Alejandro Déciga-Alcaraz, Adriana Martínez-Cuazitl, Mónica Maribel Mata-Miranda, Gustavo Jesús Vázquez-Zapién, Jose Pedraza-Chaverri, Yolanda Irasema Chirino, and Marisol Orozco-Ibarra. 2020. "Astrocytes Are More Vulnerable than Neurons to Silicon Dioxide Nanoparticle Toxicity in Vitro" Toxics 8, no. 3: 51. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics8030051