
Blood Toxic Elements and Effects on Plasma Vitamins and Carotenoids in Two Wild Bird Species: Turdus merula and Columba livia

 , , , ,
, , , ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
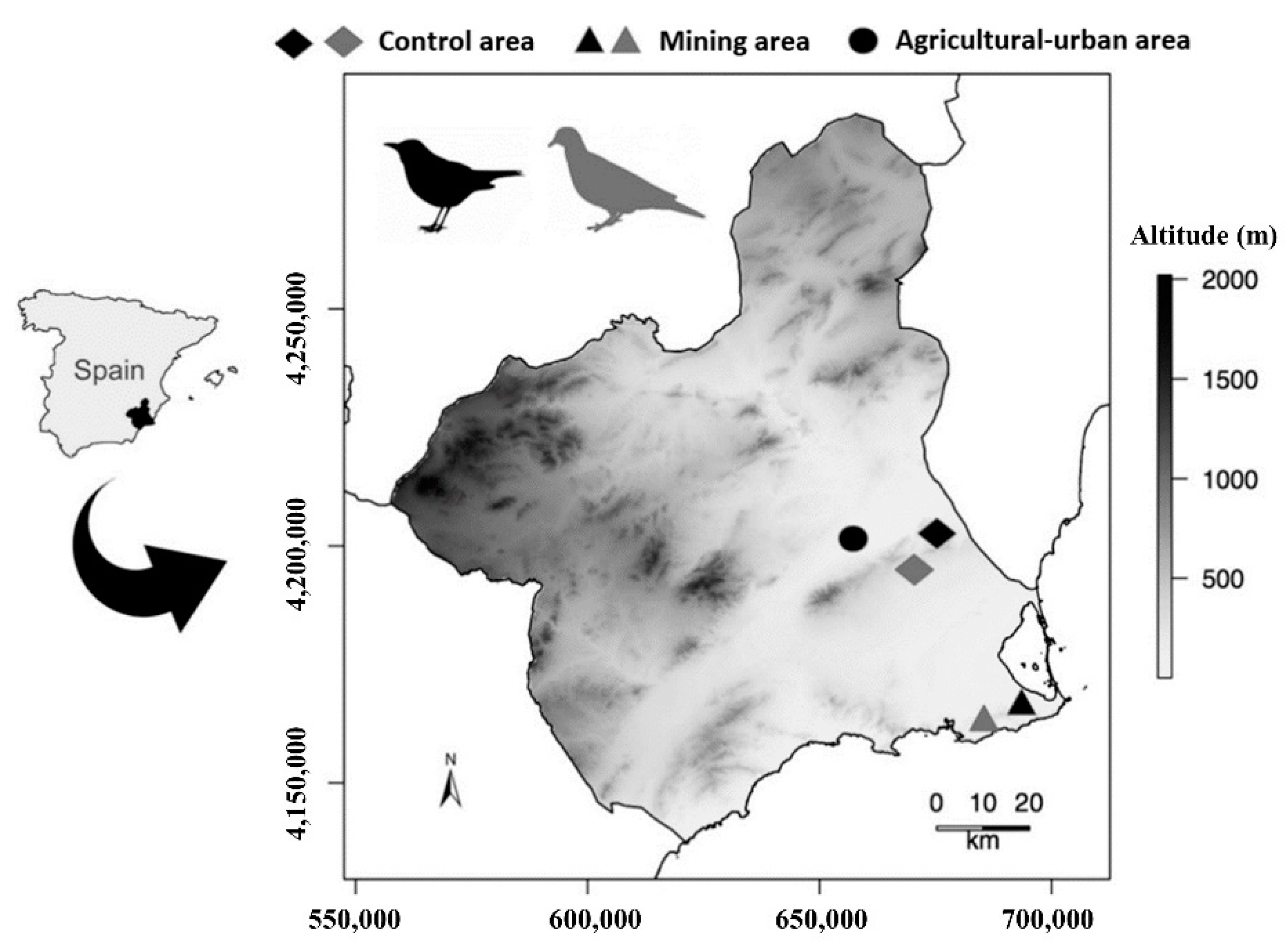
2.1. Study Area and Pollution Scenarios
2.2. Bird Sampling, Measurements, and Trace Element and Vitamin Analysis
2.3. Statistical Procedures
3. Results
4. Discussion
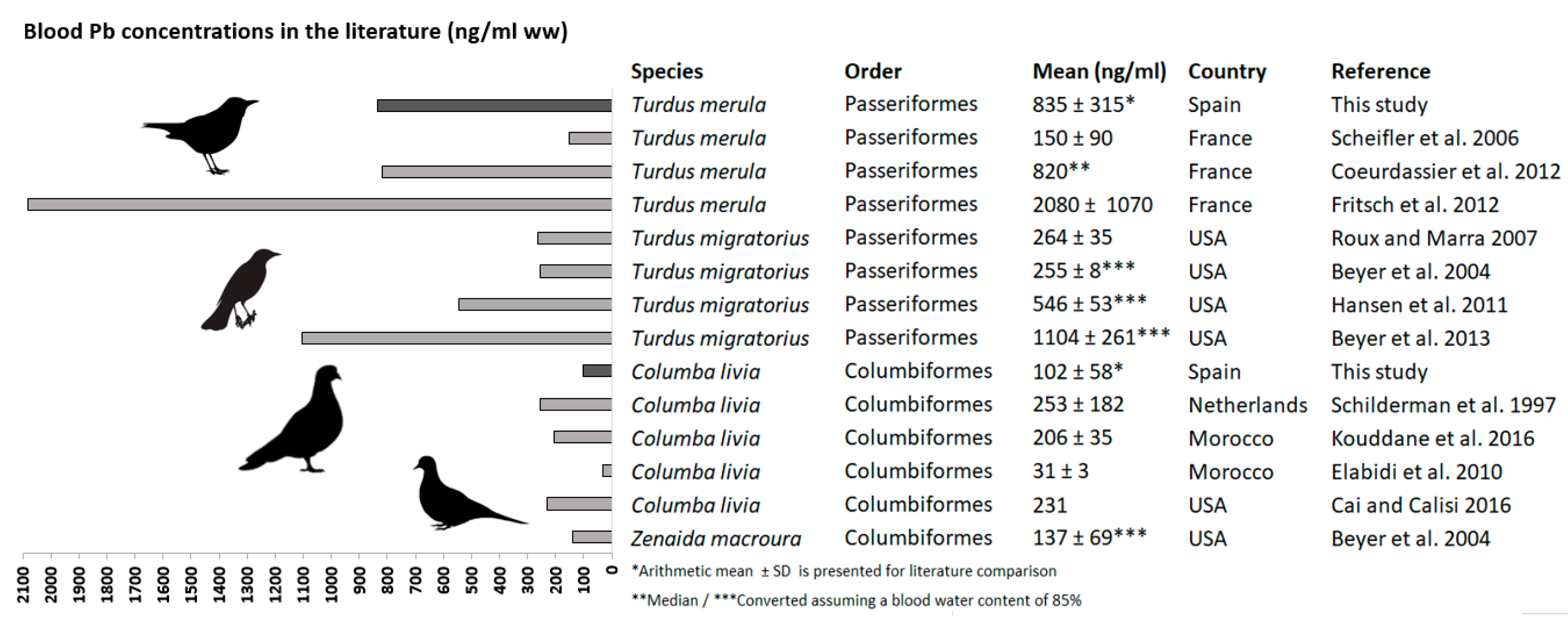
4.1. Element Concentrations in Blood
4.2. Element-Driven Effects on Lutein and Vitamin Levels, as well as Hematocrit and Body Measurements
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| As | Ba | Cd | Co | Cr | Cu | Fe | Hg | Mn | Mo | Pb | Se | Sr | V | Zn | HT | Ret | Toc | Lut | Mass | Wing | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| As | −0.062 | 0.69 *** | 0.65 *** | 0.18 | 0.27 | 0.27 | 0.42 ** | 0.062 | −0.20 | 0.54 *** | 0.47 ** | −0.19 | N/A | 0.11 | 0.028 | −0.54 *** | −0.22 | −0.10 | 0.25 | 0.29 | |
| Ba | N/A | −0.072 | 0.056 | 0.13 | 0.086 | −0.26 | −0.28 | 0.46 ** | −0.13 | 0.12 | −0.12 | 0.39 * | N/A | 0.069 | −0.29 | 0.28 | −0.13 | −0.17 | −0.14 | −0.15 | |
| Cd | 0.42 * | N/A | 0.50 *** | 0.26 | 0.031 | 0.0035 | 0.25 | 0.23 | −0.34 * | 0.71 *** | 0.20 | −0.34 * | N/A | −0.0024 | −0.011 | −0.48 ** | −0.25 | −0.18 | 0.36 * | 0.25 | |
| Co | N/A | N/A | N/A | 0.41 ** | 0.18 | 0.013 | 0.39 * | 0.25 | −0.22 | 0.29 | 0.42 ** | −0.034 | N/A | 0.25 | −0.14 | −0.20 | −0.20 | 0.0017 | 0.19 | 0.15 | |
| Cr | N/A | N/A | N/A | N/A | 0.17 | 0.071 | 0.092 | 0.19 | −0.24 | 0.11 | 0.12 | −0.11 | N/A | 0.22 | −0.25 | −0.012 | −0.28 | 0.040 | 0.14 | 0.21 | |
| Cu | 0.30 | N/A | 0.29 | N/A | N/A | 0.30 | 0.16 | 0.052 | 0.010 | 0.23 | −0.056 | 0.11 | N/A | 0.22 | 0.11 | −0.22 | 0.036 | −0.022 | 0.018 | −0.041 | |
| Fe | 0.37 | N/A | 0.42 * | N/A | N/A | 0.63 *** | 0.16 | −0.23 | −0.21 | −0.090 | 0.26 | −0.14 | N/A | 0.30 | 0.58 *** | −0.059 | 0.28 | 0.31 * | −0.063 | −0.012 | |
| Hg | N/A | N/A | N/A | N/A | N/A | N/A | N/A | −0.21 | 0.41 ** | −0.15 | 0.62 *** | 0.17 | N/A | 0.084 | 0.29 | −0.17 | 0.12 | 0.30 | 0.12 | −0.17 | |
| Mn | 0.16 | N/A | 0.21 | N/A | N/A | 0.34 | 0.28 | N/A | −0.15 | 0.44 ** | −0.30 | −0.075 | N/A | 0.20 | −0.25 | −0.063 | −0.21 | −0.33 * | 0.11 | −0.13 | |
| Mo | 0.38 | N/A | 0.21 | N/A | N/A | 0.50 ** | 0.53 ** | N/A | 0.35 | −0.43 ** | 0.26 | 0.39 * | N/A | 0.061 | 0.16 | 0.10 | 0.25 | 0.28 | −0.22 | −0.29 | |
| Pb | 0.56 ** | N/A | 0.39 * | N/A | N/A | 0.17 | 0.46 * | N/A | 0.16 | 0.48 * | −0.27 | −0.51 *** | N/A | 0.13 | −0.064 | −0.47 ** | −0.25 | −0.34 * | 0.26 | 0.24 | |
| Se | −0.054 | N/A | 0.17 | N/A | N/A | 0.37 | 0.49 ** | N/A | 0.14 | 0.40 * | 0.39 * | 0.27 | N/A | 0.18 | 0.062 | −0.0013 | 0.046 | 0.28 | 0.12 | 0.18 | |
| Sr | −0.034 | N/A | −0.044 | N/A | N/A | 0.58 ** | 0.36 | N/A | 0.50 ** | 0.42 * | −0.14 | −0.029 | N/A | −0.0032 | −0.088 | 0.13 | −0.013 | −0.048 | −0.16 | −0.24 | |
| V | 0.35 | N/A | 0.28 | N/A | N/A | 0.19 | 0.37 | N/A | 0.075 | 0.56 ** | 0.54 ** | 0.29 | 0.14 | N/A | N/A | N/A | N/A | N/A | N/A | N/A | |
| Zn | 0.19 | N/A | 0.41 * | N/A | N/A | 0.61 *** | 0.86 *** | N/A | 0.36 | 0.59 ** | 0.42 * | 0.51 ** | 0.42 * | 0.50 ** | 0.19 | 0.17 | 0.25 | 0.22 | −0.13 | 0.054 | |
| HT | 0.20 | N/A | 0.55 * | N/A | N/A | 0.24 | 0.65 ** | N/A | 0.19 | 0.088 | 0.22 | 0.47 * | −0.22 | 0.17 | 0.61 ** | −0.048 | 0.29 | 0.21 | −0.034 | −0.23 | |
| Ret | −0.29 | N/A | 0.23 | N/A | N/A | −0.094 | 0.20 | N/A | −0.24 | −0.17 | 0.038 | 0.26 | −0.17 | 0.062 | 0.26 | 0.61 ** | 0.47 ** | 0.39 * | −0.26 | −0.20 | |
| Toc | 0.42 * | N/A | 0.0023 | N/A | N/A | 0.26 | 0.29 | N/A | 0.35 | 0.21 | 0.013 | −0.25 | 0.19 | −0.030 | 0.10 | 0.26 | −0.13 | 0.68 *** | −0.50 ** | −0.15 | |
| Lut | 0.16 | N/A | −0.14 | N/A | N/A | −0.032 | 0.15 | N/A | −0.027 | −0.0090 | −0.030 | −0.20 | 0.023 | 0.027 | 0.15 | 0.10 | 0.31 | 0.54 ** | −0.43 ** | −0.22 | |
| Mass | 0.12 | N/A | 0.52 ** | N/A | N/A | −0.025 | 0.32 | N/A | 0.082 | −0.073 | 0.12 | 0.12 | −0.063 | 0.26 | 0.41 * | 0.15 | 0.28 | −0.072 | 0.025 | 0.20 | |
| Wing | 0.033 | N/A | 0.30 | N/A | N/A | −0.27 | 0.13 | N/A | −0.18 | −0.32 | 0.063 | 0.12 | −0.39 * | 0.17 | 0.16 | 0.41 * | 0.40 * | −0.24 | 0.098 | 0.68 *** |
References
- Ratcliffe, D.A. Changes attributable to pesticides in egg breakage frequency and eggshell thickness in some British birds. J. Appl. Ecol. 1970, 7, 67–115. [Google Scholar] [CrossRef]
- Sánchez-Virosta, P.; Espín, S.; García-Fernández, A.J.; Eeva, T. A Review on Exposure and Effects of Arsenic in Passerine Birds. Sci. Total Environ. 2015, 512, 506–525. [Google Scholar] [CrossRef]
- Burger, J.; Gochfeld, M. Risk, Mercury Levels, and Birds: Relating Adverse Laboratory Effects to Field Biomonitoring. Environ. Res. 1997, 75, 160–172. [Google Scholar] [CrossRef]
- Pain, D.J.; Mateo, R.; Green, R.E. Effects of Lead from Ammunition on Birds and Other Wildlife: A Review and Update. Ambio 2019, 48, 935–953. [Google Scholar] [CrossRef] [Green Version]
- Espín, S.; Sánchez-Virosta, P. A Review of Metal-Induced Effects on Vitamins A, E and D3 in Birds. Ecotoxicology 2020, 30, 1–16. [Google Scholar] [CrossRef]
- ATSDR. The ATSDR 2019 Substance Priority List; Agency for Toxic Substances and Disease Registry, U.S. Public Health Service: Washington, DC, USA, 2019.
- Eeva, T.; Hasselquist, D.; Langefors, Å.; Tummeleht, L.; Nikinmaa, M.; Ilmonen, P. Pollution Related Effects in Immune Function and Stress in a Free-Living Population of Pied Flycatcher Ficedula hypoleuca. J. Avian Biol. 2005, 36, 405–412. [Google Scholar] [CrossRef] [Green Version]
- Espín, S.; Ruiz, S.; Sánchez-Virosta, P.; Salminen, J.-P.; Eeva, T. Effects of Experimental Calcium Availability and Anthropogenic Metal Pollution on Eggshell Characteristics and Yolk Carotenoid and Vitamin Levels in Two Passerine Birds. Chemosphere 2016, 151, 189–201. [Google Scholar] [CrossRef] [PubMed]
- Espín, S.; Martínez-López, E.; Jiménez, P.; María-Mojica, P.; García-Fernández, A.J. Interspecific Differences in the Antioxidant Capacity of Two Laridae Species Exposed to Metals. Environ. Res. 2016, 147, 115–124. [Google Scholar] [CrossRef]
- Vallverdú-Coll, N.; Mateo, R.; Mougeot, F.; Ortiz-Santaliestra, M.E. Immunotoxic Effects of Lead on Birds. Sci. Total Environ. 2019, 689, 505–515. [Google Scholar] [CrossRef]
- Whitney, M.C.; Cristol, D.A. Impacts of Sublethal Mercury Exposure on Birds: A Detailed Review. Rev. Environ. Contam. Toxicol. 2018, 244, 113–163. [Google Scholar] [CrossRef]
- Ruiz, S.; Espín, S.; Rainio, M.; Ruuskanen, S.; Salminen, J.-P.; Lilley, T.M.; Eeva, T. Effects of Dietary Lead Exposure on Vitamin Levels in Great Tit Nestlings—An Experimental Manipulation. Environ. Pollut. 2016, 213, 688–697. [Google Scholar] [CrossRef]
- Sánchez-Virosta, P.; Espín, S.; Ruiz, S.; Salminen, J.-P.; García-Fernández, A.J.; Eeva, T. Experimental Manipulation of Dietary Arsenic Levels in Great Tit Nestlings: Accumulation Pattern and Effects on Growth, Survival and Plasma Biochemistry. Environ. Pollut. 2018, 233, 764–773. [Google Scholar] [CrossRef]
- Hussain, M.; Mumtaz, S. E-Waste: Impacts, Issues and Management Strategies. Rev. Environ. Health 2014, 29, 53–58. [Google Scholar] [CrossRef]
- Tansel, B. From Electronic Consumer Products to E-Wastes: Global Outlook, Waste Quantities, Recycling Challenges. Environ. Int. 2017, 98, 35–45. [Google Scholar] [CrossRef]
- Sánchez-Virosta, P.; León-Ortega, M.; Calvo, J.F.; Camarero, P.R.; Mateo, R.; Zumbado, M.; Luzardo, O.P.; Eeva, T.; García-Fernández, A.J.; Espín, S. Blood Concentrations of 50 Elements in Eagle Owl (Bubo bubo) at Different Contamination Scenarios and Related Effects on Plasma Vitamin Levels. Environ. Pollut. 2020, 265, 115012. [Google Scholar] [CrossRef] [PubMed]
- Espín, S.; Sánchez-Virosta, P.; Zamora-Marín, J.M.; León-Ortega, M.; Jiménez, P.; Zumbado, M.; Luzardo, O.P.; Eeva, T.; García-Fernández, A.J. Toxic Elements in Blood of Red-Necked Nightjars (Caprimulgus ruficollis) Inhabiting Differently Polluted Environments. Environ. Pollut. 2020, 262, 114334. [Google Scholar] [CrossRef] [PubMed]
- Espín, S.; Sánchez-Virosta, P.; Zamora-Marín, J.M.; León-Ortega, M.; Jiménez, P.; Zamora-López, A.; Camarero, P.R.; Mateo, R.; Eeva, T.; García-Fernández, A.J. Physiological Effects of Toxic Elements on a Wild Nightjar Species. Environ. Pollut. 2020, 263, 114568. [Google Scholar] [CrossRef]
- Britton, G. Structure and Properties of Carotenoids in Relation to Function. FASEB J. 1995, 9, 1551–1558. [Google Scholar] [CrossRef] [PubMed]
- Costantini, D.; Møller, A.P. Carotenoids Are Minor Antioxidants for Birds. Funct. Ecol. 2008, 22, 367–370. [Google Scholar] [CrossRef]
- Koch, R.E.; Kavazis, A.N.; Hasselquist, D.; Hood, W.R.; Zhang, Y.; Toomey, M.B.; Hill, G.E. No Evidence That Carotenoid Pigments Boost Either Immune or Antioxidant Defenses in a Songbird. Nat. Commun. 2018, 9, 491. [Google Scholar] [CrossRef]
- Tanumihardjo, S.A. Vitamin A: Biomarkers of Nutrition for Development. Am. J. Clin. Nutr. 2011, 94, 658S–665S. [Google Scholar] [CrossRef] [Green Version]
- Zile, M.H. Function of Vitamin A in Vertebrate Embryonic Development. J. Nutr. 2001, 131, 705–708. [Google Scholar] [CrossRef] [Green Version]
- Traber, M.G.; Atkinson, J. Vitamin E, Antioxidant and Nothing More. Free Radic. Biol. Med. 2007, 43, 4–15. [Google Scholar] [CrossRef] [Green Version]
- Rizvi, S.; Raza, S.T.; Ahmed, F.; Ahmad, A.; Abbas, S.; Mahdi, F. The Role of Vitamin E in Human Health and Some Diseases. Sultan Qaboos Univ. Med. J. 2014, 14, e157–e165. [Google Scholar]
- Evans, K.L.; Hatchwell, B.J.; Parnell, M.; Gaston, K.J. A Conceptual Framework for the Colonisation of Urban Areas: The Blackbird Turdus merula as a Case Study. Biol. Rev. 2010, 85, 643–667. [Google Scholar] [CrossRef]
- Hatchwell, B.J.; Chamberlain, D.E.; Perrins, C.M. The Reproductive Success of Blackbirds Turdus merula in Relation to Habitat Structure and Choice of Nest Site. Ibis 2008, 138, 256–262. [Google Scholar] [CrossRef]
- Aparicio, R.J. Mirlo Común—Turdus merula. In Enciclopedia Virtual de los Vertebrados Españoles; Salvador, A., Morales, M.B., Eds.; Museo Nacional de Ciencias Naturales CSIC: Madrid, Spain, 2016. [Google Scholar]
- Chamberlain, D.E.; Hatchwell, B.J.; Perrins, C.M. Importance of Feeding Ecology to the Reproductive Success of Blackbirds Turdus merula Nesting in Rural Habitats. Ibis 1999, 141, 415–427. [Google Scholar] [CrossRef]
- Hernández, A. Blackcaps Sylvia atricapilla and Blackbirds Turdus merula Feeding Their Nestlings and Fledglings on Fleshly Fruit. Folia Zool. 2005, 54, 379–388. [Google Scholar]
- Scheifler, R.; Coeurdassier, M.; Morilhat, C.; Bernard, N.; Faivre, B.; Flicoteaux, P.; Giraudoux, P.; Noël, M.; Piotte, P.; Rieffel, D.; et al. Lead Concentrations in Feathers and Blood of Common Blackbirds (Turdus merula) and in Earthworms Inhabiting Unpolluted and Moderately Polluted Urban Areas. Sci. Total Environ. 2006, 371, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Fritsch, C.; Coeurdassier, M.; Faivre, B.; Baurand, P.E.; Giraudoux, P.; van den Brink, N.W.; Scheifler, R. Influence of Landscape Composition and Diversity on Contaminant Flux in Terrestrial Food Webs: A Case Study of Trace Metal Transfer to European Blackbirds Turdus merula. Sci. Total Environ. 2012, 432, 275–287. [Google Scholar] [CrossRef]
- Stukenholtz, E.; Hailu, T.; Childers, S.; Leatherwood, C.; Evans, L.; Roulain, D.; Townsley, D.; Treider, M.; Neal Platt II, R.; Ray, D.; et al. Ecology of Feral Pigeons: Population Monitoring, Resource Selection, and Management Practices. In Wildlife Population Monitoring; IntechOpen: Dallas, TX, USA, 2019. [Google Scholar]
- Spennemann, D.H.R.; Watson, M.J. Dietary Habits of Urban Pigeons (Columba livia) and Implications of Excreta PH—A Review. Eur. J. Ecol. 2017, 3, 27–41. [Google Scholar] [CrossRef] [Green Version]
- Antonio García, M.T.; Martinez-Conde, E.; Corpas Vazquez, I. Lead Levels of Feral Pigeons (Columba livia) from Madrid (Spain). Environ. Pollut. 1988, 54, 89–96. [Google Scholar] [CrossRef]
- Nam, D.-H.; Lee, D.-P. Monitoring for Pb and Cd Pollution Using Feral Pigeons in Rural, Urban, and Industrial Environments of Korea. Sci. Total Environ. 2006, 357, 288–295. [Google Scholar] [CrossRef]
- Valladares-Faundez, P.; Cáceres Tapia, G.; Valdés Saavedra, J. Content of lead, cadmium and arsenic in biological tissues of the feral pigeons (Columba livia) present in an urban area previously contaminated with mining residues. Rev. Int. Contam. Ambient. 2020, 36, 485–490. [Google Scholar] [CrossRef]
- Espín, S.; Martínez-López, E.; León-Ortega, M.; Martínez, J.E.; García-Fernández, A.J. Oxidative Stress Biomarkers in Eurasian Eagle Owls (Bubo bubo) in Three Different Scenarios of Heavy Metal Exposure. Environ. Res. 2014, 131, 134–144. [Google Scholar] [CrossRef]
- Conesa, H.M.; Schulin, R. The Cartagena–La Unión Mining District (SE Spain): A Review of Environmental Problems and Emerging Phytoremediation Solutions after Fifteen Years Research. J. Environ. Monit. 2010, 12, 1225–1233. [Google Scholar] [CrossRef]
- García-Fernández, A.J.; Sánchez-García, J.A.; Jiménez-Montalbán, P.; Luna, A. Lead and Cadmium in Wild Birds in Southeastern Spain. Environ. Toxicol. Chem. 1995, 14, 2049–2058. [Google Scholar] [CrossRef]
- Espín, S.; Martínez-López, E.; León-Ortega, M.; Calvo, J.F.; García-Fernández, A.J. Factors That Influence Mercury Concentrations in Nestling Eagle Owls (Bubo bubo). Sci. Total Environ. 2014, 470–471, 1132–1139. [Google Scholar] [CrossRef] [PubMed]
- Demongin, L. (Ed.) Identification Guide to Birds in the Hand; SEPEC: Paris, France, 2016.
- Svensson, L. Identification Guide to European Passerines; British Trust for Ornithology: Thelford, UK, 1996; ISBN 978-91-630-1118-4. [Google Scholar]
- Jackson, H.D. A Field Survey to Investigate Why Nightjars Frequent Roads at Night. Ostrich 2003, 74, 97–101. [Google Scholar] [CrossRef]
- Espín, S.; Andevski, J.; Duke, G.; Eulaers, I.; Gómez-Ramírez, P.; Hallgrimsson, G.T.; Helander, B.; Herzke, D.; Jaspers, V.L.B.; Krone, O.; et al. A Schematic Sampling Protocol for Contaminant Monitoring in Raptors. Ambio 2021, 50, 95–100. [Google Scholar] [CrossRef]
- González-Antuña, A.; Camacho, M.; Henríquez-Hernández, L.A.; Boada, L.D.; Almeida-González, M.; Zumbado, M.; Luzardo, O.P. Simultaneous Quantification of 49 Elements Associated to E-Waste in Human Blood by ICP-MS for Routine Analysis. MethodsX 2017, 4, 328–334. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Estival, J.; Martínez-Haro, M.; Martín-Hernando, M.A.P.; Mateo, R. Sub-Chronic Effects of Nitrate in Drinking Water on Red-Legged Partridge (Alectoris rufa): Oxidative Stress and T-Cell Mediated Immune Function. Environ. Res. 2010, 110, 469–475. [Google Scholar] [CrossRef]
- Croghan, C.W.; Egeghy, P.P. Methods of Dealing with Values below the Limit of Detection Using SAS; US Environmental Protection Agency: St. Petersburg, FL, USA, 2003.
- Conesa, H.; Schulin, R.; Nowack, B. Mining Landscape: A Cultural Tourist Opportunity or an Environmental Problem?: The Study Case of the Cartagena–La Unión Mining District (SE Spain). Ecol. Econ. 2008, 64, 690–700. [Google Scholar] [CrossRef]
- Pavetti, F.G.; Conesa-Alcaraz, M.; Faz-Cano, A.; Arnaldos, R.; García, G. Contaminación Por Metales Pesados En La Rambla de El Gorguel (SE Península Ibérica). Terra 2006, 24, 171–178. [Google Scholar]
- Gómez-Ramírez, P.; Martínez-López, E.; María-Mojica, P.; León-Ortega, M.; Garcia-Fernandez, A.J. Blood lead levels and δ-ALAD inhibition in nestlings of Eurasian Eagle Owl (Bubo bubo) to assess lead exposure associated to an abandoned mining area. Ecotoxicology 2011, 20, 131–138. [Google Scholar] [CrossRef]
- García-Fernández, A.J.; Motas-Guzmán, M.; Navas, I.; María-Mojica, P.; Luna, A.; Sánchez-García, J.A. Environmental Exposure and Distribution of Lead in Four Species of Raptors in Southeastern Spain. Arch. Environ. Contam. Toxicol. 1997, 33, 76–82. [Google Scholar] [CrossRef]
- Coeurdassier, M.; Fritsch, C.; Faivre, B.; Crini, N.; Scheifler, R. Partitioning of Cd and Pb in the Blood of European Blackbirds (Turdus merula) from a Smelter Contaminated Site and Use for Biomonitoring. Chemosphere 2012, 87, 1368–1373. [Google Scholar] [CrossRef]
- Beyer, W.N.; Franson, J.C.; French, J.B.; May, T.; Rattner, B.A.; Shearn-Bochsler, V.I.; Warner, S.E.; Weber, J.; Mosby, D. Toxic Exposure of Songbirds to Lead in the Southeast Missouri Lead Mining District. Arch. Environ. Contam. Toxicol. 2013, 65, 598–610. [Google Scholar] [CrossRef] [Green Version]
- Roux, K.E.; Marra, P.P. The Presence and Impact of Environmental Lead in Passerine Birds along an Urban to Rural Land Use Gradient. Arch. Environ. Contam. Toxicol. 2007, 53, 261–268. [Google Scholar] [CrossRef]
- Beyer, W.N.; Dalgarn, J.; Dudding, S.; French, J.B.; Mateo, R.; Miesner, J.; Sileo, L.; Spann, J. Zinc and Lead Poisoning in Wild Birds in the Tri-State Mining District (Oklahoma, Kansas, and Missouri). Arch. Environ. Contam. Toxicol. 2004, 48, 108–117. [Google Scholar] [CrossRef]
- Hansen, J.A.; Audet, D.; Spears, B.L.; Healy, K.A.; Brazzle, R.E.; Hoffman, D.J.; Dailey, A.; Beyer, W.N. Lead Exposure and Poisoning of Songbirds Using the Coeur d’Alene River Basin, Idaho, USA. Integr. Environ. Assess. Manag. 2011, 7, 587–595. [Google Scholar] [CrossRef] [PubMed]
- Cai, F.; Calisi, R.M. Seasons and Neighborhoods of High Lead Toxicity in New York City: The Feral Pigeon as a Bioindicator. Chemosphere 2016, 161, 274–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schilderman, P.A.E.L.; Hoogewerff, J.A.; van Schooten, F.-J.; Maas, L.M.; Moonen, E.J.C.; van Os, B.J.; van Wijnen, J.H.; Kleinjans, J.C.S. Possible Relevance of Pigeons as an Indicator Species for Monitoring Air Pollution. Environ. Health Perspect. 1997, 105, 322–330. [Google Scholar] [CrossRef]
- Kouddane, N.; Mouhir, L.; Fekhaoui, M.; Elabidi, A.; Benaakame, R. Monitoring Air Pollution at Mohammedia (Morocco): Pb, Cd and Zn in the Blood of Pigeons (Columba livia). Ecotoxicology 2016, 25, 720–726. [Google Scholar] [CrossRef]
- Tsipoura, N.; Burger, J.; Feltes, R.; Yacabucci, J.; Mizrahi, D.; Jeitner, C.; Gochfeld, M. Metal Concentrations in Three Species of Passerine Birds Breeding in the Hackensack Meadowlands of New Jersey. Environ. Res. 2008, 107, 218–228. [Google Scholar] [CrossRef] [PubMed]
- Gall, J.E.; Boyd, R.S.; Rajakaruna, N. Transfer of Heavy Metals through Terrestrial Food Webs: A Review. Environ. Monit. Assess. 2015, 187, 201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbasi, N.A.; Jaspers, V.L.B.; Chaudhry, M.J.I.; Ali, S.; Malik, R.N. Influence of Taxa, Trophic Level, and Location on Bioaccumulation of Toxic Metals in Bird’s Feathers: A Preliminary Biomonitoring Study Using Multiple Bird Species from Pakistan. Chemosphere 2015, 120, 527–537. [Google Scholar] [CrossRef]
- Ruiz-Chancho, M.J.; López-Sánchez, J.F.; Schmeisser, E.; Goessler, W.; Francesconi, K.A.; Rubio, R. Arsenic Speciation in Plants Growing in Arsenic-Contaminated Sites. Chemosphere 2008, 71, 1522–1530. [Google Scholar] [CrossRef]
- Martínez-Sánchez, M.J.; Martínez-López, S.; García-Lorenzo, M.L.; Martínez-Martínez, L.B.; Pérez-Sirvent, C. Evaluation of Arsenic in Soils and Plant Uptake Using Various Chemical Extraction Methods in Soils Affected by Old Mining Activities. Geoderma 2011, 160, 535–541. [Google Scholar] [CrossRef]
- Williams, P.N.; Price, A.H.; Raab, A.; Hossain, S.A.; Feldmann, J.; Meharg, A.A. Variation in Arsenic Speciation and Concentration in Paddy Rice Related to Dietary Exposure. Environ. Sci. Technol. 2005, 39, 5531–5540. [Google Scholar] [CrossRef]
- Alam, M.Z.; Hoque, M.A.; Ahammed, G.J.; McGee, R.; Carpenter-Boggs, L. Arsenic Accumulation in Lentil (Lens culinaris) Genotypes and Risk Associated with the Consumption of Grains. Sci. Rep. 2019, 9, 9431. [Google Scholar] [CrossRef] [Green Version]
- Vahter, M.; Marafante, E. Effects of Low Dietary Intake of Methionine, Choline or Proteins on the Biotransformation of Arsenite in the Rabbit. Toxicol. Lett. 1987, 37, 41–46. [Google Scholar] [CrossRef]
- Van Hemert, C.; Handel, C. Elements in Whole Blood of Northwestern Crows (Corvus caurinus) in Alaska, USA: No Evidence for an Association with Beak Deformities. J. Wildl. Dis. 2016, 52, 713–718. [Google Scholar] [CrossRef] [PubMed]
- Marie, P.J.; Hott, M.; Modrowski, D.; De Pollak, C.; Guillemain, J.; Deloffre, P.; Tsouderos, Y. An Uncoupling Agent Containing Strontium Prevents Bone Loss by Depressing Bone Resorption and Maintaining Bone Formation in Estrogen-Deficient Rats. J. Bone Miner. Res. 1993, 8, 607–615. [Google Scholar] [CrossRef] [PubMed]
- Maciejewska, K.; Drzazga, Z.; Kaszuba, M. Role of Trace Elements (Zn, Sr, Fe) in Bone Development: Energy Dispersive X-Ray Fluorescence Study of Rat Bone and Tooth Tissue. BioFactors 2014, 40, 425–435. [Google Scholar] [CrossRef]
- Sila-Asna, M.; Bunyaratvej, A.; Maeda, S.; Kitaguchi, H.; Bunyaratavej, N. Osteoblast differentiation and bone formation gene expression in strontium-inducing bone marrow mesenchymal stem cell. Kobe J. Med. Sci. 2007, 53, 25–35. [Google Scholar]
- PRTR Pollutant Release and Transfer Register. Registro Estatal de Emisiones y Fuentes Contaminantes; Ministerio para la Transición Ecológica y el Reto Demográfico: Madrid, Spain, 2021.
- Scheuhammer, A.M.; Basu, N.; Burgess, N.M.; Elliott, J.E.; Campbell, G.D.; Wayland, M.; Champoux, L.; Rodrigue, J. Relationships among Mercury, Selenium, and Neurochemical Parameters in Common Loons (Gavia immer) and Bald Eagles (Haliaeetus leucocephalus). Ecotoxicology 2008, 17, 93–101. [Google Scholar] [CrossRef]
- Cuvin-Aralar, M.L.; Furness, R.W. Mercury and Selenium Interaction: A Review. Ecotoxicol. Environ. Saf. 1991, 21, 348–364. [Google Scholar] [CrossRef]
- Franson, J.C.; Pain, D.J. Lead in birds. In Environmental Contaminants in Biota: Interpreting Tissue Concentrations; CRC Press Taylor & Francis: Boca Raton, FL, USA, 2011; pp. 563–593. [Google Scholar]
- Espín, S.; Martínez-López, E.; Jiménez, P.; María-Mojica, P.; García-Fernández, A.J. Delta-Aminolevulinic Acid Dehydratase (ΔALAD) Activity in Four Free-Living Bird Species Exposed to Different Levels of Lead under Natural Conditions. Environ. Res. 2015, 137, 185–198. [Google Scholar] [CrossRef]
- Espín, S.; Martínez-López, E.; Jiménez, P.; María-Mojica, P.; García-Fernández, A.J. Effects of Heavy Metals on Biomarkers for Oxidative Stress in Griffon Vulture (Gyps fulvus). Environ. Res. 2014, 129, 59–68. [Google Scholar] [CrossRef]
- Vallverdú-Coll, N.; Mougeot, F.; Ortiz-Santaliestra, M.E.; Castaño, C.; Santiago-Moreno, J.; Mateo, R. Effects of Lead Exposure on Sperm Quality and Reproductive Success in an Avian Model. Environ. Sci. Technol. 2016, 50, 12484–12492. [Google Scholar] [CrossRef]
- Defo, M.A.; Spear, P.A.; Couture, P. Consequences of Metal Exposure on Retinoid Metabolism in Vertebrates: A Review. Toxicol. Lett. 2014, 225, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Vallverdú-Coll, N.; Ortiz-Santaliestra, M.E.; Mougeot, F.; Vidal, D.; Mateo, R. Sublethal Pb Exposure Produces Season-Dependent Effects on Immune Response, Oxidative Balance and Investment in Carotenoid-Based Coloration in Red-Legged Partridges. Environ. Sci. Technol. 2015, 49, 3839–3850. [Google Scholar] [CrossRef]
- Ortiz-Santaliestra, M.E.; Resano-Mayor, J.; Hernández-Matías, A.; Rodríguez-Estival, J.; Camarero, P.R.; Moleón, M.; Real, J.; Mateo, R. Pollutant Accumulation Patterns in Nestlings of an Avian Top Predator: Biochemical and Metabolic Effects. Sci. Total Environ. 2015, 538, 692–702. [Google Scholar] [CrossRef]
- Martínez-Haro, M.; Green, A.J.; Mateo, R. Effects of Lead Exposure on Oxidative Stress Biomarkers and Plasma Biochemistry in Waterbirds in the Field. Environ. Res. 2011, 111, 530–538. [Google Scholar] [CrossRef] [Green Version]
- Hargitai, R.; Nagy, G.; Nyiri, Z.; Bervoets, L.; Eke, Z.; Eens, M.; Török, J. Effects of Breeding Habitat (Woodland versus Urban) and Metal Pollution on the Egg Characteristics of Great Tits (Parus major). Sci. Total Environ. 2016, 544, 31–38. [Google Scholar] [CrossRef]
- Ruiz, S.R.; Espín, S.; Sánchez-Virosta, P.; Salminen, J.-P.; Lilley, T.M.; Eeva, T. Vitamin Profiles in Two Free-Living Passerine Birds under a Metal Pollution Gradient—A Calcium Supplementation Experiment. Ecotoxicol. Environ. Saf. 2017, 138, 242–252. [Google Scholar] [CrossRef]
- Negro, J.J.; Figuerola, J.; Garrido, J.; Green, A.J. Fat Stores in Birds: An Overlooked Sink for Carotenoid Pigments? Funct. Ecol. 2001, 15, 297–303. [Google Scholar] [CrossRef]
- Hõrak, P.; Zilmer, M.; Saks, L.; Ots, I.; Karu, U.; Zilmer, K. Antioxidant Protection, Carotenoids and the Costs of Immune Challenge in Greenfinches. J. Exp. Biol. 2006, 209, 4329–4338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinha, S.; Mukherji, S.; Dutta, J. Effect of Manganese Toxicity on Pigment Content, Hill Activity and Photosynthetic Rate of Vigna radiata, L. Wilczek Seedlings. J. Environ. Biol. 2002, 23, 253–257. [Google Scholar] [PubMed]
- Saidi, I.; Nawel, N.; Djebali, W. Role of Selenium in Preventing Manganese Toxicity in Sunflower (Helianthus annuus) Seedling. S. Afr. J. Bot. 2014, 94, 88–94. [Google Scholar] [CrossRef] [Green Version]



| Element | Group b | LOQ | T. merula (n = 42) | C. livia (n = 27) | ||||
|---|---|---|---|---|---|---|---|---|
| Range (min—max) | Median | %<LOQ | Range (min—max) | Median | %<LOQ | |||
| Aluminum (Al) | 2 | 38.426 | <LOQ—2010 | 16.9 | 95 | <LOQ—365 | 15.0 | 96 |
| Antimony (Sb) | 2 | 0.010 | <LOQ—7.04 | 0.0060 | 95 | N/A | <LOQ | 100 |
| Arsenic (As) | 2 | 0.008 | <LOQ—44.2 | 2.56 | 2 | 0.686—72.2 | 2.33 | 0 |
| Barium (Ba) | 2 | 1.016 | 4.64—44.2 | 15.0 | 0 | <LOQ—56.4 | 7.12 | 44 |
| Beryllium (Be) | 2 | 0.050 | <LOQ—0.597 | 0.0208 | 88 | <LOQ—0.543 | 0.0317 | 96 |
| Bismuth (Bi) | 4 | 0.050 | <LOQ—0.464 | 0.0204 | 95 | N/A | <LOQ | 100 |
| Cadmium (Cd) | 2 | 0.010 | <LOQ—2.61 | 0.254 | 19 | <LOQ—1.08 | 0.328 | 19 |
| Cerium (Ce) | 3 | 0.050 | <LOQ—1.92 | 0.554 | 40 | <LOQ—1.12 | 0.0258 | 96 |
| Chromium (Cr) | 1, 2 | 0.229 | <LOQ—12.3 | 1.10 | 12 | <LOQ—11.3 | 0.138 | 81 |
| Cobalt (Co) | 1, 2 | 0.011 | 0.350—2.72 | 1.01 | 0 | <LOQ—1.52 | 0.821 | 30 |
| Copper (Cu) | 1, 2 | 1.724 | 137—310 | 200 | 0 | 153—299 | 225 | 0 |
| Dysprosium (Dy) | 3 | 0.005 | <LOQ—0.0409 | 0.0027 | 88 | <LOQ—0.0294 | 0.0026 | 89 |
| Erbium (Er) | 3 | 0.050 | N/A | <LOQ | 100 | N/A | <LOQ | 100 |
| Europium (Eu) | 3 | 0.005 | <LOQ—0.0122 | 0.0035 | 71 | <LOQ—0.0103 | 0.0019 | 96 |
| Gadolinium (Gd) | 3 | 0.005 | <LOQ—0.0523 | 0.0036 | 62 | <LOQ—0.0610 | 0.0027 | 93 |
| Gallium (Ga) | 4 | 0.050 | <LOQ—0.365 | 0.127 | 29 | <LOQ—0.292 | 0.173 | 33 |
| Gold (Au) | 4 | 0.005 | <LOQ—2.08 | 0.0025 | 95 | N/A | <LOQ | 100 |
| Holmium (Ho) | 3 | 0.005 | <LOQ—0.0064 | 0.0024 | 95 | <LOQ—0.0061 | 0.0030 | 93 |
| Indium (In) | 4 | 0.005 | <LOQ—0.0402 | 0.0035 | 60 | <LOQ—0.0419 | 0.0027 | 85 |
| Iron (Fe) | 1 | 24.645 | 246,000—410,000 | 330,000 | 0 | 221,000—476,000 | 400,000 | 0 |
| Lanthanum (La) | 3 | 0.100 | <LOQ—0.285 | 0.0539 | 81 | <LOQ—0.591 | 0.0435 | 96 |
| Lead (Pb) | 2 | 0.361 | 5.36—1350 | 114 | 0 | <LOQ—252 | 30.6 | 7 |
| Lutetium (Lu) | 3 | 0.005 | N/A | <LOQ | 100 | N/A | <LOQ | 100 |
| Manganese (Mn) | 1, 2 | 0.371 | 7.84—53.8 | 16.8 | 0 | 10.5—94.9 | 17.9 | 0 |
| Mercury (Hg) | 2 | 0.010 | 4.34—197 | 33.7 | 0 | <LOQ—24.4 | 0.0070 | 74 |
| Molybdenum (Mo) | 1 | 0.148 | 3.23—40.6 | 18.1 | 0 | <LOQ—29.1 | 14.1 | 4 |
| Neodymium (Nd) | 3 | 0.005 | <LOQ—0.282 | 0.0023 | 83 | N/A | <LOQ | 100 |
| Nickel (Ni) | 1, 2 | 7.946 | N/A | <LOQ | 100 | <LOQ—158 | 4.18 | 93 |
| Niobium (Nb) | 4 | 0.005 | <LOQ—0.256 | 0.0032 | 76 | <LOQ—0.320 | 0.0025 | 96 |
| Osmium (Os) | 4 | 0.005 | N/A | <LOQ | 100 | <LOQ—0.0059 | 0.0030 | 70 |
| Palladium (Pd) | 2 | 0.010 | N/A | <LOQ | 100 | <LOQ—0.0611 | 0.0050 | 74 |
| Platinum (Pt) | 4 | 0.005 | N/A | <LOQ | 100 | <LOQ—0.0772 | 0.0047 | 56 |
| Praseodymium (Pr) | 3 | 0.005 | <LOQ—0.0798 | 0.0029 | 90 | N/A | <LOQ | 100 |
| Ruthenium (Ru) | 4 | 0.005 | <LOQ—0.0114 | 0.0031 | 95 | <LOQ—0.0111 | 0.0025 | 89 |
| Samarium (Sm) | 3 | 0.005 | <LOQ—0.0626 | 0.0027 | 95 | <LOQ—0.0491 | 0.0020 | 96 |
| Selenium (Se) | 1, 2 | 0.153 | 93.6—10,700 | 520 | 0 | 139—621 | 330 | 0 |
| Silver (Ag) | 2 | 0.100 | <LOQ—3.09 | 0.0833 | 64 | <LOQ—0.697 | 0.0393 | 89 |
| Strontium (Sr)c | 2 | 0.439 | 28.5—162 | 58.7 | 0 | 22.4—153 | 49.5 | 0 |
| Tantalum (Ta) | 4 | 0.005 | <LOQ—0.219 | 0.0047 | 60 | N/A | <LOQ | 100 |
| Terbium (Tb) | 3 | 0.005 | N/A | <LOQ | 100 | N/A | <LOQ | 100 |
| Thallium (Tl) | 2 | 0.050 | <LOQ—0.914 | 0.0750 | 33 | <LOQ—0.748 | 0.0456 | 59 |
| Thorium (Th) | 2 | 0.050 | <LOQ—0.0599 | 0.0281 | 93 | <LOQ—0.182 | 0.0329 | 93 |
| Thulium (Tm) | 3 | 0.005 | N/A | <LOQ | 100 | N/A | <LOQ | 100 |
| Tin (Sn) | 2 | 0.010 | <LOQ—5.42 | 0.170 | 45 | <LOQ—2.74 | 0.0051 | 85 |
| Titanium (Ti) | 4 | 0.757 | <LOQ—23.4 | 14.5 | 43 | <LOQ—62.6 | 0.521 | 67 |
| Uranium (U) | 2 | 0.050 | N/A | <LOQ | 100 | <LOQ—0.0785 | 0.0302 | 96 |
| Vanadium (V) | 2 | 0.050 | <LOQ—1.29 | 0.0358 | 67 | <LOQ—8.41 | 2.20 | 7 |
| Ytterbium (Yb) | 3 | 0.005 | <LOQ—0.0147 | 0.0020 | 76 | <LOQ—0.0103 | 0.0034 | 78 |
| Yttrium (Y) | 3 | 0.005 | <LOQ—0.195 | 0.0038 | 62 | <LOQ—0.198 | 0.0026 | 96 |
| Zinc (Zn) | 1, 2 | 51.031 | 4030—5980 | 4950 | 0 | 2190—7320 | 5600 | 0 |
| Mean | LM | |||||
|---|---|---|---|---|---|---|
| T. merula | Variable | Agric.-urban (CL) | Mining (CL) | Control (CL) | F df | p |
| Arsenic (As) * | 2.87 a (1.34–6.15) | 3.81 a (2.01–7.21) | 0.494 b (0.238–1.03) | 9.98 2,39 | 0.0003 | |
| Barium (Ba) * | 8.81 a (6.57–11.8) | 17.6 b (13.7–22.5) | 16.5 b (12.5–21.9) | 7.57 2,39 | 0.0017 | |
| Cadmium (Cd) * | 0.185 a (0.0722–0.474) | 0.523 a (0.237–1.15) | 0.0147 b (0.00597–0.0364) | 18.5 2,39 | <0.0001 | |
| Chromium (Cr) * | 0.716 (0.362–1.41) | 1.28 (0.720–2.26) | 0.650 (0.338–1.25) | 1.49 2,39 | 0.24 | |
| Cobalt (Co) * | 1.04 (0.809–1.33) | 1.15 (0.931–1.41) | 0.808 (0.637–1.03) | 2.56 2,39 | 0.090 | |
| Copper (Cu) | 219 (198–239) | 206 (188–223) | 188 (168–208) | 2.46 2,39 | 0.099 | |
| Iron (Fe) | 336,000 (310,000–363,000) | 327,000 (304,000–349,000) | 336,000 (310,000–361,000) | 0.22 2,39 | 0.81 | |
| Lead (Pb) * | 109 a (80.4–148) | 779 b (603–1010) | 13.4 c (9.98–17.9) | 224 2,39 | <0.0001 | |
| Manganese (Mn) * | 14.8 a (12.0–18.3) | 21.9 b (18.3–26.1) | 14.9 a (12.1–18.2) | 5.79 2,39 | 0.0063 | |
| Mercury (Hg) * | 77.5 a (46.0–131) | 21.5 b (13.9–33.4) | 29.6 b (17.9–48.8) | 7.49 2,39 | 0.0018 | |
| Molybdenum (Mo) | 26.6 a (23.5–29.7) | 11.1 b (8.47–13.7) | 19.8 c (16.8–22.8) | 30.5 2,39 | <0.0001 | |
| Selenium (Se) * | 1020 a (578–1810) | 415 b (257–671) | 788 ab (455–1360) | 3.32 2,39 | 0.047 | |
| Strontium (Sr) * | 66.1 a (55.7–78.4) | 50.2 b (43.5–58.0) | 79.7 a (67.7–94.0) | 9.46 2,39 | 0.0004 | |
| Zinc (Zn) * | 5040 (4760–5340) | 4940 (4710–5190) | 4820 (4560–5090) | 0.69 2,39 | 0.51 | |
| Hematocrit (%) | 41.9 (38.2–45.5) | 37.2 (34.2–40.3) | 39.1 (35.6–42.6) | 1.96 2,39 | 0.15 | |
| Body mass (g) | 82.8 (79.3–86.3) | 84.5 (81.6–87.5) | 80.9 (77.6–84.3) | 1.31 2,39 | 0.28 | |
| Wing length (mm) ** | 122 (120–124) | 124 (123–126) | 123 (120–125) | 1.55 2,26 | 0.23 | |
| Retinol (µM/mL) * | 8.92 a (7.59–10.5) | 9.54 a (8.26–11.0) | 14.3 b (12.3–16.7) | 11.3 2,37 | 0.0002 | |
| Tocopherol (µM/mL) | 47.3 a (37.9–56.8) | 31.7 b (23.2–40.1) | 45.0 ab (35.9–54.1) | 3.79 2,37 | 0.032 | |
| Lutein (µM/mL) * | 15.1 a (9.55–23.9) | 6.40 b (4.24–9.66) | 12.2 ab (7.82–18.9) | 4.47 2,37 | 0.018 | |
| C. livia | Variable | Agric.-urban (CL) | Mining (CL) | Control (CL) | F df | p |
| Arsenic (As) * | N/A | 12.5 (6.47–24.2) | 1.93 (1.16–3.20) | 21.4 1,25 | <0.0001 | |
| Cadmium (Cd) * | N/A | 0.478 (0.145–1.58) | 0.0799 (0.0320–0.200) | 5.99 1,25 | 0.022 | |
| Copper (Cu) | N/A | 224 (199–249) | 215 (196–235) | 0.31 1,25 | 0.58 | |
| Iron (Fe) | N/A | 408,000 (371,000–446,000) | 391,000 (362,000–420,000) | 0.58 1,25 | 0.45 | |
| Lead (Pb) * | N/A | 90.6 (35.8–229) | 9.02 (4.43–18.4) | 16.5 1,25 | 0.0004 | |
| Manganese (Mn) * | N/A | 23.0 (16.0–33.2) | 18.9 (14.3–25.0) | 0.78 1,25 | 0.39 | |
| Molybdenum (Mo) | N/A | 15.9 (11.9–19.8) | 13.1 (10.0–16.1) | 1.34 1,25 | 0.26 | |
| Selenium (Se) * | N/A | 397 (322–490) | 312 (266–366) | 3.58 1,25 | 0.070 | |
| Strontium (Sr) * | N/A | 38.8 (29.3–51.5) | 59.3 (47.7–73.6) | 5.96 1,25 | 0.022 | |
| Vanadium (V) * | N/A | 2.34 (0.985–5.54) | 1.28 (0.661–2.48) | 1.29 1,25 | 0.27 | |
| Zinc (Zn) * | N/A | 5400 (4610–6330) | 5170 (4580–5850) | 0.20 1,25 | 0.66 | |
| Hematocrit (%) *** | N/A | 64.0 (55.7–72.3) | 57.6 (54.1–61.1) | 2.25 1,18 | 0.15 | |
| Body mass (g) | N/A | 341 (311–371) | 330 (307–353) | 1.25 1,25 | 0.42 | |
| Wing length (mm) ** | N/A | 233 (229–237) | 230 (226–234) | 1.20 1,13 | 0.29 | |
| Retinol (µM/mL) | N/A | 7.96 (6.17–9.74) | 9.53 (8.17–10.9) | 2.09 1,25 | 0.16 | |
| Tocopherol (µM/mL) * | N/A | 32.5 (21.2–49.9) | 33.0 (23.8–45.8) | 0.00 1,25 | 0.95 | |
| Lutein (µM/mL) | N/A | 10.5 (4.88–16.2) | 13.9 (9.58–18.2) | 0.96 1,25 | 0.34 | |
| F (Estimate ± SE) | |||||
|---|---|---|---|---|---|
| T. merula | n | PC1 (As+, Cd+, Pb+) | PC2 (Hg+, Mn−) | Body mass | Wing length |
| Hematocrit (%) | 42 | 0.08 (0.19 ± 0.65) | 4.07 (1.62 ± 0.80) ° | 0.00 (−0.01 ± 0.17) | 2.18 (−0.37 ± 0.25) |
| Retinol (μM/mL) a | 40 | 14.1 (−0.052 ± 0.014) *** | 0.65 (−0.013 ± 0.016) | 0.20 (−0.0016 ± 0.0035) | 0.01 (0.00058 ± 0.0053) |
| Tocopherol (μM/mL) | 40 | 0.79 (−1.52 ± 1.70) | 1.39 (2.37 ± 2.01) | 8.85 (−1.26 ± 0.42) ** | 0.00 (−0.038 ± 0.64) |
| Lutein (μM/mL) a | 40 | 0.22 (−0.017 ± 0.035) | 7.84 (0.12 ± 0.041) ** | 7.18 (−0.023 ± 0.087) * | 0.67 (−0.011 ± 0.013) |
| C. livia | n | PC1 (As+, Cd+, Pb+) | Body mass | Wing length | |
| Hematocrit | 20 | 3.82 (2.05 ± 1.05) ° | 0.00 (0.00056 ± 0.041) | 2.63 (0.33 ± 0.20) | |
| Retinol (μM/mL) | 27 | 0.32 (−0.23 ± 0.40) | 0.05 (0.0036 ± 0.017) | 2.02 (0.12 ± 0.084) | |
| Tocopherol (μM/mL) a | 27 | 1.07 (0.043 ± 0.042) | 0.08 (0.00050 ± 0.0017) | 1.51 (−0.011 ± 0.0086) | |
| Lutein (μM/mL) | 27 | 0.02 (−0.21 ± 1.36) | 0.05 (−0.012 ± 0.057) | 0.27 (0.15 ± 0.28) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-Virosta, P.; Zamora-Marín, J.M.; León-Ortega, M.; Jiménez, P.J.; Rivas, S.; Sánchez-Morales, L.; Camarero, P.R.; Mateo, R.; Zumbado, M.; Luzardo, O.P.; et al. Blood Toxic Elements and Effects on Plasma Vitamins and Carotenoids in Two Wild Bird Species: Turdus merula and Columba livia. Toxics 2021, 9, 219. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics9090219
Sánchez-Virosta P, Zamora-Marín JM, León-Ortega M, Jiménez PJ, Rivas S, Sánchez-Morales L, Camarero PR, Mateo R, Zumbado M, Luzardo OP, et al. Blood Toxic Elements and Effects on Plasma Vitamins and Carotenoids in Two Wild Bird Species: Turdus merula and Columba livia. Toxics. 2021; 9(9):219. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics9090219
Chicago/Turabian StyleSánchez-Virosta, Pablo, José Manuel Zamora-Marín, Mario León-Ortega, Pedro J. Jiménez, Silvia Rivas, Lidia Sánchez-Morales, Pablo R. Camarero, Rafael Mateo, Manuel Zumbado, Octavio P. Luzardo, and et al. 2021. "Blood Toxic Elements and Effects on Plasma Vitamins and Carotenoids in Two Wild Bird Species: Turdus merula and Columba livia" Toxics 9, no. 9: 219. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics9090219