Teratogenic, Oxidative Stress and Behavioural Outcomes of Three Fungicides of Natural Origin (Equisetum arvense, Mimosa tenuiflora, Thymol) on Zebrafish (Danio rerio)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Animals
2.3. Concentration Determination
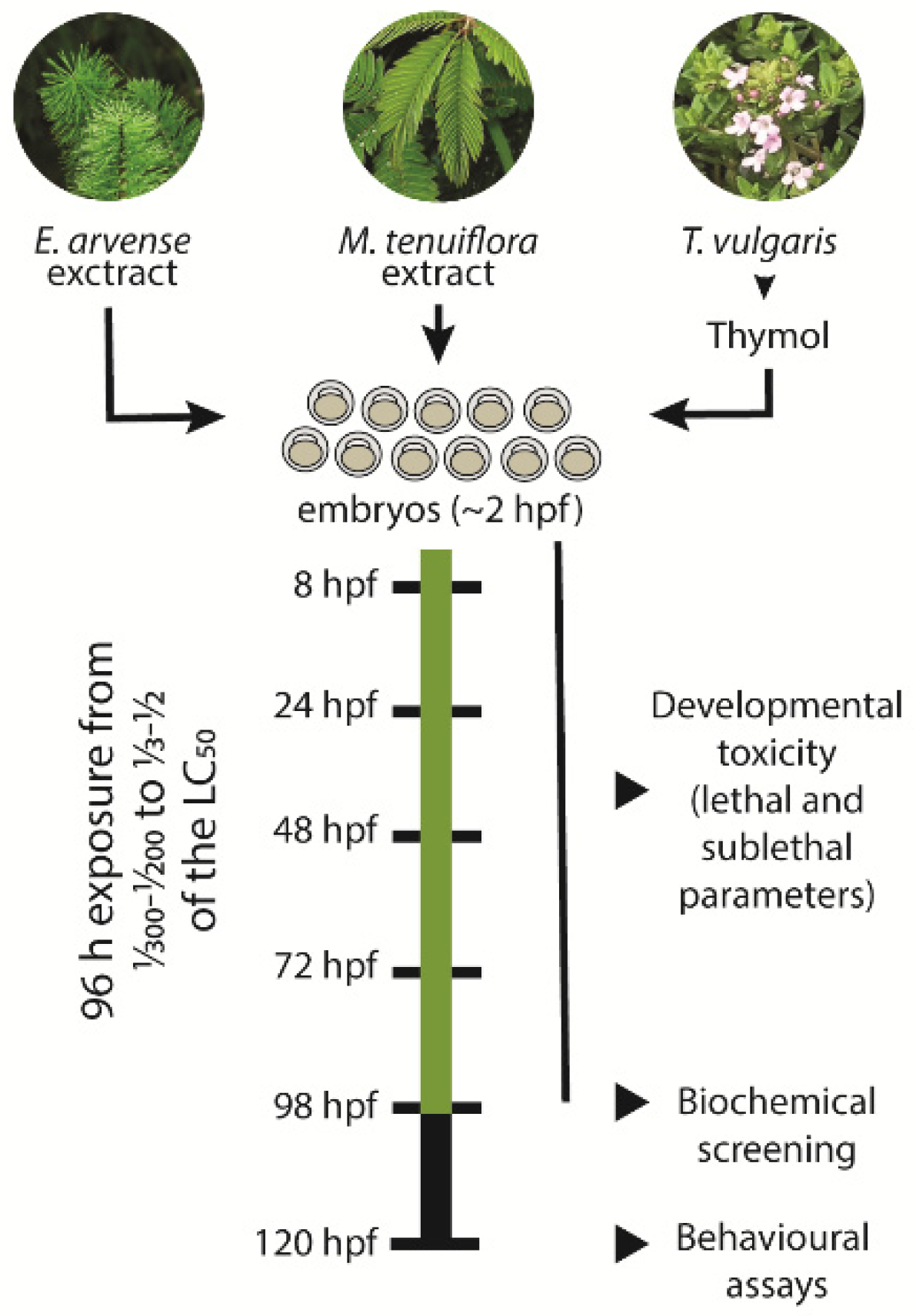
2.4. Embryo Toxicity
2.5. Biochemical Analysis
2.6. Locomotor Behaviour Analysis
2.7. Statistical Analysis
3. Results
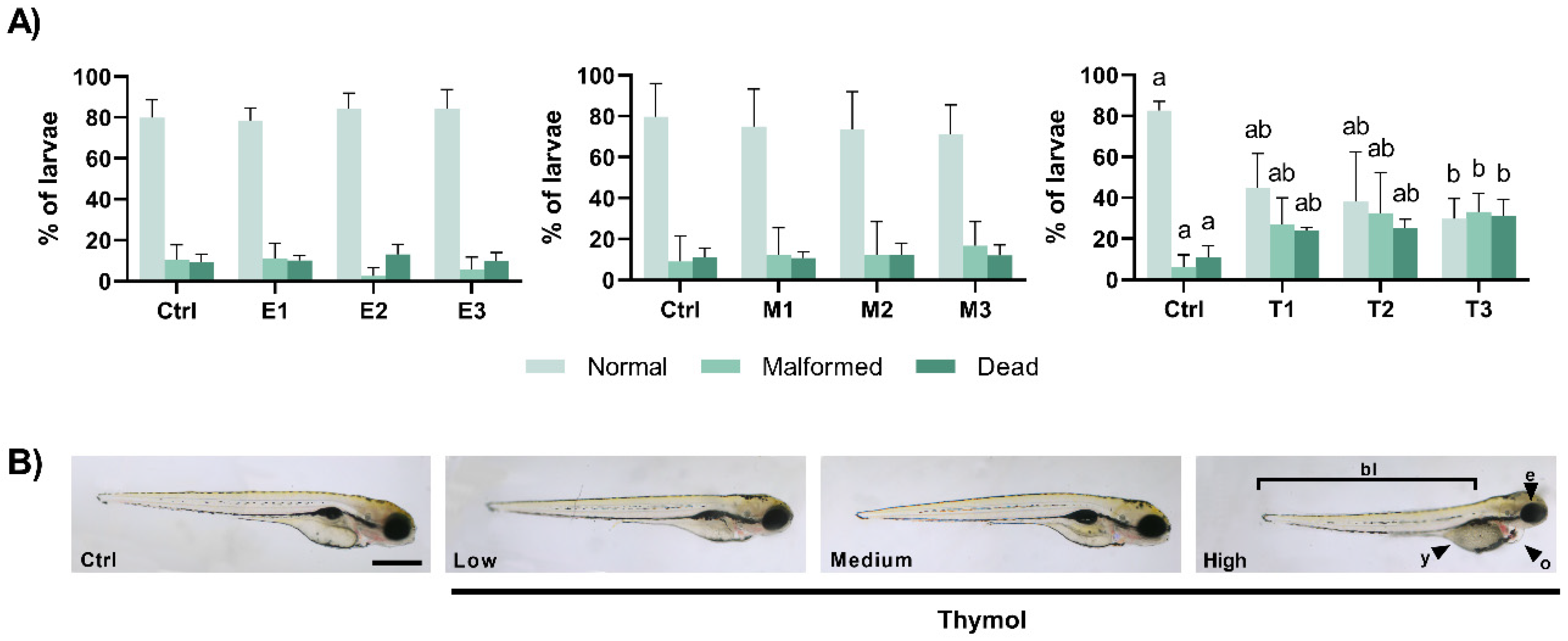
3.1. Teratogenic Effects of Phyto-Fungicide Formulations
3.2. Biochemical Markers Affected by the Phyto-Fungicide Exposure
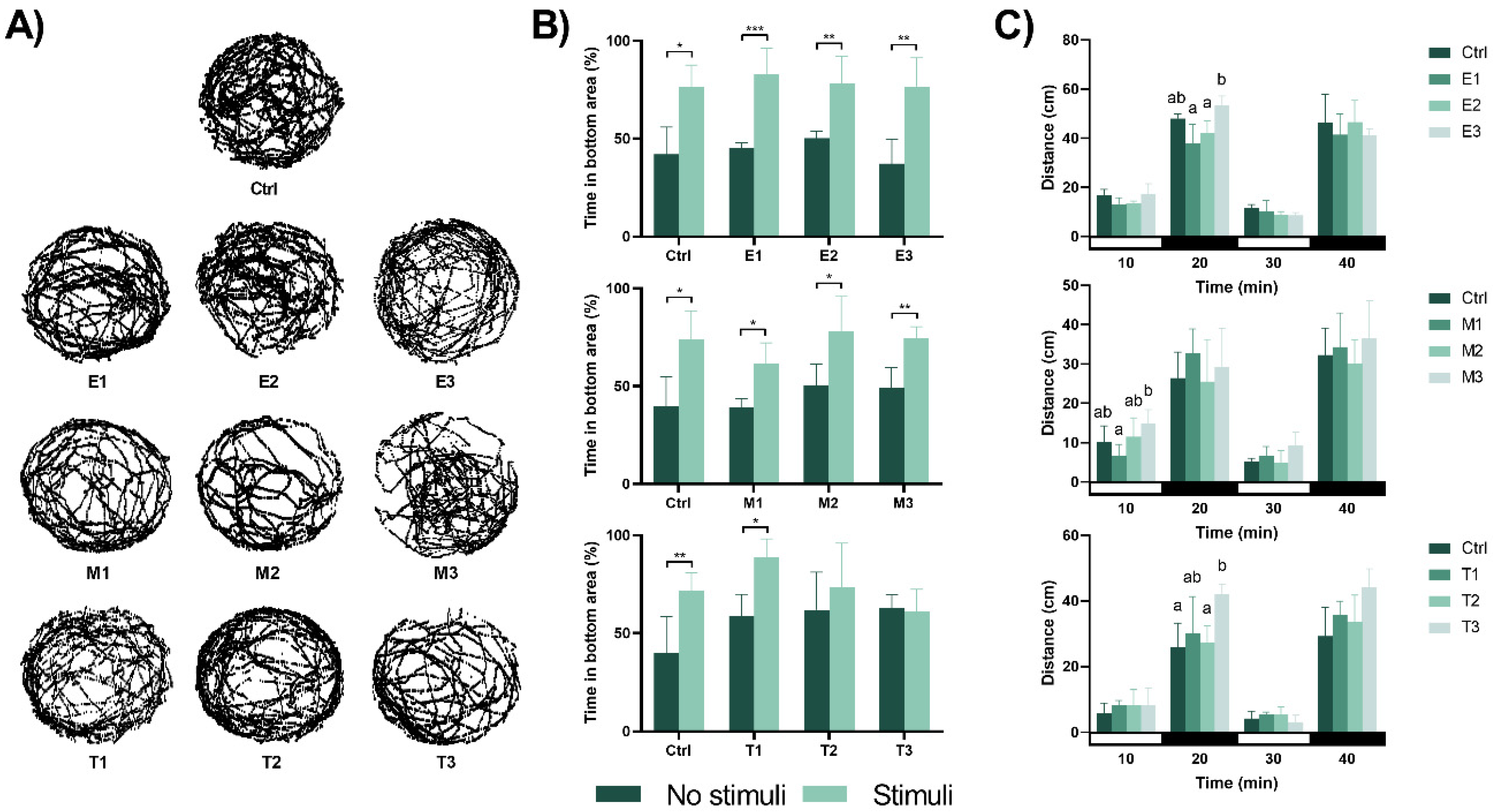
3.3. Behavioural Responses Induced by the Different Formulations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jeschke, P. Progress of modern agricultural chemistry and future prospects. Pest Manag. Sci. 2016, 72, 433–455. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, F.P. Pesticides, environment, and food safety. Food Energy Secur. 2017, 6, 48–60. [Google Scholar] [CrossRef]
- Gonzalez-Rey, M.; Tapie, N.; Le Menach, K.; Devier, M.H.; Budzinski, H.; Bebianno, M.J. Occurrence of pharmaceutical compounds and pesticides in aquatic systems. Mar. Pollut. Bull. 2015, 96, 384–400. [Google Scholar] [CrossRef] [PubMed]
- Zubrod, J.P.; Bundschuh, M.; Arts, G.; Bruhl, C.A.; Imfeld, G.; Knabel, A.; Payraudeau, S.; Rasmussen, J.J.; Rohr, J.; Scharmuller, A.; et al. Fungicides: An overlooked pesticide class? Environ. Sci. Technol. 2019, 53, 3347–3365. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Jennings, A. Worldwide regulations of standard values of pesticides for human health risk control: A review. Int. J. Environ. Res. Public Health 2017, 14, 826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbey, J.A.; Percival, D.; Abbey, L.; Asiedu, S.K.; Prithiviraj, B.; Schilder, A. Biofungicides as alternative to synthetic fungicide control of grey mould (Botrytis cinerea)—Prospects and challenges. Biocontrol Sci. Technol. 2019, 29, 241–262. [Google Scholar] [CrossRef]
- Lorsbach, B.A.; Sparks, T.C.; Cicchillo, R.M.; Garizi, N.V.; Hahn, D.R.; Meyer, K.G. Natural products: A strategic lead generation approach in crop protection discovery. Pest Manag. Sci. 2019, 75, 2301–2309. [Google Scholar] [CrossRef] [PubMed]
- Palou, L.; Ali, A.; Fallik, E.; Romanazzi, G. GRAS, plant- and animal-derived compounds as alternatives to conventional fungicides for the control of postharvest diseases of fresh horticultural produce. Postharvest Biol. Technol. 2016, 122, 41–52. [Google Scholar] [CrossRef]
- Marrone, P.G. Pesticidal natural products—Status and future potential. Pest Manag. Sci. 2019, 75, 2325–2340. [Google Scholar] [CrossRef]
- Pavela, R.; Benelli, G. Essential oils as ecofriendly biopesticides? Challenges and constraints. Trends Plant Sci. 2016, 21, 1000–1007. [Google Scholar] [CrossRef]
- Damalas, C.A.; Koutroubas, S.D. Botanical pesticides for eco-friendly pest management: Drawbacks and limitations. In Pesticides in Crop Production: Physiological and Biochemical Action; Wiley: Chichester, UK, 2020; pp. 181–193. [Google Scholar]
- Vagner, M.; Zambonino-Infante, J.L.; Mazurais, D. Fish facing global change: Are early stages the lifeline? Mar. Environ. Res. 2019, 147, 159–178. [Google Scholar] [CrossRef] [PubMed]
- Marchand, P.A. Basic substances under EU pesticide regulation: An opportunity for organic production? Org. Farming 2017, 3, 16–19. [Google Scholar] [CrossRef] [Green Version]
- Ortega Garcia, J.A.; Angulo, M.G.; Sobrino-Najul, E.J.; Soldin, O.P.; Mira, A.P.; Martinez-Salcedo, E.; Claudio, L. Prenatal exposure of a girl with autism spectrum disorder to ‘horsetail’ (Equisetum arvense) herbal remedy and alcohol: A case report. J. Med. Case Rep. 2011, 5, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medeiros, R.M.; de Figueiredo, A.P.; Benicio, T.M.; Dantas, F.P.; Riet-Correa, F. Teratogenicity of Mimosa tenuiflora seeds to pregnant rats. Toxicon 2008, 51, 316–319. [Google Scholar] [CrossRef]
- Pimentel, L.A.; Correa, F.R.; Gardner, D.; Panter, K.E.; Dantas, A.F.; Medeiros, R.M.; Mota, R.A.; Araujo, J.A. Mimosa tenuiflora as a cause of malformations in ruminants in the northeastern Brazilian semiarid rangelands. Vet. Pathol. 2007, 44, 928–931. [Google Scholar] [CrossRef]
- Marchese, A.; Orhan, I.E.; Daglia, M.; Barbieri, R.; Di Lorenzo, A.; Nabavi, S.F.; Gortzi, O.; Izadi, M.; Nabavi, S.M. Antibacterial and antifungal activities of thymol: A brief review of the literature. Food Chem. 2016, 210, 402–414. [Google Scholar] [CrossRef]
- Verrett, M.J.; Scott, W.F.; Reynaldo, E.F.; Alterman, E.K.; Thomas, C.A. Toxicity and teratogenicity of food additive chemicals in the developing chicken embryo. Toxicol. Appl. Pharmacol. 1980, 56, 265–273. [Google Scholar] [CrossRef]
- Krishnan, M.; Kim, D.K.; Gie Kim, S.; Kang, S.C. Thymol exposure mediates pro-oxidant shift by regulating Nrf2 and apoptotic events in zebrafish (Danio rerio) embryos. Environ. Toxicol. Pharmacol. 2019, 65, 1–8. [Google Scholar] [CrossRef]
- He, T.; Li, X.; Wang, X.; Xu, X.; Yan, X.; Li, X.; Sun, S.; Dong, Y.; Ren, X.; Liu, X.; et al. Chemical composition and anti-oxidant potential on essential oils of Thymus quinquecostatus Celak. from Loess Plateau in China, regulating Nrf2/Keap1 signaling pathway in zebrafish. Sci. Rep. 2020, 10, 11280. [Google Scholar] [CrossRef]
- Capatina, L.; Todirascu-Ciornea, E.; Napoli, E.M.; Ruberto, G.; Hritcu, L.; Dumitru, G. Thymus vulgaris essential oil protects zebrafish against cognitive dysfunction by regulating cholinergic and antioxidants systems. Antioxidants 2020, 9, 1083. [Google Scholar] [CrossRef]
- Garcia, D.; Ramos, A.J.; Sanchis, V.; Marín, S. Equisetum arvense hydro-alcoholic extract: Phenolic composition and antifungal and antimycotoxigenic effect against Aspergillus flavus and Fusarium verticillioides in stored maize. J. Sci. Food Agric. 2013, 93, 2248–2253. [Google Scholar] [CrossRef] [PubMed]
- Llorens, E.; Mateu, M.; González-Hernández, A.I.; Agustí-Brisach, C.; García-Agustín, P.; Lapeña, L.; Vicedo, B. Extract of Mimosa tenuiflora and Quercus robur as potential eco-friendly management tool against Sclerotinia sclerotiorum in Lactuca sativa enhancing the natural plant defences. Eur. J. Plant Pathol. 2019, 153, 1105–1118. [Google Scholar] [CrossRef]
- Hinton, D.E.; Kullman, S.W.; Hardman, R.C.; Volz, D.C.; Chen, P.J.; Carney, M.; Bencic, D.C. Resolving mechanisms of toxicity while pursuing ecotoxicological relevance? Mar. Pollut. Bull. 2005, 51, 635–648. [Google Scholar] [CrossRef] [PubMed]
- Bambino, K.; Chu, J. Zebrafish in toxicology and environmental health. Curr. Top. Dev. Biol. 2017, 124, 331–367. [Google Scholar] [CrossRef] [Green Version]
- Magyary, I. Recent advances and future trends in zebrafish bioassays for aquatic ecotoxicology. Ecocycles 2018, 4, 12–18. [Google Scholar] [CrossRef]
- Scholz, S.; Fischer, S.; Gundel, U.; Kuster, E.; Luckenbach, T.; Voelker, D. The zebrafish embryo model in environmental risk assessment—Applications beyond acute toxicity testing. Environ. Sci. Pollut. Res. Int. 2008, 15, 394–404. [Google Scholar] [CrossRef]
- Bugel, S.M.; Tanguay, R.L.; Planchart, A. Zebrafish: A marvel of high-throughput biology for 21(st) century toxicology. Curr. Environ. Health Rep. 2014, 1, 341–352. [Google Scholar] [CrossRef] [Green Version]
- Domingues, I.; Oliveira, R.; Lourenco, J.; Grisolia, C.K.; Mendo, S.; Soares, A.M. Biomarkers as a tool to assess effects of chromium (VI): Comparison of responses in zebrafish early life stages and adults. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2010, 152, 338–345. [Google Scholar] [CrossRef]
- Sisman, T. Early life stage and genetic toxicity of stannous chloride on zebrafish embryos and adults: Toxic effects of tin on zebrafish. Environ. Toxicol. 2011, 26, 240–249. [Google Scholar] [CrossRef]
- Knobel, M.; Busser, F.J.; Rico-Rico, A.; Kramer, N.I.; Hermens, J.L.; Hafner, C.; Tanneberger, K.; Schirmer, K.; Scholz, S. Predicting adult fish acute lethality with the zebrafish embryo: Relevance of test duration, endpoints, compound properties, and exposure concentration analysis. Environ. Sci. Technol. 2012, 46, 9690–9700. [Google Scholar] [CrossRef]
- OECD Guidelines. Test No. 236: Fish Embryo Acute Toxicity (FET) Test. In OECD Guidelines for the Testing of Chemicals, Section 2; OECD: Paris, France, 2013; pp. 1–22. [Google Scholar] [CrossRef]
- Felix, L.M.; Serafim, C.; Valentim, A.M.; Antunes, L.M.; Campos, S.; Matos, M.; Coimbra, A.M. Embryonic stage-dependent teratogenicity of ketamine in zebrafish (Danio rerio). Chem. Res. Toxicol. 2016, 29, 1298–1309. [Google Scholar] [CrossRef] [PubMed]
- Felix, L.M.; Serafim, C.; Martins, M.J.; Valentim, A.M.; Antunes, L.M.; Matos, M.; Coimbra, A.M. Morphological and behavioral responses of zebrafish after 24 h of ketamine embryonic exposure. Toxicol. Appl. Pharmacol. 2017, 321, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Vieira, R.; Venancio, C.A.S.; Felix, L.M. Toxic effects of a mancozeb-containing commercial formulation at environmental relevant concentrations on zebrafish embryonic development. Environ. Sci. Pollut. Res. Int. 2020, 27, 21174–21187. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, W.; Burgeot, T.; Porcher, J.M. A novel “Integrated Biomarker Response” calculation based on reference deviation concept. Environ. Sci. Pollut. Res. Int. 2013, 20, 2721–2725. [Google Scholar] [CrossRef] [PubMed]
- Felix, L.M.; Antunes, L.M.; Coimbra, A.M.; Valentim, A.M. Behavioral alterations of zebrafish larvae after early embryonic exposure to ketamine. Psychopharmacology 2017, 234, 549–558. [Google Scholar] [CrossRef]
- Andrade, T.S.; Henriques, J.F.; Almeida, A.R.; Machado, A.L.; Koba, O.; Giang, P.T.; Soares, A.; Domingues, I. Carbendazim exposure induces developmental, biochemical and behavioural disturbance in zebrafish embryos. Aquat. Toxicol. 2016, 170, 390–399. [Google Scholar] [CrossRef] [Green Version]
- Chahardehi, A.M.; Arsad, H.; Lim, V. Zebrafish as a successful animal model for screening toxicity of medicinal plants. Plants 2020, 9, 1345. [Google Scholar] [CrossRef]
- Andreu, V.; Levert, A.; Amiot, A.; Cousin, A.; Aveline, N.; Bertrand, C. Chemical composition and antifungal activity of plant extracts traditionally used in organic and biodynamic farming. Environ. Sci. Pollut. Res. Int. 2018, 25, 29971–29982. [Google Scholar] [CrossRef]
- Schier, A.F. Axis formation and patterning in zebrafish. Curr. Opin. Genet. Dev. 2001, 11, 393–404. [Google Scholar] [CrossRef]
- Salazar, V.S.; Gamer, L.W.; Rosen, V. BMP signalling in skeletal development, disease and repair. Nat. Rev. Endocrinol. 2016, 12, 203–221. [Google Scholar] [CrossRef]
- Reynolds, K.; Kumari, P.; Sepulveda Rincon, L.; Gu, R.; Ji, Y.; Kumar, S.; Zhou, C.J. Wnt signaling in orofacial clefts: Crosstalk, pathogenesis and models. Dis. Model. Mech. 2019, 12, dmm037051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, R.; Chen, Z.Z. Thymol inhibits cell migration and invasion by downregulating the activation of PI3K/AKT and ERK pathways in human colon cancer cells. Trop. J. Pharm. Res. 2017, 16, 2895–2901. [Google Scholar] [CrossRef] [Green Version]
- Akieda, Y.; Ogamino, S.; Furuie, H.; Ishitani, S.; Akiyoshi, R.; Nogami, J.; Masuda, T.; Shimizu, N.; Ohkawa, Y.; Ishitani, T. Cell competition corrects noisy Wnt morphogen gradients to achieve robust patterning in the zebrafish embryo. Nat. Commun. 2019, 10, 4710. [Google Scholar] [CrossRef] [PubMed]
- Hikasa, H.; Sokol, S.Y. Wnt signaling in vertebrate axis specification. Cold Spring Harb. Perspect. Biol. 2013, 5, a007955. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.S.; Cui, W. Proliferation, survival and metabolism: The role of PI3K/AKT/mTOR signalling in pluripotency and cell fate determination. Development 2016, 143, 3050–3060. [Google Scholar] [CrossRef] [Green Version]
- Finkielsztein, A.; Kelly, G.M. Altering PI3K-Akt signalling in zebrafish embryos affects PTEN phosphorylation and gastrulation. Biol. Cell 2009, 101, 661–678. [Google Scholar] [CrossRef]
- Koundouros, N.; Poulogiannis, G. Phosphoinositide 3-Kinase/Akt signaling and redox metabolism in cancer. Front. Oncol. 2018, 8, 160. [Google Scholar] [CrossRef]
- Hahn, M.E.; Timme-Laragy, A.R.; Karchner, S.I.; Stegeman, J.J. Nrf2 and Nrf2-related proteins in development and developmental toxicity: Insights from studies in zebrafish (Danio rerio). Free Radic. Biol. Med. 2015, 88, 275–289. [Google Scholar] [CrossRef] [Green Version]
- Leung, L.; Kwong, M.; Hou, S.; Lee, C.; Chan, J.Y. Deficiency of the Nrf1 and Nrf2 transcription factors results in early embryonic lethality and severe oxidative stress. J. Biol. Chem. 2003, 278, 48021–48029. [Google Scholar] [CrossRef] [Green Version]
- Dennery, P.A. Oxidative stress in development: Nature or nurture? Free Radic. Biol. Med. 2010, 49, 1147–1151. [Google Scholar] [CrossRef]
- Massarsky, A.; Kozal, J.S.; Di Giulio, R.T. Glutathione and zebrafish: Old assays to address a current issue. Chemosphere 2017, 168, 707–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timme-Laragy, A.R.; Goldstone, J.V.; Imhoff, B.R.; Stegeman, J.J.; Hahn, M.E.; Hansen, J.M. Glutathione redox dynamics and expression of glutathione-related genes in the developing embryo. Free Radic. Biol. Med. 2013, 65, 89–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tierbach, A.; Groh, K.J.; Schonenberger, R.; Schirmer, K.; Suter, M.J. Glutathione S-Transferase protein expression in different life stages of zebrafish (Danio rerio). Toxicol. Sci. 2018, 162, 702–712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schafer, F.Q.; Buettner, G.R. Redox environment of the cell as viewed through the redox state of the glutathione disulfide/glutathione couple. Free Radic. Biol. Med. 2001, 30, 1191–1212. [Google Scholar] [CrossRef]
- Liu, Y.; Sepich, D.S.; Solnica-Krezel, L. Stat3/Cdc25a-dependent cell proliferation promotes embryonic axis extension during zebrafish gastrulation. PLoS Genet. 2017, 13, e1006564. [Google Scholar] [CrossRef]
- Murakami, S.; Motohashi, H. Roles of Nrf2 in cell proliferation and differentiation. Free Radic. Biol. Med. 2015, 88, 168–178. [Google Scholar] [CrossRef] [Green Version]
- Ozkan, A.; Erdogan, A. A comparative study of the antioxidant/prooxidant effects of carvacrol and thymol at various concentrations on membrane and DNA of parental and drug resistant H1299 cells. Nat. Prod. Commun. 2012, 7, 1557–1560. [Google Scholar] [CrossRef] [Green Version]
- Guvenc, M.; Cellat, M.; Gokcek, I.; Yavas, I.; Yurdagul Ozsoy, S. Effects of thymol and carvacrol on sperm quality and oxidant/antioxidant balance in rats. Arch. Physiol. Biochem. 2019, 125, 396–403. [Google Scholar] [CrossRef]
- Kandil, S.; Brennan, L.; McBean, G.J. Glutathione depletion causes a JNK and p38MAPK-mediated increase in expression of cystathionine-gamma-lyase and upregulation of the transsulfuration pathway in C6 glioma cells. Neurochem. Int. 2010, 56, 611–619. [Google Scholar] [CrossRef]
- Day, R.M.; Suzuki, Y.J. Cell proliferation, reactive oxygen and cellular glutathione. Dose Response 2006, 3, 425–442. [Google Scholar] [CrossRef]
- Obrador, E.; Navarro, J.; Mompo, J.; Asensi, M.; Pellicer, J.A.; Estrela, J.M. Glutathione and the rate of cellular proliferation determine tumour cell sensitivity to tumour necrosis factor in vivo. Biochem. J. 1997, 325 Pt 1, 183–189. [Google Scholar] [CrossRef] [Green Version]
- Glisic, B.; Mihaljevic, I.; Popovic, M.; Zaja, R.; Loncar, J.; Fent, K.; Kovacevic, R.; Smital, T. Characterization of glutathione-S-transferases in zebrafish (Danio rerio). Aquat. Toxicol. 2015, 158, 50–62. [Google Scholar] [CrossRef] [PubMed]
- Glavan, G.; Novak, S.; Bozic, J.; Jemec Kokalj, A. Comparison of sublethal effects of natural acaricides carvacrol and thymol on honeybees. Pestic. Biochem. Physiol. 2020, 166, 104567. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, K.; Wada, K.; Tanaka, Y.; Yoshimura, T.; Matuoka, K.; Anno, T. Thyme (Thymus vulgaris L.) leaves and its constituents increase the activities of xenobiotic-metabolizing enzymes in mouse liver. J. Med. Food 2005, 8, 184–189. [Google Scholar] [CrossRef] [PubMed]
- Kumrungsee, N.; Pluempanupat, W.; Koul, O.; Bullangpoti, V. Toxicity of essential oil compounds against diamondback moth, Plutella xylostella, and their impact on detoxification enzyme activities. J. Pest Sci. 2014, 87, 721–729. [Google Scholar] [CrossRef]
- Shahriari, M.; Zibaee, A.; Sahebzadeh, N.; Shamakhi, L. Effects of alpha-pinene, trans-anethole, and thymol as the essential oil constituents on antioxidant system and acetylcholine esterase of Ephestia kuehniella Zeller (Lepidoptera: Pyralidae). Pestic. Biochem. Physiol. 2018, 150, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Laborde, E. Glutathione transferases as mediators of signaling pathways involved in cell proliferation and cell death. Cell Death Differ. 2010, 17, 1373–1380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, Y.; Zhou, Y.; Xiong, Z.; Zou, L.; Jiang, M.; Luo, Z.; Wen, S.; Liu, W.; Liu, S.; Li, W. Involvement of JNK in the embryonic development and organogenesis in zebrafish. Mar. Biotechnol. (NY) 2013, 15, 716–725. [Google Scholar] [CrossRef]
- Valesio, E.G.; Zhang, H.; Zhang, C. Exposure to the JNK inhibitor SP600125 (anthrapyrazolone) during early zebrafish development results in morphological defects. J. Appl. Toxicol. 2013, 33, 32–40. [Google Scholar] [CrossRef]
- Liang, D.; Li, F.; Fu, Y.; Cao, Y.; Song, X.; Wang, T.; Wang, W.; Guo, M.; Zhou, E.; Li, D.; et al. Thymol inhibits LPS-stimulated inflammatory response via down-regulation of NF-kappaB and MAPK signaling pathways in mouse mammary epithelial cells. Inflammation 2014, 37, 214–222. [Google Scholar] [CrossRef]
- Gholijani, N.; Gharagozloo, M.; Farjadian, S.; Amirghofran, Z. Modulatory effects of thymol and carvacrol on inflammatory transcription factors in lipopolysaccharide-treated macrophages. J. Immunotoxicol. 2016, 13, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Colwill, R.M.; Creton, R. Imaging escape and avoidance behavior in zebrafish larvae. Rev. Neurosci. 2011, 22, 63–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jesuthasan, S. Fear, anxiety, and control in the zebrafish. Dev. Neurobiol. 2012, 72, 395–403. [Google Scholar] [CrossRef] [PubMed]
- Capibaribe, V.C.C.; Vasconcelos Mallmann, A.S.; Lopes, I.S.; Oliveira, I.C.M.; de Oliveira, N.F.; Chaves, R.C.; Fernandes, M.L.; de Araujo, M.A.; da Silva, D.M.A.; Valentim, J.T.; et al. Thymol reverses depression-like behaviour and upregulates hippocampal BDNF levels in chronic corticosterone-induced depression model in female mice. J. Pharm. Pharmacol. 2019, 71, 1774–1783. [Google Scholar] [CrossRef]
- Deng, X.Y.; Li, H.Y.; Chen, J.J.; Li, R.P.; Qu, R.; Fu, Q.; Ma, S.P. Thymol produces an antidepressant-like effect in a chronic unpredictable mild stress model of depression in mice. Behav. Brain Res. 2015, 291, 12–19. [Google Scholar] [CrossRef]
- Okamoto, H.; Agetsuma, M.; Aizawa, H. Genetic dissection of the zebrafish habenula, a possible switching board for selection of behavioral strategy to cope with fear and anxiety. Dev. Neurobiol. 2012, 72, 386–394. [Google Scholar] [CrossRef] [Green Version]
- Agetsuma, M.; Aizawa, H.; Aoki, T.; Nakayama, R.; Takahoko, M.; Goto, M.; Sassa, T.; Amo, R.; Shiraki, T.; Kawakami, K.; et al. The habenula is crucial for experience-dependent modification of fear responses in zebrafish. Nat. Neurosci. 2010, 13, 1354–1356. [Google Scholar] [CrossRef]
- Lee, A.; Mathuru, A.S.; Teh, C.; Kibat, C.; Korzh, V.; Penney, T.B.; Jesuthasan, S. The habenula prevents helpless behavior in larval zebrafish. Curr. Biol. 2010, 20, 2211–2216. [Google Scholar] [CrossRef] [Green Version]
- Fore, S.; Acuña-Hinrichsen, F.; Mutlu, K.A.; Bartoszek, E.M.; Serneels, B.; Faturos, N.G.; Chau, K.T.P.; Cosacak, M.I.; Verdugo, C.D.; Palumbo, F.; et al. Functional properties of habenular neurons are determined by developmental stage and sequential neurogenesis. Sci. Adv. 2020, 6, eaaz3173. [Google Scholar] [CrossRef]
- Schmidt, R.; Strahle, U.; Scholpp, S. Neurogenesis in zebrafish—From embryo to adult. Neural Dev. 2013, 8, 3. [Google Scholar] [CrossRef] [Green Version]
- Amura, C.R.; Marek, L.; Winn, R.A.; Heasley, L.E. Inhibited neurogenesis in JNK1-deficient embryonic stem cells. Mol. Cell. Biol. 2005, 25, 10791–10802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro-Torres, R.D.; Landa, J.; Rabaza, M.; Busquets, O.; Olloquequi, J.; Ettcheto, M.; Beas-Zarate, C.; Folch, J.; Camins, A.; Auladell, C.; et al. JNK Isoforms Are Involved in the Control of Adult Hippocampal Neurogenesis in Mice, Both in Physiological Conditions and in an Experimental Model of Temporal Lobe Epilepsy. Mol. Neurobiol. 2019, 56, 5856–5865. [Google Scholar] [CrossRef] [PubMed]
- Coffey, E.T. Nuclear and cytosolic JNK signalling in neurons. Nat. Rev. Neurosci. 2014, 15, 285–299. [Google Scholar] [CrossRef] [PubMed]
- Hollos, P.; Marchisella, F.; Coffey, E.T. JNK Regulation of depression and anxiety. Brain Plast. 2018, 3, 145–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phyto-fungicide | Group | 24 hpf | 48 hpf | 72 hpf | 98 hpf | |||
|---|---|---|---|---|---|---|---|---|
| Spontaneous Movements (units/min) | Heart Rate (beats/min) | Hatching Rate (%) | Body Length (mm) | Yolk Area (mm2) | Heart Area (mm2) | Eye Area (mm2) | ||
| E. arvense | Ctrl | 2.0 (2.0–2.5) | 123.6 ± 1.82 | 70.0 (65.5–71.5) | 3.75 (3.71–3.77) | 0.21 (0.20–0.21) | 0.034 ± 0.001 | 0.082 (0.082–0.085) |
| E1 | 2.0 (2.0–3.0) | 124.2 ± 3.42 | 70.0 (66.0–71.0) | 3.74 (3.72–3.78) | 0.21 (0.21–0.22) | 0.033 ± 0.003 | 0.083 (0.081–0.084) | |
| E2 | 1.0 (1.0–2.0) | 117.0 ± 3.39 | 68.0 (63.5–71.5) | 3.53 (3.49–3.65) | 0.22 (0.22–0.22) | 0.035 ± 0.003 | 0.075 (0.074–0.079) | |
| E3 | 2.0 (2.0–2.5) | 121.2 ± 6.57 | 68.0 (61.5–68.5) | 3.72 (3.65–3.80) | 0.20 (0.20–0.21) | 0.032 ± 0.004 | 0.081 (0.079–0.085) | |
| M. tenuiflora | Ctrl | 2.0 (0.5–2.0) | 113.2 ± 7.19 | 62.5 ± 7.7 | 3.56 ± 0.20 | 0.22 (0.21–0.23) | 0.038 ± 0.005 | 0.081 ± 0.015 |
| M1 | 1.0 (1.0–1.5) | 115.0 ± 9.82 | 65.6 ± 5.3 | 3.48 ± 0.26 | 0.24 (0.23–0.24) | 0.035 ± 0.006 | 0.086 ± 0.015 | |
| M2 | 2.0 (1.0–2.0) | 120.0 ± 5.92 | 56.4 ± 13 | 3.65 ± 0.14 | 0.24 (0.22–0.25) | 0.039 ± 0.005 | 0.088 ± 0.010 | |
| M3 | 1.0 (0.5–2.5) | 118.8 ± 11.4 | 54.0 ± 16 | 3.60 ± 0.15 | 0.25 (0.22–0.25) | 0.038 ± 0.005 | 0.087 ± 0.011 | |
| Thymol | Ctrl | 2.0 (0.5–2.0) | 144.2 ± 21.7 | 60.8 ± 7.6 | 3.47 ± 0.10 a | 0.21 (0.20–0.22) a | 0.031 ± 0.001 a | 0.079 (0.078–0.082) a |
| T1 | 0.0 (0.0–2.5) | 159.6 ± 5.32 | 58.6 ± 3.1 | 3.44 ± 0.09 a,b | 0.21 (0.20–0.23) a | 0.037 ± 0.002 ab | 0.072 (0.061–0.079) a,b | |
| T2 | 1.0 (0.0–1.0) | 165.6 ± 5.68 | 59.4 ± 5.4 | 3.32 ± 0.05 b | 0.23 (0.22–0.24) a,b | 0.037 ± 0.006 a,b | 0.070 (0.063–0.073) a,b | |
| T3 | 2.0 (1.5–2.0) | 145.4 ± 20.5 | 59.0 ± 5.2 | 3.30 ± 0.04 b | 0.25 (0.24–0.28) b | 0.038 ± 0.003 b | 0.067 (0.062–0.073) b | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vieira, R.; Venâncio, C.; Félix, L. Teratogenic, Oxidative Stress and Behavioural Outcomes of Three Fungicides of Natural Origin (Equisetum arvense, Mimosa tenuiflora, Thymol) on Zebrafish (Danio rerio). Toxics 2021, 9, 8. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics9010008
Vieira R, Venâncio C, Félix L. Teratogenic, Oxidative Stress and Behavioural Outcomes of Three Fungicides of Natural Origin (Equisetum arvense, Mimosa tenuiflora, Thymol) on Zebrafish (Danio rerio). Toxics. 2021; 9(1):8. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics9010008
Chicago/Turabian StyleVieira, Raquel, Carlos Venâncio, and Luís Félix. 2021. "Teratogenic, Oxidative Stress and Behavioural Outcomes of Three Fungicides of Natural Origin (Equisetum arvense, Mimosa tenuiflora, Thymol) on Zebrafish (Danio rerio)" Toxics 9, no. 1: 8. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics9010008