
Early Pregnancy Exposure to Ambient Air Pollution among Late-Onset Preeclamptic Cases Is Associated with Placental DNA Hypomethylation of Specific Genes and Slower Placental Maturation


 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
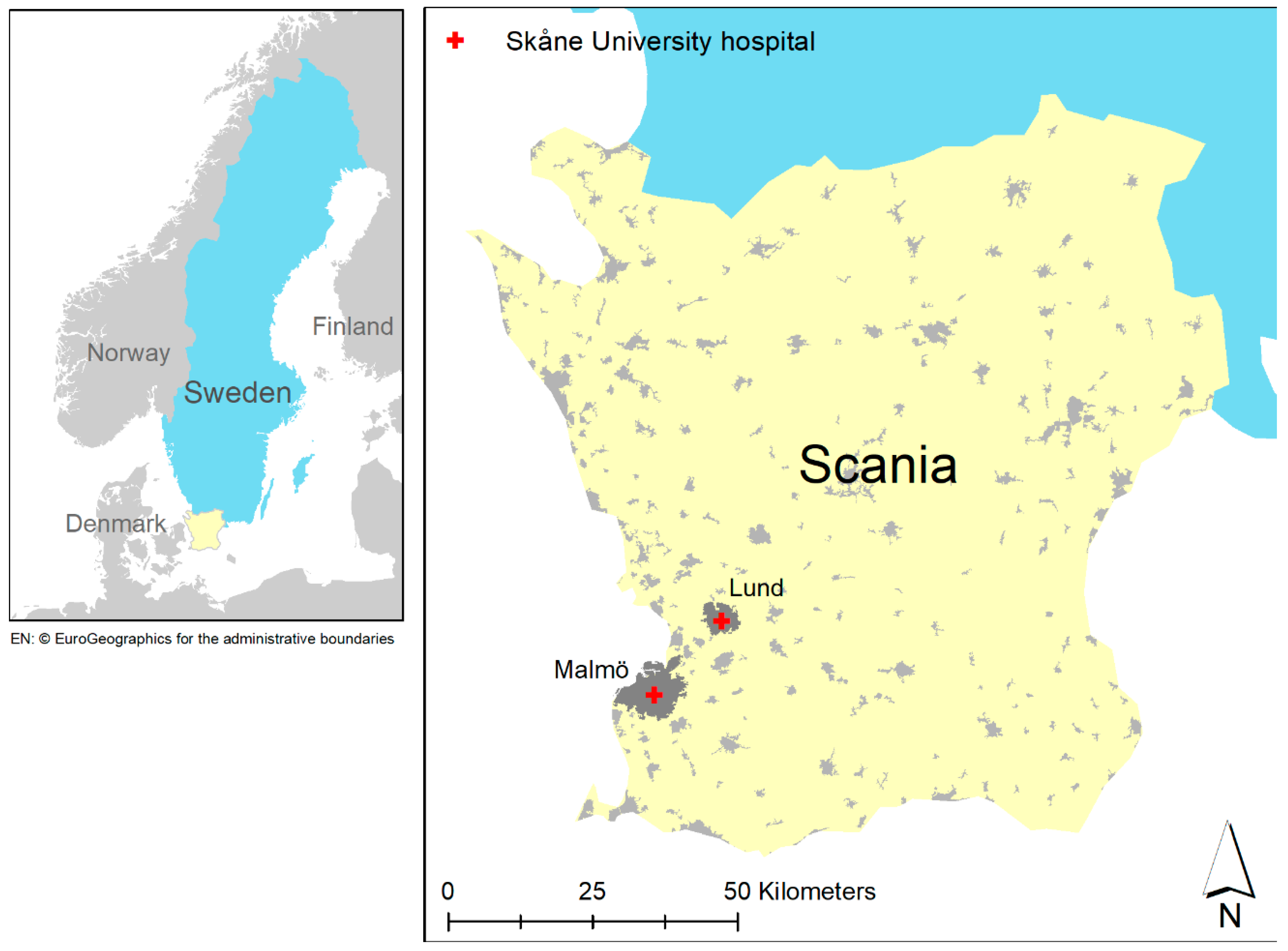
2.1. Study Population
2.2. Air Pollution Exposure Assessment
2.3. Study Design
2.4. Association between NOx Exposure and PE
2.5. Biological Sample Collection
2.6. DNA and RNA Extraction
2.7. Placental DNA Methylation Laboratory Analyses
2.8. Placental DNA Methylation Preprocessing
2.9. Predicted Gestational Age Acceleration/Deceleration and Associations with PE and NOx Exposure
2.10. DNA Methylation Preprocessing and Data Analysis
2.11. In Silico Analyses: Interaction Network Analyses and Gene Ontology Analyses for DNA Methylation
2.12. RNA Sequencing Analysis
3. Results
3.1. Characteristics of the Study Participants
3.2. Association between NOx Exposure and PE Status
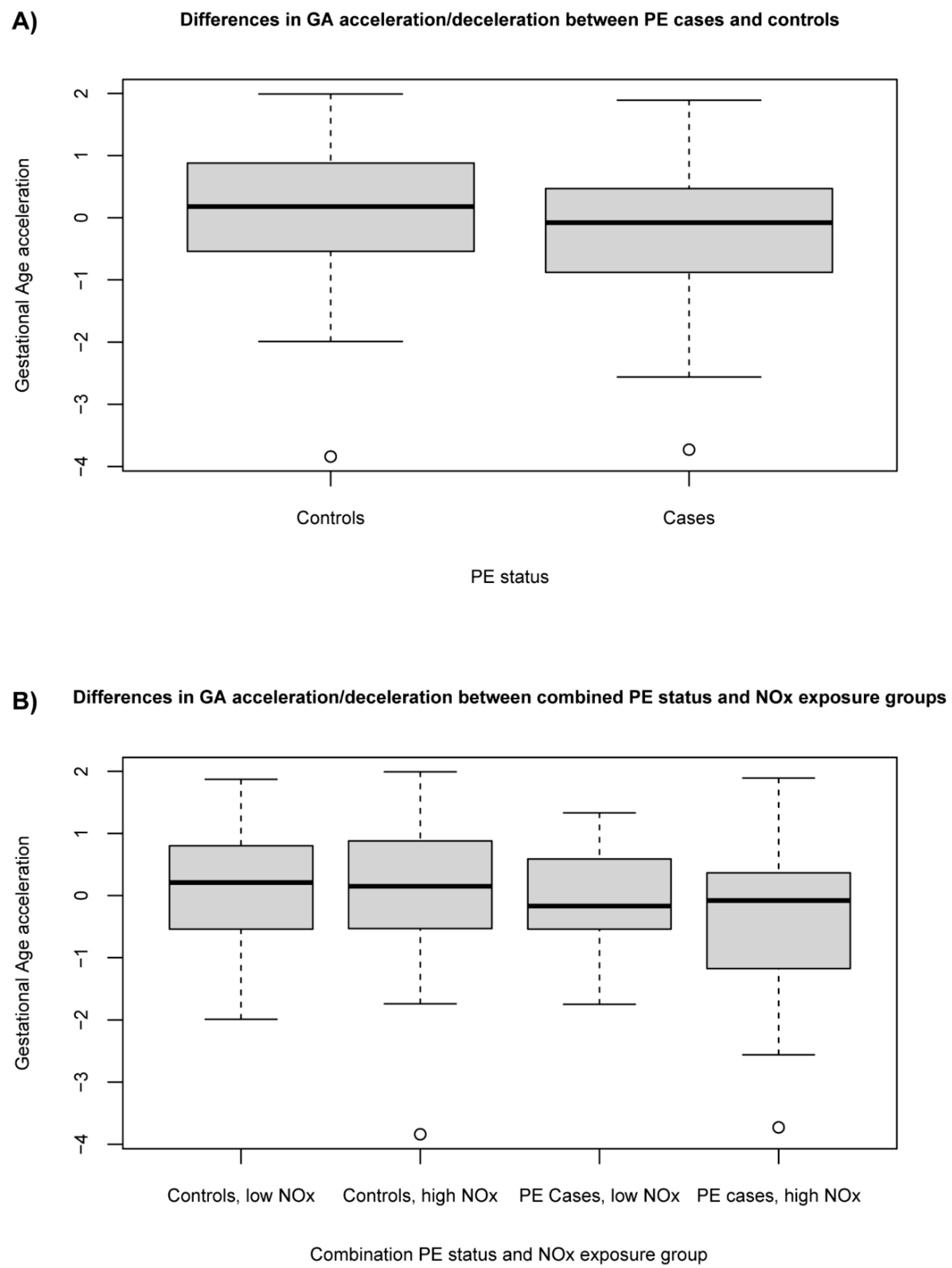
3.3. Association between Exposure to Ambient NOx and PE Status with Gestational Age Acceleration/Deceleration
3.4. Differentially Methylated Positions Associated with PE Status and NOx Exposure
3.5. In Silico Analyses: Interaction Network Analyses and Gene Ontology Analyses for DNA Methylation
3.6. RNA Sequencing: Analysis of Differentially Expressed Genes and Gene Ontology
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Khan, N.; Andrade, W.; De Castro, H.; Wright, A.; Wright, D.; Nicolaides, K.H. Impact of New Definitions of Pre-Eclampsia on Incidence and Performance of First-Trimester Screening. Ultrasound Obstet. Gynecol. 2020, 55, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Umesawa, M.; Kobashi, G. Epidemiology of Hypertensive Disorders in Pregnancy: Prevalence, Risk Factors, Predictors and Prognosis. Hypertens. Res. 2017, 40, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Thornton, C.; Dahlen, H.; Korda, A.; Hennessy, A. The Incidence of Preeclampsia and Eclampsia and Associated Maternal Mortality in Australia from Population-Linked Datasets: 2000–2008. Am. J. Obstet. Gynecol. 2013, 208, 476.e1–476.e5. [Google Scholar] [CrossRef]
- MacKay, A.P.; Berg, C.J.; Atrash, H.K. Pregnancy-Related Mortality from Preeclampsia and Eclampsia. Obstet. Gynecol. 2001, 97, 533–538. [Google Scholar] [CrossRef] [PubMed]
- Burton, G.J.; Fowden, A.L.; Thornburg, K.L. Placental Origins of Chronic Disease. Physiol. Rev. 2016, 96, 1509–1565. [Google Scholar] [CrossRef]
- Davis, E.F.; Lazdam, M.; Lewandowski, A.J.; Worton, S.A.; Kelly, B.; Kenworthy, Y.; Adwani, S.; Wilkinson, A.R.; McCormick, K.; Sargent, I.; et al. Cardiovascular Risk Factors in Children and Young Adults Born to Preeclamptic Pregnancies: A Systematic Review. Pediatrics 2012, 129, e1552–e1561. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.A.; Magee, L.A.; Kenny, L.C.; Karumanchi, S.A.; McCarthy, F.P.; Saito, S.; Hall, D.R.; Warren, C.E.; Adoyi, G.; Ishaku, S. Hypertensive Disorders of Pregnancy. Hypertension 2018, 72, 24–43. [Google Scholar] [CrossRef] [Green Version]
- Redman, C.W.G.; Staff, A.C.; Roberts, J.M. Syncytiotrophoblast Stress in Preeclampsia: The Convergence Point for Multiple Pathways. Am. J. Obstet. Gynecol. 2021, S0002–9378, 31115-7. [Google Scholar] [CrossRef] [PubMed]
- Raymond, D.; Peterson, E. A Critical Review of Early-Onset and Late-Onset Preeclampsia. Obstet. Gynecol. Surv. 2011, 66, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Taylor, B.D.; Ness, R.B.; Klebanoff, M.A.; Tang, G.; Roberts, J.M.; Hougaard, D.M.; Skogstrand, K.; Haggerty, C.L. The Impact of Female Fetal Sex on Preeclampsia and the Maternal Immune Milieu. Pregnancy Hypertens. 2018, 12, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Elsmén, E.; Källén, K.; Maršál, K.; Hellström-Westas, L. Fetal Gender and Gestational-Age-Related Incidence of Pre-Eclampsia. Acta Obstet. Gynecol. Scand. 2006, 85, 1285–1291. [Google Scholar] [CrossRef] [PubMed]
- Gyselaers, W. Preeclampsia Is a Syndrome with a Cascade of Pathophysiologic Events. J. Clin. Med. 2020, 9, 2245. [Google Scholar] [CrossRef]
- Pedersen, M.; Stayner, L.; Slama, R.; Sorensen, M.; Figueras, F.; Nieuwenhuijsen, M.J.; Raaschou-Nielsen, O.; Dadvand, P. Ambient Air Pollution and Pregnancy-Induced Hypertensive Disorders: A Systematic Review and Meta-Analysis. Hypertension 2014, 64, 494–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malmqvist, E.; Jakobsson, K.; Tinnerberg, H.; Rignell-Hydbom, A.; Rylander, L. Gestational Diabetes and Preeclampsia in Association with Air Pollution at Levels below Current Air Quality Guidelines. Environ. Health Perspect. 2013, 121, 488–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nääv, Å.; Erlandsson, L.; Isaxon, C.; Åsander Frostner, E.; Ehinger, J.; Sporre, M.K.; Krais, A.M.; Strandberg, B.; Lundh, T.; Elmér, E.; et al. Urban PM2.5 Induces Cellular Toxicity, Hormone Dysregulation, Oxidative Damage, Inflammation, and Mitochondrial Interference in the HRT8 Trophoblast Cell Line. Front. Endocrinol. 2020, 11, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Familari, M.; Nääv, Å.; Erlandsson, L.; de Iongh, R.U.; Isaxon, C.; Strandberg, B.; Lundh, T.; Hansson, S.R.; Malmqvist, E. Exposure of Trophoblast Cells to Fine Particulate Matter Air Pollution Leads to Growth Inhibition, Inflammation and ER Stress. PLoS ONE 2019, 14, e0218799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Ambient Air Pollution: A Global Assessment of Exposure and Burden of Disease. Clean Air J. 2016, 26, 6. [Google Scholar] [CrossRef]
- Bai, W.; Li, Y.; Niu, Y.; Ding, Y.; Yu, X.; Zhu, B.; Duan, R.; Duan, H.; Kou, C.; Li, Y.; et al. Association between Ambient Air Pollution and Pregnancy Complications: A Systematic Review and Meta-Analysis of Cohort Studies. Environ. Res. 2020, 185, 109471. [Google Scholar] [CrossRef] [PubMed]
- Bianco-Miotto, T.; Mayne, B.T.; Buckberry, S.; Breen, J.; Rodriguez Lopez, C.M.; Roberts, C.T. Recent Progress towards Understanding the Role of DNA Methylation in Human Placental Development. Reproduction 2016, 152, R23–R30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collas, P.; Noer, A.; Timoskainen, S. Programming the Genome in Embryonic and Somatic Stem Cells. J. Cell. Mol. Med. 2007, 11, 602–620. [Google Scholar] [CrossRef] [Green Version]
- Jirtle, R.L.; Skinner, M.K. Environmental Epigenomics and Disease Susceptibility. Nat. Rev. Genetics 2007, 8, 253–262. [Google Scholar] [CrossRef]
- Saenen, N.D.; Martens, D.S.; Neven, K.Y.; Alfano, R.; Bové, H.; Janssen, B.G.; Roels, H.A.; Plusquin, M.; Vrijens, K.; Nawrot, T.S. Air Pollution-Induced Placental Alterations: An Interplay of Oxidative Stress, Epigenetics, and the Aging Phenotype? Clin. Epigenet. 2019, 11, 124. [Google Scholar] [CrossRef] [PubMed]
- Cirkovic, A.; Garovic, V.; Milin Lazovic, J.; Milicevic, O.; Savic, M.; Rajovic, N.; Aleksic, N.; Weissgerber, T.; Stefanovic, A.; Stanisavljevic, D.; et al. Systematic Review Supports the Role of DNA Methylation in the Pathophysiology of Preeclampsia: A Call for Analytical and Methodological Standardization. Biol. Sex Differ. 2020, 11, 36. [Google Scholar] [CrossRef]
- Neven, K.Y.; Saenen, N.D.; Tarantini, L.; Janssen, B.G.; Lefebvre, W.; Vanpoucke, C.; Bollati, V.; Nawrot, T.S. Placental Promoter Methylation of DNA Repair Genes and Prenatal Exposure to Particulate Air Pollution: An ENVIR ON AGE Cohort Study. Lancet Planet. Health 2018, 2, e174–e183. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. WHO Global Air Quality Guidelines: Particulate Matter (PM2.5 and PM10), Ozone, Nitrogen Dioxide, Sulfur Dioxide and Carbon Monoxideitle; World Health Organization: Geneva, Switzerland, 2021; ISBN 9789240034228. [Google Scholar]
- Abraham, E.; Rousseaux, S.; Agier, L.; Giorgis-Allemand, L.; Tost, J.; Galineau, J.; Hulin, A.; Siroux, V.; Vaiman, D.; Charles, M.-A.; et al. Pregnancy Exposure to Atmospheric Pollution and Meteorological Conditions and Placental DNA Methylation. Environ. Int. 2018, 118, 334–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, L.S.; Redman, C. The Role of Cellular Senescence in Ageing of the Placenta. Placenta 2017, 52, 139–145. [Google Scholar] [CrossRef] [Green Version]
- Horvath, S. DNA Methylation Age of Human Tissues and Cell Types. Genome Biol. 2013, 14, R115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.; Choufani, S.; Weksberg, R.; Wilson, S.L.; Yuan, V.; Burt, A.; Marsit, C.; Lu, A.T.; Ritz, B.; Bohlin, J.; et al. Placental Epigenetic Clocks: Estimating Gestational Age Using Placental DNA Methylation Levels. Aging 2019, 11, 4238–4253. [Google Scholar] [CrossRef]
- Opsis. 2006. Available online: http://Www.Opsis.Se/ (accessed on 5 November 2021).
- Mandakh, Y.; Rittner, R.; Flanagan, E.; Oudin, A.; Isaxon, C.; Familari, M.; Hansson, S.R.; Malmqvist, E. Maternal Exposure to Ambient Air Pollution and Risk of Preeclampsia: A Population-Based Cohort Study in Scania, Sweden. Int. J. Environ. Res. Public Health 2020, 17, 1744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iodice, S.; Hoxha, M.; Ferrari, L.; Carbone, I.F.; Anceschi, C.; Miragoli, M.; Pesatori, A.C.; Persico, N.; Bollati, V. Particulate Air Pollution, Blood Mitochondrial DNA Copy Number, and Telomere Length in Mothers in the First Trimester of Pregnancy: Effects on Fetal Growth. Oxidative Med. Cell. Longev. 2018, 2018, 5162905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martens, D.S.; Cox, B.; Janssen, B.G.; Clemente, D.B.P.; Gasparrini, A.; Vanpoucke, C.; Lefebvre, W.; Roels, H.A.; Plusquin, M.; Nawrot, T.S. Prenatal Air Pollution and Newborns’ Predisposition to Accelerated Biological Aging. JAMA Pediatrics 2017, 171, 1160. [Google Scholar] [CrossRef] [PubMed]
- Melody, S.M.; Wills, K.; Knibbs, L.D.; Ford, J.; Venn, A.; Johnston, F. Maternal Exposure to Ambient Air Pollution and Pregnancy Complications in Victoria, Australia. Int. J. Environ. Res. Public Health 2020, 17, 2572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, T.J.; Butcher, L.M.; Feber, A.; Teschendorff, A.E.; Chakravarthy, A.R.; Wojdacz, T.K.; Beck, S. ChAMP: 450 k Chip Analysis Methylation Pipeline. Bioinformatics 2014, 30, 428–430. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Morris, T.J.; Webster, A.P.; Yang, Z.; Beck, S.; Feber, A.; Teschendorff, A.E. ChAMP: Updated Methylation Analysis Pipeline for Illumina BeadChips. Bioinformatics 2017, 33, 3982–3984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fortin, J.-P.; Triche, T.J.; Hansen, K.D. Preprocessing, Normalization and Integration of the Illumina HumanMethylationEPIC Array. bioRxiv 2017, 15, 558–560. [Google Scholar] [CrossRef] [PubMed]
- Aryee, M.J.; Jaffe, A.E.; Corrada-Bravo, H.; Ladd-Acosta, C.; Feinberg, A.P.; Hansen, K.D.; Irizarry, R.A. Minfi: A Flexible and Comprehensive Bioconductor Package for the Analysis of Infinium DNA Methylation Microarrays. Bioinformatics 2014, 30, 1363–1369. [Google Scholar] [CrossRef] [Green Version]
- Teschendorff, A.E.; Marabita, F.; Lechner, M.; Bartlett, T.; Tegner, J.; Gomez-Cabrero, D.; Beck, S. A Beta-Mixture Quantile Normalization Method for Correcting Probe Design Bias in Illumina Infinium 450 k DNA Methylation Data. Bioinformatics 2013, 29, 189–196. [Google Scholar] [CrossRef] [Green Version]
- Yuan, V.; Hui, D.; Yin, Y.; Peñaherrera, M.S.; Beristain, A.G.; Robinson, W.P. Cell-Specific Characterization of the Placental Methylome. BMC Genom. 2021, 22, 6. [Google Scholar] [CrossRef]
- Khouja, J.N.; Simpkin, A.J.; O’Keeffe, L.M.; Wade, K.H.; Houtepen, L.C.; Relton, C.L.; Suderman, M.; Howe, L.D. Epigenetic Gestational Age Acceleration: A Prospective Cohort Study Investigating Associations with Familial, Sociodemographic and Birth Characteristics. Clin. Epigenet. 2018, 10, 86. [Google Scholar] [CrossRef]
- Johnson, W.E.; Li, C.; Rabinovic, A. Adjusting Batch Effects in Microarray Expression Data Using Empirical Bayes Methods. Biostatistics 2007, 8, 118–127. [Google Scholar] [CrossRef]
- Leek, J.T.; Johnson, W.E.; Parker, H.S.; Fertig, E.J.; Jaffe, A.E.; Storey, J.D.; Zhang, Y.; Torres, L.C. Sva: Surrogate Variable Analysis. R Package Version 3.24.4. 2017. Available online: https://bioconductor.org/packages/release/bioc/html/sva.html (accessed on 18 November 2021).
- Gude, N.M.; Roberts, C.T.; Kalionis, B.; King, R.G. Growth and Function of the Normal Human Placenta. Thromb. Res. 2004, 114, 397–407. [Google Scholar] [CrossRef]
- Reyes, L.; Golos, T.G. Hofbauer Cells: Their Role in Healthy and Complicated Pregnancy. Front. Immunol. 2018, 9, 2628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. Limma Powers Differential Expression Analyses for RNA-Sequencing and Microarray Studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef] [PubMed]
- Phipson, B.; Maksimovic, J.; Oshlack, A. MissMethyl: An R Package for Analyzing Data from Illumina’s HumanMethylation450 Platform. Bioinformatics 2016, 32, 286–288. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. 2010. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc (accessed on 19 November 2021).
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.-C.; Mendell, J.T.; Salzberg, S.L. StringTie Enables Improved Reconstruction of a Transcriptome from RNA-Seq Reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frazer, K.A.; Ballinger, D.G.; Cox, D.R.; Hinds, D.A.; Stuve, L.L.; Gibbs, R.A.; Belmont, J.W.; Boudreau, A.; Hardenbol, P.; Leal, S.M.; et al. A Second Generation Human Haplotype Map of over 3.1 Million SNPs. Nature 2007, 449, 851–861. [Google Scholar] [CrossRef] [PubMed]
- Altshuler, D.M.; Gibbs, R.A.; Peltonen, L.; Schaffner, S.F.; Yu, F.; Dermitzakis, E.; Bonnen, P.E.; De Bakker, P.I.W.; Deloukas, P.; Gabriel, S.B.; et al. Integrating Common and Rare Genetic Variation in Diverse Human Populations. Nature 2010, 467, 52–58. [Google Scholar] [CrossRef]
- Seidmann, L.; Suhan, T.; Unger, R.; Gerein, V.; Kirkpatrick, C.J. Transient CD15-Positive Endothelial Phenotype in the Human Placenta Correlates with Physiological and Pathological Fetoplacental Immaturity. Eur. J. Obstet. Gynecol. Reprod. Biol. 2014, 180, 172–179. [Google Scholar] [CrossRef] [PubMed]
- Costanzo, V.; Bardelli, A.; Siena, S.; Abrignani, S. Exploring the Links between Cancer and Placenta Development. Open Biol. 2018, 8, 180081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holtan, S.G.; Creedon, D.J.; Haluska, P.; Markovic, S.N. Cancer and Pregnancy: Parallels in Growth, Invasion, and Immune Modulation and Implications for Cancer Therapeutic Agents. Mayo Clin. Proc. 2009, 84, 985–1000. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, M.M.S.; Salgado, C.M.; Viana, L.R.; Gomes-Marcondes, M.C.C. Pregnancy and Cancer: Cellular Biology and Mechanisms Affecting the Placenta. Cancers 2021, 13, 1667. [Google Scholar] [CrossRef] [PubMed]
- Meinhardt, G.; Haider, S.; Kunihs, V.; Saleh, L.; Pollheimer, J.; Fiala, C.; Hetey, S.; Feher, Z.; Szilagyi, A.; Than, N.G.; et al. Pivotal Role of the Transcriptional Co-Activator YAP in Trophoblast Stemness of the Developing Human Placenta. Proc. Natl. Acad. Sci. USA 2020, 117, 13562–13570. [Google Scholar] [CrossRef] [PubMed]
- Zha, W.; Guan, S.; Liu, N.; Li, Y.; Tian, Y.; Chen, Y.; Wang, Y.; Wu, F. Let-7a Inhibits Bcl-Xl and YAP1 Expression to Induce Apoptosis of Trophoblast Cells in Early-Onset Severe Preeclampsia. Sci. Total Environ. 2020, 745, 139919. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Xu, P.; Luo, X.; Hu, M.; Liu, Y.; Yang, Y.; Peng, W.; Bai, Y.; Chen, X.; Tan, B.; et al. Phosphorylation of Yes-Associated Protein Impairs Trophoblast Invasion and Migration: Implications for the Pathogenesis of Fetal Growth Restriction†. Biol. Reprod. 2020, 103, 866–879. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Na, Q.; Huang, L.; Song, G.; Jin, F.; Li, Y.; Hou, Y.; Kang, D.; Qiao, C. YAP Is Decreased in Preeclampsia and Regulates Invasion and Apoptosis of HTR-8/SVneo. Reprod. Sci. 2018, 25, 1382–1393. [Google Scholar] [CrossRef]
- Liu, R.; Wei, C.; Ma, Q.; Wang, W. Hippo-YAP1 Signaling Pathway and Severe Preeclampsia (SPE) in the Chinese Population. Pregnancy Hypertens. 2020, 19, 1–10. [Google Scholar] [CrossRef]
- Turowski, G.; Vogel, M. Re-View and View on Maturation Disorders in the Placenta. Apmis 2018, 126, 602–612. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Cui, D.; Ren, J.; Wang, K.; Zeng, T.; Gao, L. CACNA2D3 Is Downregulated in Gliomas and Functions as a Tumor Suppressor. Mol. Carcinog. 2017, 56, 945–959. [Google Scholar] [CrossRef]
- Baczyk, D.; Kingdom, J.C.P.; Uhlén, P. Calcium Signaling in Placenta. Cell Calcium 2011, 49, 350–356. [Google Scholar] [CrossRef] [PubMed]
- Jakoubek, V.; Bíbová, J.; Hampl, V. Voltage-Gated Calcium Channels Mediate Hypoxic Vasoconstriction in the Human Placenta. Placenta 2006, 27, 1030–1033. [Google Scholar] [CrossRef]
- Hansson, S.R.; Chen, Y.; Brodszki, J.; Chen, M.; Hernandez-Andrade, E.; Inman, J.M.; Kozhich, O.A.; Larsson, I.; Marsal, K.; Medstrand, P.; et al. Gene Expression Profiling of Human Placentas from Preeclamptic and Normotensive Pregnancies. Mol. Hum. Reprod. 2006, 12, 169–179. [Google Scholar] [CrossRef] [PubMed]
- Li, H.Q.; Fan, J.J.; Li, X.H.; Bao, D. MiR-507 Inhibits the Growth and Invasion of Trophoblasts by Targeting CAMK4. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 5856–5862. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Xiang, Y.; Zhou, X.; Zheng, X.; Zhang, H.; Zhang, X.; Zhang, J.; He, L.; Zhao, X. Epigenome-Wide Association Data Implicate Fetal/Maternal Adaptations Contributing to Clinical Outcomes in Preeclampsia. Epigenomics 2019, 11, 1003–1019. [Google Scholar] [CrossRef] [Green Version]
- Schoentgen, F.; Jonic, S. PEBP1/RKIP Behavior: A Mirror of Actin-Membrane Organization. Cell. Mol. Life Sci. 2020, 77, 859–874. [Google Scholar] [CrossRef] [PubMed]
- Ciarmela, P.; Marzioni, D.; Islam, M.S.; Gray, P.C.; Terracciano, L.; Lorenzi, T.; Todros, T.; Petraglia, F.; Castellucci, M. Possible Role of RKIP in Cytotrophoblast Migration: Immunohistochemical and in Vitro Studies. J. Cell. Physiol. 2012, 227, 1821–1828. [Google Scholar] [CrossRef] [PubMed]
- Goswami, D.; Tannetta, D.S.; Magee, L.A.; Fuchisawa, A.; Redman, C.W.; Sargent, I.L.; von Dadelszen, P. Excess Syncytiotrophoblast Microparticle Shedding Is a Feature of Early-Onset Pre-Eclampsia, but Not Normotensive Intrauterine Growth Restriction. Placenta 2006, 27, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Frémont, S.; Hammich, H.; Bai, J.; Wioland, H.; Klinkert, K.; Rocancourt, M.; Kikuti, C.; Stroebel, D.; Romet-Lemonne, G.; Pylypenko, O.; et al. Oxidation of F-Actin Controls the Terminal Steps of Cytokinesis. Nat. Commun. 2017, 8, 14528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, W.; Wang, Y.; Gu, L.; Duan, B.; Cui, J.; Zhang, Y.; Chen, Y.; Sun, S.; Dong, J.; Du, J. MICAL1 Controls Cell Invasive Phenotype via Regulating Oxidative Stress in Breast Cancer Cells. BMC Cancer 2016, 16, 489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Li, Y. The Pseudogene URAHP Promotes Proliferation and Regulates the Pathogenesis of Preeclampsia. Am. J. Transl. Res. 2020, 12, 4715–4727. [Google Scholar] [PubMed]
- Gracilla, D.E.; Korla, P.K.; Lai, M.; Chiang, A.; Liou, W.; Sheu, J.J. Overexpression of Wild Type or a Q311E Mutant MB21D2 Promotes a Pro-oncogenic Phenotype in HNSCC. Mol. Oncol. 2020, 14, 3065–3082. [Google Scholar] [CrossRef] [PubMed]
- Fabriek, B.O.; Dijkstra, C.D.; van den Berg, T.K. The Macrophage Scavenger Receptor CD163. Immunobiology 2005, 210, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Joerink, M.; Rindsjo, E.; van Riel, B.; Alm, J.; Papadogiannakis, N. Placental Macrophage (Hofbauer Cell) Polarization Is Independent of Maternal Allergen-Sensitization and Presence of Chorioamnionitis. Placenta 2011, 32, 380–385. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Buhimschi, I.A.; Buhimschi, C.S.; Tadesse, S.; Norwitz, E.; Niven-Fairchild, T.; Huang, S.T.; Guller, S. Decreased Levels of Folate Receptor-Beta and Reduced Numbers of Fetal Macrophages (Hofbauer Cells) in Placentas from Pregnancies with Severe Pre-Eclampsia. Am. J. Reprod. Immunol. 2013, 70, 104–115. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.W.; Cho, E.H.; Choi, S.Y.; Lee, Y.K.; Park, J.H.; Kim, M.K.; Park, J.Y.; Choi, H.J.; Lee, J.I.; Ko, H.M.; et al. DC-SIGN Expression in Hofbauer Cells May Play an Important Role in Immune Tolerance in Fetal Chorionic Villi during the Development of Preeclampsia. J. Reprod. Immunol. 2017, 124, 30–37. [Google Scholar] [CrossRef]
- Munn, D.H.; Zhou, M.; Attwood, J.T.; Bondarev, I.; Conway, S.J.; Marshall, B.; Brown, C.; Mellor, A.L. Prevention of Allogeneic Fetal Rejection by Tryptophan Catabolism. Science 1998, 281, 1191–1193. [Google Scholar] [CrossRef]
- Broekhuizen, M.; Klein, T.; Hitzerd, E.; de Rijke, Y.B.; Schoenmakers, S.; Sedlmayr, P.; Danser, A.H.J.; Merkus, D.; Reiss, I.K.M. L-Tryptophan–Induced Vasodilation Is Enhanced in Preeclampsia. Hypertension 2020, 76, 184–194. [Google Scholar] [CrossRef]
- Keaton, S.A.; Heilman, P.; Bryleva, E.Y.; Madaj, Z.; Krzyzanowski, S.; Grit, J.; Miller, E.S.; Jalmby, M.; Kalapotharakos, G.; Racicot, K.; et al. Altered Tryptophan Catabolism in Placentas From Women With Pre-Eclampsia. Int. J. Tryptophan Res. 2019, 12, 1178646919840321. [Google Scholar] [CrossRef]
- Hanswijk, S.I.; Spoelder, M.; Shan, L.; Verheij, M.M.M.; Muilwijk, O.G.; Li, W.; Liu, C.; Kolk, S.M.; Homberg, J.R. Gestational Factors throughout Fetal Neurodevelopment: The Serotonin Link. Int. J. Mol. Sci. 2020, 21, 5850. [Google Scholar] [CrossRef]
- Yu, J.; Madison, J.M.; Mundlos, S.; Winston, F.; Olsen, B.R. Characterization of a Human Homologue of TheSaccharomyces CerevisiaeTranscription Factor Spt3 (SUPT3H). Genomics 1998, 53, 90–96. [Google Scholar] [CrossRef] [Green Version]
- Perez-Garcia, V.; Fineberg, E.; Wilson, R.; Murray, A.; Mazzeo, C.I.; Tudor, C.; Sienerth, A.; White, J.K.; Tuck, E.; Ryder, E.J.; et al. Placentation Defects Are Highly Prevalent in Embryonic Lethal Mouse Mutants. Nature 2018, 555, 463–468. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, S.L.; Mathew, P. Alpha2-Antiplasmin and Its Deficiency: Fibrinolysis out of Balance. Haemophilia 2008, 14, 1250–1254. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Okada, K.; Okamoto, C.; Ueshima, S.; Matsuo, O. Alpha2-Antiplasmin Is a Critical Regulator of Angiotensin II-Mediated Vascular Remodeling. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 1257–1262. [Google Scholar] [CrossRef] [Green Version]
- Shao, X.; Wang, Y.; Liu, Y.; Guo, X.; Li, D.; Huo, R.; Jia, W.; Cao, G.; Li, Y.-X.; Liu, M.; et al. Association of Imbalanced Sex Hormone Production with Excessive Procoagulation Factor SerpinF2 in Preeclampsia. J. Hypertens. 2019, 37, 197–205. [Google Scholar] [CrossRef]
- Jin, G.; Ma, H.; Wu, C.; Dai, J.; Zhang, R.; Shi, Y.; Lu, J.; Miao, X.; Wang, M.; Zhou, Y.; et al. Genetic Variants at 6p21.1 and 7p15.3 Are Associated with Risk of Multiple Cancers in Han Chinese. Am. J. Hum. Genet. 2012, 91, 928–934. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.; Liu, K.; Huang, J.; Sun, Q.; Shao, C.; Luo, J.; Xu, L.; Shen, Y.; Ren, B. FAM111B, a Direct Target of P53, Promotes the Malignant Process of Lung Adenocarcinoma. OncoTargets Ther. 2019, 12, 2829–2842. [Google Scholar] [CrossRef] [Green Version]
- Kawasaki, K.; Nojima, S.; Hijiki, S.; Tahara, S.; Ohshima, K.; Matsui, T.; Hori, Y.; Kurashige, M.; Umeda, D.; Kiyokawa, H.; et al. FAM111B Enhances Proliferation of KRAS-Driven Lung Adenocarcinoma by Degrading P16. Cancer Sci. 2020, 111, 2635–2646. [Google Scholar] [CrossRef]
- Liu, C.; Cummins, T.R.; Tyrrell, L.; Black, J.A.; Waxman, S.G.; Dib-Hajj, S.D. CAP-1A Is a Novel Linker That Binds Clathrin and the Voltage-Gated Sodium Channel Nav1.8. Mol. Cell. Neurosci. 2005, 28, 636–649. [Google Scholar] [CrossRef] [PubMed]
- Cronqvist, T.; Erlandsson, L.; Tannetta, D.; Hansson, S.R. Placental Syncytiotrophoblast Extracellular Vesicles Enter Primary Endothelial Cells through Clathrin-Mediated Endocytosis. Placenta 2020, 100, 133–141. [Google Scholar] [CrossRef]
- Fisher, S.J. Why Is Placentation Abnormal in Preeclampsia? Am. J. Obstet. Gynecol. 2015, 213, S115–S122. [Google Scholar] [CrossRef] [Green Version]
- Isaevska, E.; Moccia, C.; Asta, F.; Cibella, F.; Gagliardi, L.; Ronfani, L.; Rusconi, F.; Stazi, M.A.; Richiardi, L. Exposure to Ambient Air Pollution in the First 1000 Days of Life and Alterations in the DNA Methylome and Telomere Length in Children: A Systematic Review. Environ. Res. 2021, 193, 110504. [Google Scholar] [CrossRef]
- Uhlen, M.; Fagerberg, L.; Hallstrom, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, A.; Kampf, C.; Sjostedt, E.; Asplund, A.; et al. Proteomics. Tissue-Based Map of the Human Proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef]
- Stelzer, G.; Rosen, N.; Plaschkes, I.; Zimmerman, S.; Twik, M.; Fishilevich, S.; Stein, T.I.; Nudel, R.; Lieder, I.; Mazor, Y.; et al. The GeneCards Suite: From Gene Data Mining to Disease Genome Sequence Analyses. Curr. Protoc. Bioinform. 2016, 54, 1.30.1–1.30.33. [Google Scholar] [CrossRef]
- Detchokul, S.; Williams, E.D.; Parker, M.W.; Frauman, A.G. Tetraspanins as Regulators of the Tumour Microenvironment: Implications for Metastasis and Therapeutic Strategies. Br. J. Pharmacol. 2014, 171, 5462–5490. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Zucker, B.; Zhang, S.; Elias, S.; Zhu, Y.; Chen, H.; Ding, T.; Li, Y.; Sun, Y.; Lou, J.; et al. Migrasome Formation Is Mediated by Assembly of Micron-Scale Tetraspanin Macrodomains. Nat. Cell Biol. 2019, 21, 991–1002. [Google Scholar] [CrossRef]
- Thomas, J.R.; Appios, A.; Zhao, X.; Dutkiewicz, R.; Donde, M.; Lee, C.Y.C.; Naidu, P.; Lee, C.; Cerveira, J.; Liu, B.; et al. Phenotypic and Functional Characterization of First-Trimester Human Placental Macrophages, Hofbauer Cells. J. Exp. Med. 2021, 218, e20200891. [Google Scholar] [CrossRef]
- Shindo-Okada, N.; Iigo, M. Expression of the Arp11 Gene Suppresses the Tumorigenicity of PC-14 Human Lung Adenocarcinoma Cells. Biochem. Biophys. Res. Commun. 2003, 312, 889–896. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yao, X.; Fischer, L.; Abenza, J.F.; Penalva, M.A.; Xiang, X. The P25 Subunit of the Dynactin Complex Is Required for Dynein-Early Endosome Interaction. J. Cell Biol. 2011, 193, 1245–1255. [Google Scholar] [CrossRef] [Green Version]
- Henrichs, V.; Grycova, L.; Barinka, C.; Nahacka, Z.; Neuzil, J.; Diez, S.; Rohlena, J.; Braun, M.; Lansky, Z. Mitochondria-Adaptor TRAK1 Promotes Kinesin-1 Driven Transport in Crowded Environments. Nat. Commun. 2020, 11, 3123. [Google Scholar] [CrossRef] [PubMed]
- Fenton, A.R.; Jongens, T.A.; Holzbaur, E.L.F. Mitochondrial Adaptor TRAK2 Activates and Functionally Links Opposing Kinesin and Dynein Motors. Nat. Commun. 2021, 12, 4578. [Google Scholar] [CrossRef] [PubMed]
- Sprecher, E.; Itin, P.; Whittock, N.V.; McGrath, J.A.; Meyer, R.; DiGiovanna, J.J.; Bale, S.J.; Uitto, J.; Richard, G. Refined Mapping of Naegeli-Franceschetti-Jadassohn Syndrome to a 6 CM Interval on Chromosome 17q11.2-Q21 and Investigation of Candidate Genes. J. Investig. Dermatol. 2002, 119, 692–698. [Google Scholar] [CrossRef]
- Han, H.J.; Sudo, K.; Inazawa, J.; Nakamura, Y. Isolation and Mapping of a Human Gene (RABL) Encoding a Small GTP-Binding Protein Homologous to the Ras-Related RAB Gene. Cytogenet. Cell Genet. 1996, 73, 137–139. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.I.; Nam, I.K.; Lee, D.K.; Bhandari, S.; Charton, L.; Kwak, S.; Lim, J.Y.; Hong, K.; Kim, S.J.; Lee, J.N.; et al. Slc25a17 Acts as a Peroxisomal Coenzyme A Transporter and Regulates Multiorgan Development in Zebrafish. J. Cell. Physiol. 2020, 235, 151–165. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Lv, X.; Huang, C.; Hua, G.; Ma, B.; Chen, X.; Angeletti, P.C.; Dong, J.; Zhou, J.; Wang, Z.; et al. YAP1-LATS2 Feedback Loop Dictates Senescent or Malignant Cell Fate to Maintain Tissue Homeostasis. EMBO Rep. 2019, 20, e44948. [Google Scholar] [CrossRef]
- Wills, Z.P.; Mandel-Brehm, C.; Mardinly, A.R.; McCord, A.E.; Giger, R.J.; Greenberg, M.E. The Nogo Receptor Family Restricts Synapse Number in the Developing Hippocampus. Neuron 2012, 73, 466–481. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.F.; Olsson, P.; Wolfgang, C.D.; Bera, T.K.; Duray, P.; Lee, B.; Pastan, I. PRAC: A Novel Small Nuclear Protein That Is Specifically Expressed in Human Prostate and Colon. Prostate 2001, 47, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Yang, X.; Zhang, L.; Duan, Z.; Jia, L.; Wang, P.; Shi, Y.; Li, Y.; Gao, J. Decreased Expression and Activation of Stat3 in Severe Preeclampsia. J. Mol. Histol. 2015, 46, 205–219. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, X.; Wang, J.; Zhang, L. The Decreased Expression of Stat3 and P-Stat3 in Preeclampsia-like Rat Placenta. J. Mol. Histol. 2018, 49, 175–183. [Google Scholar] [CrossRef]
- Zhang, X.L.; Pan, J.; Wang, G.; Li, H.; Wang, J.L.; Jiang, X.M.; Zhao, X.Y. Effect of STAT3 Mediated Epigenetic Regulation in Pre-Eclampsia: An Analysis of Trial Data. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 5843–5855. [Google Scholar] [CrossRef]
- Kalisch-Smith, J.I.; Simmons, D.G.; Dickinson, H.; Moritz, K.M. Review: Sexual Dimorphism in the Formation, Function and Adaptation of the Placenta. Placenta 2017, 54, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Kalisch-Smith, J.I.; Simmons, D.G.; Pantaleon, M.; Moritz, K.M. Sex Differences in Rat Placental Development: From Pre-Implantation to Late Gestation. Biol. Sex Differ. 2017, 8, 17. [Google Scholar] [CrossRef] [Green Version]
- Rosenfeld, C.S. Sex-Specific Placental Responses in Fetal Development. Endocrinology 2015, 156, 3422–3434. [Google Scholar] [CrossRef] [Green Version]
- Seidmann, L.; Kamyshanskiy, Y.; Martin, S.Z.; Fruth, A.; Roth, W. Immaturity for Gestational Age of Microvasculature and Placental Barrier in Term Placentas with High Weight. Eur. J. Obstet. Gynecol. Reprod. Biol. 2017, 215, 134–140. [Google Scholar] [CrossRef]
- Benirschke, K.; Kaufmann, P. Three-Dimensional Aspects of Villous Maldevelopment. In Pathology of the Human Placenta; Springer: New York, NY, USA, 1995; pp. 167–181. [Google Scholar]
- Fujii, T.; Nagamatsu, T.; Morita, K.; Schust, D.J.; Iriyama, T.; Komatsu, A.; Osuga, Y.; Fujii, T. Enhanced HIF2α Expression during Human Trophoblast Differentiation into Syncytiotrophoblast Suppresses Transcription of Placental Growth Factor. Sci. Rep. 2017, 7, 12455. [Google Scholar] [CrossRef] [Green Version]
- Roland, C.S.; Hu, J.; Ren, C.-E.; Chen, H.; Li, J.; Varvoutis, M.S.; Leaphart, L.W.; Byck, D.B.; Zhu, X.; Jiang, S.-W. Morphological Changes of Placental Syncytium and Their Implications for the Pathogenesis of Preeclampsia. Cell. Mol. Life Sci. 2016, 73, 365–376. [Google Scholar] [CrossRef] [Green Version]
- Workalemahu, T.; Ouidir, M.; Shrestha, D.; Wu, J.; Grantz, K.L.; Tekola-Ayele, F. Differential DNA Methylation in Placenta Associated With Maternal Blood Pressure During Pregnancy. Hypertension 2020, 75, 1117–1124. [Google Scholar] [CrossRef] [PubMed]
- Fox, R.; Kitt, J.; Leeson, P.; Aye, C.Y.L.; Lewandowski, A.J. Preeclampsia: Risk Factors, Diagnosis, Management, and the Cardiovascular Impact on the Offspring. J. Clin. Med. 2019, 8, 1625. [Google Scholar] [CrossRef] [Green Version]
- Girchenko, P.; Lahti, J.; Czamara, D.; Knight, A.K.; Jones, M.J.; Suarez, A.; Hämäläinen, E.; Kajantie, E.; Laivuori, H.; Villa, P.M.; et al. Associations between Maternal Risk Factors of Adverse Pregnancy and Birth Outcomes and the Offspring Epigenetic Clock of Gestational Age at Birth. Clin. Epigenet. 2017, 9, 49. [Google Scholar] [CrossRef] [Green Version]
- Mayne, B.T.; Leemaqz, S.Y.; Smith, A.K.; Breen, J.; Roberts, C.T.; Bianco-Miotto, T. Accelerated Placental Aging in Early Onset Preeclampsia Pregnancies Identified by DNA Methylation. Epigenomics 2016, 9, 279–289. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Du, L.; Qiao, Y.; Zhang, X.; Zheng, W.; Wu, Q.; Chen, Y.; Zhu, G.; Liu, Y.; Bian, Z.; et al. Ferroptosis Is Governed by Differential Regulation of Transcription in Liver Cancer. Redox Biol. 2019, 24, 101211. [Google Scholar] [CrossRef]
- Brunton, H.; Caligiuri, G.; Cunningham, R.; Upstill-Goddard, R.; Bailey, U.-M.; Garner, I.M.; Nourse, C.; Dreyer, S.; Jones, M.; Moran-Jones, K.; et al. HNF4A and GATA6 Loss Reveals Therapeutically Actionable Subtypes in Pancreatic Cancer. Cell Rep. 2020, 31, 107625. [Google Scholar] [CrossRef]
- Hsu, J.; Sage, J. Novel Functions for the Transcription Factor E2F4 in Development and Disease. Cell Cycle 2016, 15, 3183–3190. [Google Scholar] [CrossRef] [PubMed]
- Bhawe, K.; Roy, D. Interplay between NRF1, E2F4 and MYC Transcription Factors Regulating Common Target Genes Contributes to Cancer Development and Progression. Cell. Oncol. 2018, 41, 465–484. [Google Scholar] [CrossRef]
- Tseng, L.; Tang, M.; Wang, Z.; Mazella, J. Progesterone Receptor (HPR) Upregulates the Fibronectin Promoter Activity in Human Decidual Fibroblasts. DNA Cell Biol. 2003, 22, 633–640. [Google Scholar] [CrossRef]
- O’Rawe, J.A.; Wu, Y.; Dorfel, M.J.; Rope, A.F.; Au, P.Y.; Parboosingh, J.S.; Moon, S.; Kousi, M.; Kosma, K.; Smith, C.S.; et al. TAF1 Variants Are Associated with Dysmorphic Features, Intellectual Disability, and Neurological Manifestations. Am. J. Hum. Genet. 2015, 97, 922–932. [Google Scholar] [CrossRef] [Green Version]
- Cheng, H.; Capponi, S.; Wakeling, E.; Marchi, E.; Li, Q.; Zhao, M.; Weng, C.; Piatek, S.G.; Ahlfors, H.; Kleyner, R.; et al. Missense Variants in TAF1 and Developmental Phenotypes: Challenges of Determining Pathogenicity. Hum. Mutat. 2020, 41, 449–464. [Google Scholar] [CrossRef]
- Oh, H.R.; An, C.H.; Yoo, N.J.; Lee, S.H. Frameshift Mutations in the Mononucleotide Repeats of TAF1 and TAF1L Genes in Gastric and Colorectal Cancers with Regional Heterogeneity. Pathol. Oncol. Res. 2017, 23, 125–130. [Google Scholar] [CrossRef]
- Fuke, C.; Shimabukuro, M.; Petronis, A.; Sugimoto, J.; Oda, T.; Miura, K.; Miyazaki, T.; Ogura, C.; Okazaki, Y.; Jinno, Y. Age Related Changes in 5-Methylcytosine Content in Human Peripheral Leukocytes and Placentas: An HPLC-Based Study. Ann. Hum. Genet. 2004, 68, 196–204. [Google Scholar] [CrossRef]
- Novakovic, B.; Yuen, R.K.; Gordon, L.; Penaherrera, M.S.; Sharkey, A.; Moffett, A.; Craig, J.M.; Robinson, W.P.; Saffery, R. Evidence for Widespread Changes in Promoter Methylation Profile in Human Placenta in Response to Increasing Gestational Age and Environmental/Stochastic Factors. BMC Genom. 2011, 12, 529. [Google Scholar] [CrossRef] [Green Version]
- Maccani, J.; Maccani, M. Altered Placental DNA Methylation Patterns Associated with Maternal Smoking: Current Perspectives. Adv. Genom. Genet. 2015, 2015, 205. [Google Scholar] [CrossRef] [Green Version]
- Suter, M.; Ma, J.; Harris, A.; Patterson, L.; Brown, K.A.; Shope, C.; Showalter, L.; Abramovici, A.; Aagaard-Tillery, K.M. Maternal Tobacco Use Modestly Alters Correlated Epigenome-Wide Placental DNA Methylation and Gene Expression. Epigenetics 2011, 6, 1284–1294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikeska, T.; Craig, J.M. DNA Methylation Biomarkers: Cancer and Beyond. Genes 2014, 5, 821–864. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Characteristics | PE Cases, N = 29 (%) | Controls, N = 82 (%) | p-Value |
|---|---|---|---|
| Maternal age (years) | 31.2 ± 5.6 | 30.2 ± 3.9 | 0.30 |
| <35 | 20 (69.0) | 70 (85.4) | 0.053 |
| 35≤ | 9 (31.0) | 12 (14.6) | |
| Maternal pregestational body mass index (BMI) (kg/m2) | |||
| <18.5 | 0 (0) | 1 (1.2) | 0.062 |
| 18.5–24.9 | 11 (37.9) | 44 (53.7) | |
| 25–29.9 | 7 (24.1) | 25 (30.5) | |
| 30≤ | 11 (37.9) | 12 (14.6) | |
| Maternal parity | |||
| Nullipara | 17 (58.6) | 60 (73.2) | 0.26 |
| Primipara | 10 (34.5) | 16 (19.5) | |
| Multipara | 2 (6.9) | 6 (7.3) | |
| Previous gestational hypertension | 0 (0) | 2 (2.4) | 0.40 |
| Previous preeclampsia | 6 (20.7) | 6 (7.3) | 0.046 |
| Mode of delivery | |||
| Vaginal | 22 (75.9) | 67 (81.7) | 0.50 |
| Caesarean | 7 (24.1) | 15 (18.3) | |
| Fetal sex | |||
| Male | 16 (55.2) | 43 (52.4) | 0.80 |
| Female | 13 (44.8) | 39 (47.6) | |
| Preterm birth (<week 37) | 4 (13.8) | 4 (4.9) | 0.11 |
| Fetal weight (g) | 3485 ± 562 | 3282 ± 753 | |
| Year of birth | |||
| 2008 | 6 (20.7) | 15 (18.3) | 0.11 |
| 2009 | 8 (27.6) | 21 (25.6) | |
| 2010 | 2 (6.9) | 0 | |
| 2011 | 2 (6.9) | 19 (23.2) | |
| 2014 | 5 (17.2) | 13 (15.9) | |
| 2015 | 6 (20.7) | 14 (17.1) | |
| Gestational age by week | 38.5 ± 2.1 | 39.5 ± 1.7 | 0.014 |
| CpG | Chr | Gene | Gene Name | 2log FC (95% CI) a | Beta b | q-Value |
|---|---|---|---|---|---|---|
| Controls with low NOx exposure (reference) vs. PE cases with high NOx exposure | ||||||
| cg27596779 | 5 | NAc | 0.78 (0.72, 0.86) | 0.64 | 0.028 | |
| cg26672098 | 2 | NA | 0.75 (0.68, 0.83) | 0.34 | 0.028 | |
| cg07806361 | 3 | CACNA2D3 | Calcium voltage-gated channel auxiliary subunit alpha 2 delta 3 | 0.62 (0.53, 0.73) | 0.72 | 0.031 |
| cg23935220 | 20 | RPRD1B | Regulation of nuclear pre-mRNA domain containing 1B | 0.77 (0.70, 0.85) | 0.18 | 0.034 |
| cg05694331 | 1 | SH3D21 | SH3 domain-containing protein 21 | 0.78 (0.71, 0.85) | 0.68 | 0.034 |
| cg15999356 | 11 | YAP1 | Yes1-associated transcriptional regulator | 0.77 (0.70, 0,85) | 0.66 | 0.036 |
| Controls with high NOx exposure (reference) vs. PE cases with high NOx exposure | ||||||
| cg17283620 | 2 | HAAO | 3-hydroxyanthranilate 3,4-dioxygenase | 0.1 (0.05, 0.21) | 0.14 | 0.005 |
| cg26672098 | 2 | NA | 0.74 (0.67, 0.81) | 0.34 | 0.006 | |
| cg24832457 | 15 | NA | 0.73 (0.66, 0.81) | 0.56 | 0.010 | |
| cg15534461 | 12 | PEBP1 | phosphatidylethanolamine binding protein 1 | 0.70 (0.62, 0.8) | 0.69 | 0.021 |
| cg18531351 | 15 | NA | 0.77 (0.7, 0.84) | 0.61 | 0.021 | |
| cg02404739 | 11 | FAM111B | FAM111 trypsin-like peptidase B | 2.58 (1.85, 3.63) | 0.16 | 0.021 |
| cg07715379 | 3 | PLCXD2 | phosphatidylinositol specific phospholipase C X domain containing 2 | 0.76 (0.68, 0.84) | 0.5 | 0.042 |
| cg18235274 | 12 | CD163 | CD163 molecule, also known as M130 | 0.68 (0.59, 0.78) | 0.83 | 0.042 |
| cg23502295 | 6 | MICAL1 | microtubule associated monooxygenase, calponin, and LIM domain containing 1 | 0.77 (0.70, 0.85) | 0.83 | 0.042 |
| cg15999356 | 11 | YAP1 | Yes1 associated transcriptional regulator | 0.77 (0.74, 0.87) | 0.66 | 0.042 |
| cg07249517 | 3 | MB21D2 | Mab-21 domain containing 2 also known as:C3orf59 | 0.80 (0.74, 0.87) | 0.56 | 0.047 |
| cg08493590 | 6 | SUPT3H | SPT3 homolog, SAGA, and STAGA complex component | 0.74 (0.66, 0.83) | 0.66 | 0.047 |
| cg02171814 | 17 | SERPINF2 | serpin family F member 2 | 0.74 (0.66, 0.82) | 0.61 | 0.047 |
| Controls with high NOx exposure (reference) vs. PE cases with low NOx exposure | ||||||
| cg18904784 | 4 | SCLT1 | sodium channel and clathrin linker 1 | 0.54 (0.45, 0.67) | 0.02 | 0.031 |
| Female pregnancies only—Controls with high NOx exposure (reference) vs. PE cases with high NOx exposure | ||||||
| cg11398400 | 17 | RTN4RL1 | reticulon 4 receptor like 1 | 2.71 (2, 3.66) | 0.95 | 0.011 |
| cg09234983 | 13 | LATS2 | large tumor suppressor kinase 2 | 0.59 (0.5, 0.69) | 0.39 | 0.011 |
| cg22343476 | 1 | NA | 0.46 (0.36, 0.59) | 0.24 | 0.011 | |
| cg08568550 | 11 | C11orf16 | chromosome 11 open reading frame 16 | 0.65 (0.56, 0.74) | 0.57 | 0.011 |
| cg24474409 | 7 | ACTR3C | actin related protein 3C | 0.47 (0.37, 0.59) | 0.74 | 0.011 |
| cg17764946 | 3 | TRAK1 | trafficking kinesin protein 1 | 0.46 (0.36, 0.59) | 0.88 | 0.011 |
| cg04219544 | 17 | KRT24 | keratin 24 | 0.57 (0.48, 0.68) | 0.3 | 0.011 |
| cg24551459 | 17 | RAB5C | member RAS oncogene family | 0.46 (0.35, 0.59) | 0.74 | 0.011 |
| cg14620234 | 9 | NA | 0.67 (0.59, 0.76) | 0.31 | 0.015 | |
| cg11173246 | 11 | TSPAN4 | tetraspanin 4 | 0.44 (0.33, 0.57) | 0.3 | 0.015 |
| cg08157194 | 22 | SLC25A17 | Solute Carrier Family 25 Member 17 | 0.6 (0.51, 0.71) | 0.55 | 0.016 |
| cg04574034 | 17 | PRAC | PRAC small nuclear protein | 2.88 (2.03, 4.08) | 0.8 | 0.016 |
| cg24874090 | 8 | NA | 0.62 (0.53, 0.73) | 0.64 | 0.019 | |
| Female pregnancies only—Controls with high NOx exposure (reference) vs. PE cases with low NOx exposure | ||||||
| cg16162930 | 1 | NA | 0.10 (0.05, 0.20) | 0.57 | 0.015 | |
| Male pregnancies only—Controls with high NOx exposure (reference) vs. PE cases with low NOx exposure | ||||||
| cg09804439 | 17 | STAT3 | Signal transducer and activator of transcription 3 | 0.15 (0.09, 0.26) | 0.02 | 0.004 |
| Gene | Gene Name | 2logFC (95% CI) a | p-Value (Unadjusted) | q-Value |
|---|---|---|---|---|
| ESCO2 | establishment of sister chromatid cohesion N-acetyltransferase 2 | 2.4 (1.7, 3.3) | 0.00002 | 0.33 |
| MUC20P1 | mucin 20, cell surface associated pseudogene 1 | 19.0 (6.1, 59.4) | 0.00003 | 0.33 |
| LINC01003 | long intergenic non-protein coding RNA 1003 | 0.58 (0.47, 0.73) | 0.0001 | 0.64 |
| KCNA4 | potassium voltage-gated channel subfamily A member 4 | 3.6 (1.9, 6.7) | 0.0004 | 0.94 |
| TTC3 | tetratricopeptide repeat domain 3 | 1.4 (1.2, 1.6) | 0.0005 | 0.94 |
| ABCC13 | ATP binding cassette subfamily C member 13 (pseudogene) | 0.36 (0.21, 0.60) | 0.0006 | 0.94 |
| GNAI1 | G protein subunit alpha i1 | 0.72 (0.61, 0.86) | 0.0009 | 0.94 |
| SRRM3 | serine/arginine repetitive matrix 3 | 0.44 (0.27, 0.69) | 0.0013 | 0.94 |
| AL451007.2 | 0.29 (0.15, 0.58) | 0.0014 | 0.94 | |
| PCDHB18P | protocadherin beta 18 pseudogene | 3.3 (1.7, 6.7) | 0.0016 | 0.94 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Engström, K.; Mandakh, Y.; Garmire, L.; Masoumi, Z.; Isaxon, C.; Malmqvist, E.; Erlandsson, L.; Hansson, S.R. Early Pregnancy Exposure to Ambient Air Pollution among Late-Onset Preeclamptic Cases Is Associated with Placental DNA Hypomethylation of Specific Genes and Slower Placental Maturation. Toxics 2021, 9, 338. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics9120338
Engström K, Mandakh Y, Garmire L, Masoumi Z, Isaxon C, Malmqvist E, Erlandsson L, Hansson SR. Early Pregnancy Exposure to Ambient Air Pollution among Late-Onset Preeclamptic Cases Is Associated with Placental DNA Hypomethylation of Specific Genes and Slower Placental Maturation. Toxics. 2021; 9(12):338. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics9120338
Chicago/Turabian StyleEngström, Karin, Yumjirmaa Mandakh, Lana Garmire, Zahra Masoumi, Christina Isaxon, Ebba Malmqvist, Lena Erlandsson, and Stefan R. Hansson. 2021. "Early Pregnancy Exposure to Ambient Air Pollution among Late-Onset Preeclamptic Cases Is Associated with Placental DNA Hypomethylation of Specific Genes and Slower Placental Maturation" Toxics 9, no. 12: 338. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics9120338