Assessment of In Vitro Bioaccessibility and In Vivo Oral Bioavailability as Complementary Tools to Better Understand the Effect of Cooking on Methylmercury, Arsenic, and Selenium in Tuna



Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals, Surgeries, and Treatments
2.2. Blood Collection and Analysis
2.3. In Vitro Digestion Experiment
2.4. Total Mercury Analysis
2.5. MeHg Analysis
2.6. Other Metal(loid)s
2.7. Arsenic Speciation
2.8. Quality Control for Metal(loid) Analyses
2.9. Data Handling and Statistics
3. Results
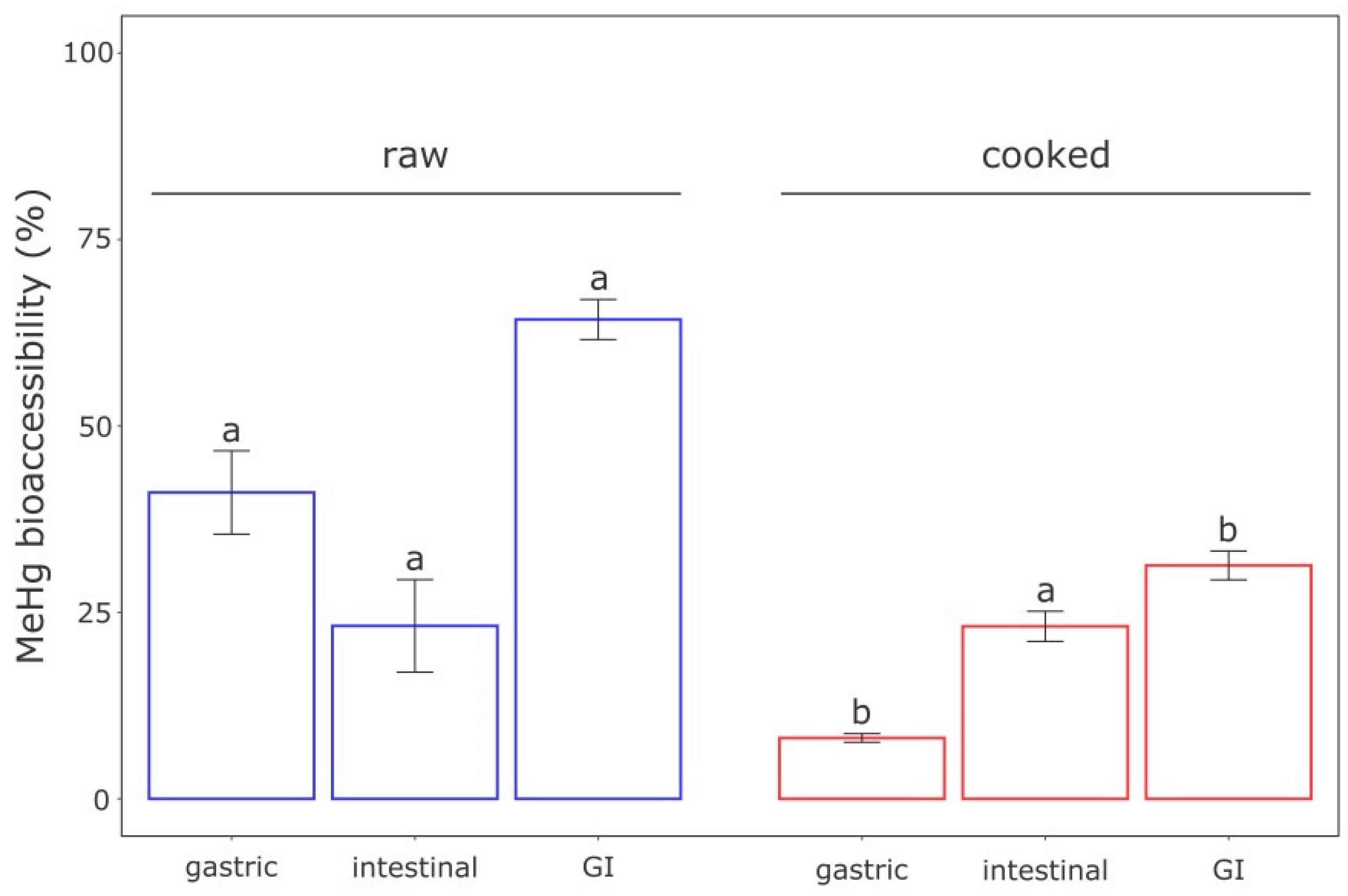
3.1. In Vitro Bioaccessibility of MeHg
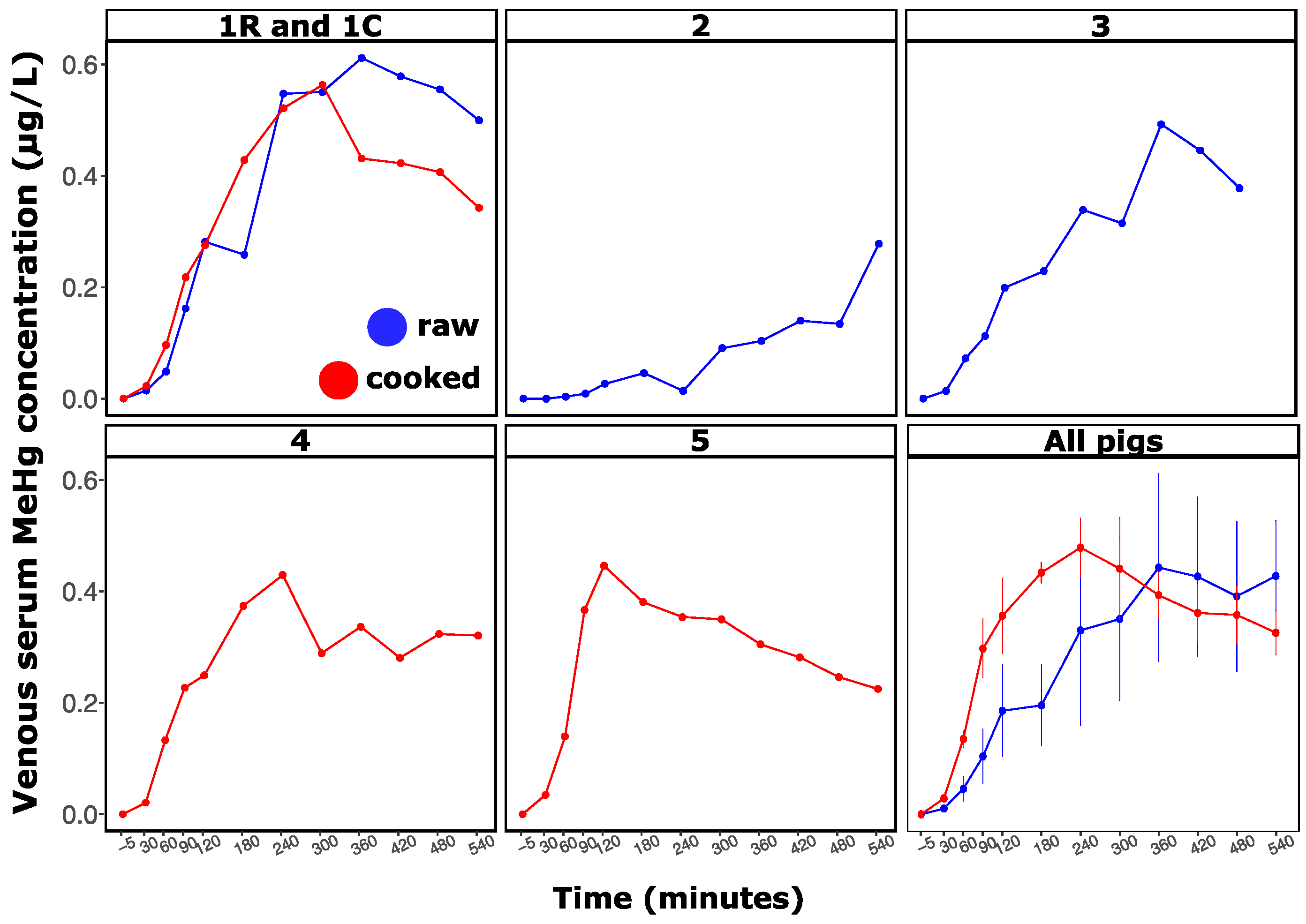
3.2. Effect of Cooking on Methylmercury, Arsenic, and Selenium Kinetics in the Pig Digestive System
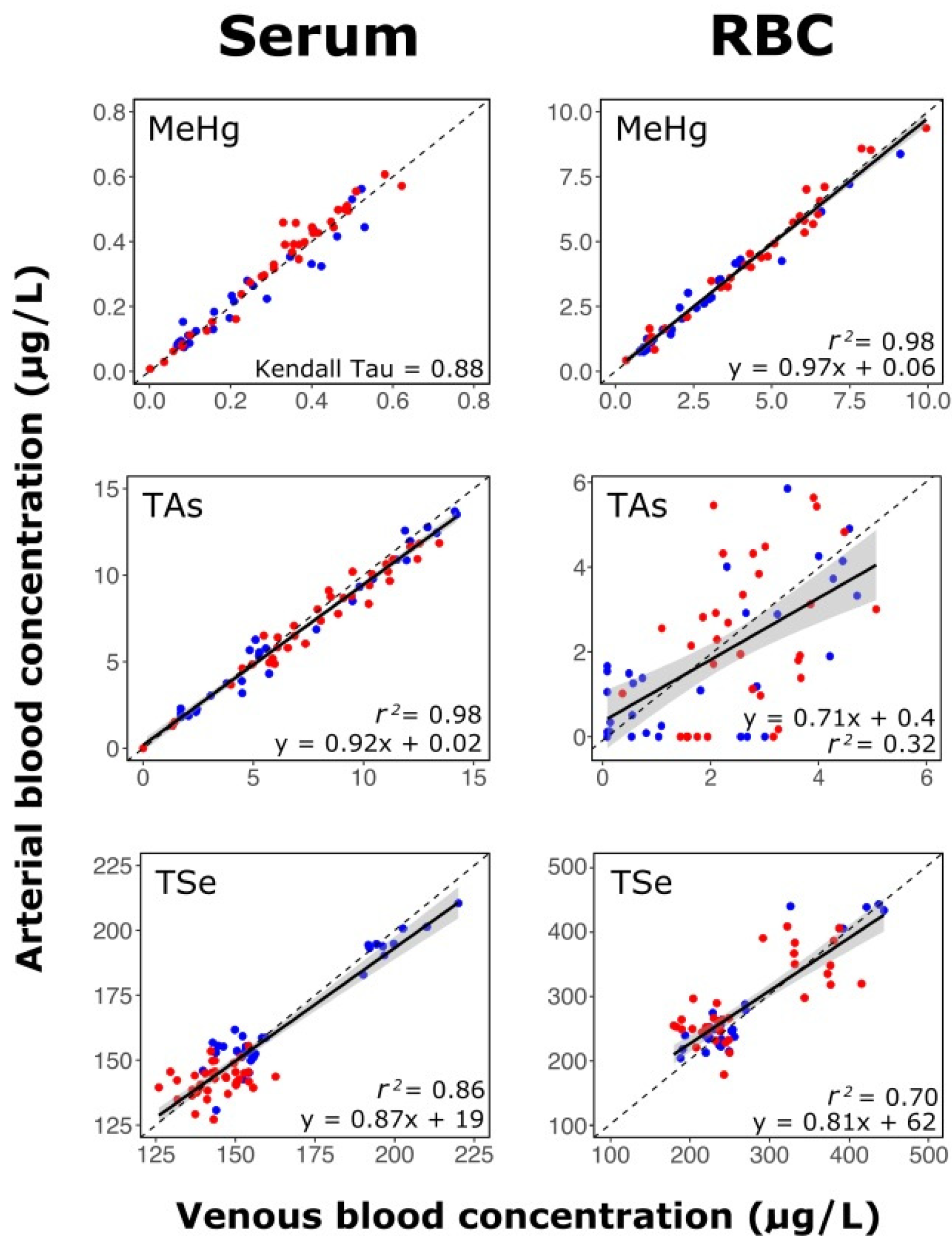
3.3. Stability of Methylmercury, Arsenic, and Selenium Distribution between Blood Compartments
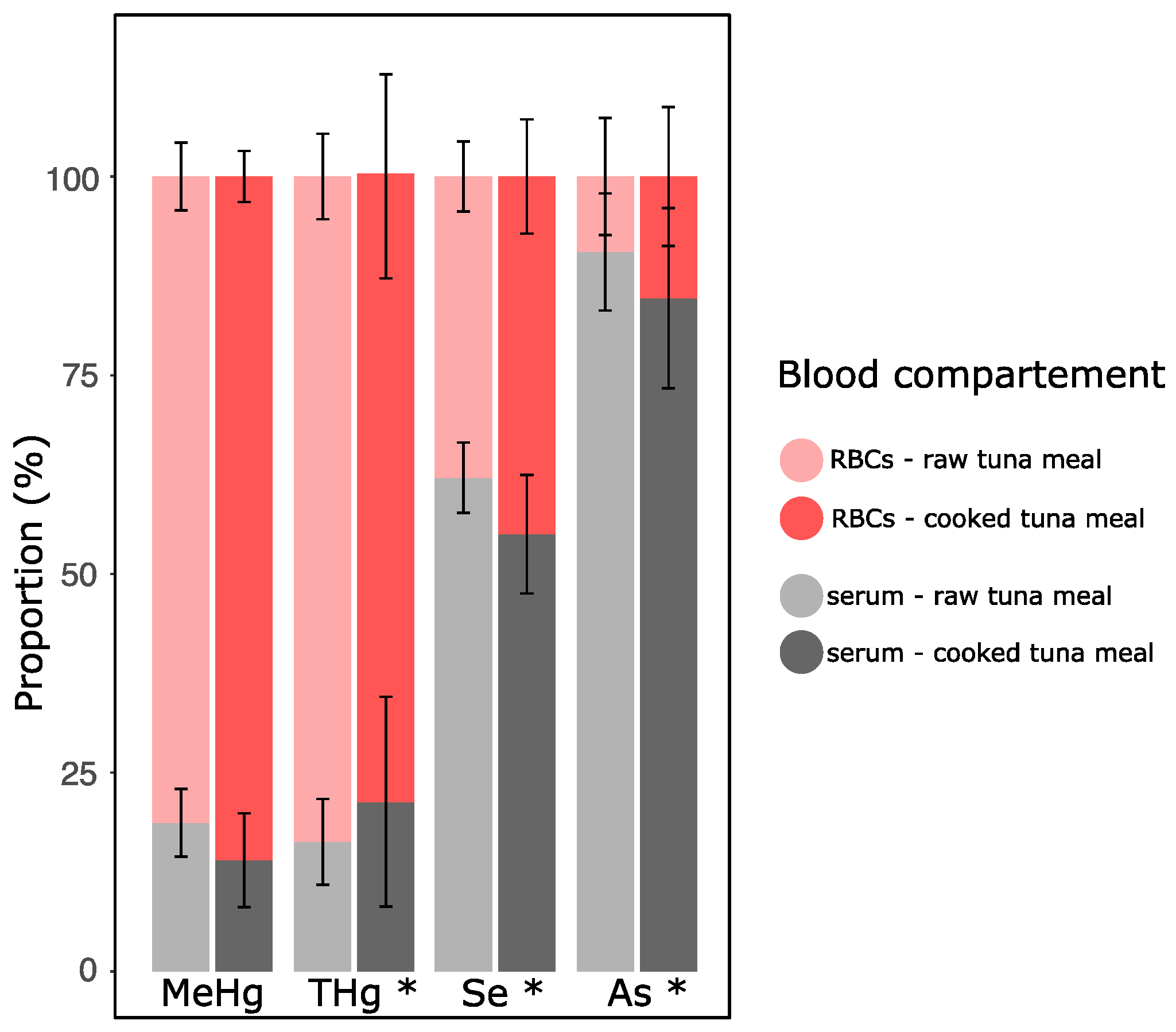
3.4. Cooking Effect on Metal(loid) Distribution between Blood Compartments
4. Discussion
4.1. Differing Effects of Cooking on the In Vitro and In Vivo Model Results
4.2. Cooking Does Not Influence the MeHg Fraction in Blood
4.3. Cooking Affects Methylmercury and Arsenic Kinetics of Oral Bioavailability in Blood
4.4. Partitioning of MeHg in Blood Compartments Differed for As and Se
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO/WHO. Report of the Joint FAO/WHO Expert Consultation on the Risks and Benefits of Fish Consumption; World Health Organization: Rome, Italy, 2010; Volume FIPM/R978. [Google Scholar]
- National Research Council. Toxicological Effects of Methylmercury; National Academy Press: Washington, DC, USA, 2000. [Google Scholar]
- Bourdineaud, J.P.; Fujimura, M.; Laclau, M.; Sawada, M.; Yasutake, A. Deleterious effects in mice of fish-associated methylmercury contained in a diet mimicking the Western populations’ average fish consumption. Environ. Int. 2011, 37, 303–313. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority. Scientific Opinion on the Risk for Public Health Related to the Presence of Mercury and Methylmercury in Food; European Food Safety Authority: Parma, Italy, 2012; Volume 10. [Google Scholar]
- Health Canada. Health Risk Assessment of Mercury in Fish and the Health Benefits of Fish Consumption; Health Canada: Ottawa, ON, Canada, 2007.
- Magalhães, M.C.; Costa, V.; Menezes, G.M.; Pinho, M.R.; Santos, R.S.; Monteiro, L.R. Intra- and inter-specific variability in total and methylmercury bioaccumulation by eight marine fish species from the Azores. Mar. Pollut. Bull. 2007, 54, 1654–1662. [Google Scholar] [CrossRef] [PubMed]
- Cappon, C.J.; Smith, J.C. Chemical form and distribution of mercury and selenium in canned tuna. J. Appl. Toxicol. 1982, 2, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Storelli, M.M.; Giacominelli-Stuffler, R.; Storelli, A.; Marcotrigiano, G.O. Accumulation of mercury, cadmium, lead and arsenic in swordfish and bluefin tuna from the Mediterranean Sea: A comparative study. Mar. Pollut. Bull. 2005, 50, 1004–1007. [Google Scholar] [CrossRef] [PubMed]
- Peterson, S.A.; Ralston, N.V.C.; Whanger, P.D.; Oldfield, J.E.; Mosher, W.D. Selenium and mercury interactions with emphasis on fish tissue. Environ. Bioindic. 2009, 4, 318–334. [Google Scholar] [CrossRef]
- Yamashita, Y.; Amlund, H.; Suzuki, T.; Hara, T.; Hossain, M.A.; Yabu, T.; Touhata, K.; Yamashita, M. Selenoneine, total selenium, and total mercury content in the muscle of fishes. Fish. Sci. 2011, 77, 679–686. [Google Scholar] [CrossRef]
- Cabañero, A.I.; Carvalho, C.; Madrid, Y.; Batoréu, C.; Cámara, C. Quantification and speciation of mercury and selenium in fish samples of high consumption in Spain and Portugal. Biol. Trace Elem. Res. 2005, 103, 17–35. [Google Scholar] [CrossRef]
- Zeng, H.; Uthus, E.O.; Combs, G.F. Mechanistic aspects of the interaction between selenium and arsenic. J. Inorg. Biochem. 2005, 99, 1269–1274. [Google Scholar] [CrossRef]
- Leufroy, A.; Noël, L.; Dufailly, V.; Beauchemin, D.; Guérin, T. Determination of seven arsenic species in seafood by ion exchange chromatography coupled to inductively coupled plasma-mass spectrometry following microwave assisted extraction: Method validation and occurrence data. Talanta 2011, 83, 770–779. [Google Scholar] [CrossRef]
- De Rosemond, S.; Xie, Q.; Liber, K. Arsenic concentration and speciation in five freshwater fish species from Back Bay near Yellowknife, NT, Canada. Environ. Monit. Assess. 2008, 147, 199–210. [Google Scholar] [CrossRef]
- Shah, A.Q.; Kazi, T.G.; Baig, J.A.; Arain, M.B.; Afridi, H.I.; Kandhro, G.A.; Wadhwa, S.K.; Kolachi, N.F. Determination of inorganic arsenic species (As3+ and As5+) in muscle tissues of fish species by electrothermal atomic absorption spectrometry (ETAAS). Food Chem. 2010, 119, 840–844. [Google Scholar] [CrossRef]
- Charette, T.; Rosabal, M.; Amyot, M. Mapping metal (Hg, As, Se), lipid and protein levels within fish muscular system in two fish species (Striped Bass and Northern Pike). Chemosphere 2021, 265, 129036. [Google Scholar] [CrossRef] [PubMed]
- Taylor, V.; Goodale, B.; Raab, A.; Schwerdtle, T.; Reimer, K.; Conklin, S.; Karagas, M.R.; Francesconi, K.A. Human exposure to organic arsenic species from seafood. Sci. Total Environ. 2016, 580, 266–282. [Google Scholar] [CrossRef] [PubMed]
- El-Begearmi, M.M.; Ganther, H.E.; Sunde, M.L. Dietary interaction between methylmercury, selenium, arsenic, and sulfur amino acids in Japanese quail. Poult. Sci. 1982, 61, 272–279. [Google Scholar] [CrossRef]
- Korbas, M.; Percy, A.J.; Gailer, J.; George, G.N. A possible molecular link between the toxicological effects of arsenic, selenium and methylmercury: Methylmercury(II) seleno bis(S-glutathionyl) arsenic(III). J. Biol. Inorg. Chem. 2008, 13, 461–470. [Google Scholar] [CrossRef]
- Moreda-Pineiro, J.; Moreda-Pineiro, A.; Romaris-Hortas, V.; Moscoso-Pérez, C.; Lopez-Mahia, P.; Muniategui-Lorenzo, S.; Bermejo-Barrera, P.; Prada-Rodriguez, D. In-vivo and in-vitro testing to assess the bioaccessibility and the bioavailability of arsenic, selenium and mercury species in food samples. TrAC Trends Anal. Chem. 2011, 30, 324–345. [Google Scholar] [CrossRef]
- Van de Wiele, T.R.; Oomen, A.G.; Wragg, J.; Cave, M.; Minekus, M.; Hack, A.; Cornelis, C.; Rompelburg, C.J.M.; De Zwart, L.L.; Klinck, B.; et al. Comparison of five in vitro digestion models to in vivo experimental results: Lead bioaccessibility in the human gastrointestinal tract. J. Environ. Sci. Heal. Part A Toxichazardous Subst. Environ. Eng. 2007, 42, 1203–1211. [Google Scholar] [CrossRef]
- Juhasz, A.L.; Smith, E.; Weber, J.; Rees, M.; Rofe, A.; Kuchel, T.; Sansom, L.; Naidu, R. Comparison of in vivo and in vitro methodologies for the assessment of arsenic bioavailability in contaminated soils. Chemosphere 2007, 69, 961–966. [Google Scholar] [CrossRef]
- Juhasz, A.L.; Weber, J.; Smith, E.; Naidu, R.; Rees, M.; Rofe, A.; Kuchel, T.; Sansom, L. Assessment of four commonly employed in vitro arsenic bioaccessibility assays for predicting in vivo relative arsenic bioavailability in contaminated soils. Environ. Sci. Technol. 2009, 43, 9487–9494. [Google Scholar] [CrossRef]
- Rees, M.; Sansom, L.; Rofe, A.; Juhasz, A.L.; Smith, E.; Weber, J.; Naidu, R.; Kuchel, T. Principles and application of an in vivo swine assay for the determination of arsenic bioavailability in contaminated matrices. Environ. Geochem. Health 2009, 31, 167–177. [Google Scholar] [CrossRef]
- Li, W.; Wang, W.X. In vivo oral bioavailability of fish mercury and comparison with in vitro bioaccessibility. Sci. Total Environ. 2019, 683, 648–658. [Google Scholar] [CrossRef] [PubMed]
- Anacleto, P.; Barbosa, V.; Alves, R.N.; Maulvault, A.L.; Bronze, M.R.; Marques, A. Green tea infusion reduces mercury bioaccessibility and dietary exposure from raw and cooked fish. Food Chem. Toxicol. 2020, 145, 135577. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.; Nunes, L.M.; Greenfield, B.K.; Qin, Z.; Yang, Q.; Huang, L.; Bu, W.; Zhong, H. Bioaccessibility-corrected risk assessment of urban dietary methylmercury exposure via fish and rice consumption in China. Sci. Total Environ. 2018, 630, 222–230. [Google Scholar] [CrossRef] [PubMed]
- Girard, C.; Charette, T.; Leclerc, M.; Shapiro, B.J.; Amyot, M. Cooking and co-ingested polyphenols reduce in vitro methylmercury bioaccessibility from fish and may alter exposure in humans. Sci. Total Environ. 2017, 616–617, 863–874. [Google Scholar] [CrossRef]
- Ouédraogo, O.; Amyot, M. Effects of various cooking methods and food components on bioaccessibility of mercury from fish. Environ. Res. 2011, 111, 1064–1069. [Google Scholar] [CrossRef]
- Afonso, C.; Costa, S.; Cardoso, C.; Oliveira, R.; Lourenço, H.M.; Viula, A.; Batista, I. Benefits and risks associated with consumption of raw, cooked, and canned tuna (Thunnus spp.) based on the bioaccessibility of selenium and methylmercury. Environ. Res. 2015, 143, 130–137. [Google Scholar] [CrossRef]
- Brodkorb, A.; Egger, L.; Alminger, M.; Alvito, P.; Assunção, R.; Ballance, S.; Bohn, T.; Bourlieu-Lacanal, C.; Boutrou, R.; Carrière, F.; et al. INFOGEST static in vitro simulation of gastrointestinal food digestion. Nat. Protoc. 2019, 14, 991–1014. [Google Scholar] [CrossRef]
- Mahaffey, K.R.; Clickner, R.P.; Bodurow, C.C. Blood organic mercury and dietary mercury intake: National Health and Nutrition Examination Survey, 1999 and 2000. Environ. Health Perspect. 2004, 112, 562–570. [Google Scholar] [CrossRef] [Green Version]
- Sanzo, J.M.; Dorronsoro, M.; Amiano, P.; Amurrio, A.; Aguinagalde, F.X.; Azpiri, M.A.; EPIC Group of Spain. Estimation and validation of mercury intake associated with fish consumption in an EPIC cohort of Spain. Public Health Nutr. 2001, 4, 981–988. [Google Scholar] [CrossRef] [Green Version]
- Shibata, Y.; Yoshinaga, J.; Morita, M. Detection of arsenobetaine in human blood. Appl. Organomet. Chem. 1994, 8, 249–251. [Google Scholar] [CrossRef]
- Ito, K.; Palmer, C.D.; Steuerwald, A.J.; Parsons, P.J. Determination of five arsenic species in whole blood by liquid chromatography coupled with inductively coupled plasma mass spectrometry. J. Anal. At. Spectrom. 2010, 25, 1334–1342. [Google Scholar] [CrossRef]
- Kaise, T.; Watanabe, S.; Itoh, K. The acute toxicity of arsenobetaine. Chemosphere 1985, 14, 1327–1332. [Google Scholar] [CrossRef]
- Carrier, G.; Bouchard, M.; Brunet, R.C.; Caza, M. A toxicokinetic model for predicting the tissue distribution and elimination of organic and inorganic mercury following exposure to methyl mercury in animals and humans. II. Application and validation of the model in humans. Toxicol. Appl. Pharmacol. 2001, 171, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Hooda, S.; Matte, J.J.; Wilkinson, C.W.; Zijlstra, R.T. Technical note: An improved surgical model for the long-term studies of kinetics and quantification of nutrient absorption in swine. J. Anim. Sci. 2009, 87, 2013–2019. [Google Scholar] [CrossRef] [PubMed]
- Dalto, D.B.; Audet, I.; Girard, C.L.; Matte, J.J. Bioavailability of vitamin B12 from dairy products using a pig model. Nutrients 2018, 10, 1134. [Google Scholar] [CrossRef] [Green Version]
- Health Canada. Safe Cooking Temperatures. Available online: https://www.canada.ca/en/health-canada/services/general-food-safety-tips/safe-internal-cooking-temperatures.html (accessed on 12 December 2020).
- Strathe, A.B.; Danfær, A.; Chwalibog, A. A dynamic model of digestion and absorption in pigs. Anim. Feed Sci. Technol. 2008, 143, 328–371. [Google Scholar] [CrossRef]
- Davis, S.S.; Illum, L.; Hinchcliffe, M. Gastrointestinal transit of dosage forms in the pig. J. Pharm. Pharmacol. 2001, 53, 33–39. [Google Scholar] [CrossRef]
- DeSesso, J.M.; Jacobson, C.F. Anatomical and physiological parameters affecting gastrointestinal absorption in humans and rats. Food Chem. Toxicol. 2001, 39, 209–228. [Google Scholar] [CrossRef]
- Manet, L. Hématologie; Éditions m.: Paris, France, 1969. [Google Scholar]
- Ruby, M.V.; Davis, A.; Schoof, R.; Eberle, S.; Sellstone, C.M. Estimation of lead and arsenic bioavailability using a physiologically based extraction test. Environ. Sci. Technol. 1996, 30, 422–430. [Google Scholar] [CrossRef]
- Smith, M.E.; Morton, D.G. The Digestive System, 2nd ed.; Churchill Livingstone Elsevier: Toronto, ON, Canada, 2010. [Google Scholar]
- U.S. EPA. System Requirements and Design for the Integrated Exposure Uptake Biokinetic Model for Lead in Children (IEUBK) Windows® Version—32-Bit Version, Appendix C, IEUBKwin Parameter Dictionary, EPA OSWER No. 9285.7-43; U.S. Environmental Protection Agency: Washington, DC, USA, 2007.
- Ito, K.; Goessler, W.; Gürleyük, H.; Wels, B.; Palmer, C.D.; Verostek, M.F.; Parsons, P.J. An interlaboratory study of arsenic speciation analysis of whole blood. J. Anal. At. Spectrom. 2011, 26, 1740–1745. [Google Scholar] [CrossRef]
- Taleshi, M.S.; Edmonds, J.S.; Goessler, W.; Ruiz-Chancho, M.J.; Raber, G.; Jensen, K.B.; Francesconi, K.A. Arsenic-containing lipids are natural constituents of sashimi tuna. Environ. Sci. Technol. 2010, 44, 1478–1483. [Google Scholar] [CrossRef] [PubMed]
- Fournier, F.; Karasov, W.H.; Kenow, K.P.; Meyer, M.W.; Hines, R.K. The oral bioavailability and toxicokinetics of methylmercury in common loon (Gavia immer) chicks. Comp. Biochem. Physiol. Mol. Integr. Physiol. 2002, 133, 703–714. [Google Scholar] [CrossRef]
- Kershaw, T.G.; Dhahir, P.H.; Clarkson, T.W. The relationship between blood levels and dose of methylmercury in man. Arch. Environ. Health 1980, 35, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Wolfensohn, S.; Lloyd, M. Handbook of Laboratory Animal Management and Welfare, 4th ed.; Wiley-Blackwell: Chichester, UK, 2013. [Google Scholar]
- Ramasawmy, R.; Rogers, T.; Alcantar, M.A.; McGuirt, D.R.; Khan, J.M.; Kellman, P.; Xue, H.; Faranesh, A.Z.; Campbell-Washburn, A.E.; Lederman, R.J.; et al. Blood volume measurement using cardiovascular magnetic resonance and ferumoxytol: Preclinical validation. J. Cardiovasc. Magn. Reson. 2018, 20, 62. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.L.; Davit, B.; Lionberger, R.; Wahba, Z.; Ahn, H.Y.; Yu, L.X. Using partial area for evaluation of bioavailability and bioequivalence. Pharm. Res. 2011, 28, 1939–1947. [Google Scholar] [CrossRef] [PubMed]
- Cohen-Barak, O.; Barkay, H.; Rasamoelisolo, M.; Butler, K.; Yamada, K.; Bassan, M.; Yoon, E.; Spiegelstein, O. Assessment of the Pharmacokinetics, Pharmacodynamics, and Safety of Single Doses of TV-1106, a Long-Acting Growth Hormone, in Healthy Japanese and Caucasian Subjects. Clin. Pharmacol. Drug Dev. 2017, 6, 331–342. [Google Scholar] [CrossRef] [PubMed]
- Purves, R.D. Optimum numerical integration methods for estimation of area-under-the-curve (AUC) and area-under-the-moment-curve (AUMC). J. Pharmacokinet. Biopharm. 1992, 20, 211–226. [Google Scholar] [CrossRef] [PubMed]
- Afonso, C.; Costa, S.; Cardoso, C.; Coelho, I.; Castanheira, I.; Lourenço, H.; Gonçalves, S.; Oliveira, R.; Carvalho, M.L.; Martins, M.F.; et al. Bioaccessibility in risk-benefit analysis of raw and cooked seabream consumption. J. Food Compos. Anal. 2018, 68, 118–127. [Google Scholar] [CrossRef]
- He, M.; Ke, C.H.; Wang, W.X. Effects of cooking and subcellular distribution on the bioaccessibility of trace elements in two marine fish species. J. Agric. Food Chem. 2010, 58, 3517–3523. [Google Scholar] [CrossRef]
- Jadán Piedra, C.; Sánchez, V.; Vélez, D.; Devesa, V. Reduction of mercury bioaccessibility using dietary strategies. LWT Food Sci. Technol. 2016, 71, 10–16. [Google Scholar] [CrossRef]
- Torres-Escribano, S.; Velez, D.; Montoro, R.; Vélez, D.; Montoro, R. Mercury and methylmercury bioaccessibility in swordfish. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2010, 27, 327–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davies, K.J.A. Protein damage and degradation by oxygen radicals. I: General aspects. J. Biol. Chem. 1987, 262, 9895–9901. [Google Scholar] [CrossRef]
- Stadtman, E.R. Oxidation of free amino acids and amino acids residues in proteins by radiolysis and by metal-catalyzed reactions. Annu. Rev. Biochem. 1993, 62, 797–821. [Google Scholar] [CrossRef] [PubMed]
- Santé-Lhoutellier, V.; Astruc, T.; Marinova, P.; Greve, E.; Gatellier, P. Effect of meat cooking on physicochemical state and in vitro digestibility of myofibrillar proteins. J. Agric. Food Chem. 2008, 56, 1488–1494. [Google Scholar] [CrossRef]
- Grune, T.; Jung, T.; Merker, K.; Davies, K.J.A. Decreased proteolysis caused by protein aggregates, inclusion bodies, plaques, lipofuscin, ceroid, and aggresomes during oxidative stress, aging, and disease. Int. J. Biochem. Cell Biol. 2004, 36, 2519–2530. [Google Scholar] [CrossRef] [PubMed]
- Nong, Q.; Dong, H.; Liu, Y.; Liu, L.; He, B.; Huang, Y.; Jiang, J.; Luan, T.; Chen, B.; Hu, L. Characterization of the mercury-binding proteins in tuna and salmon sashimi: Implications for health risk of mercury in food. Chemosphere 2020, 263, 1–8. [Google Scholar] [CrossRef]
- Venugopal, V.; Shahidi, F. Structure and composition of fish muscle. Food Rev. Int. 1996, 12, 175–197. [Google Scholar] [CrossRef]
- Oberli, M.; Marsset-Baglieri, A.; Airinei, G.; Santé-Lhoutellier, V.; Khodorova, N.; Rémond, D.; Foucault-Simonin, A.; Piedcoq, J.; Tomé, D.; Fromentin, G.; et al. High true ileal digestibility but not postprandial utilization of nitrogen from bovine meat protein in humans is moderately decreased by high-temperature, long-duration cooking. J. Nutr. 2015, 145, 2221–2228. [Google Scholar] [CrossRef] [Green Version]
- Harris, H.H.; Pickering, I.J.; George, G.N. The chemical form of mercury in fish. Science 2003, 301, 1203. [Google Scholar] [CrossRef] [Green Version]
- Ballatori, N. Transport of toxic metals by molecular mimicry. Environ. Health Perspect. 2002, 110, 689–694. [Google Scholar] [CrossRef] [Green Version]
- Broer, S. Amino acid transport across mammalian intestinal and renal epithelia. Am. Physiol. Soc. 2008, 88, 249–286. [Google Scholar] [CrossRef]
- Vázquez, M.; Vélez, D.; Devesa, V. In vitro characterization of the intestinal absorption of methylmercury using a caco-2 cell model. Chem. Res. Toxicol. 2014, 27, 942–943. [Google Scholar] [CrossRef] [PubMed]
- Vázquez, M.; Vélez, D.; Devesa, V. Participation of b0,+ and B0,+ systems in the transport of mercury bound to cysteine in intestinal cells. Toxicol. Res. (Camb). 2015, 4, 895–900. [Google Scholar] [CrossRef]
- Bartell, S.M.; Ponce, R.A.; Sanga, R.N.; Faustman, E.M. Human variability in mercury toxicokinetics and steady state biomarker ratios. Environ. Res. 2000, 84, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Health Canada. Le mercure. Votre santé et l’environnement. Outil de ressources; Health Canada: Ottawa, ON, Canada, 2004.
- Sherlock, J.; Hislop, J.; Newton, D.; Topping, G.; Whittle, K. Elevation of mercury in human blood from controlled chronic ingestion of methylmercury in fish. Hum. Toxicol. 1984, 3, 117–131. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, L.R.; Furness, R.W. Kinetics, dose-response, and excretion of methylmercury in free-living adult Cory’s shearwaters. Environ. Sci. Technol. 2001, 35, 739–746. [Google Scholar] [CrossRef] [PubMed]
- Naganuma, A.; Koyama, Y.; Imura, N. Behavior of methylmercury in mammalian erythrocytes. Top. Catal. 1980, 54, 405–410. [Google Scholar] [CrossRef]
- Ehrlein, H.-J.; Pröve, J. Effect of viscosity of test meals on gastric emptying in gogs. Q. J. Exp. Physiol. 1982, 67, 419–425. [Google Scholar] [CrossRef]
- Liu, Y.M.; Lin., T.S.; Lanier, T.C. Thermal denaturation and aggregation of actomyosin from atlantic croaker. J. Food Sci. 1982, 47, 1916–1920. [Google Scholar] [CrossRef]
- Tam, G.K.H.; Charbonneau, S.M.; Bryce, F.; Sandi, E. Excretion of a single oral dose of fish-arsenic in man. Bull. Environ. Contam. Toxicol. 1982, 28, 669–673. [Google Scholar] [CrossRef]
- Lehmann, B.; Ebeling, E.; Alsen-Hinrichs, C. Kinetics of arsenic in human blood after a fish meal. Gesundheitswesen 2001, 63, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Kurokawa, S.; Berry, M.J. Selenium. Role of the essential metalloid in health. In Interrelations between Essential Metal Ions and Human Diseases; Springer: New York, NY, USA, 2013; Volume 23, pp. 1–7. ISBN 6176321972. [Google Scholar]
- Fairweather-Tait, S.J.; Collings, R.; Hurst, R. Selenium bioavailability: Current knowledge and future research requirements. Am. J. Clin. Nutr. 2010, 91, 1484–1491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalto, D.B.; Matte, J.J. Effects of different sources and levels of dietary iron and selenium on the postprandial net portal appearance of these minerals in growing pigs. J. Anim. Sci. 2020, 98, 1–9. [Google Scholar] [CrossRef]
- Jotty, K.; Ojeda, M.L.; Nogales, F.; Murillo, M.L.; Carreras, O. Selenium dietary supplementation as a mechanism to restore hepatic selenoprotein regulation in rat pups exposed to alcohol. Alcohol 2013, 47, 545–552. [Google Scholar] [CrossRef]
- Thomas, D.J.; Smith, J.C. Effects of coadministered low-molecular-weight thiol compounds on short-term distribution of methyl mercury in the rat. Toxicol. Appl. Pharmacol. 1982, 62, 104–110. [Google Scholar] [CrossRef]
- Naganuma, A.; Imura, N. Methylmercury binds to a low molecular weight substance in rabbit and human erythrocytes. Toxicol. Appl. Pharmacol. 1979, 47, 613–616. [Google Scholar] [CrossRef]
- Wang, F.; Lemes, M.; Khan, M.A.K. Metallomics of mercury: Role of thiol- and selenol-containing biomolecules. In Environmental Chemistry and Toxicology of Mercury; Liu, G., Cai, Y., O’Driscoll, N., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2012; pp. 517–544. ISBN 9780470578728. [Google Scholar]
- Nogara, P.A.; Oliveira, C.S.; Schmitz, G.L.; Piquini, P.C.; Farina, M.; Aschner, M.; Rocha, J.B.T. Methylmercury’s chemistry: From the environment to the mammalian brain. Biochim. Biophys. Acta Gen. Subj. 2019, 1863, 1–23. [Google Scholar] [CrossRef]
- Rabenstein, D.L.; Isab, A.A. A proton nuclear magnetic resonance study of the interaction of mercury with intact human erythrocytes. BBA Mol. Cell Res. 1982, 721, 374–384. [Google Scholar] [CrossRef]
- Ancora, S.; Rossi, R.; Di Simplicio, P.; Lusini, L.; Leonzio, C. In vitro study of methylmercury in blood of bottlenose dolphins (Tursiops truncatus). Arch. Environ. Contam. Toxicol. 2002, 42, 348–353. [Google Scholar] [CrossRef]
- Pei, K.L.; Gailer, J. Probing the interaction of arsenobetaine with blood plasma constituents in vitro: An SEC-ICP-AES study. Metallomics 2009, 1, 403–408. [Google Scholar] [CrossRef]
- Little, M.; Achouba, A.; Dumas, P.; Ouellet, N.; Ayotte, P.; Lemire, M. Determinants of selenoneine concentration in red blood cells of Inuit from Nunavik (Northern Québec, Canada). Environ. Int. 2019, 127, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Achouba, A.; Dumas, P.; Ouellet, N.; Lemire, M.; Ayotte, P. Plasma levels of selenium-containing proteins in Inuit adults from Nunavik. Environ. Int. 2016, 96, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, W.; Zhao, J.; Lin, X.; Liu, J.; Cui, L.; Gao, Y.; Zhang, T.L.; Li, B.; Li, Y.F. Selenoprotein P as the major transporter for mercury in serum from methylmercury-poisoned rats. J. Trace Elem. Med. Biol. 2018, 50, 589–595. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, Y.; Yamashita, M. Identification of a novel selenium-containing compound, selenoneine, as the predominant chemical form of organic selenium in the blood of bluefin tuna. J. Biol. Chem. 2010, 285, 18134–18138. [Google Scholar] [CrossRef] [Green Version]
- Yamashita, M.; Yamashita, Y.; Suzuki, T.; Kani, Y.; Mizusawa, N.; Imamura, S.; Takemoto, K.; Hara, T.; Hossain, M.A.; Yabu, T.; et al. Selenoneine, a novel selenium-containing compound, mediates detoxification mechanisms against methylmercury accumulation and toxicity in zebrafish embryo. Mar. Biotechnol. 2013, 15, 559–570. [Google Scholar] [CrossRef] [Green Version]
- Achouba, A.; Dumas, P.; Ouellet, N.; Little, M.; Lemire, M.; Ayotte, P. Selenoneine is a major selenium species in beluga skin and red blood cells of Inuit from Nunavik. Chemosphere 2019, 229, 549–558. [Google Scholar] [CrossRef]
- Shi, W.; Chance, M.R. Metallomics and metalloproteomics. Cell. Mol. Life Sci. 2008, 65, 3040–3048. [Google Scholar] [CrossRef]
- Canadian Council on Animal Care (CCAC). Guide to the Care and Use of Experimental Animals; Canadian Council on Animal Care: Ottawa, ON, Canada, 2009. [Google Scholar]
- National Farm Animal Care Council (NFACC). Code of Practice for Care and Handling of Pigs; National Farm Animal Care Council: Ottawa, ON, Canada, 2014. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Basal Diet, Nutrients 1. | Amount (%) | |||||
|---|---|---|---|---|---|---|
| Corn | 55.4 | |||||
| Soybean meal 48% | 15.1 | |||||
| Wheat | 15.0 | |||||
| Barley | 1.3 | |||||
| Distillers dried grain with solubles | 9.8 | |||||
| Amino acids | 3.5 | |||||
| Basal Diet, Contaminants 2 (n = 5) | Levels (ng/g) | |||||
| THg | 2.6 ± 1.3 | |||||
| TSe | 58.4 ± 7.5 | |||||
| TAs | ND | |||||
| Treatment 3 (n = 6) | Pig Number | MeHg (µg) | %MeHg 4 | TSe (µg) | TAs (µg) | %AsB 5 |
| Raw | 1R *, 2, 3 | 118.8 ± 26.7 | 94 ± 8% | 319.2 ± 39.4 | 508.8 ± 16.3 | 73 ± 9% |
| Cooked | 1C *, 4, 5 | 139.2 ± 9.4 | 93 ± 3% | 409.4 ± 45.4 | 598.4 ± 83 | 96 ± 5% |
| Raw | Cooked | |||||
|---|---|---|---|---|---|---|
| Pig Number | 1R * | 2 | 3 | 1C * | 4 | 5 |
| Preconsumption values (μg/L) | 0.20 | 0.07 | 0.09 | 0.06 | 0.08 | 0.002 |
| AUC(0–540) (μg × min/L/μg) | 1.7 | 0.3 | 1.8 | 1.5 | 1.2 | 1.1 |
| Cmax (t0 corrected) (μg/L) | 0.6 | 0.3 | 0.5 | 0.6 | 0.4 | 0.5 |
| Tmax (min) | 360 | NA † | 360 | 300 | 240 | 120 |
| Intake (μg) | 130 | 138 | 88 | 137 | 149 | 131 |
| Dose (μg/kg bw) | 2.5 | 2.4 | 1.7 | 2.6 | 2.3 | 2.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Charette, T.; Bueno Dalto, D.; Rosabal, M.; Matte, J.J.; Amyot, M. Assessment of In Vitro Bioaccessibility and In Vivo Oral Bioavailability as Complementary Tools to Better Understand the Effect of Cooking on Methylmercury, Arsenic, and Selenium in Tuna. Toxics 2021, 9, 27. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics9020027
Charette T, Bueno Dalto D, Rosabal M, Matte JJ, Amyot M. Assessment of In Vitro Bioaccessibility and In Vivo Oral Bioavailability as Complementary Tools to Better Understand the Effect of Cooking on Methylmercury, Arsenic, and Selenium in Tuna. Toxics. 2021; 9(2):27. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics9020027
Chicago/Turabian StyleCharette, Tania, Danyel Bueno Dalto, Maikel Rosabal, J. Jacques Matte, and Marc Amyot. 2021. "Assessment of In Vitro Bioaccessibility and In Vivo Oral Bioavailability as Complementary Tools to Better Understand the Effect of Cooking on Methylmercury, Arsenic, and Selenium in Tuna" Toxics 9, no. 2: 27. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics9020027