
Modulation by Ozone of Glucocorticoid-Regulating Factors in the Lungs in Relation to Stress Axis Reactivity

1
Environmental Health Science and Research Bureau, Healthy Environments and Consumer Safety Branch, Health Canada, Ottawa, ON K1A 0K9, Canada
2
Department of Biochemistry, Microbiology, and Immunology, Faculty of Medicine, University of Ottawa, Ottawa, ON K1H 8M5, Canada
*
Author to whom correspondence should be addressed.
Toxics 2021, 9(11), 290; https://0-doi-org.brum.beds.ac.uk/10.3390/toxics9110290
Submission received: 25 September 2021
/
Revised: 27 October 2021
/
Accepted: 29 October 2021
/
Published: 3 November 2021
(This article belongs to the Special Issue Effects of Exposure to Air Pollution on Respiratory Health)
{kind=link}
{kind=link}
{kind=link}
Abstract
:Exposure to air pollutants increases levels of circulating glucocorticoid stress hormones that exert profound effects relevant to health and disease. However, the nature and magnitude of tissue-level effects are modulated by factors that regulate local glucocorticoid activity; accordingly, inter-individual differences could contribute to susceptibility. In the present study, we characterized effects of ozone (O3) inhalation on glucocorticoid-regulating factors in the lungs of rat strains with contrasting hypothalamic–pituitary–adrenal stress axis responses. Hyper-responsive Fischer (F344) and less responsive Lewis (LEW) rats were exposed to air or 0.8 ppm O3 for 4 h by nose-only inhalation. Levels of the high-specificity and -affinity corticosteroid-binding globulin protein increased in the lungs of both strains proportional to the rise in corticosterone levels following O3 exposure. Ozone reduced the ratio of 11β-hydroxysteroid dehydrogenase type 1 (HSDB1)/HSDB2 mRNA in the lungs of F344 but not LEW, indicating strain-specific transcriptional regulation of the major glucocorticoid metabolism factors that control tissue-level action. Intercellular adhesion molecule (ICAM)-1 and total elastase activity were increased by O3 in both strains, consistent with extravasation and tissue remodeling processes following injury. However, mRNA levels of inflammatory markers were significantly higher in the lungs of O3-exposed LEW compared to F344. The data show that strain differences in the glucocorticoid response to O3 are accompanied by corresponding changes in regulatory factors, and that these effects are collectively associated with a differential inflammatory response to O3. Innate differences in glucocorticoid regulatory factors may modulate the pulmonary effects of inhaled pollutants, thereby contributing to differential susceptibility.
1. Introduction
A growing number of studies have demonstrated that health effects of air pollutants vary significantly within the population; however, mechanisms underlying inter-individual differences in susceptibility remain unclear. Glucocorticoids are major stress hormones produced by the adrenal cortex under regulation of the hypothalamic-pituitary adrenal (HPA) axis [1,2]. Normal adrenal cortical function is essential to resolve systemic and local inflammation, and dysfunction leads to various pathophysiological conditions [3,4]. Acute exposure to air pollutants (ozone [O3], particulate matter, diesel exhaust) has been demonstrated to increase concentrations of plasma glucocorticoids (corticosterone in rodents, cortisol in humans) [5,6,7,8]. We have shown previously that pharmacological inhibition of glucocorticoid synthesis increased O3-induced lung inflammatory responses, suggesting that glucocorticoids play a key role in regulating air pollutant-induced lung inflammatory responses [9]. Moreover, innate differences in stress axis function are associated with differential glucocorticoid and lung inflammatory response to O3 [10].
Although such studies have shown a relationship between circulating glucocorticoid levels and effects in the lungs, it is clear that local factors are critical regulators of glucocorticoid action [11,12,13]. Major factors that alter glucocorticoid availability in the lungs include binding proteins (e.g., corticosteroid -binding globulin [CBG] and albumin), glucocorticoid receptors, and/or 11β-hydroxysteroid dehydrogenase (HSDB)1 and HSDB2 enzymes [11,12,14]. While a number of studies have demonstrated that endogenous glucocorticoids modify lung inflammatory responses [15,16], less attention has been given to local factors in the lungs that may modify glucocorticoid effects.
The present study aimed to investigate how O3 inhalation regulates factors that control glucocorticoid activity in the lungs, and how inter-individual differences in response are reflected in differences in O3-induced inflammatory responses. Fischer 344 (F344) rats have a hyper-responsive stress axis, whereas Lewis (LEW) rats tend to be hypo-responsive [17,18]. In turn, LEW have been found to be susceptible to inflammatory diseases, whereas F344 exhibit resilience [19,20]. Moreover, F344- and LEW-like phenotypes are reported in humans [21,22,23], making them a useful model for investigating modifying factors of glucocorticoid responses in lungs and how these factors contribute to health effects of O3.
2. Material and Methods
2.1. Experimental Model
All experimental methods were reviewed and approved by the Animal Care Committee of Health Canada (Identification code: ACC#2017-004; Date of approval: 8 February 2017), and adhered to the Canadian Council on Animal Care guidelines for humane animal use. Details on housing and experimental procedures were described previously [10]. Briefly, male F344 and LEW rats (~250–300 g) were purchased from Charles River (St. Constant, QC, Canada), and progressively trained in nose-only exposure tubes over 7 consecutive days for acclimatization prior to the study. Nose-only exposures to clean air or 0.8 ppm O3 (800 ± 13 ppb) were performed for 4 h (n = 5/group), followed by anesthetization by isoflurane (5% at 1.5 L of O2/min) and euthanizing by exsanguination immediately at the end of the exposure.
2.2. Biological Samples
The lungs were washed-out with warm saline (37 °C) at 30 mL/kg body weight to obtain bronchoalveolar lavage fluid (BALF). Lavage samples were centrifuged (491× g for 10 min at 8 °C) to pellet cells, and the supernatant was frozen at −80 °C for subsequent analyses. The lungs were excised and frozen at −80 °C for total RNA extraction and transcript analysis.
2.3. Lavage Corticosteroid-Binding Globulin and Soluble Intracellular Adhesion Molecule-1 (ICAM-1)
Lavage corticosteroid-binding globulin and ICAM-1 were quantified using commercially available immunoassay kits (MyBioSource Rat Corticosteroid-binding globulin ELISA Kit, San Diego, CA, USA and Invitrogen ICAM-1 Rat ELISA Kit; Thermo Fisher Scientific, Waltham, MA, USA) according to the manufacturer’s protocols.
2.4. Total Elastase Activity
Total elastase activity in BALF was measured using an elastase activity assay kit (Cayman Chemicals, Ann Arbor, MI, USA). Briefly, elastase activity was measured using a substrate (Z-Ala-Ala-Ala-Ala 2Rh110) and production of the highly fluorescent product (compound R110) was measured using 485 nm excitation and 525 nm emission wavelengths in a fluorescence plate reader (SpectraMax®, Molecular Devices, San Jose, CA, USA).
2.5. RNA Extraction
RNA extraction of lung tissue samples was performed using Trizol reagent according to manufacturer instructions (Invitrogen Canada Inc., Burlington, ON, Canada). Total RNA was quantified using the Quant-iT™ RiboGreen® RNA Assay Kit (Molecular Probes, Eugene, OR, USA), and quality was verified (RNA quality index (RQI)/RNA integrity number greater than 7.5) using Experion™ RNA StdSens analysis kits (Bio-Rad Laboratories Ltd., Mississauga, ON, Canada).
2.6. Gene Expression Analyses
Complementary DNA (cDNA) synthesis was performed using MultiScribe reverse transcriptase and random hexamers (Applied Biosystems, Mississauga, ON, Canada). Universal Probe Library design software (Roche Diagnostics, Laval, QC, Canada) was used for primer design (Table S1), and reaction efficiency (>90%) was verified using a cDNA dilution series. PCR reactions were performed in duplicate on 384-well plates using BrightGreen 2× qPCR MasterMix (Applied Biological Materials Inc. Richmond, BC, Canada) in a spectrofluorometric thermal cycler (Lightcycler 480, Roche Diagnostics) according to the following protocol: 3 min incubation at 95 °C followed by 50 cycles of denaturation at 95 °C, annealing at 60 °C, and elongation at 72 °C, each for 10 s. Product purity was verified by confirming a single peak for each analyte via melt curve analysis. Relative gene expression was calculated using β-actin as the reference gene according to the delta-delta Ct method [24].
2.7. Statistical Analyses
Sigma-Plot 12.5 (Systat Software Inc., San Jose, CA, USA) was used for all statistical analyses and generation of figures. Data were transformed to meet the requirements of normality and equal variance as warranted. Significant effects were determined by two-way ANOVA (factors Ozone (0, 0.8 ppm) and Strain (F344, LEW)) followed by the Holm–Sidak multiple comparison procedure (α = 0.05). Pearson correlation was conducted to test the relationships between lavage corticosterone and CBG.
3. Results
3.1. Effects of O3 on Glucocorticoid-Regulating Factors in the Lungs
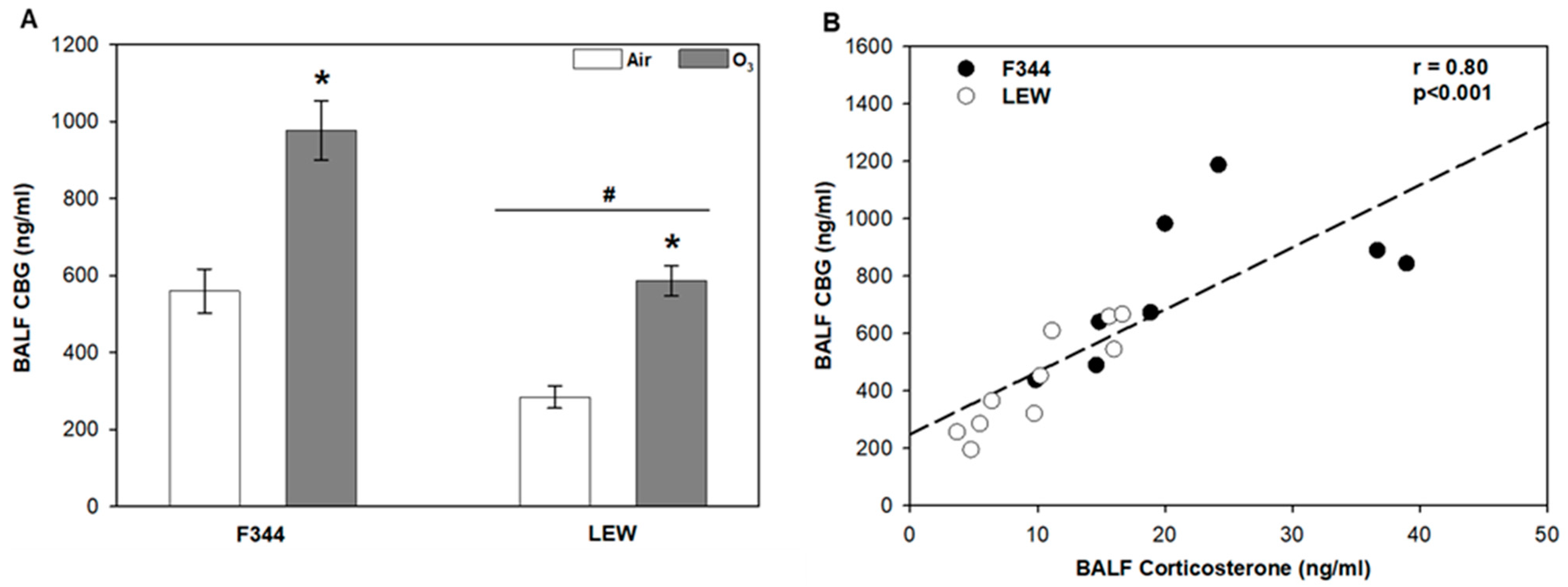
Levels of the high-specificity and -affinity corticosteroid-binding globulin (CBG) were elevated in F344 compared to LEW (Strain main effect, p < 0.001; Figure 1A). Exposure to O3 significantly increased lavage CBG (~1.7–2 fold) in both F344 and LEW, albeit with lower overall levels in LEW (Ozone main effect, p < 0.001). The profiles were similar to corticosterone profiles in plasma and lung lavage described previously [10,25]. Overall, there was a significant positive association between corticosterone and CBG recovered by bronchoalveolar lavage (r = 0.80; p < 0.001; Figure 1B).
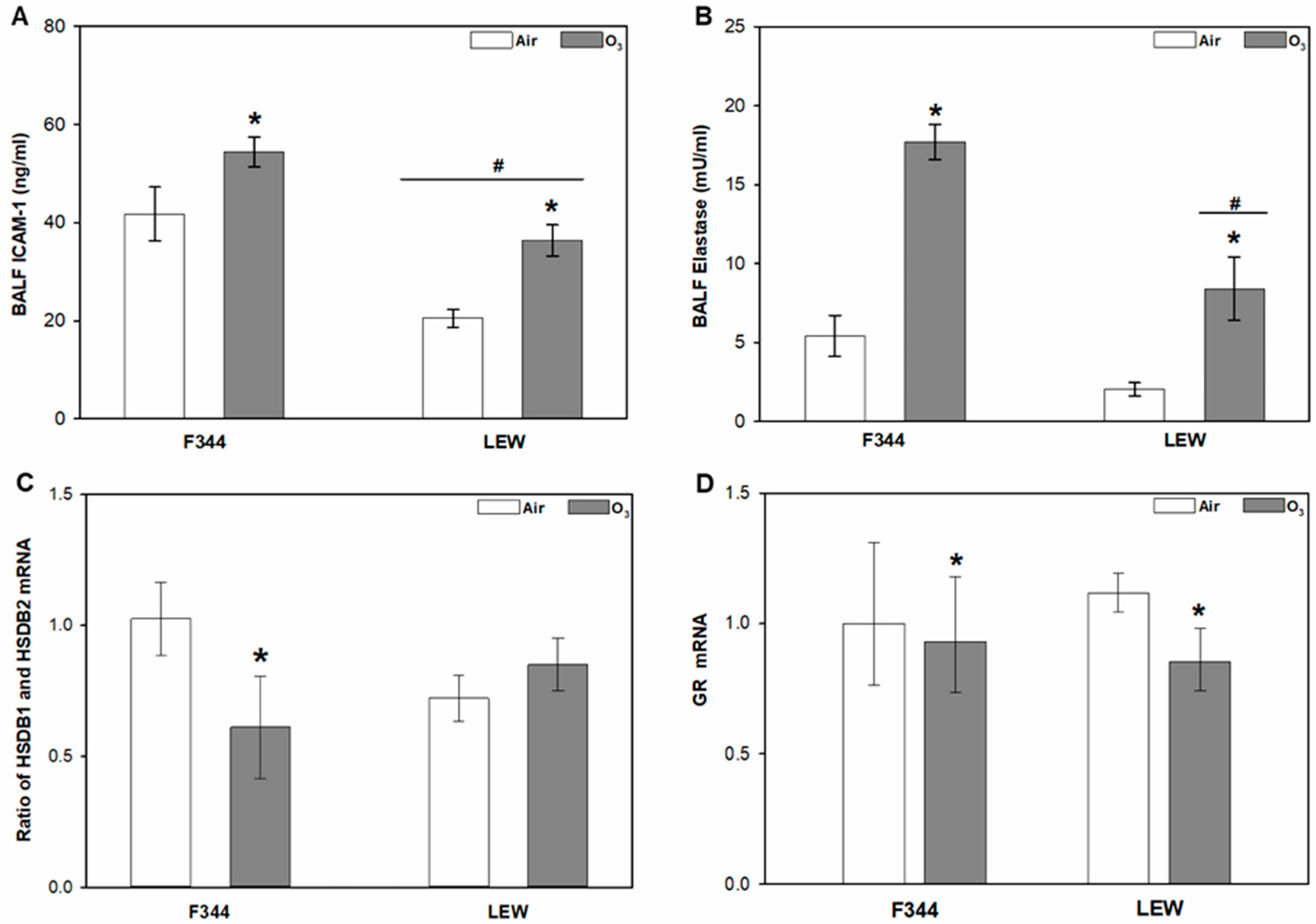
In addition to binding of glucocorticoids, other factors controlling glucocorticoid entry and action in the lungs include regulation of lung permeability, glucocorticoid metabolism, and glucocorticoid receptors. Levels of ICAM-1, a factor regulating inflammatory response and lung permeability, were significantly higher in F344 compared to LEW, and were increased by O3 in both strains (Strain main effect, p < 0.001; Ozone main effect, p < 0.001; Figure 2A). Ozone exposure also increased total elastase activity in lavage collected from both F344 and LEW when compared to their air-exposed counterparts, with greater elastase activity observed in F344 compared to LEW (Ozone × Strain interactions; p = 0.048; Figure 2B). The mRNA ratio of the glucocorticoid metabolism factors HSDB1 and HSDB2 was reduced by approximately 40% by O3 in F344, an effect not observed in LEW (Ozone × Strain interaction; p = 0.03; Figure 2C). Overall, mRNA levels of the glucocorticoid receptor (GR) were decreased after O3 exposure (Ozone main effect, p < 0.01; Figure 2D), but no Strain effect was observed. No effect of Ozone or Strain was observed for mineralocorticoid receptor (MCR) gene expression (Figure S1).
3.2. O3-Dependent Inflammatory Signaling in the Lungs
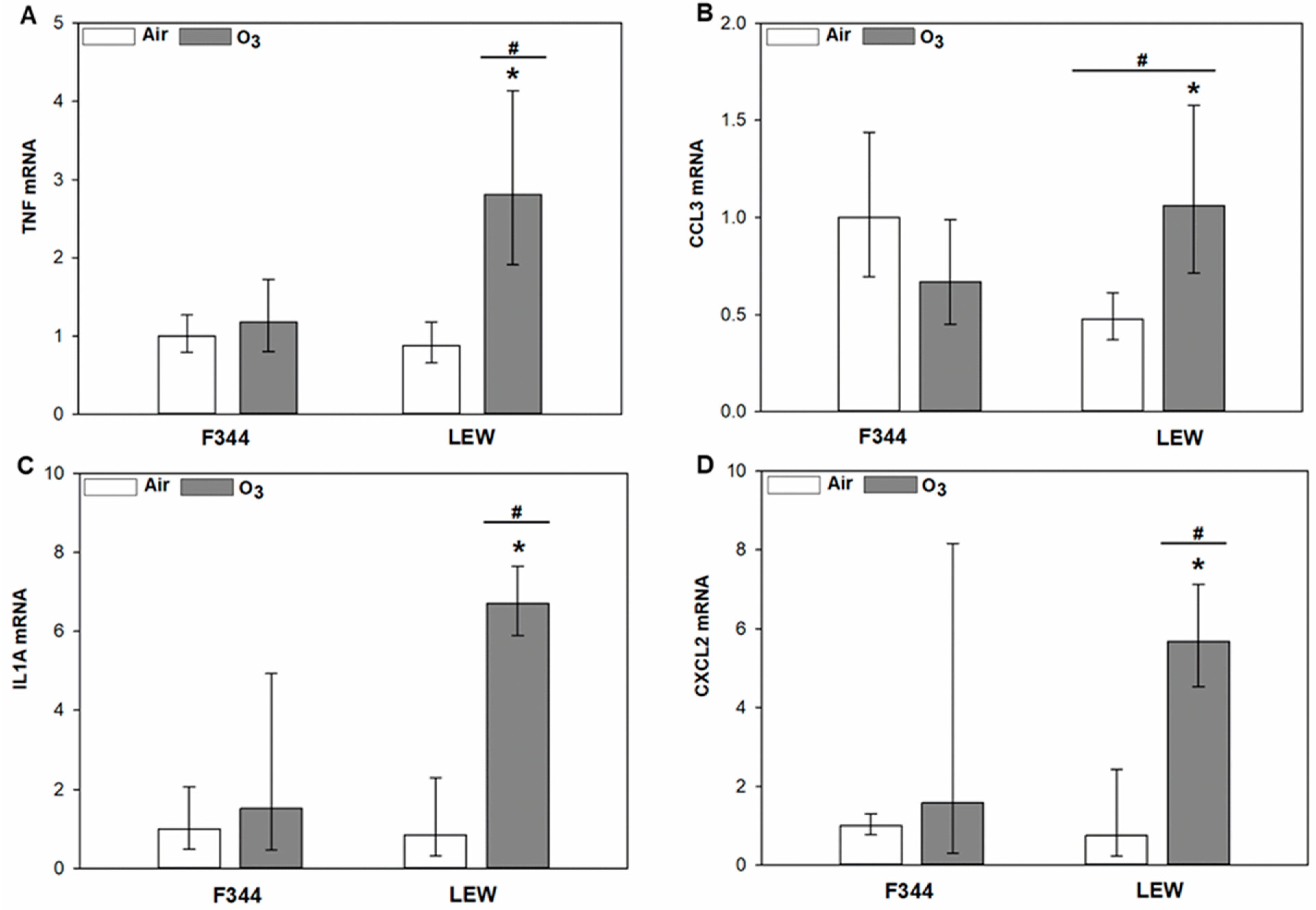
Strain-dependent differences in the response of inflammatory genes to exposure were observed in lung tissue samples (Figure 3A–D). Compared to F344, the response of pro-inflammatory genes was significantly higher in LEW after O3. Significant Ozone × Strain interactions were observed for tumour necrosis factor (TNF; p < 0.001), C-C motif chemokine ligand 3 (CCL3; p = 0.001), interleukin-1α (IL1A; p = 0.01), and C-X-C motif chemokine ligand 2 (CXCL2; p = 0.006). Expression of inflammatory genes increased by 2.2–7.9 fold in O3 exposed LEW when compared to their air exposure counterparts.
4. Discussion
Immunomodulatory effects of endogenous glucocorticoids are well understood, and it has been demonstrated that impaired glucocorticoid release or signaling can modify lung inflammatory processes [16,26,27]. Since the ability of glucocorticoids to regulate inflammation is linked to their availability at sites of inflammation, factors that modify glucocorticoid availability in the lungs could impact air pollutant-induced lung inflammatory responses. We have previously demonstrated that exposure to O3 increases levels of the glucocorticoid corticosterone in the lungs of rat strains with distinct stress axis responsiveness, but that the profile of response differed by strain [25]. This prompted us to investigate how factors that modify glucocorticoid availability in the lungs differ across strains with contrasting stress axis responsiveness, and how exposure to an air pollutant may impact these factors. In the present study, we demonstrated that O3 exposure modified several factors that affect local glucocorticoid availability in the lungs; a subset of effects differed by strain. The data suggest that glucocorticoid regulatory factors may contribute to pulmonary effects of air pollutants, providing insight into a potential mechanism underlying differential sensitivity to pollutant exposure.
The major factors that affect glucocorticoid availability in the lungs include glucocorticoid-binding proteins, neutrophil elastase, and glucocorticoid metabolism enzymes HSDB1 and 2. Although it is suggested that only free glucocorticoids enter peripheral tissues [28], plasma glucocorticoid-binding proteins (CBG and albumin) are reported in the lungs, and their levels vary between individuals and are impacted by various diseases [12,28,29,30]. The ozone-dependent increase in CBG recovered in bronchoalveolar lavage fluid generally tracked levels of corticosterone, with higher levels observed in F344 compared to LEW. We have previously shown that acute exposure to 0.8 ppm O3 decreased total cell recovery by bronchoalveolar lavage in both strains immediately after exposure, consistent with increased production of adhesion factors; at the same time, we observed higher levels of lung injury markers in F344 compared to LEW [10]. In the present study, soluble ICAM-1, a marker of injury and lung permeability [31], was increased by exposure to O3 in both F344 and LEW, in line with the notion that glucocorticoid-binding proteins may enter from circulation after acute O3 exposure. The lungs are also a site of CBG synthesis [28,30], suggesting the potential for involvement of local production in regulating glucocorticoid activity.
It has been demonstrated that neutrophil elastase cleaves a reactive center loop of CBG, thereby enabling the local release of glucocorticoids at the site of inflammation [12,28]. In the present study, lavage elastase activity was significantly increased in F344 and LEW after O3 exposure, with more pronounced activity observed in F344. However, the substrate used was not specific to neutrophil or macrophage elastase activity; given the lack of neutrophils recovered by lung lavage at this early time point [25], it is possible that the present study measured macrophage elastase (MMP12) activity instead of neutrophil elastase. Although macrophage elastase (MMP-12) is reported to degrade extracellular matrix components including elastin and is implicated in tissue remodelling processes [32], there is no evidence that macrophage elastase cleaves CBG. Nevertheless, increased MMP12 activity may contribute to lung permeability that results in an influx of plasma glucocorticoid-binding proteins to the lungs and that in turn impact local glucocorticoid availability [33].
Regulation of other factors implicated in tissue-level control of glucocorticoid action may also be important. 11β-hydroxysteroid dehydrogenases (HSDB1 and 2) are considered intracellular regulators of tissue glucocorticoid action [14], as these enzymes control intracellular metabolism of endogenous glucocorticoids. It has been demonstrated that levels of these enzymes can vary with disease states, and considerable inter-individual variability exists in HSDB activity in humans [14,34]. In the present study, O3 exposure reduced the ratio of HSDB1/HSDB2 mRNA in the lungs of F344 but not LEW, suggesting strain-specific differences in the regulation of glucocorticoid metabolism in these animals. Glucocorticoid signalling in target tissues is, in turn, mediated primarily through binding to receptors; moreover, there is significant inter-individual variability in GR expression, and chronic impairment in GR function can lead to lung glucocorticoid resistance [35,36]. Here, O3 exposure downregulated lung GR expression in both F344 and LEW, consistent with known effects of glucocorticoid exposure [11,37]. The decrease in lung GR mRNA suggests that despite the strain differences in CBG and glucocorticoid metabolism factors, lung glucocorticoid levels in both rat strains reached a threshold sufficient to induce biological effects.
Differential regulation by air pollutants of factors important to the control of glucocorticoid may have physiological relevance. We have previously shown that pharmacological inhibition of the O3-dependent increase in corticosterone was associated with increased levels of inflammatory cytokines in the lungs [9]. In the present study, expression of several pro-inflammatory genes was increased by O3 in the lungs of LEW but not F344. These inter-strain differences may be due to the differences in both the levels of glucocorticoids and those factors that regulate their entry and activity. Indeed, it has been demonstrated that glucocorticoids at lower levels facilitate inflammatory responses, whereas at higher levels they dampen such responses [38]. Accordingly, the results are consistent with the notion that the higher glucocorticoid levels in F344 rats reduce lung inflammatory responses to O3 in F344, compared to less inhibition by the lower glucocorticoid levels in LEW. In a randomised double-blind crossover study, we recently showed that short-term exposure to diesel exhaust increased plasma cortisol levels, with the greatest effects seen in those with asthma or with risk alleles for antioxidant genes [7]. Investigation of inter-individual differences in glucocorticoid response and regulation in the lungs may provide insight into factors affecting the course and treatment of inflammatory lung diseases.
Our study provides evidence that local glucocorticoid-regulatory factors are affected by O3 exposure and, therefore, may play a role in the regulation of glucocorticoid signaling in the lungs following pollutant exposure. Furthermore, our results suggest that inter-individual differences in stress axis function may contribute to differential inflammatory responses following pollutant exposure. Given the important role played by glucocorticoids in the regulation of lung inflammatory responses [15,16,39], variability in glucocorticoid regulatory factors in lungs that subsequently modify glucocorticoid availability could contribute to differential susceptibility to air pollutants.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/toxics9110290/s1, Figure S1: Expression of a glucocorticoid-regulating gene (mineralocorticoid receptor; MCR) in lung tissue of Fischer (F344) and Lewis (LEW) rats after exposure to air or 0.8 ppm ozone (O3) for 4 h. Real-time PCR results are expressed as geometric mean of fold-change relative to the F344 air control ±95% confidence intervals, Table S1: Gene-specific primer sequences for the quantitative real-time PCR used in this study.
Author Contributions
J.T.: Investigation; Formal Analysis; Writing—Original Draft Preparation. E.M.T.: Conceptualization; Funding Acquisition; Methodology; Supervision; Writing—Review and Editing. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by Health Canada through the Clean Air Regulatory Agenda and the Addressing Air Pollution Horizontal Initiative.
Institutional Review Board Statement
All experimental methods were reviewed and approved by the Animal Care Committee of Health Canada (Identification code: ACC#2017-004; Date of approval: 8 February 2017), and adhered to the Canadian Council on Animal Care guidelines for humane animal use.
Acknowledgments
Authors would like to thank Josée Guénette, Kevin Curtin, Alain Filiatreault, Marjolaine Godbout-Cheliak, Dalibor Breznan, Martha Navarro, Sandra Brenton, Scott Smith, and Michelle Lalande for assistance with the study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Katsu, Y.; Iguchi, T. Subchapter 95A—Corticosterone. In Handbook of Hormones; Takei, Y., Ando, H., Tsutsui, K., Eds.; Academic Press: San Diego, CA, USA, 2016. [Google Scholar] [CrossRef]
- Uchoa, E.T.; Aguilera, G.; Herman, J.P.; Fiedler, J.L.; Deak, T.; Cordeiro de Sousa, M.B. Novel Aspects of Hypothalamic-Pituitary-Adrenal Axis Regulation and Glucocorticoid Actions. J. Neuroendocrinol. 2014, 26, 557–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chrousos, G.P. The Hypothalamic–Pituitary–Adrenal Axis and Immune-Mediated Inflammation. N. Engl. J. Med. 1995, 332, 1351–1363. [Google Scholar] [CrossRef] [PubMed]
- Silverman, M.N.; Sternberg, E.M. Glucocorticoid Regulation of Inflammation and Its Functional Correlates: From HPA Axis to Glucocorticoid Receptor Dysfunction. Ann. N. Y. Acad. Sci. 2012, 1261, 55–63. [Google Scholar] [CrossRef]
- Li, H.; Cai, J.; Chen, R.; Zhao, Z.; Ying, Z.; Wang, L.; Chen, J.; Hao, K.; Kinney, P.L.; Chen, H.; et al. Particulate Matter Exposure and Stress Hormone Levels: A Randomized, Double-Blind, Crossover Trial of Air Purification. Circulation 2017, 136, 618–627. [Google Scholar] [CrossRef]
- Miller, D.B.; Ghio, A.J.; Karoly, E.D.; Bell, L.N.; Snow, S.J.; Madden, M.C.; Soukup, J.; Cascio, W.E.; Gilmour, M.I.; Kodavanti, U.P. Ozone Exposure Increases Circulating Stress Hormones and Lipid Metabolites in Humans. Am. J. Respir. Crit. Care Med. 2016, 193, 1382–1391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomson, E.M.; Filiatreault, A.; Williams, A.; Rider, C.F.; Carlsten, C. Exposure to Diesel Exhaust and Plasma Cortisol Response: A Randomized Double-Blind Crossover Study. Environ. Health Perspect. 2021, 129, 037701. [Google Scholar] [CrossRef] [PubMed]
- Thomson, E.M.; Vladisavljevic, D.; Mohottalage, S.; Kumarathasan, P.; Vincent, R. Mapping Acute Systemic Effects of Inhaled Particulate Matter and Ozone: Multiorgan Gene Expression and Glucocorticoid Activity. Toxicol. Sci. 2013, 135, 169–181. [Google Scholar] [CrossRef] [Green Version]
- Thomson, E.M.; Pal, S.; Guénette, J.; Wade, M.G.; Atlas, E.; Holloway, A.C.; Williams, A.; Vincent, R. Ozone Inhalation Provokes Glucocorticoid-Dependent and -Independent Effects on Inflammatory and Metabolic Pathways. Toxicol. Sci. 2016, 152, 17–28. [Google Scholar] [CrossRef] [Green Version]
- Thomas, J.; Guénette, J.; Thomson, E.M. Stress Axis Variability Is Associated with Differential Ozone-Induced Lung Inflammatory Signaling and Injury Biomarker Response. Environ. Res. 2018, 167, 751–758. [Google Scholar] [CrossRef] [PubMed]
- Kadmiel, M.; Cidlowski, J.A. Glucocorticoid Receptor Signaling in Health and Disease. Trends Pharmacol. Sci. 2013, 34, 518–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moisan, M.-P.; Castanon, N. Emerging Role of Corticosteroid-Binding Globulin in Glucocorticoid-Driven Metabolic Disorders. Front. Endocrinol. 2016, 7, 160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pemberton, P.A.; Stein, P.E.; Pepys, M.B.; Potter, J.M.; Carrell, R.W. Hormone Binding Globulins Undergo Serpin Conformational Change in Inflammation. Nature 1988, 336, 257–258. [Google Scholar] [CrossRef] [PubMed]
- Chapman, K.; Holmes, M.; Seckl, J. 11β-Hydroxysteroid Dehydrogenases: Intracellular Gate-Keepers of Tissue Glucocorticoid Action. Physiol. Rev. 2013, 93, 1139–1206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watterberg, K.L.; Scott, S.M.; Backstrom, C.; Gifford, K.L.; Cook, K.L. Links between Early Adrenal Function and Respiratory Outcome in Preterm Infants: Airway Inflammation and Patent Ductus Arteriosus. Pediatrics 2000, 105, 320–324. [Google Scholar] [CrossRef]
- Zhang, S.; Shen, Z.; Hu, G.; Liu, R.; Zhang, X. Effects of Endogenous Glucocorticoids on Allergic Inflammation and TH1/TH2 Balance in Airway Allergic Disease. Ann. Allergy Asthma Immunol. 2009, 103, 525–534. [Google Scholar] [CrossRef]
- Cohen, H.; Zohar, J.; Gidron, Y.; Matar, M.A.; Belkind, D.; Loewenthal, U.; Kozlovsky, N.; Kaplan, Z. Blunted HPA Axis Response to Stress Influences Susceptibility to Posttraumatic Stress Response in Rats. Biol. Psychiatry 2006, 59, 1208–1218. [Google Scholar] [CrossRef] [PubMed]
- Dhabhar, F.S.; McEwen, B.S.; Spencer, R.L. Stress Response, Adrenal Steroid Receptor Levels and Corticosteroid-Binding Globulin Levels - a Comparison between Sprague-Dawley, Fischer 344 and Lewis Rats. Brain Res. 1993, 616, 89–98. [Google Scholar] [CrossRef]
- Karalis, K.; Crofford, L.; Wilder, R.L.; Chrousos, G.P. Glucocorticoid and/or Glucocorticoid Antagonist Effects in Inflammatory Disease-Susceptible Lewis Rats and Inflammatory Disease-Resistant Fischer Rats. Endocrinology 1995, 136, 3107–3112. [Google Scholar] [CrossRef] [PubMed]
- Sternberg, E.M.; Hill, J.M.; Chrousos, G.P.; Kamilaris, T.; Listwak, S.J.; Gold, P.W.; Wilder, R.L. Inflammatory Mediator-Induced Hypothalamic-Pituitary-Adrenal Axis Activation Is Defective in Streptococcal Cell Wall Arthritis-Susceptible Lewis Rats. Proc. Natl. Acad. Sci. USA 1989, 86, 2374–2378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charmandari, E.; Tsigos, C.; Chrousos, G. Endocrinology of the Stress Response. Annu. Rev. Physiol. 2005, 67, 259–284. [Google Scholar] [CrossRef]
- Ebner, K.; Singewald, N. Individual Differences in Stress Susceptibility and Stress Inhibitory Mechanisms. Curr. Opin. Behav. Sci. 2017, 14, 54–64. [Google Scholar] [CrossRef]
- Elenkov, I.J.; Kvetnansky, R.; Hashiramoto, A.; Bakalov, V.K.; Link, A.A.; Zachman, K.; Crane, M.; Jezova, D.; Rovensky, J.; Dimitrov, M.A.; et al. Low- versus High-Baseline Epinephrine Output Shapes Opposite Innate Cytokine Profiles: Presence of Lewis- and Fischer-like Neurohormonal Immune Phenotypes in Humans? J. Immunol. 2008, 181, 1737–1745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative Pcr and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Thomas, J.; Stalker, A.; Breznan, D.; Thomson, E.M. Ozone-Dependent Increases in Lung Glucocorticoids and Macrophage Response: Effect Modification by Innate Stress Axis Function. Environ. Toxicol. Pharmacol. 2021, 86, 103662. [Google Scholar] [CrossRef] [PubMed]
- Baker, R.W.; Walker, B.R.; Shaw, R.J.; Honour, J.W.; Jessop, D.S.; Lightman, S.L.; Zumla, A.; Rook, G.a.W. Increased Cortisol: Cortisone Ratio in Acute Pulmonary Tuberculosis. Am. J. Respir. Crit. Care. Med. 2000, 162, 1641–1647. [Google Scholar] [CrossRef]
- Corvol, H.; Nathan, N.; Charlier, C.; Chadelat, K.; Le Rouzic, P.; Tabary, O.; Fauroux, B.; Henrion-Caude, A.; Feingold, J.; Boelle, P.-Y.; et al. Glucocorticoid Receptor Gene Polymorphisms Associated with Progression of Lung Disease in Young Patients with Cystic Fibrosis. Respir. Res. 2007, 8, 88. [Google Scholar] [CrossRef] [Green Version]
- Hammond, G.L. Plasma Steroid-Binding Proteins: Primary Gatekeepers of Steroid Hormone Action. J. Endocrinol. 2016, 230, R13–R25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aman, J.; van der Heijden, M.; van Lingen, A.; Girbes, A.R.J.; van Nieuw Amerongen, G.P.; van Hinsbergh, V.W.M.; Groeneveld, A.B.J. Plasma Protein Levels Are Markers of Pulmonary Vascular Permeability and Degree of Lung Injury in Critically Ill Patients with or at Risk for Acute Lung Injury/Acute Respiratory Distress Syndrome. Crit. Care Med. 2011, 39, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Gulfo, J.; Ledda, A.; Gea-Sorlí, S.; Bonjoch, L.; Closa, D.; Grasa, M.; Esteve, M. New Roles for Corticosteroid Binding Globulin and Opposite Expression Profiles in Lung and Liver. PLoS ONE 2016, 11, e0146497. [Google Scholar] [CrossRef] [Green Version]
- Spadaro, S.; Fogagnolo, A.; Campo, G.; Zucchetti, O.; Verri, M.; Ottaviani, I.; Tunstall, T.; Grasso, S.; Scaramuzzo, V.; Murgolo, F.; et al. Markers of Endothelial and Epithelial Pulmonary Injury in Mechanically Ventilated COVID-19 ICU Patients. Crit. Care 2021, 25, 74. [Google Scholar] [CrossRef]
- Nénan, S.; Boichot, E.; Lagente, V.; Bertrand, C.P. Macrophage Elastase (MMP-12): A pro-Inflammatory Mediator? Mem. Inst. Oswaldo Cruz 2005, 100, 167–172. [Google Scholar] [CrossRef] [Green Version]
- Elkington, P.T.G.; Friedland, J.S. Matrix Metalloproteinases in Destructive Pulmonary Pathology. Thorax 2006, 61, 259–266. [Google Scholar] [CrossRef] [Green Version]
- Soldan, M.; Nagel, G.; Losekam, M.; Ernst, M.; Maser, E. Interindividual Variability in the Expression and NNK Carbonyl Reductase Activity of 11β-Hydroxysteroid Dehydrogenase 1 in Human Lung. Cancer Lett. 1999, 145, 49–56. [Google Scholar] [CrossRef]
- Rider, C.F.; Carlsten, C. Air Pollution and Resistance to Inhaled Glucocorticoids: Evidence, Mechanisms and Gaps to Fill. Pharmacol. Ther. 2019, 194, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Song, Q.-Q.; Xie, W.-Y.; Tang, Y.-J.; Zhang, J.; Liu, J. Genetic Variation in the Glucocorticoid Pathway Involved in Interindividual Differences in the Glucocorticoid Treatment. Pharmacogenomics 2017, 18, 293–316. [Google Scholar] [CrossRef]
- Bekhbat, M.; Rowson, S.A.; Neigh, G.N. Checks and Balances: The Glucocorticoid Receptor and NFκB in Good Times and Bad. Front. Neuroendocrinol. 2017, 46, 15–31. [Google Scholar] [CrossRef] [PubMed]
- Cain, D.W.; Cidlowski, J.A. Immune Regulation by Glucocorticoids. Nat. Rev. Immunol. 2017, 17, 233–247. [Google Scholar] [CrossRef] [PubMed]
- Coutinho, A.E.; Chapman, K.E. The Anti-Inflammatory and Immunosuppressive Effects of Glucocorticoids, Recent Developments and Mechanistic Insights. Mol. Cell. Endocrinol. 2011, 335, 2–13. [Google Scholar] [CrossRef]
Figure 1.
Strain comparison of lung lavage corticosteroid-binding globulin (CBG) after ozone inhalation. A. CBG levels in bronchoalveolar lavage fluid (BALF) in Fischer (F344) and Lewis (LEW) exposed to air or 0.8 ppm ozone (O3) for 4 h. * Ozone main effect, p < 0.001; # Strain main effect, p < 0.001; two-way analysis of variance (ANOVA) with Holm–Sidak post-hoc test; n = 4–5 rats/strain/treatment. Data are expressed as mean ± SEM. B. Relationship between BALF corticosterone and CBG in F344 and LEW (Pearson correlation).
Figure 1.
Strain comparison of lung lavage corticosteroid-binding globulin (CBG) after ozone inhalation. A. CBG levels in bronchoalveolar lavage fluid (BALF) in Fischer (F344) and Lewis (LEW) exposed to air or 0.8 ppm ozone (O3) for 4 h. * Ozone main effect, p < 0.001; # Strain main effect, p < 0.001; two-way analysis of variance (ANOVA) with Holm–Sidak post-hoc test; n = 4–5 rats/strain/treatment. Data are expressed as mean ± SEM. B. Relationship between BALF corticosterone and CBG in F344 and LEW (Pearson correlation).

Figure 2.
Glucocorticoid-regulating factors in the lungs in Fischer (F344) and Lewis (LEW) rats after exposure to air or 0.8 ppm ozone (O3) for 4 h. (A) Protein levels in bronchoalveolar lavage fluid (BALF) of intercellular adhesion molecule-1 (ICAM-1); Ozone (p = 0.001) and Strain (p < 0.001) main effects. (B) Elastase activity in BALF; Ozone × Strain interaction, p = 0.048. (C) 11β-Hydroxysteroid dehydrogenase 1 (HSDB1) and HSDB2 ratio (calculated from the mRNA abundance data); Ozone main effect, p = 0.030. (D) Glucocorticoid receptor (GR) mRNA levels in lung tissue; Ozone main effect, p = 0.01. * 0.8 vs. 0 ppm ozone within given strain (except for GR where an asterisk indicates the Ozone main effect across strains); # LEW vs. F344 within indicated exposure level; two-way ANOVA followed by Holm–Sidak post-hoc test; n = 4–5 rats. For (A–C), data are expressed as mean ± SEM and for (D) data are expressed as geometric mean of fold-change relative to the F344 air control ± 95% confidence intervals.
Figure 2.
Glucocorticoid-regulating factors in the lungs in Fischer (F344) and Lewis (LEW) rats after exposure to air or 0.8 ppm ozone (O3) for 4 h. (A) Protein levels in bronchoalveolar lavage fluid (BALF) of intercellular adhesion molecule-1 (ICAM-1); Ozone (p = 0.001) and Strain (p < 0.001) main effects. (B) Elastase activity in BALF; Ozone × Strain interaction, p = 0.048. (C) 11β-Hydroxysteroid dehydrogenase 1 (HSDB1) and HSDB2 ratio (calculated from the mRNA abundance data); Ozone main effect, p = 0.030. (D) Glucocorticoid receptor (GR) mRNA levels in lung tissue; Ozone main effect, p = 0.01. * 0.8 vs. 0 ppm ozone within given strain (except for GR where an asterisk indicates the Ozone main effect across strains); # LEW vs. F344 within indicated exposure level; two-way ANOVA followed by Holm–Sidak post-hoc test; n = 4–5 rats. For (A–C), data are expressed as mean ± SEM and for (D) data are expressed as geometric mean of fold-change relative to the F344 air control ± 95% confidence intervals.

Figure 3.
Expression of inflammatory genes in the lungs of Fischer (F344) and Lewis (LEW) rats after exposure to air or 0.8 ppm ozone (O3) for 4 h. Real-time PCR results are expressed as geometric mean of fold-change relative to the F344 air control ± 95% confidence intervals. (A). TNF. Ozone × Strain interaction, p < 0.001. (B). CCL3. Ozone × Strain interaction, p = 0.001. (C). IL1A. Ozone × Strain interaction, p = 0.01. (D). CXCL2. Ozone × Strain interaction, p = 0.006. * 0.8 vs. 0 ppm ozone within given strain and # LEW vs. F344 within indicated exposure level (two-way ANOVA followed by Holm–Sidak post-hoc test; n = 4–5 rats).
Figure 3.
Expression of inflammatory genes in the lungs of Fischer (F344) and Lewis (LEW) rats after exposure to air or 0.8 ppm ozone (O3) for 4 h. Real-time PCR results are expressed as geometric mean of fold-change relative to the F344 air control ± 95% confidence intervals. (A). TNF. Ozone × Strain interaction, p < 0.001. (B). CCL3. Ozone × Strain interaction, p = 0.001. (C). IL1A. Ozone × Strain interaction, p = 0.01. (D). CXCL2. Ozone × Strain interaction, p = 0.006. * 0.8 vs. 0 ppm ozone within given strain and # LEW vs. F344 within indicated exposure level (two-way ANOVA followed by Holm–Sidak post-hoc test; n = 4–5 rats).

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Thomas, J.; Thomson, E.M. Modulation by Ozone of Glucocorticoid-Regulating Factors in the Lungs in Relation to Stress Axis Reactivity. Toxics 2021, 9, 290. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics9110290
AMA Style
Thomas J, Thomson EM. Modulation by Ozone of Glucocorticoid-Regulating Factors in the Lungs in Relation to Stress Axis Reactivity. Toxics. 2021; 9(11):290. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics9110290
Chicago/Turabian StyleThomas, Jith, and Errol M. Thomson. 2021. "Modulation by Ozone of Glucocorticoid-Regulating Factors in the Lungs in Relation to Stress Axis Reactivity" Toxics 9, no. 11: 290. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics9110290
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.