
Gossypol Exhibited Higher Detrimental Effect on Ruminal Fermentation Characteristics of Low-Forage in Comparison with High-Forage Mixed Feeds



Abstract
:1. Introduction
2. Materials and Methods
2.1. Gossypol Solution Preparation
2.2. Rumen Fluid Preparation
2.3. Experimental Design and In Vitro Incubation
2.4. Sampling and Analysis
2.4.1. Sample Collection
2.4.2. Chemical Analysis and Calculations
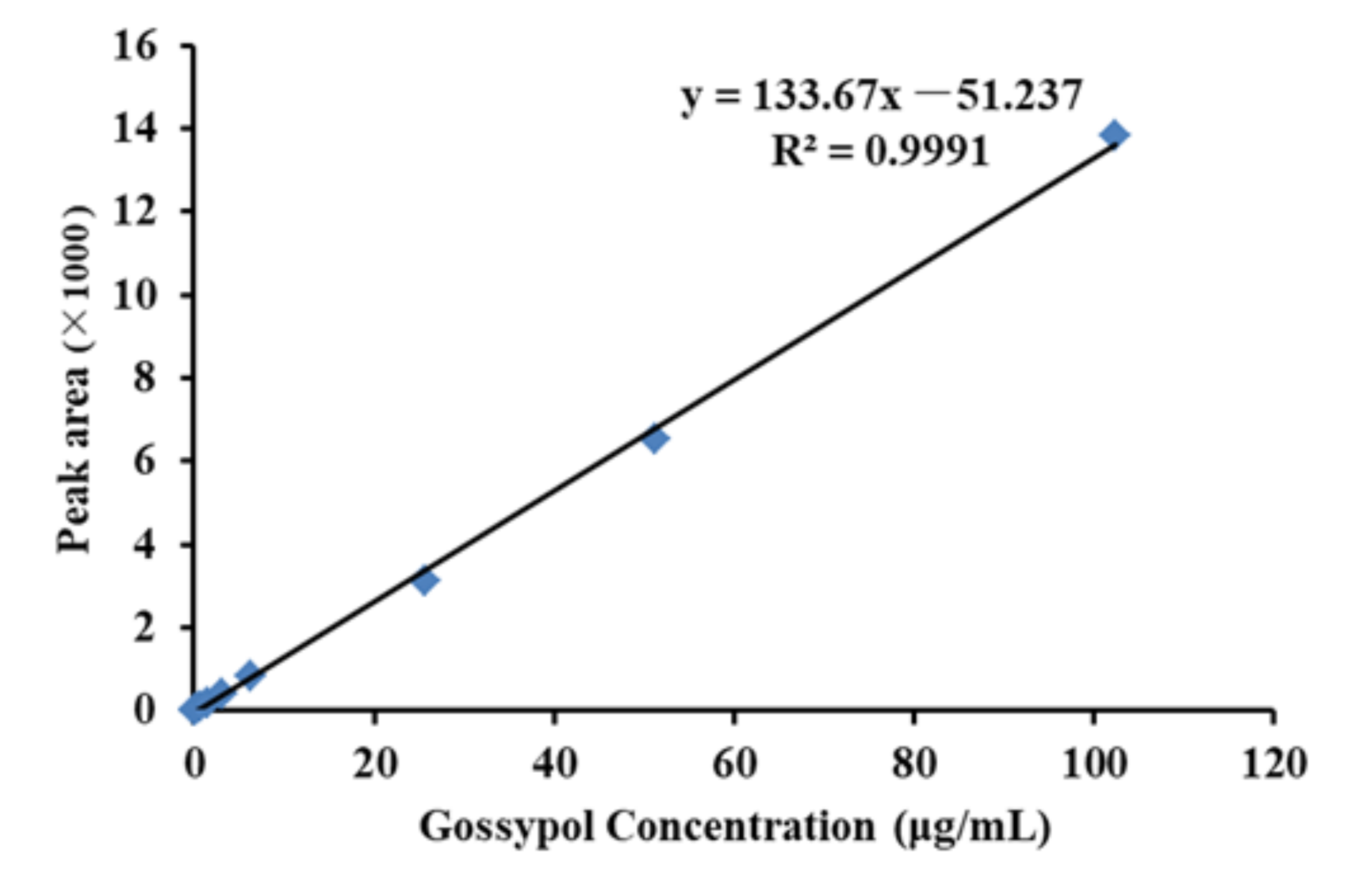
2.4.3. Gossypol Extraction and High-Performance Liquid Chromatography Analysis
2.4.4. DNA Extraction and Real-Time PCR Amplification of 16S RNA Genes
2.4.5. Preparation of Standard Plasmid for Real-Time PCR
2.5. Statistical Analysis
3. Results and Discussion
3.1. Effect of Gossypol on IVDMD and Kinetic Gas Production
3.2. Effect of Gossypol on In Vitro Rumen Fermentation Characteristics
3.3. Effect of Gossypol on Rumen Microbial Population
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Santos, J.E.P.; Villaseňor, M.; Depeters, E.J.; Robinson, P.H.; Baldwin, B.C., Jr. Type of cottonseed and level of gossypol in diets of lactating dairy cows: Effects on lactation performance and plasma gossypol. J. Dairy Sci. 2002, 85, 1491–1501. [Google Scholar] [CrossRef]
- Wang, X.; Howell, C.P.; Chen, F.; Yin, J.J.; Jiang, Y.M. Gossypol—A polyphenolic compound from cotton plant. Adv. Food Nutr. Res. 2009, 58, 215–263. [Google Scholar] [PubMed]
- Reiser, R.; Fu, H.C. The mechanism of gossypol detoxification by ruminant animals. J. Nutr. 1962, 76, 215–218. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.; Wang, Y. The nutrition value and metabolism of cottonseed in ruminants. Feed Res. 2011, 4, 13–16. [Google Scholar]
- Câmara, A.C.L.; Vale, A.M.D.; Mattoso, C.R.S.; Melo, M.M.; Soto-Blanco, B. Effects of gossypol from cottonseed cake on the blood profile in sheep. Trop. Anim. Health Pro. 2016, 48, 1–6. [Google Scholar] [CrossRef]
- Zbidah, M.; Lupescu, A.; Shaik, N.; Lang, F. Gossypol-induced suicidal erythrocyte death. Toxicology 2012, 302, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Higginbotham, G.E.; Santos, J.E.P.; Depeters, E.J. Effects of pelleting whole cottonseed on plasma gossypol, rumen metabolites, and performance of lactating dairy cows. Prof. Anim. Sci. 2004, 20, 413–421. [Google Scholar] [CrossRef]
- Santos, J.; Villasenor, M.; Robinson, P.H.; DePeters, E.J.; Holmberg, C.A. Type of cottonseed and level of gossypol in diets of lactating dairy cows: Plasma gossypol, health, and reproductive performance. J. Dairy Sci. 2003, 86, 892–905. [Google Scholar] [CrossRef]
- Chenoweth, P.J.; Chase, C.C., Jr.; Risco, C.A.; Larsen, R.E. Characterization of gossypol-induced sperm abnormalities in bulls. Theriogenology 2000, 53, 1193–1203. [Google Scholar] [CrossRef]
- Hassan, M.E.; Smith, G.W.; Ott, R.S.; Faulkner, D.B.; Firkins, L.D.; Ehrhart, E.J.; Schaeffer, D.J. Reversibility of the reproductive toxicity of gossypol in peripubertal bulls. Theriogenology 2004, 61, 1171–1179. [Google Scholar] [CrossRef]
- Yuan, Y.Y.; Shi, Q.X. Inhibition of hamster sperm acrosomal enzyme by gossypol is closely associated with the decrease in fertilization capacity. Contraception 2005, 62, 229–236. [Google Scholar] [CrossRef]
- Knutsen, H.K.; Barregård, L.; Bignami, M.; Brüschweiler, B.; Ceccatelli, S.; Dinovi, M.; Edler, L.; Kraupp, B.G.; Hogstrand, H.; Hoogenboom, L.; et al. Presence of free gossypol in whole cottonseed. EFSA J. 2017, 15, 4850–4865. [Google Scholar]
- Russell, J.B.; Rychlik, J.L. Factors that alter rumen microbial ecology. Science 2001, 292, 1119–1122. [Google Scholar] [CrossRef]
- Menke, K.H.; Steingass, H. Estimation of the energetic feed value obtained from chemical analysis and in vitro gas production using rumen fluid. Anim. Res. Dev. 1988, 28, 7–55. [Google Scholar]
- Zhang, D.F.; Yang, H.J. In vitro ruminal methanogenesis of a hay-rich substrate in response to different combination supplements of nitrocompounds; pyromellitic diimide and 2-bromoethanesulphonate. Anim. Feed Sci. Technol. 2011, 163, 20–32. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis of AOAC International, 17th ed.; Association of Official Analytical Chemists: Gaithersburg, MD, USA, 2005. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Verdouw, H.; Vanechteld, C.J.A.; Dekkers, E.M.J. Ammonia determination based on indophenol formation with sodium salicylate. Water Res. 1978, 12, 399–402. [Google Scholar] [CrossRef]
- Cui, J.H.; Yang, H.J.; Yu, C.Q.; Bai, S.; Wu, T.T.; Song, S.S.; Sun, W.; Shao, X.M.; Jiang, L.S. Effect of urea fertilization on biomass yield, chemical composition, in vitro rumen digestibility and fermentation characteristics of straw of highland barley planted in Tibet. J. Agric. Sci. 2016, 154, 151–164. [Google Scholar] [CrossRef]
- Groot, J.C.; Cone, J.W.; Williams, B.A. Multiphasic analysis of gas production kinetics for in vitro fermentation of ruminant feeds. Anim. Feed Sci. Technol. 1996, 64, 77–89. [Google Scholar] [CrossRef]
- SAS. Statistical Analytical System (SAS) Users Guides: Statistics; Version 8.2; Statistica Analysis Institute: Cary, NC, USA, 1999. [Google Scholar]
- Ørskov, E.R. Manipulation of rumen fermentation for maximum food utilization. World Rev. Nutr. Diet. 1975, 22, 152–182. [Google Scholar]
- Zhong, R.Z. Performance and Composition of Milk Fat of Holstein Cows Fed Whole Cottonseed During early Lactation. Master’s Thesis, Hebei Agricultural University, Shijiazhuang, China, 2007. [Google Scholar]
- Denman, S.E.; Mc Sweeney, C.S. Development of a real-time PCR assay for monitoring anaerobic fungal and cellulolytic bacterial populations within the rumen. FEMS Microbiol. Ecol. 2006, 58, 572–582. [Google Scholar] [CrossRef] [PubMed]
- Koike, S.; Kobayashi, Y. Development and use of competitive PCR assays for the rumen cellulolytic bacteria: Fibrobacter succinogenes, Ruminococcus albus and Ruminococcus flavefaciens. FEMS Microbiol. Lett. 2001, 204, 361–366. [Google Scholar] [CrossRef]
- Yang, S. Effect of Soybean Oil and Linseed Oil Supplementation on Population of Ruminal Bacteria and Fermentation Parameters in Dairy Cows. Ph.D. Thesis, Chinese Academy of Agricultural Sciences, Beijing, China, 2007. [Google Scholar]
- Stevenson, D.M.; Weimer, P. Dominance of Prevotella and low abundance of classical ruminal bacterial species in the bovine rumen revealed by relative quantification real-time PCR. Appl. Microbiol. Biotechnol. 2007, 75, 165–174. [Google Scholar] [CrossRef]
- Sylvester, J.T.; Karnati, S.K.; Yu, Z.; Morrison, M.; Firkins, J.L. Development of an assay to quantify rumen ciliate protozoal biomass in cows using realtime PCR. J. Nutr. 2004, 134, 3378–3384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DePeters, E.J.; Bath, D.L. Canola meal vensus cottonseed meal as the protein supplementin dairy diets. J. Dairy Sci. 1986, 58, 572–582. [Google Scholar]
- Van Soest, P.J. Nutritional Ecology of the Ruminant; Comstock, Cornell University Press: NewYork, NY, USA, 1982. [Google Scholar]
- Bergman, E.N. Energy contribution of volatile fatty acids from the gastrointestinal tract in various species. Physiol. Rev. 1990, 70, 567–590. [Google Scholar] [CrossRef] [Green Version]
- Herdt, T.H. Metabolic diseases of ruminant livestock: Fuel homeostasis in the ruminants. Vet. Clin. North Am. Food Anim. 1988, Pract4, 213–231. [Google Scholar] [CrossRef]
- Moss, A.R.; Jouany, J.; Newbold, J. Methane Production by Ruminants: Its Contribution to Global Warming; Annales de Zootechnie; Institut National de la Recherché Agronomique: Paris, France, 2000. [Google Scholar]
- Forsberg, C.W.; Cheng, K.J.; White, B.A. Polysaccharide degradation in the rumen and large intestine. In Gastrointestinal Microbiology; Mackie, R.I., White, B.A., Eds.; Springer: Boston, MA, USA, 1997; pp. 319–379. [Google Scholar]
- Michalet-Doreau, B.; Fernandez, I.; Peyron, C.; Millet, L.; Fonty, G. Fibrolytic activities and cellulolytic bacterial community structure in the solid and liquid phases of rumen contents. Reprod. Nutr. Dev. 2001, 41, 187–194. [Google Scholar] [CrossRef] [Green Version]
- Michalet-Doreau, B.; Fernandez, I.; Fonty, G. A comparison of enzymatic and molecular approaches to characterize the cellulolytic microbial ecosystems of the rumen and the cecum. J. Anim. Sci. 2002, 80, 790–796. [Google Scholar] [CrossRef] [PubMed]
- Koike, S.; Yoshitani, S.; Kobayashi, Y.; Tanaka, K. Kinetics of in sacco fiber-attachment of representative ruminal cellulolytic bacteria monitored by competitive PCR. J. Dairy Sci. 2003, 86, 1429–1435. [Google Scholar] [CrossRef]
- Willems, A.; Collins, M.D. Butyrivibrio. In Bergey’s Manual of Systematics of Archaea and Bacteria; Whitman, W.B., Ed.; John Wiley & Sons: New York, NY, USA, 2015; pp. 1–20. [Google Scholar]
- Hungate, R.E. The Rumen and its Microbes; Academic Press: New York, NY, USA, 1966. [Google Scholar]
- Evans, J.D.; Martin, S.A. Factors affecting lactate and malate utilization by Selenomonas ruminantium. Appl. Environ. Microbiol. 1997, 63, 4853–4858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgavi, D.P.; Sakurada, M.; Tomita, Y.; Onodera, R. Presence in rumen bacterial and protozoal populations of enzymes capable of degrading fungal cell walls. Microbiology 1994, 140, 631–636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, A.G.; Lloyd, D. Biological activities of symbiotic and parasitic protozoa and fungi in low-oxygen environments. Adv. Microb. Ecol. 1993, 13, 211–262. [Google Scholar]
- Yang, X.; Guo, J.L.; Sun, J.Y. Biodegradation of free-gossypol by a new fungus isolated from cotton planted soil. Afr. J. Microbiol. Res. 2011, 5, 3066–3072. [Google Scholar]



{kind=link}
{kind=link}
| Item 1 | HF | LF |
|---|---|---|
| Crude protein | 7.5 | 8.0 |
| Neutral detergent fiber | 45.0 | 33.0 |
| Acid detergent fiber | 25.6 | 18.4 |
| Ether extract | 3.5 | 4.0 |
| Starch | 28.4 | 41.2 |
| Target Groups | Sequence 5′–3′ | Product Size (bp) | References |
|---|---|---|---|
| F. succinogenes | F: GTTCGGAATTACTGGGCGTAAAR: CGCCTGCCCCTGAACTATC | 121 | [24] |
| R. flavefaciens | F:CGAACGGAGATAATTTGAGTTTACTTAGGR: CGGTCTCTGTATGTTATGAGGTATTACC | 132 | [24] |
| R. albus | F: CCCTAAAAGCAGTCTTAGTTCGR: CCTCCTTGCGGTTAGAACA | 175 | [25] |
| B. fibrisolvens | F:TAACATGAGAGTTTGATCCTGGCTCR:CGTTACTCACCCGTCCGC | 136 | [26] |
| P. ruminicola | F:GAAAGTCGGATTAATGCTCTATGTTGR:CATCCTATAGCGGTAAACCTTTGG | 74 | [27] |
| S. ruminantium | F:CAATAAGCATTCCGCCTGGGR:TTCACTCAATGTCAAGCCCTGG | 138 | [27] |
| Protozoa | F:GCTTTCGWTGGTAGTGTATTR:CTTGCCCTCYAATCGTWCT | 223 | [28] |
| Fungi | F:GAGGAAGTAAAAGTCGTAACAAGGTTTCR:CAAATTCACAAAGGGTAGGATGATT | 120 | [24] |
| Total bacteria | F:CGGCAACGAGCGCAACCCR:CCATTGTAGCACGTGTGTAGCC | 146 | [25] |
| Item 1 | IVDMD | GP48 | Gas Production Kinetic | Gas Composition (%) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| A | B | C | AGPR | CO2 | CH4 | H2 | ||||
| Ration | HF 2 | 53.5 b | 130.5 | 147.8 | 1.1 | 4.7 | 10.4 a | 83.0 a | 15.6 | 0.2 |
| LF 2 | 65.0 a | 118.6 | 131.7 | 1.1 | 4.7 | 8.0 b | 82.2 b | 16.3 | 0.2 | |
| SEM 3 | 0.50 | 4.17 | 7.94 | 0.03 | 0.56 | 0.78 | 0.20 | 0.20 | 0.01 | |
| Gossypol Dose (mg/g) | 0 | 60.6 | 140.5 a | 156.3 a | 1.1 ab | 5.1 | 9.2 | 83.4 a | 15.4 b | 0.20 a |
| 0.25 | 59.4 | 128.2 ab | 152.8 a | 1.0 b | 5.4 | 9.0 | 83.0 a | 16.0 ab | 0.18 ab | |
| 0.5 | 58.9 | 118.8 bc | 136.1 ab | 1.0 b | 4.9 | 8.0 | 82.0 b | 16.7 a | 0.16 b | |
| 0.75 | 58.6 | 111.8 c | 116.3 b | 1.2 a | 3.6 | 10.4 | 81.6 b | 15.8 b | 0.15 b | |
| SEM 3 | 0.70 | 4.62 | 9.71 | 0.03 | 0.75 | 1.17 | 0.30 | 0.20 | 0.01 | |
| p-Value | Ration | <0.01 | 0.02 | 0.17 | 0.13 | 0.96 | 0.04 | 0.04 | 0.06 | 0.81 |
| Dose | 0.50 | <0.01 | 0.05 | 0.02 | 0.39 | 0.36 | <0.01 | 0.02 | 0.01 | |
| I 4 | 0.90 | 0.58 | 0.75 | 0.28 | 0.56 | 0.27 | 0.90 | 0.04 | 0.80 | |
| L 4 | 0.63 | <0.01 | <0.01 | 0.08 | 0.17 | 0.43 | <0.01 | 0.18 | <0.01 | |
| Q 4 | 0.18 | 0.54 | 0.44 | 0.01 | 0.32 | 0.19 | 0.90 | <0.01 | 0.40 | |
| Item 1 | Ration | SEM 3 | Gossypol (mg/g) | SEM 3 | p-Value | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| HF 2 | LF 2 | 0 | 0.25 | 0.5 | 0.75 | Ration | Dose | I 4 | L 4 | Q 4 | |||
| pH | 6.68 a | 6.61 b | 0.01 | 6.62 b | 6.64 ab | 6.66 ab | 6.67 a | 0.02 | <0.01 | 0.09 | 0.99 | 0.01 | 0.87 |
| NH3-N (mg/dL) | 40.6 a | 37.3 b | 0.31 | 39.3 | 37.9 | 39.0 | 39.2 | 0.40 | <0.01 | 0.22 | 0.05 | 0.91 | 0.07 |
| MCP (μg/mL) | 91.5 | 91.0 | 2.25 | 105.4 a | 91.1 b | 85.5 b | 83.0 b | 3.19 | 0.89 | <0.01 | 0.68 | <0.01 | 0.02 |
| tVFA (mM) | 93.1 a | 85.9 b | 1.09 | 89.6 ab | 90.8 a | 91.8 a | 85.8 b | 1.55 | <0.01 | 0.06 | 0.53 | 0.15 | 0.03 |
| Ace (%) | 51.9 a | 47.2 b | 0.60 | 50.5 a | 49.9 a | 51.1 a | 46.9 b | 0.82 | <0.01 | 0.01 | 0.15 | 0.01 | 0.03 |
| Pro (%) | 18.7 b | 19.7 a | 0.23 | 18.9 b | 20.3 a | 19.3 b | 18.5 b | 0.33 | 0.01 | <0.01 | 0.23 | 0.10 | <0.01 |
| But (%) | 13.0 | 13.0 | 0.16 | 12.4 b | 13.2 a | 13.5 a | 12.9 ab | 0.22 | 0.97 | 0.01 | 0.17 | 0.06 | <0.01 |
| Val (%) | 2.1 | 2.0 | 0.03 | 2.0 | 2.1 | 2.1 | 2.1 | 0.05 | 0.14 | 0.93 | 0.09 | 0.55 | 0.83 |
| BCVFA (%) | 6.0 a | 5.3 b | 0.12 | 6.0 a | 5.0 b | 6.1 a | 5.6 a | 0.16 | <0.01 | <0.01 | 0.03 | 0.96 | 0.13 |
| NGR | 3.6 | 3.6 | 0.03 | 3.6 b | 3.5 b | 3.8 a | 3.6 b | 0.04 | 0.62 | <0.01 | 0.77 | 0.21 | 0.58 |
| FE | 0.75 | 0.78 | 0.01 | 0.79 | 0.76 | 0.75 | 0.76 | 0.012 | 0.14 | 0.31 | 0.31 | 0.18 | 0.34 |
| DG (%) | 97.6 a | 95.9 b | 0.12 | - | 96.8 b | 97.7 a | 95.8 c | 0.15 | <0.01 | <0.01 | 0.66 | <0.01 | <0.01 |
| Item | Ration | SEM 2 | Gossypol (mg/g) | SEM 2 | p-Value | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| HF 1 | LF 1 | 0 | 0.25 | 0.5 | 0.75 | Ration | Dose | I 3 | L 3 | Q 3 | |||
| F. succinogenes | 6.75 a | 6.50 b | 0.025 | 7.15 a | 6.34 d | 6.48 c | 6.61 b | 0.034 | <0.01 | <0.01 | 0.05 | <0.01 | <0.01 |
| R. flavefaciens | 6.78 | 6.68 | 0.080 | 6.67 b | 6.73 b | 6.41 b | 7.11 a | 0.110 | 0.41 | <0.01 | 0.99 | 0.08 | 0.02 |
| R. albus | 6.32 | 6.25 | 0.021 | 6.42 a | 6.28 b | 6.24 b | 6.21 b | 0.026 | 0.07 | <0.01 | 0.82 | <0.01 | 0.11 |
| B. fibrisolvens | 8.50 b | 8.58 a | 0.016 | 8.72 a | 8.54 b | 8.49 b | 8.40 c | 0.022 | <0.01 | <0.01 | 0.60 | <0.01 | 0.11 |
| P. ruminicola | 7.18 b | 7.26 a | 0.022 | 7.40 a | 7.01 d | 7.25 b | 7.14 c | 0.028 | 0.03 | <0.01 | 0.02 | <0.01 | <0.01 |
| S. ruminantium | 6.09 b | 6.20 a | 0.028 | 6.23 a | 6.05 b | 6.15 ab | 6.15 ab | 0.034 | 0.01 | 0.06 | 0.62 | 0.51 | 0.06 |
| Protozoa | 4.97 a | 4.89 b | 0.016 | 4.76 c | 4.87 b | 4.94 b | 5.15 a | 0.022 | <0.01 | <0.01 | 0.82 | <0.01 | 0.07 |
| Fungi | 3.36 a | 3.21 b | 0.016 | 3.45 a | 3.35 b | 3.26 c | 3.09 d | 0.023 | <0.01 | <0.01 | 0.31 | <0.01 | 0.14 |
| Total bacteria | 9.82 b | 9.93 a | 0.035 | 9.78 b | 9.83 b | 9.86 b | 10.04 a | 0.049 | <0.01 | <0.01 | <0.01 | <0.01 | 0.04 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, W.-K.; Wang, Y.-L.; Li, W.-J.; Wu, Q.-C.; Li, S.-L.; Yang, H.-J. Gossypol Exhibited Higher Detrimental Effect on Ruminal Fermentation Characteristics of Low-Forage in Comparison with High-Forage Mixed Feeds. Toxics 2021, 9, 51. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics9030051
Wang W-K, Wang Y-L, Li W-J, Wu Q-C, Li S-L, Yang H-J. Gossypol Exhibited Higher Detrimental Effect on Ruminal Fermentation Characteristics of Low-Forage in Comparison with High-Forage Mixed Feeds. Toxics. 2021; 9(3):51. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics9030051
Chicago/Turabian StyleWang, Wei-Kang, Yan-Lu Wang, Wen-Juan Li, Qi-Chao Wu, Sheng-Li Li, and Hong-Jian Yang. 2021. "Gossypol Exhibited Higher Detrimental Effect on Ruminal Fermentation Characteristics of Low-Forage in Comparison with High-Forage Mixed Feeds" Toxics 9, no. 3: 51. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics9030051