
Cigarette Smoke Extract Produces Superoxide in Aqueous Media by Reacting with Bicarbonate

 , and
, and 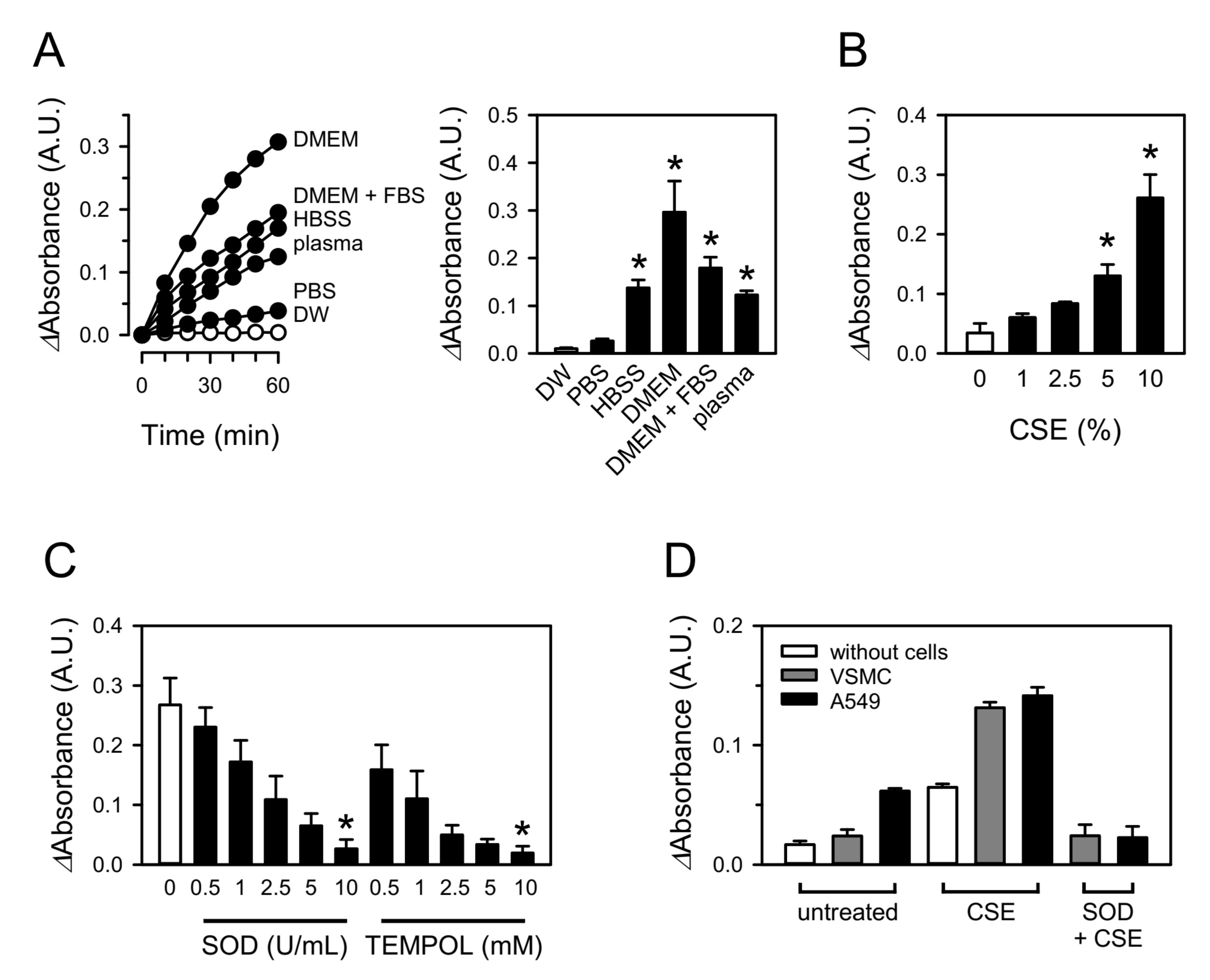{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Animals
2.3. Preparation of Blood Plasma
2.4. Preparation of CSE and TPM
2.5. Cells and Cell Culture
2.6. Superoxide Measurement Using WST-1
2.7. EPR Spectroscopy
2.8. Assessment of Protein Carbonylation by CSE
2.9. Statistical Analyses
3. Results
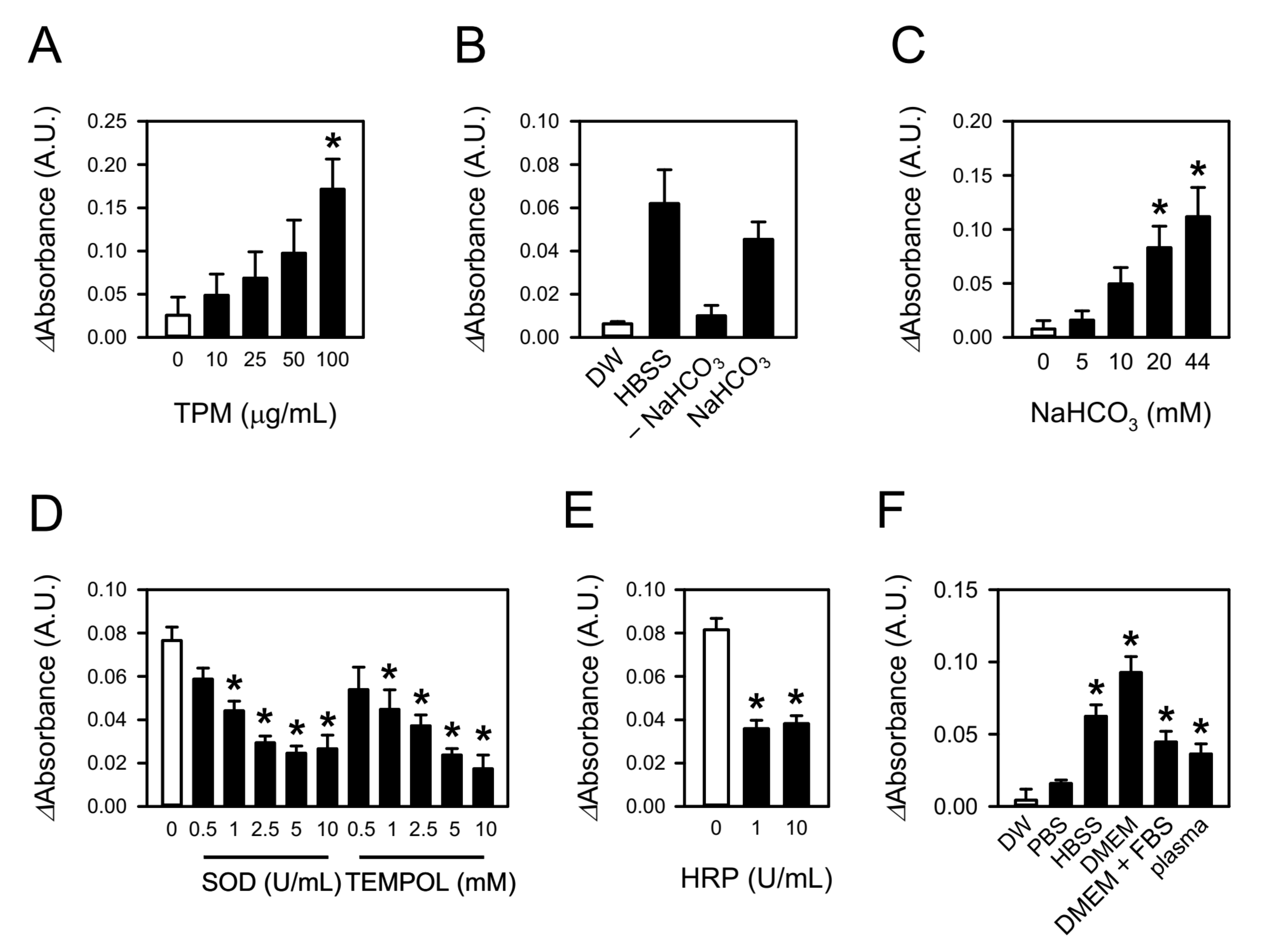
3.1. CSE Produces Superoxide in Aqueous Solutions
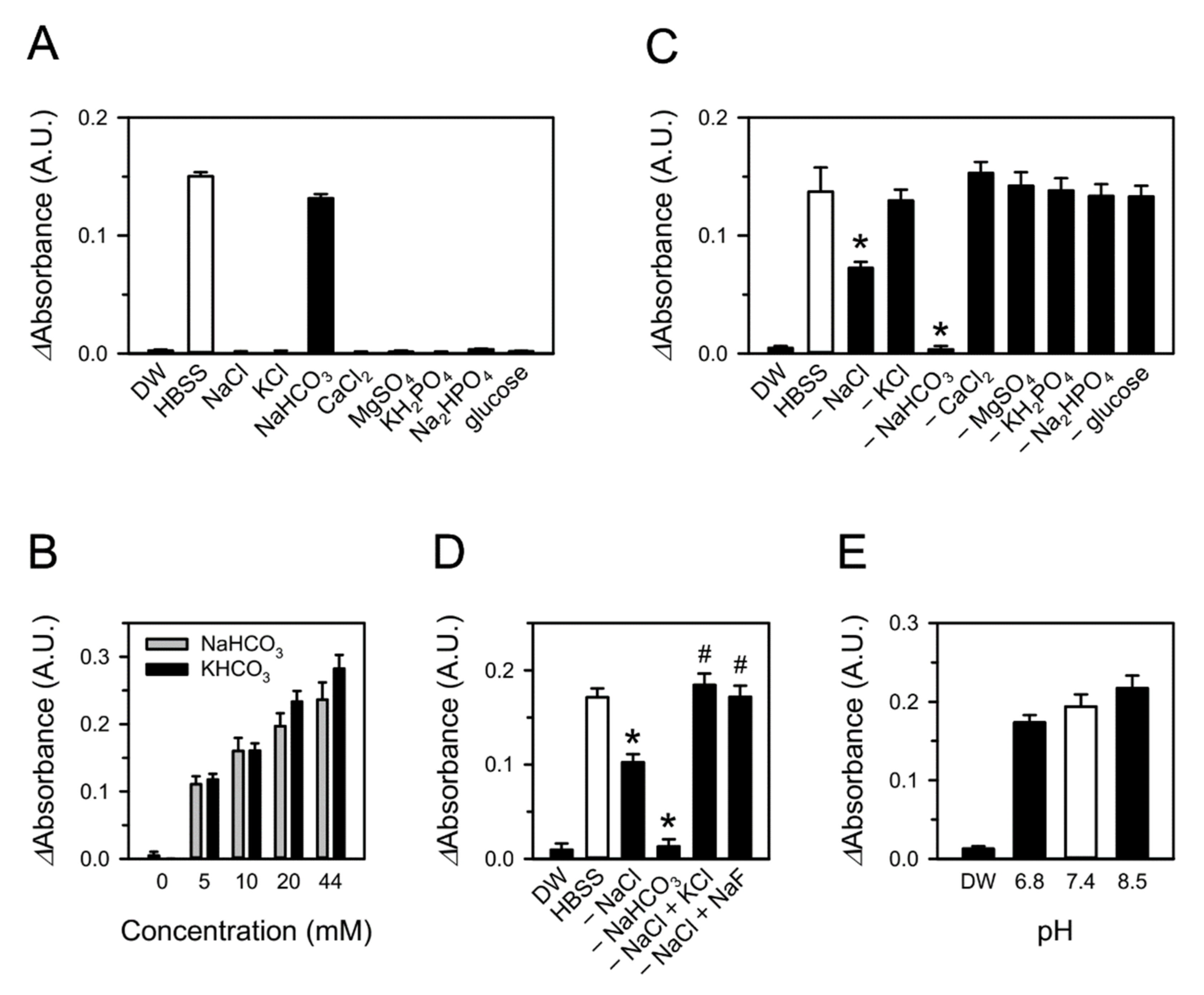
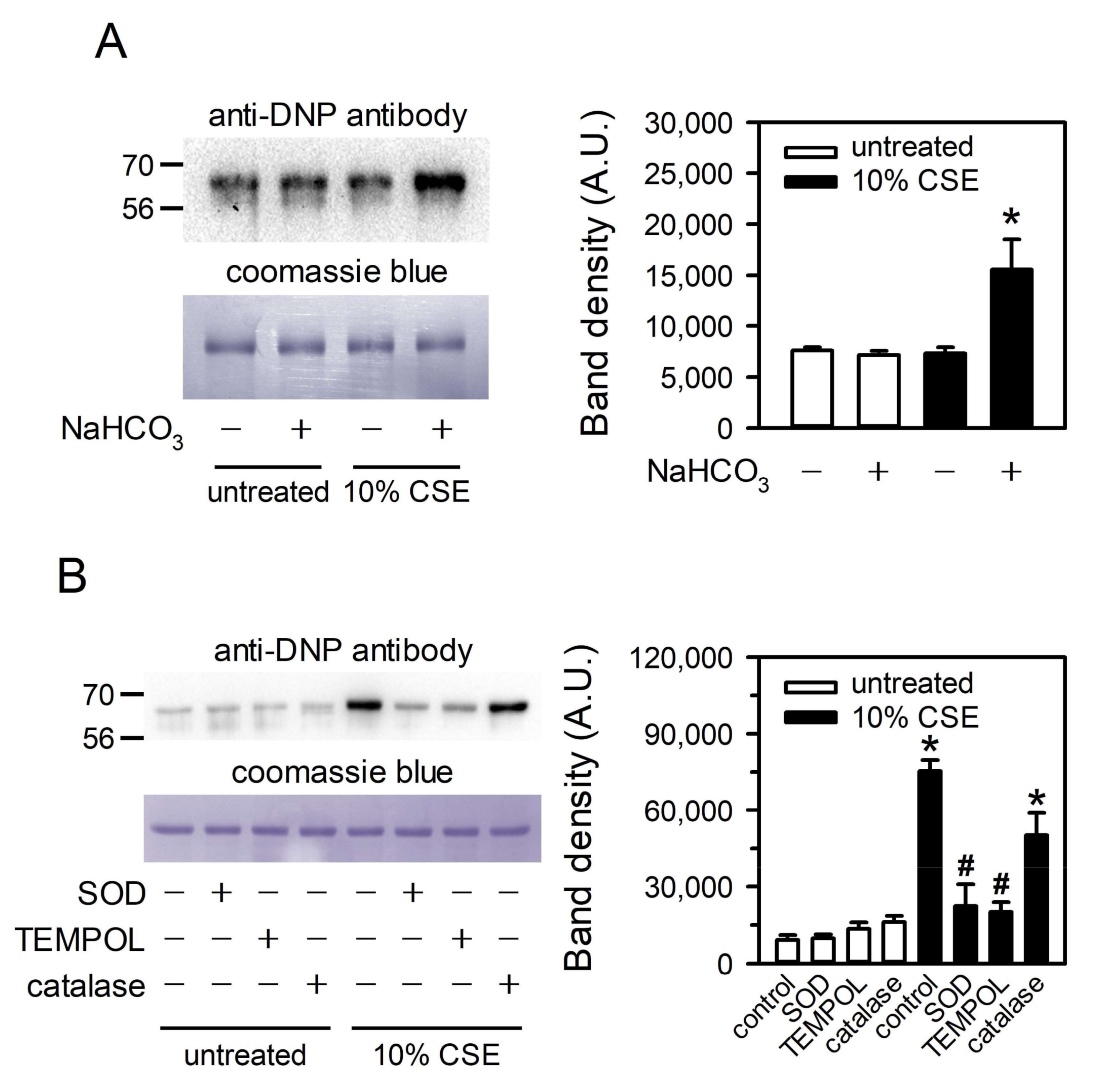
3.2. Bicarbonate in Aqueous Media Is Involved in CSE-Induced Superoxide Production
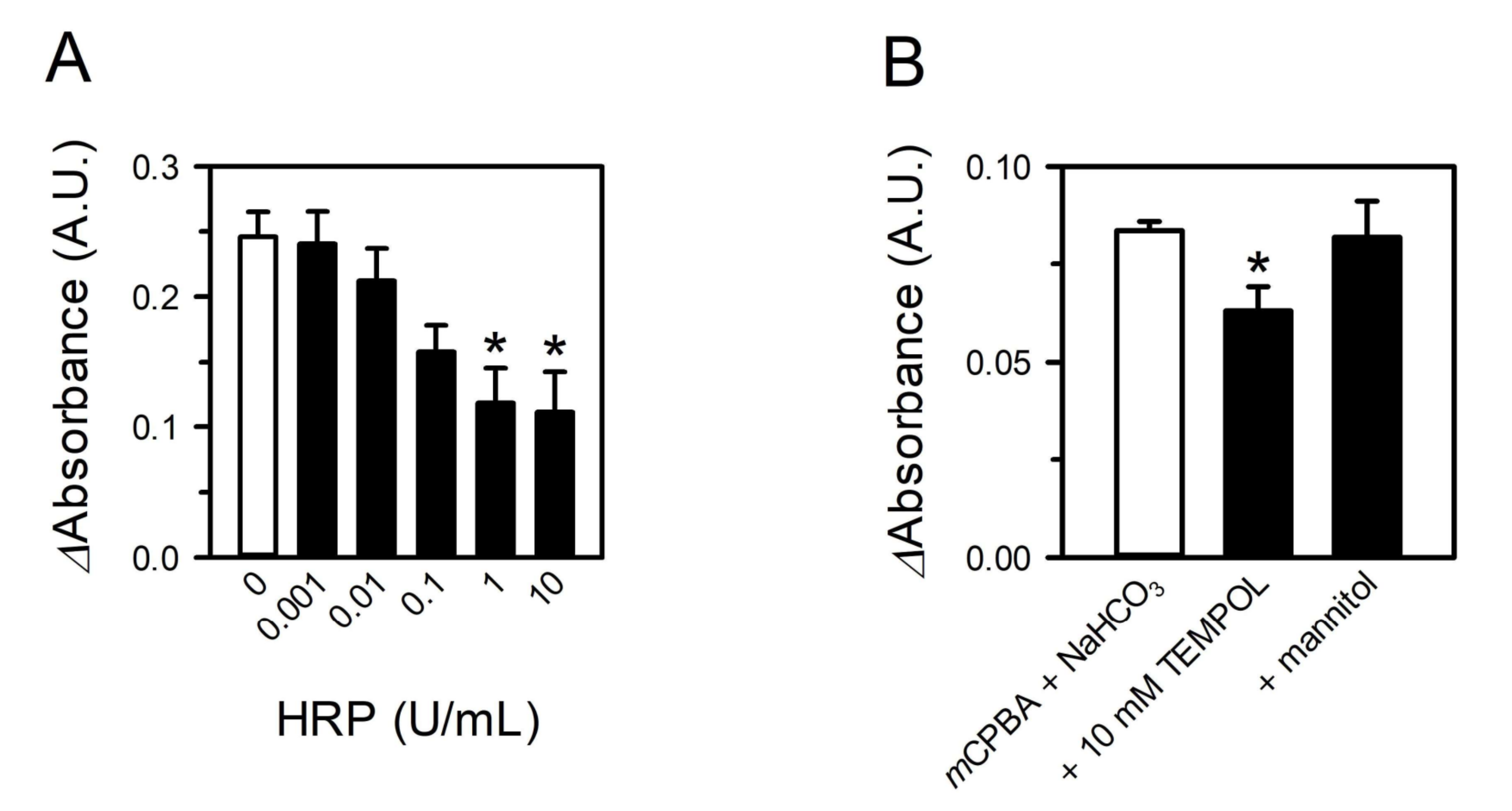
3.3. Peroxides in CSE Contribute to Superoxide Production by Reacting with Bicarbonate
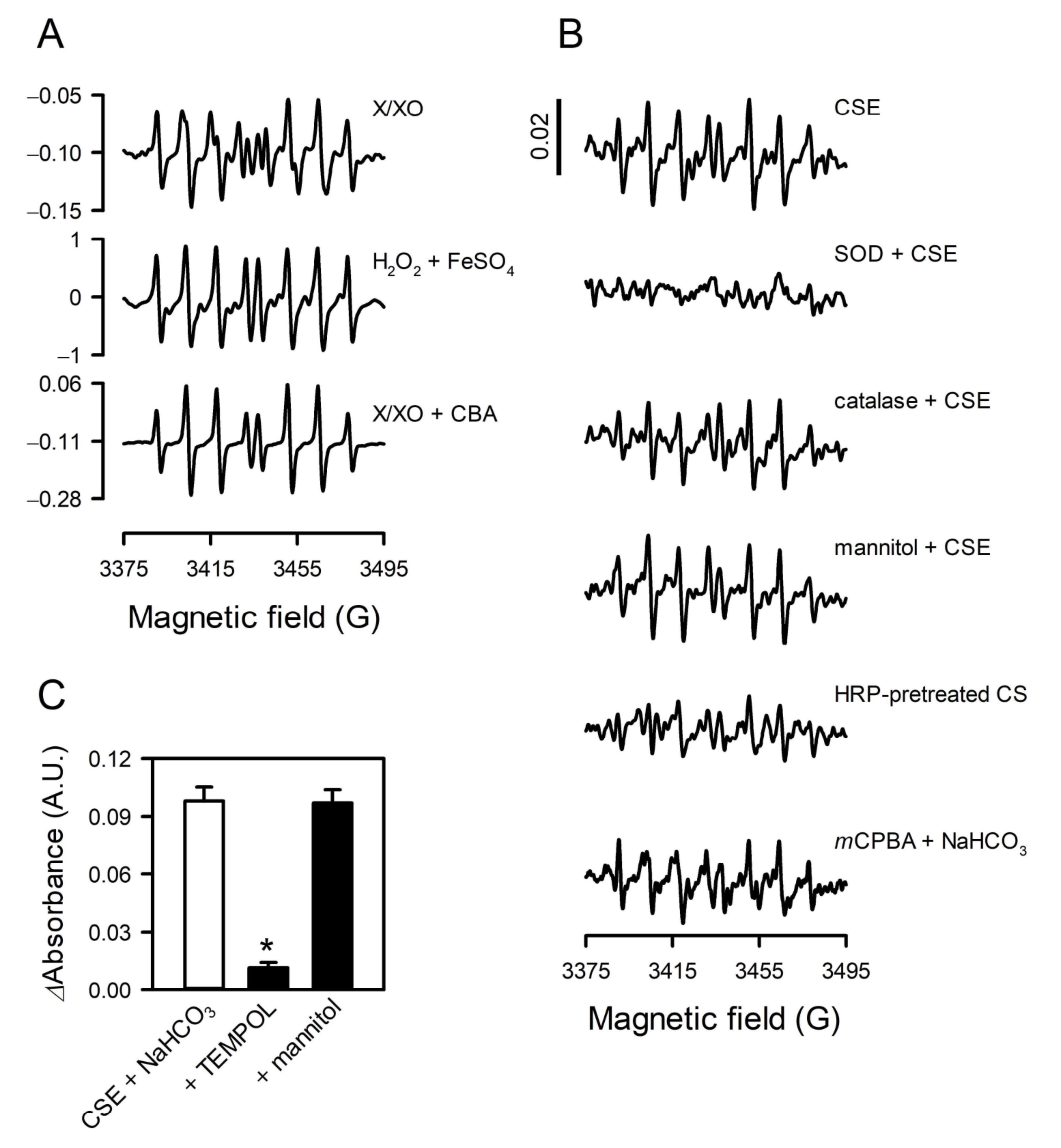
3.4. The Species of Reactive Oxygen Was Reconfirmed to Be Superoxide
3.5. TPM Also Generates Superoxide by Reacting with Bicarbonate
3.6. CSE-Generated Superoxide Is Capable of Inducing Oxidative Modification of Albumin
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- US Department of Health and Human Services. The Health Consequences of Smoking–50 Years of Progress: A Report of the Surgeon General; US Department of Health and Human Services: Washington, DC, USA, 2014.
- Wright, C. Standardized methods for the regulation of cigarette-smoke constituents. Trends Anal. Chem. 2015, 66, 118–127. [Google Scholar] [CrossRef] [Green Version]
- Ambrose, J.A.; Barua, R.S. The pathophysiology of cigarette smoking and cardiovascular disease: An update. J. Am. Coll. Cardiol. 2004, 43, 1731–1737. [Google Scholar] [CrossRef] [Green Version]
- Pryor, W.A.; Stone, K. Oxidants in cigarette smoke radicals, hydrogen oeroxide, peroxynitrate, and peroxynitritea. Ann. N. Y. Acad. Sci. 1993, 686, 12–27. [Google Scholar] [CrossRef]
- Pryor, W.A.; Hales, B.J.; Premovic, P.I.; Church, D.F. The radicals in cigarette tar: Their nature and suggested physiological implications. Science 1983, 220, 425–427. [Google Scholar] [CrossRef] [PubMed]
- Fetterman, J.L.; Sammy, M.J.; Ballinger, S.W. Mitochondrial toxicity of tobacco smoke and air pollution. Toxicology 2017, 391, 18–33. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Han, C.H.; Lee, M.Y. NADPH oxidase and the cardiovascular toxicity associated with smoking. Toxicol. Res. 2014, 30, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.H.; Park, J.M.; Lee, C.H.; Kim, B.; Choi, K.C.; Choi, S.J.; Lee, K.; Lee, M.Y. NADPH oxidase (NOX) 1 mediates cigarette smoke-induced superoxide generation in rat vascular smooth muscle cells. Toxicol. In Vitro 2017, 38, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Church, D.F.; Pryor, W.A. Free-radical chemistry of cigarette smoke and its toxicological implications. Environ. Health Perspect. 1985, 64, 111–126. [Google Scholar] [CrossRef] [PubMed]
- Kalyanaraman, B.; Perez-Reyes, E.; Mason, R.P. Spin-trapping and direct electron spin resonance investigations of the redox metabolism of quinone anticancer drugs. Biochim. Biophys. Acta Gen. Subj. 1980, 630, 119–130. [Google Scholar] [CrossRef]
- Tanigawa, T.; Yoshikawa, T.; Takahashi, S.; Naito, Y.; Kondo, M. Spin trapping of superoxide in aqueous solutions of fresh and aged cigarette smoke. Free Radic. Biol. Med. 1994, 17, 361–365. [Google Scholar] [CrossRef]
- Takanami, Y.; Nakayama, T. Evaluation of superoxide anion radicals generated from an aqueous extract of particulate phase cigarette smoke by electron spin resonance using 5,5-dimethyl-1-pyrroline-N-oxide. Biosci. Biotechnol. Biochem. 2011, 75, 34–39. [Google Scholar] [CrossRef] [Green Version]
- Jaccard, G.; Djoko, D.T.; Korneliou, A.; Stabbert, R.; Belushkin, M.; Esposito, M. Mainstream smoke constituents and in vitro toxicity comparative analysis of 3R4F and 1R6F reference cigarettes. Toxicol. Rep. 2019, 6, 222–231. [Google Scholar] [CrossRef] [PubMed]
- Kunzi, L.; Holt, G.E. Cigarette smoke activates the parthanatos pathway of cell death in human bronchial epithelial cells. Cell Death Discov. 2019, 5, 127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, C.W.; Go, R.E.; Lee, H.M.; Hwang, K.A.; Lee, K.; Kim, B.; Lee, M.Y.; Choi, K.C. Cigarette smoke extracts induced the colon cancer migration via regulating epithelial mesenchymal transition and metastatic genes in human colon cancer cells. Environ. Toxicol. 2017, 32, 690–704. [Google Scholar] [CrossRef] [PubMed]
- Yu, K.N.; Kim, H.J.; Kim, S.; Dawaadamdin, O.; Lee, A.Y.; Hong, S.H.; Chang, S.H.; Choi, S.J.; Shim, S.M.; Lee, K.; et al. Cigarette smoking condensate disrupts endoplasmic reticulum-golgi network homeostasis through GOLPH3 expression in normal lung epithelial cells. Nicotine Tob. Res. 2016, 18, 1877–1885. [Google Scholar] [CrossRef] [PubMed]
- Zielonka, J.; Cheng, G.; Zielonka, M.; Ganesh, T.; Sun, A.; Joseph, J.; Michalski, R.; O’Brien, W.J.; Lambeth, J.D.; Kalyanaraman, B. High-throughput assays for superoxide and hydrogen peroxide: Design of a screening workflow to identify inhibitors of NADPH oxidases. J. Biol. Chem. 2014, 289, 16176–16189. [Google Scholar] [CrossRef] [Green Version]
- Maghzal, G.J.; Krause, K.H.; Stocker, R.; Jaquet, V. Detection of reactive oxygen species derived from the family of NOX NADPH oxidases. Free Radic. Biol. Med. 2012, 53, 1903–1918. [Google Scholar] [CrossRef]
- Chen, H.; Lin, L.; Lin, Z.; Guo, G.; Lin, J.M. Chemiluminescence arising from the decomposition of peroxymonocarbonate and enhanced by CdTe quantum dots. J. Phys. Chem. A 2010, 114, 10049–10058. [Google Scholar] [CrossRef]
- Zhao, S.; Xi, H.; Zuo, Y.; Wang, Q.; Wang, Z.; Yan, Z. Bicarbonate-activated hydrogen peroxide and efficient decontamination of toxic sulfur mustard and nerve gas simulants. J. Hazard. Mater. 2018, 344, 136–145. [Google Scholar] [CrossRef]
- Dikalov, S.; Jiang, J.; Mason, R.P. Characterization of the high-resolution ESR spectra of superoxide radical adducts of 5-(diethoxyphosphoryl)-5-methyl-1-pyrroline N-oxide (DEPMPO) and 5,5-dimethyl-1-pyrroline N-oxide (DMPO). Analysis of conformational exchange. Free Radic. Res. 2005, 39, 825–836. [Google Scholar] [CrossRef]
- Roubaud, V.; Sankarapandi, S.; Kuppusamy, P.; Tordo, P.; Zweier, J.L. Quantitative measurement of superoxide generation using the spin trap 5-(diethoxyphosphoryl)-5-methyl-1-pyrroline-N-oxide. Anal. Biochem. 1997, 247, 404–411. [Google Scholar] [CrossRef] [PubMed]
- Sikora, A.; Zielonka, J.; Lopez, M.; Dybala-Defratyka, A.; Joseph, J.; Marcinek, A.; Kalyanaraman, B. Reaction between peroxynitrite and boronates: EPR spin-trapping, HPLC analyses, and quantum mechanical study of the free radical pathway. Chem. Res. Toxicol. 2011, 24, 687–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hogg, N.; Zielonka, J.; Kalyanaraman, B. Chapter 3—Detection of nitric oxide and peroxynitrite in biological systems: A state-of-the-art review. In Nitric Oxide, 3rd ed.; Ignarro, L.J., Freeman, B.A., Eds.; Academic Press: Boston, MA, USA, 2017; pp. 23–44. [Google Scholar]
- Johnson, M.D.; Schilz, J.; Djordjevic, M.V.; Rice, J.R.; Shields, P.G. Evaluation of in vitro assays for assessing the toxicity of cigarette smoke and smokeless tobacco. Cancer Epidemiol. Biomark. Prev. 2009, 18, 3263–3304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medinas, D.B.; Cerchiaro, G.; Trindade, D.F.; Augusto, O. The carbonate radical and related oxidants derived from bicarbonate buffer. IUBMB Life 2007, 59, 255–262. [Google Scholar] [CrossRef]
- Sabatini, S.; Kurtzman, N.A. Bicarbonate therapy in severe metabolic acidosis. J. Am. Soc. Nephrol. 2009, 20, 692–695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quintanilla, A.P. Acute acid-base disorders. Postgrad. Med. 1976, 60, 75–83. [Google Scholar] [CrossRef]
- Kamel, K.S.; Halperin, M.L. Chapter 8—Respiratory acid–base disturbances. In Fluid, Electrolyte and Acid-Base Physiology, 5th ed.; Kamel, K.S., Halperin, M.L., Eds.; Elsevier: Philadelphia, PA, USA, 2017; pp. 199–211. [Google Scholar]
- Effros, R.M.; Chinard, F.P. The in vivo pH of the extravascular space of the lung. J. Clin. Investig. 1969, 48, 1983–1996. [Google Scholar] [CrossRef] [PubMed]
- Bridges, R.J. Mechanisms of bicarbonate secretion: Lessons from the airways. Cold Spring Harb. Perspect. Med. 2012, 2, a015016. [Google Scholar] [CrossRef] [Green Version]
- Engelking, L.R. Chapter 90—Respiratory acidosis. In Textbook of Veterinary Physiological Chemistry, 3rd ed.; Engelking, L.R., Ed.; Academic Press: Boston, MA, USA, 2015; pp. 584–589. [Google Scholar]
- Rialp, G.; Raurich, J.M.; Llompart-Pou, J.A.; Ayestaran, I.; Ibanez, J. Respiratory CO2 response depends on plasma bicarbonate concentration in mechanically ventilated patients. Med. Intensiv. 2014, 38, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Kreindler, J.L.; Bertrand, C.A.; Lee, R.J.; Karasic, T.; Aujla, S.; Pilewski, J.M.; Frizzell, R.A.; Kolls, J.K. Interleukin-17A induces bicarbonate secretion in normal human bronchial epithelial cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 2009, 296, L257–L266. [Google Scholar] [CrossRef] [Green Version]
- Xiong, J.; Tian, J.; Zhou, L.; Le, Y.; Sun, Y. Interleukin-17A deficiency attenuated emphysema and bone loss in mice exposed to cigarette smoke. Int. J. Chron. Obstruct. Pulmon. Dis. 2020, 15, 301–310. [Google Scholar] [CrossRef] [Green Version]
- Cosgrove, J.P.; Borish, E.T.; Church, D.F.; Pryor, W.A. The metal-mediated formation of hydroxyl radical by aqueous extracts of cigarette tar. Biochem. Biophys. Res. Commun. 1985, 132, 390–396. [Google Scholar] [CrossRef]
- Nakayama, T.; Kodama, M.; Nagata, C. Generation of hydrogen peroxide and superoxide anion radical from cigarette smoke. Gann 1984, 75, 95–98. [Google Scholar]
- Besson, E.; Gastaldi, S.; Bloch, E.; Zielonka, J.; Zielonka, M.; Kalyanaraman, B.; Aslan, S.; Karoui, H.; Rockenbauer, A.; Ouari, O.; et al. Embedding cyclic nitrone in mesoporous silica particles for EPR spin trapping of superoxide and other radicals. Analyst 2019, 144, 4194–4203. [Google Scholar] [CrossRef]
- Moreno, J.J.; Foroozesh, M.; Church, D.F.; Pryor, W.A. Release of iron from ferritin by aqueous extracts of cigarette smoke. Chem. Res. Toxicol. 1992, 5, 116–123. [Google Scholar] [CrossRef]
- Wesselius, L.J.; Nelson, M.E.; Skikne, B.S. Increased release of ferritin and iron by iron-loaded alveolar macrophages in cigarette smokers. Am. J. Respir. Crit. Care Med. 1994, 150, 690–695. [Google Scholar] [CrossRef]
- Brun-Pascaud, M.; Gaudebout, C.; Blayo, M.C.; Pocidalo, J.J. Arterial blood gases and acid-base status in awake rats. Respir. Physiol. 1982, 48, 45–57. [Google Scholar] [CrossRef]
- Stocker, R. Antioxidant defenses in human blood plasma and extra-cellular fluids. Arch. Biochem. Biophys. 2016, 595, 136–139. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, J.-M.; Jeong, H.; Seo, Y.-S.; Do, V.Q.; Choi, S.-J.; Lee, K.; Choi, K.-C.; Choi, W.J.; Lee, M.-Y. Cigarette Smoke Extract Produces Superoxide in Aqueous Media by Reacting with Bicarbonate. Toxics 2021, 9, 316. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics9110316
Park J-M, Jeong H, Seo Y-S, Do VQ, Choi S-J, Lee K, Choi K-C, Choi WJ, Lee M-Y. Cigarette Smoke Extract Produces Superoxide in Aqueous Media by Reacting with Bicarbonate. Toxics. 2021; 9(11):316. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics9110316
Chicago/Turabian StylePark, Jung-Min, Haerin Jeong, Yoon-Seok Seo, Van Quan Do, Seong-Jin Choi, Kyuhong Lee, Kyung-Chul Choi, Won Jun Choi, and Moo-Yeol Lee. 2021. "Cigarette Smoke Extract Produces Superoxide in Aqueous Media by Reacting with Bicarbonate" Toxics 9, no. 11: 316. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics9110316