Zebrafish as an Emerging Model for Bioassay-Guided Natural Product Drug Discovery for Neurological Disorders




1
Molecular and Nanomedicine Research Unit (MNRU), Centre for Nanoscience and Nanotechnology (CNSNT), Sathyabama Institute of Science and Technology, Chennai 600119, Tamil Nadu, India
2
School of Health Sciences, Purdue University, West Lafayette, IN 47907, USA
*
Authors to whom correspondence should be addressed.
Medicines 2019, 6(2), 61; https://0-doi-org.brum.beds.ac.uk/10.3390/medicines6020061
Submission received: 29 April 2019
/
Revised: 26 May 2019
/
Accepted: 27 May 2019
/
Published: 30 May 2019
(This article belongs to the Special Issue Biological Efficacy of Natural Products against Noncommunicable Diseases)
Abstract
:Most neurodegenerative diseases are currently incurable, with large social and economic impacts. Recently, there has been renewed interest in investigating natural products in the modern drug discovery paradigm as novel, bioactive small molecules. Moreover, the discovery of potential therapies for neurological disorders is challenging and involves developing optimized animal models for drug screening. In contemporary biomedicine, the growing need to develop experimental models to obtain a detailed understanding of malady conditions and to portray pioneering treatments has resulted in the application of zebrafish to close the gap between in vitro and in vivo assays. Zebrafish in pharmacogenetics and neuropharmacology are rapidly becoming a widely used organism. Brain function, dysfunction, genetic, and pharmacological modulation considerations are enhanced by both larval and adult zebrafish. Bioassay-guided identification of natural products using zebrafish presents as an attractive strategy for generating new lead compounds. Here, we see evidence that the zebrafish’s central nervous system is suitable for modeling human neurological disease and we review and evaluate natural product research using zebrafish as a vertebrate model platform to systematically identify bioactive natural products. Finally, we review recently developed zebrafish models of neurological disorders that have the potential to be applied in this field of research.
1. Introduction
Central nervous system (CNS) diseases and disorders, including Alzheimer’s disease (AD), schizophrenia (SCZ), Huntington’s disease (HD), and Parkinson’s disease (PD) [1,2,3], signify a global burden on society in terms of disability, economic loss, and human suffering. Globally, more than a million people have CNS disorders [4]. CNS disorders are multifaceted diseases with unclear causes and often ineffective therapies, with only a few therapeutic drugs being clinically effective [5]. Natural products (NPs) are small molecules synthesized from living organisms (plants, bacteria, and fungi) and are similar to secondary metabolites. Among all existing sources for drug discovery against single targets of new lead compounds [6], NPs are most promising but are underutilized. Crude extracts from NPs are a complex mixture of mostly uncharacterized compounds, some of which may have unwanted effects. Worldwide, nearly 30% of all top-selling drugs are NPs or their derivatives. NPs are an excellent source of new drug-like compounds to be discovered, and their diversity of chemicals has helped to develop drugs for a wide range of neurodegenerative disorders. Most new drugs have been authorized from either NPs themselves or NPs over the past 30 years [7,8,9]. In complex NP extracts, the isolation and structural characterization of bioactive small molecules involves several new methodologies that need considerable time and effort [10,11]. Furthermore, there are several methods involved in testing NPs in high-throughput screening (HTS). Combinatorial libraries with NP-like compounds have been recently used in HTS [9].
A vital component of the drug discovery program for NPs is bioassay-guided separation. In bioassay-guided separation, each chromatographic fractionation undertakes biological evaluation for further fractionation, and only biologically active fractions are selected. The crude extracts are fractionated and evaluated in bioassays. Further fractionation is repeated until the chosen activity isolates pure compounds and then characterizes them structurally. Novel pharmaceutically active NPs had been identified through screening and fractionation of crude extracts using several presently regarded in vitro assays, collectively with (i) cell fractions, (ii) entire cellular assays, or (iii) recombinant enzymes as target molecules [12]. Notwithstanding its application for HTS identification, the biomedical relevance of the isolated active metabolites can be limited when using only enzymatic or in vitro assays. To overcome this limitation, high-resolution micro-fractionation can be coupled with high-content bioassays to further analyze the separate constituents. In contrast to cell-based reporter or enzymatic assays, high-content bioassays (e.g., phenotypic assays using some cells or organisms) allow for an impartial investigation of pharmacological activity. Many in vivo animal models offer the possibility of independent screening of biomedically relevant bioactivities. However, milligrams of compounds are required for mammalian models and are thus not ideal for in vivo platforms for micro-fractionation and rapid HPLC profiling approaches.
Moreover, many naturally derived active compounds not only play a role as drugs but also help in the development of many new model structures for synthetic molecules through combinatorial chemistry. During the last 20 years, about 50% of drugs introduced to the market have been derived indirectly or directly from small bioactive molecules. As a source of chemical diversity, unfulfilled expectations from current R&D strategies and emerging trends have led to interest in NPs [7,13]. NPs have attracted considerable attention in the treatment of CNS diseases due to their neuroprotective and therapeutic effects. NPs are excellent sources of safe, precise, and effective anti-neurotherapeutic agents and thus are useful in the development of safer substitutes to pharmaceuticals. Recent literature suggests that many bioactive compounds have both neurotrophic and neuroprotective actions [14]; therefore, for peripheral neuropathy early treatment using phytochemical approaches could be one of the important strategies in preventing many neurological disorders.
Many presently known bioactive NPs have been previously recognized for their activity-guided extract isolation through the use of in vitro assays. Biologically active NPs have been identified by physical characteristics using chromatography, mass spectrometry, and NMR spectroscopy analysis. In vivo bioassay-guided fractionation has not widely been used for the discovery of drug-like NPs, as traditional in vivo models (e.g., mice and rats) are low-throughput systems and require much larger quantities of compounds for testing in these systems.
The zebrafish (Danio rerio) provides a complementary integrative biological model for the discovery of natural drug-like products through in vivo bioassay-guided chromatographic fractions requiring only microgram quantities of individual components. Zebrafish are vertebrates, and thus are more evolutionary similar to humans compared to non-vertebrate models. Logistically speaking, zebrafish are tiny and can be kept in a small space in high numbers. Zebrafish are currently emerging as an in vivo vertebrate model system for drug discovery and functional genomics [15]. In addition to their many pharmacological and physiological similarities with mammals, zebrafish have many added advantages including small size of embryos and larvae (0.5 to 5 mm depending on the stage of development), optical transparency, rapid ex vivo development, and high fecundity (up to hundreds of offspring per day). These characteristics makes zebrafish a standalone versatile experimental in vivo model compatible with HTS and NP discovery micro-fraction techniques [16]. Furthermore, zebrafish embryos and larvae provide the convenience of using microtiter plates (96-well and even 384-well plates) to test the activity of micro-fraction isolated natural compounds. Depending on the performance of these isolated compounds, the need for only microgram quantities to initiate an initial biological reaction represents another advantage of using zebrafish as a model organism in comparison to other vertebrates (e.g., rodents, where the energy dose requirements are typically one thousand times higher) [17]. This latter feature is prime to NP discovery, as many high-resolution HPLC-based separation techniques, particularly micro-fractionation, bring about very limited pattern quantities that could otherwise be inadequate for in vivo activity analysis.
The neuroprotective activity of bioactive compounds from herbal drugs has been proven by using cellular or animal models [18,19,20]. However, effective delivery of drugs to the brain remains the main task in the discovery and development of new CNS disease treatments [21,22]. This review focuses on neurological disorders with an emphasis on neurodegenerative diseases, use of zebrafish for bioassay-guided isolation of neuroactive small molecule from NPs, and new methods to develop zebrafish neurodegenerative models that have the potential for expansion into NP drug discovery applications.
2. Neurodegenerative Diseases
Neurodegenerative diseases lead to a rapid loss of brain processes such as cognitive and/or motor neuron function, and are a major challenge facing aging populations. AD, PD, HD, and amyotrophic lateral sclerosis (ALS) are common neurodegenerative diseases. Neurodegenerative diseases share common characteristics and mechanisms despite their different clinical forms. One of these features is regional cytosolic or nuclear protein aggregation [23]. Specific features include extra cell deposition of plaques of amyloid-beta (Aβ), intracellular accumulation of inclusions of hyperphosphorylated microtubule-binding tau in AD, intracellular storage of α-synuclein in PD, inclusion of TAR DNA-binding protein (TDP)-43 transactive response in ALS, frontotemporal dementia, and polyglutamine protein aggregates in HD and other repeat CAG-polyglutamine diseases. While for some cases genetic causes have been identified, the main driver is a complex interaction of predisposition factors in genetics and the environment. In every common neurodegenerative disease condition, there is usually a mixture of hereditary and "sporadic" forms. While the identity of many mutated genes in family forms of AD, PD, and ALS is known, the function of such genes and how their mutations induce neuronal degeneration is not fully understood. Processes that cause degeneration and the death of particular neuron types are probably the most important discovery goals in the field, shaping the disease’s manifestations and defining the characteristics of all neurodegenerative diseases.
3. Using the Zebrafish Model for Neurological Disorders
The zebrafish is being progressively used to model neurodegenerative diseases and neurological disorders successfully [24,25,26,27,28,29,30,31,32,33], with promises to test potential treatments for diseases and disorders [31,34]. The zebrafish CNS is similarly arranged to that of other vertebrates, and is traditionally separated into the hindbrain, midbrain, forebrain, ascending and descending spinal cord, cranial nerves, motor spinal cord, and sensory nerves. Zebrafish neuroanatomy has been examined and described in detail elsewhere during development, as well as in adults [35,36]. The genome of the zebrafish is widely annotated [37]. The evolutionary lineage of zebrafish (teleost-bonyfish) separated about 450 million years ago from the human lineage (tetrapod) [38]. Zebrafish pairs can produce large number of embryos that make it possible to achieve relatively high-throughput screening drug studies and behavioral testing [15] with simple methods for modulating gene expression available [39,40]. Many human-associated neurodegenerative disease proteins in zebrafish are homologous, highlighting potentially preserved molecular cellular functions that can be easily examined [28] (Table 1).
3.1. Zebrafish and Alzheimer’s Disease
The most common form of irreversible neurodegenerative disorder and dementia is Alzheimer’s disease (AD). Fifty million people were estimated to live with AD in 2018, and this figure is predicted to increase to 152 million by 2050 [62]. AD’s main clinical feature is progressive memory loss, motor and speech impairment, deception, depression, and aggressive behavior [63]. There is significant neuronal damage in AD patients in numerous brain regions [64,65]. This is usually caused by extracellular deposition of amyloid-beta peptide and tau protein aggregates called neurofibrillary tangles (NFTs). Several risk factors are identified or under investigation, including both genetic and environmental factors, as potential triggers of AD. AD may be classified as familial AD (FAD, at < 65 years of age) or sporadic AD (SAD, at > 65 years of age). Most of the knowledge of AD pathogenesis has been defined by studying FAD mutations. Some of the genetic targets are precursor protein amyloid-ß (APP) and presenilins (PSEN1 and PSEN2) associated with increased FAD risk. The more common form of AD occurs sporadically (representing >90% of cases) [66]. Multi-faceted pathogenesis of SAD is associated with several risk factors such as old age and the presence of the apolipoprotein (APOE) gene ε4 allele and/or the recently identified genetic risk factor sortilin-related receptor (SORL1). SORL1 is an APOE receptor with primary expression in neurons of the brain [67].
Zebrafish have human orthologous genes that play key roles in AD. The zebrafish genes psen1 [42] and psen2 [43] are human PSEN1 and PSEN2 orthologs, respectively, whereas the genes appa and appb are human APP co-orthologs [41]. The zebrafish genome also contains orthologous genes for gamma-secretase’s complex components, PSENEN (psenen) [47], NCTN (ncstn) [48], and APH1b (aph1b). In addition, β-secretase orthologs (BACE1 and BACE2) were also identified (bace1 [44] and bace2 [45]) in zebrafish. The zebrafish genome contains co-orthologs of the microtubule-associated tau protein (MAPT) gene, which encodes tau protein (mapta, and maptb) [68]. The human APOE and SORL1 co-orthologs apoea and apoeb are also present in the zebrafish genome [37,50] and sorl1 [51], respectively.
3.2. Zebrafish and Parkinson’s Disease
Dopaminergic neuron degeneration, as well as the presence of Lewy bodies called intracytoplasmic inclusions, are neuropathological lesions associated with Parkinson’s disease (PD). Six genes associated with Parkinsonism have been identified, including Parkin, DJ-1, PINK1, α-Synuclein, UCHL-1, and LRRK2 [69]. Although predominantly a motion disorder, PD is a mixed group of neurological conditions that are not capable of producing or controlling movement and cognitive impairment [70]. The human PARK2 ortholog in zebrafish (park2) resides on chromosome 13, and encodes a protein of 458 amino acids (465 in humans) [53]. The PINK1 zebrafish ortholog has 54% similarity, and an initial study reported a severe developmental phenotype in pink1 k/d zebrafish [54]. The PARK7 zebrafish ortholog encodes a protein of 189 amino acids with 83% human DJ-1 identity [52].
3.3. Zebrafish and Huntington’s and Other Polyglutamine Diseases
Huntington’s disease (HD) is a monogenic neurodegenerative disease that follows an autosomal dominant pattern of huntingtin gene mutant form (HTT) inheritance. The mutation encodes for an abnormal trinucleotide that leads to glutamine (CAG) expansion at the HTT protein amino terminal and arises in an extended polyglutamine tract of the Huntingtin protein [71]. This causes cell death by gain of function mechanisms, like protein accumulation, mitochondrial dysfunction, and caspase activation. A decline in normal Huntingtin can also make a significant contribution to pathogenesis [72]. To try to elucidate the loss as well as the gain of function mechanisms, zebrafish models are being used. The HD cDNA homology in zebrafish was isolated as the first step towards discovering the possible role of the HD gene in initial vertebrate development [56]. This cDNA codes a predicted protein product of 3121 amino acids with a human HTT identity of 70%. Loss of developmental expression of 15/hd1 caused noticeable morphological abnormalities, including pericardial edema microcephalus and CNS necrosis [73]. Zebrafish htt is also necessary for normal pharyngeal arch cartilage development [74].
3.4. Zebrafish and Amyotrophic Lateral Sclerosis (ALS)
Amyotrophic lateral sclerosis (ALS) is characterized by protein inclusions present in the affected neurons. These protein inclusions are linked to spinal cord motor neuron loss and downward motor tracts in the brain and spinal cord. Familial ALS is fairly rare, but a gene-encoding mutation of superoxide dismutase (SOD1) inherited in an autosomal dominant motif causes 20% of such cases [75,76]. The mutations are usually prescribed by gain of function mechanisms [77]. Over 150 mutations have been discovered in SOD1, including the point mutations G93R and G85R. Recent studies also indicate a role for SOD1 in the sporadic form of ALS and propose a prion-like function of protein misfolding. Moreover, a few of the recently identified genes involved in ALS, such as FUS and TARDBP, also demonstrate a high tendency to act similar to prions in misfolding proteins.
A recent study used zebrafish to assess overexpression of SOD1 by mRNA microinjection to study ALS etiology. In this study, vascular endothelial growth factor (VEGF) overexpression rescued the SOD1-expressing zebrafish axonopathy, while VEGF morpholino knockdown exacerbated the abnormalities [27]. However, one of the limitations in working with ALS in vivo models is the lack of comprehensive methods to assess the presymptomatic course of the disease. The zebrafish provides advantages in the study of processes of early disease with rapid development and reach post-embryonic life about 3 days post fertilization (dpf), which is similar to neonatal mouse development.
3.5. Zebrafish and Schizophrenia
Schizophrenia is a severe neurodegenerative disorder with the etiology of hallucination, delusions, depression, agitated body movements, confused thoughts and snafu speech, anhedonia, lack of motivation, and speech problems. The defects of schizophrenia are caused by early development in the brain [78]. About 1% of the world’s population is affected by schizophrenia and is characterized by neuronal dysfunction, which results in deficiencies in various cognitive areas including visual and verbal memory, learning, and attention [79]. Patients with schizophrenia, as well as with HD, have impaired preimpulse inhibition (PPI) [80,81], a type of sensorimotor gaiting [82]. PPI is a neurological event where the response following shocking stimulus is defeated by a weak prestimulus or prepulse and is conserved among vertebrates. The sensorimotor zebrafish gating has been described in 6 dpf larvae for PPI testing [83]. Twin studies have a projected heritage of around 81% for schizophrenia and an environmental impact of about 11% (factors such as diet, parenting, and exposure to toxins or teratogens) [84]. A large number of cases of schizophrenia are sporadic and appear in a family without a history of the disease [85]. Many genes have been linked to schizophrenia susceptibility. Genes with a largely robust disease connection include dystrobrevin binding protein 1 (DTNBP1), neuregulin1, disrupted in schizophrenia1 (DISC1), kinesin family member 1 (KIF1), kinesin family member 17 (KIF17), SH3, multiple ankyrin repeat domains 3 (SHANK3), and NOTCH4 [86,87]. Candidate genes for schizophrenia may be vital in determining neuronal migration, neurogenesis, and cell fate [88].
Burgess and Granato [89] developed an endophenotype of schizophrenia in zebrafish PPI. Exposure to apomorphine and ketamine influences zebrafish PPI, and therefore appears to be facilitated by similar neurotransmitters as in other animals. The same study also identified five novel mutant lines with abnormal PPI responses. One of the most intensively studied genes associated with schizophrenia is DISC1, and was first identified with a high incidence of depression, schizophrenia, and bipolar disorder in a Scottish pedigree [90]. Furthermore, disc1 studies in zebrafish have provided new information on this gene’s function.
3.6. Zebrafish and Epilepsy
Epilepsy is a common neurological disease caused by unexpected seizures that can vary from a short attention interval to severe and prolonged seizures and muscle cramps [91]. Epilepsy has a pathological mechanism that is poorly understood and is a complex brain disorder with many fundamental causes [92]. Zebrafish have a multifaceted nervous system with elegant behaviors, and are prone to seizure. Adult zebrafish have a wide array of established behaviors that can be studied, making them especially suitable for model development. The pentylenetetrazole (PTZ)-induced zebrafish epileptic seizure has been used to study the mechanism of epilepsy. The affordability of both larval and adult zebrafish, which allows the ontogenesis to investigate a wider range of epilepsy-related phenomena, is also useful.
Several genetically altered zebrafish are now being used to study the behavior and brain function associated with epilepsy. Zebrafish (∼5–7 dpf) are commonly placed in multiple wells and tracked using video tracking software, continuously recorded by a camera. Mutations in two family members, Potassium Voltage-Gated Channel Subfamily Q Member 2 (KCNQ2) and Potassium Voltage-Gated Channel Subfamily Q Member 3 (KCNQ3), have been correlated with inherited neonatal epilepsy, e.g., benign family neonatal convulsions. These genes are highly expressed in zebrafish, providing support for studies of epilepsy using this vertebrate model [93].
4. Zebrafish Bioassay-Guided Isolation of Natural Product Drug Discovery
Zebrafish was first suggested by Jones and Huffmann of the Oklahoma Research Foundation as a model for in vivo drug development in 1957, and soon thereafter zebrafish were first used to examine NP bioactivities. Zebrafish bioassay-guided identification of NPs in a number of neurological disorders can be an attractive approach for generating novel lead compounds (Figure 1). Over the past decade, zebrafish as a primary model for HTS in the scope of drug discovery for NPs for neurological disease has grown [16,94,95,96]. Zebrafish model profits combined with robust chromatographic and spectroscopic methods are creating a path to discover and further develop HIT compounds from various plant extracts [97,98].
Zebrafish has recently emerged as a strong model in a wide range of applications for rapid analysis of gene function and small molecular bioactivity [15]. Zebrafish are well-suited to identify therapeutically potential bioactive NPs (Table 2). Zebrafish were first proposed over fifty years ago as an in vivo model for the discovery of small molecules of drugs [99]. This preliminary study examined the utility of zebrafish embryos and larvae to screen both NPs and synthetic compounds. Zebrafish offers the opportunity of in vivo swift microgram-scale bioactivity evaluation of small molecules, an attractive feature combined with high-resolution fractionation technologies and analytical methods like UHPLC-TOF-MS and NMR microflow. A recent example is the bioassay-guided isolation of zebrafish with six spirostane glycosylated triterpene important for decoction and methanol extract anti-sizing activity from Solanum torvum aerial parts, which was discovered by Soura Challal [95] and his colleagues. In addition, the recently identified flavonoid-trans-tephrostachin inhibitory of acetylcholinesterase has also been isolated from the leaves of Indian herb Tephrosia purpurea by a zebrafish bioassay [96].
5. Development of Zebrafish Models for Neurological Disorders
In order to study the genes associated with various neurodegenerative disorders, the zebrafish has proven to be a perfect system where the genetic material is directly injected into the fertilized embryo. For instant study of gene function, effective techniques for the manipulation of gene expression are available [105]. By inserting genes into specific tissue promoters using vectors such as the Tol2 transposase system, transgenic zebrafish can be efficiently produced [106]. The Cre/loxP [107] and GAL4-UAS [108] gene function analysis systems can also be used at specific time points to generate conditionally expressed transgenics. The major disadvantage to induce specific mutations in the zebrafish genome was the unavailability of effective previous technologies. However, transcription activator-like effector nuclease (TALENS), zinc finger nucleases (ZFNs), and type II prokaryotic CRISPR (clustered regularly short palindromic repeats)/Cas systems for targeted gene sequences have been developed in recent times and are now being applied to create zebrafish transgenic models [109,110]. Furthermore, new technologies have expanded development of adult zebrafish and cell culture-based models.
5.1. Transgenic Zebrafish Models
Due to the effortless screening of genes and small molecules in zebrafish, innovative genetic pathways that enable the development stages for isolating chemical modifiers can be obtained easily. [111,112,113,114]. More recently, it was suggested that many of these benefits could be applied to the study of human disease: high-content small molecular screens, genetic suppressor screens, in vivo disease progression observations, use of fluorescent reporters to identify interesting cell populations, and rapid hypothesis testing experiments in statistically robust larvae samples could provide valuable insight into disease pathogenesis or new therapeutic approaches [115,116]. In 2008, ZFNs were used to describe the first targeted gene knockout in zebrafish, and morpholinos were used to show gene knockdowns in neurodegenerative diseases (Table 3). For example, an appb knockdown study showed that the dramatic developmental defects in embryos and function of appb were needed for axonal outgrowth of motor neurons in zebrafish [117]. In addition, the bace1 knockout zebrafish was generated by zinc finger nucleases. bace1 mutants in the PNS are hypomyelinated, whereas the CNS is not affected [45]. Furthermore, study of the leucine-rich repeat kinase 2 (LRRK2) gene associated with PD is being studied in the zebrafish. Along with neuronal loss, the morpholino-mediated gene knockdown of lrrk2 zebrafish also caused developmental disturbances in the eyes, lens, and otic vesicles, including axis curvature defects, eye abnormalities, and edema [118]. Since then, reverse genetic tools have seen unprecedented growth rates with the introduction of TALEN and CRISPR-Cas9 systems, including an apoea knockout for AD [119] and a tardbp knockin and fus knockin for ALS [40,120] (Table 3). Further development of transgenic models using the recently developed CRISPR technique is set to unravel a greater potential for zebrafish in gene knockdown and knockin studies.
5.2. Generation of a Neurodegenerative Model Using Amyloid-β42 (Aβ42) in the Adult Zebrafish Brain
The zebrafish bear extensive regenerative ability [126], and clinically important studies are aimed at understanding the mechanisms of zebrafish regeneration. Zebrafish are excellent tools because of their CNS regenerative capacity [127,128]. Models of neurodegeneration in the adult zebrafish brain will be helpful to investigate the activation state of the neural stem/progenitor cells (NSPCs) and to identify the molecular differences between zebrafish and mammalian NSPCs to utilize them for regenerative therapies [129]. Multifaceted inflammatory conditions in neurodegenerative diseases affect microglia, neurons, and NSPCs pleiotropically [130,131]. Kizil et al. first developed a gene knockdown method based on cerebroventricular microinjection (CVMI) in vivo morpholino oligonucleotide [132] in the adult zebrafish brain (Figure 2). CVMI injection in a skull incision is capable of uniformly targeting cells near the injection site, in this case the forebrain ventricular region containing neurogenic progenitor cells. The amyloid-β42 (Aβ42) induced neurotoxicity in adult zebrafish brain using CVMI of Aβ42 derivatives [133]. One of the earliest findings in understanding the etiology of AD was the discovery of a 40-length peptide in AD brains now called Aβ, which constitute the primary component of AD-related amyloid deposits [134,135]. Aβ is produced from the amyloid precursor protein (APP) with the continuous breakdown of β- and γ-secretases [136]. Importantly, the creation of Aβ through APP’s proteolytic processing is heterogeneous, leading to variable Aβ lengths, especially at the peptide’s carboxy terminus. 40 and 42 long residues are the two main forms of Aβ produced under normal APP processing conditions (Aβ40 and Aβ42, respectively). The shorter variety of Aβ40 is the majority of the Aβ produced in a normal individual [136]. Approximately 5%–15% of the total Aβ pool is Aβ42, and it is possible to observe smaller amounts of other Aβs, both longer and shorter. Generally, the brain’s Aβ pool has 5%–15% of Aβ42, which causes reminiscent phenotypes of amyloid pathophysiology: apoptosis, microglial activation, synaptic degeneration deficiencies, and learning. Aβ42 also results in NSPC proliferation and increased neurogenesis [37]. This understanding can help to design regenerative therapy-based drug discovery for neurological disorders.
5.3. Zebrafish Cell Culture-Based Neurodegenerative Disease Models
The developing zebrafish embryo is an excellent source for culturing cells, including neural cells [137,138,139,140,141,142,143]. The technique to culture primary motor neuron (MN) in zebrafish has been developed for studying neurological disorders. The motor neuronal zebrafish cell culture was initiated at 24 hpf when the axonal development and outgrowth of MN started, allowing the development of MN axons in vivo in the context of the normal endogenous signs of the model organism, while also providing availability for an in vitro system. The zebrafish’s primary culture techniques offer another approach to examine the neuronal population. There have been reports of primary neuron culture protocols ranging from blastula stage to 19 hpf [144,145,146], but these cultures are derived from MN axon pathfinding and neuromuscular development prior to normal course. Primary MN axons in zebrafish are present at 18 hpf out of the spinal cord, while the appearance of secondary MN axonal path findings range from 26 to 34 hpf [147,148]. The brain explant cultures [149] and primary cell culture of muscle fibers [150,151,152] are possible to develop from the later development stages of zebrafish embryogenesis. The advantages of primary zebrafish cell culture provide a new foundation to develop potential therapies for neurological disorders.
A new protocol [153] outlines how the subcellular spreading and protein aggregation status of neurodegenerative disease-causing neurons from transgenic zebrafish embryos can be investigated (Figure 3). ALS and spinocerebellar ataxia type-3 (SCA3) can be studied from this protocol, as the disease-causing sarcoma-fused (FUS) and ataxin-3 proteins of the human variant gene can be expressed in the zebrafish cell culture. A combination of neuronal subtypes, including motor neurons, exhibited cultural differentiation as well as an outgrowth of neurites. The human mutant FUS mislocated from nuclei to cytosol, imitating observed in human ALS and the zebrafish FUS model. In contrast, zebrafish-grown neurons expressing human ataxin-3 with disease-associated improved polyQ repeats did not build up in nuclei as frequently reported in SCA3. Another simple and efficient protocol was recently proposed to obtain the primary cells of embryonic zebrafish [134]. By exploiting the cell-type rich resource specific fluorescent zebrafish reporter lines, different types of differentiated cells were cultivated and monitored, proving that they continued their original morphology in culture for many days and demonstrated that before cultivation, particular types of cells could be enriched with flow cytometry. This group also successfully tested several fluorescent vital colors to facilitate subcellular imaging. This technique delivers a new tool to enhance our understanding of neurodegenerative disorders pathogenesis and help the development of mechanism-based drugs for neurological disorders.
6. Conclusions
In summary, several observational studies have shown a connection between zebrafish and human neurological disorders. Zebrafish are proving to be an ideal model for screening pharmaceutical agents prior to testing in rodents. The long-term aims of this work are to clarify the mechanisms of neurodegeneration and develop new neuroprotective compounds for the treatment of neurodegenerative diseases. In adult zebrafish, the approach of neurodegeneration using Aβ peptides can also help to design regenerative therapies in the neurodegenerative situation. The described culture of neuronal cells adds a new tool to investigate neurodegenerative diseases regarding molecular and cellular mechanisms, high-quality live cell imaging, and the discovery of new therapeutic drugs for neurological disorders. The primary embryo of zebrafish and larvae culture has the potential to provide tremendous knowledge regarding various mechanisms and treatments for human disease. Zebrafish-based assays are capable of promoting the bioassay-guided fractionation of great numbers of bioactive extracts identified in these in vivo screens, thus allowing the isolation of different novel, bioactive natural products—most of which are likely to be desirable lead compounds for the development of new, potent drugs. These initial studies support zebrafish in helping to resolve a crucial bottleneck in the discovery of NPs by allowing rapid in vivo, microgram-scale, bioassay-guided fractionation analysis, and diverse natural extract dereplication studies.
Author Contributions
The writing, review, and editing of manuscript were completed by A.P., R.K.R., and J.L.F.
Funding
Financial support was provided by the Science and Engineering Research Board-Purdue University Overseas Visiting Doctoral Fellowship (SERB-OVDF) (San. No: SB/S9/Z-03/2017-XXV (2018–2019), Dated: 21 December 2018), Council of Scientific and Industrial Research (CSIR) (File number: 09/1205 (0001) 2k18 EMR-I, Dated: 2 May 2018), and Department of Biotechnology (DBT) (San. No: BT/PR6765/NNT/28/618/2012).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Devine, M.J.; Plun-Favreau, H.; Wood, N.W. Parkinson’s disease and cancer: Two wars, one front. Nat. Rev. Cancer 2011, 11, 812–823. [Google Scholar] [CrossRef]
- Tabarés-seisdedos, R.; Rubenstein, J.L. Inverse cancer comorbidity a serendipitous opportunity to gain insight into CNS disorders. Nat. Rev. Neurosci. 2013, 14, 293–304. [Google Scholar] [CrossRef]
- Behrens, M.I.; Silva, M.; Salech, F.; Ponce, D.P.; Merino, D.; Sinning, M.; Xiong, C.; Roe, M.C.; Quest, A.F.G. Inverse susceptibility to oxidative death of lymphocytes obtained from Alzheimer’s patients and skin cancer survivors: Increased apoptosis in Alzheimer’s and reduced necrosis in cancer. J. Gerontol. Ser. A Biol Sci. Med. Sci. 2012, 67, 1036–1040. [Google Scholar] [CrossRef] [PubMed]
- Hilario, E.; Álvarez, A.; Domínguez, A.; Suarez-Merino, B.; Goñi-de-Cerio, F. Central nervous system diseases and the role of the blood-brain barrier in their treatment. Neurosci. Discov. 2013, 1, 1–3. [Google Scholar]
- Gilmore, J.L.; Yi, X.; Quan, L.; Kabanov, A.V. Novel Nanomaterials for Clinical Neuroscience. J. Neuroimmune Pharmacol. 2008, 3, 83–94. [Google Scholar] [CrossRef] [Green Version]
- Rosén, J.; Gottfries, J.; Muresan, S.; Backlund, A.; Oprea, T.I. Novel Chemical Space Exploration via Natural Products. J. Med. Chem. 2009, 52, 1953–1962. [Google Scholar] [CrossRef] [Green Version]
- Koehn, F.E.; Carter, G.T. The evolving role of natural products in drug discovery. Nat. Rev. Drug Discov. 2005, 4, 206–220. [Google Scholar] [CrossRef] [PubMed]
- Harvey, A.L. Natural products in drug discovery. Drug Discov. Today 2008, 13, 894–901. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural Products As Sources of New Drugs over the 30 Years from 1981 to 2010. J. Nat. Prod. 2012, 75, 311–335. [Google Scholar] [CrossRef] [Green Version]
- Macarron, R.; Banks, M.N.; Bojanic, D.; Burns, D.J.; Cirovic, D.A.; Garyantes, T.; Green, D.V.S.; Hertzberg, R.P.; Janzen, W.P.; Paslay, J.W.; et al. Impact of high-throughput screening in biomedical research. Nat. Rev. Drug Discov. 2011, 10, 188–195. [Google Scholar] [CrossRef]
- Welsch, M.E.; Snyder, S.A.; Stockwell, B.R. Privileged scaffolds for library design and drug discovery. Curr. Opin. Chem. Biol. 2010, 14, 347–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cordell, G.A. Biodiversity and drug discovery—A symbiotic relationship. Phytochemistry 2000, 55, 463–480. [Google Scholar] [CrossRef]
- Vuorela, P.; Tammela, P.; Rauha, J.; Saikku, P.; Leinonen, M.; Vuorela, H. Natural Products in the Process of Finding New Drug Candidates. Curr. Med. Chem. 2004, 11, 1375–1389. [Google Scholar] [CrossRef]
- Testa, R.; Bonfigli, A.; Genovese, S.; De Nigris, V.; Ceriello, A. The Possible Role of Flavonoids in the Prevention of Diabetic Complications. Nutrients 2016, 8, 310. [Google Scholar] [CrossRef] [PubMed]
- Zon, L.I.; Peterson, R.T. In vivo drug discovery in the zebrafish. Nat. Rev. Drug Discov. 2005, 4, 35–44. [Google Scholar] [CrossRef]
- Crawford, A.D.; Esguerra, C.V.; de Witte, P.A.M. Fishing for Drugs from Nature: Zebrafish as a Technology Platform for Natural Product Discovery. Planta Med. 2008, 74, 624–632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, J.-F.; Yoo, H.-D.; Williams, C.T.; Garo, E.; Cremin, P.A.; Zeng, L.; Vervoort, H.C.; Lee, C.M.; Hart, S.M.; Goering, M.G.; et al. Miniaturization of the Structure Elucidation of Novel Natural Products—Two Trace Antibacterial Acylated Caprylic Alcohol Glycosides from Arctostaphylos pumila. Planta Med. 2005, 71, 176–180. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.-K.; Lee, K.; More, S.V.; Kumar, H.; Kang, S.M.; Song, S.-Y. Advances in Neuroprotective Ingredients of Medicinal Herbs by Using Cellular and Animal Models of Parkinson’s Disease. Evid. Based Complement. Altern Med. 2013, 2013, 1–15. [Google Scholar]
- Zhang, Z.J.; Cheang, L.C.V.; Wang, M.W.; Lee, S.M.Y. Quercetin exerts a neuroprotective effect through inhibition of the iNOS/NO system and pro-inflammation gene expression in PC12 cells and in zebrafish. Int. J. Mol. Med. 2011, 27, 195–203. [Google Scholar]
- Chan, H.M.; Guo, B.J.; Sa, F.; Li, S.; Lee, S.M.Y.; Zhang, Z.J.; Zheng, Y. Pharmacokinetic Study and Optimal Formulation of New Anti-Parkinson Natural Compound Schisantherin, A. Parkinsons Dis. 2015, 2015, 1–7. [Google Scholar]
- Löscher, W.; Potschka, H. Role of drug efflux transporters in the brain for drug disposition and treatment of brain diseases. Prog. Neurobiol. 2005, 76, 22–76. [Google Scholar] [CrossRef] [PubMed]
- Jeffrey, P.; Summerfield, S. Assessment of the blood–brain barrier in CNS drug discovery. Neurobiol. Dis. 2010, 37, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Taylor, J.P.; Hardy, J.; Fischbeck, K.H. Toxic proteins in neurodegenerative disease. Science 2002, 296, 1991–1996. [Google Scholar] [CrossRef] [PubMed]
- Bandmann, O.; Burton, E.A. Genetic zebrafish models of neurodegenerative diseases. Neurobiol. Dis. 2010, 40, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Babin, P.J.; Goizet, C.; Raldúa, D. Zebrafish models of human motor neuron diseases: Advantages and limitations. Prog. Neurobiol. 2014, 118, 36–58. [Google Scholar] [CrossRef]
- Lemay, N.; Hayward, L.J.; Bosco, D.A.; Brown, R.H.; Zhou, H.; Burke, C.; Kwiatkowski, T.J.; Sapp, P., Jr.; Yasek, D.M.; Brown, R.H.; Hayward, L.J., Jr. Mutant FUS proteins that cause amyotrophic lateral sclerosis incorporate into stress granules. Hum. Mol. Genet. 2010, 19, 4160–4175. [Google Scholar] [Green Version]
- Lemmens, R.; Van Hoecke, A.; Hersmus, N.; Geelen, V.; D’Hollander, I.; Thijs, V.; Van Den Bosch, L.; Carmeliet, P.; Robberecht, W. Overexpression of mutant superoxide dismutase 1 causes a motor axonopathy in the zebrafish. Hum. Mol. Genet. 2007, 16, 2359–2365. [Google Scholar] [CrossRef] [Green Version]
- Laird, A.S.; Mackovski, N.; Rinkwitz, S.; Becker, T.S.; Giacomotto, J. Tissue-specific models of spinal muscular atrophy confirm a critical role of SMN in motor neurons from embryonic to adult stages. Hum. Mol. Genet. 2016, 25, 1728–1738. [Google Scholar] [CrossRef]
- Paquet, D.; Bhat, R.; Sydow, A.; Mandelkow, E.-M.; Berg, S.; Hellberg, S.; Fälting, J.; Schmid, B.; Haass, C. A zebrafish model of tauopathy allows in vivo imaging of neuronal cell death and drug evaluation Find the latest version: Technical advance A zebrafish model of tauopathy allows in vivo imaging of neuronal cell death and drug evaluation. J. Clin. Investig. 2009, 119, 1382–1395. [Google Scholar] [CrossRef]
- Miller, V.M.; Nelson, R.F.; Gouvion, C.M.; Williams, A.; Rodriguez-Lebron, E.; Harper, S.Q.; Davidson, B.L.; Rebagliati, M.R.; Paulson, H.L. CHIP Suppresses Polyglutamine Aggregation and Toxicity In Vitro and In Vivo. J. Neurosci. 2005, 25, 9152–9161. [Google Scholar] [CrossRef] [Green Version]
- McGown, A.; McDearmid, J.R.; Panagiotaki, N.; Tong, H.; Al Mashhadi, S.; Redhead, N.; Lyon, A.N.; Beattie, C.E.; Shaw, P.J.; Ramesh, T.M. Early interneuron dysfunction in ALS: Insights from a mutant sod1 zebrafish model. Ann. Neurol. 2013, 73, 246–258. [Google Scholar] [CrossRef] [PubMed]
- Kabashi, E.; Lin, L.; Tradewell, M.L.; Dion, P.A.; Bercier, V.; Bourgouin, P.; Rochefort, D.; Hadj, S.B.; Durham, H.D.; Rouleau, C.V.V.G.A.; et al. Gain and loss of function of ALS-related mutations of TARDBP (TDP-43) cause motor deficits in vivo. Hum. Mol. Genet. 2009, 19, 671–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, Q.; Burton, E.A. Zebrafish models of Tauopathy HHS Public Access. Biochim. Biophys. Acta 2011, 1812, 353–363. [Google Scholar] [CrossRef]
- Hartl, F.U.; Kretzschmar, H.; Haass, C.; Hirschberger, T.; Giese, A.; Schmid, B.; Schiffer, N.W.; Broadley, S.A.; Tavan, P. Identification of Anti-prion Compounds as Efficient Inhibitors of Polyglutamine Protein Aggregation in a Zebrafish Model. J. Biol. Chem. 2006, 282, 9195–9203. [Google Scholar]
- Charles, B.K. Patterning the Brain of the Zebrafish Embryo. Annu. Rev. Neurosci. 2002, 16, 707–732. [Google Scholar] [CrossRef] [PubMed]
- Wullimann, M.F.; Rupp, B.; Reichert, H.; Verlag, B. Neuroanatomy of the Zebrafish Brain: A Topological Atlas; Birkhäuser: Basel, Switzerland, 1996. [Google Scholar]
- Brown, J.; Guerra-Assunção, J.A.; Saunders, D.; Willey, D.; Barker, D.; Ellwood, M.; Gordon, D.; Chow, W.; Clark, R.; Karotki, L.; et al. The zebrafish reference genome sequence and its relationship to the human genome. Nature 2013, 496, 498–503. [Google Scholar]
- Kumar, S.; Hedges, B.S. Amolecular timescalefor vertebrateevolution Sudhir. Nature 1998, 392, 917–920. [Google Scholar] [CrossRef]
- Clarke, A.R. Transgenesis Techniques: Principles and Protocols; Springer: Berlin, Germany, 2003; p. 561. [Google Scholar]
- Hruscha, A.; Krawitz, P.; Rechenberg, A.; Heinrich, V.; Hecht, J.; Haass, C.; Schmid, B. Efficient CRISPR/Cas9 genome editing with low off-target effects in zebrafish. Development 2013, 140, 4982–4987. [Google Scholar] [CrossRef] [Green Version]
- Musa, A.; Lehrach, H.; Russo, V.E. Distinct expression patterns of two zebrafish homologues of the human APP gene during embryonic development. Dev. Genes Evol. 2001, 211, 563–567. [Google Scholar] [CrossRef]
- Leimer, U.; Lun, K.; Romig, H.; Walter, J.; Grünberg, J.; Brand, M.; Haass, C. Zebrafish (Danio rerio) Presenilin Promotes Aberrant Amyloid β-Peptide Production and Requires a Critical Aspartate Residue for Its Function in Amyloidogenesis. Biochemistry 1999, 38, 13602–13609. [Google Scholar] [CrossRef]
- Groth, C.; Nornes, S.; McCarty, R.; Tamme, R.; Lardelli, M. Identification of a second presenilin gene in zebrafish with similarity to the human Alzheimer’s disease gene presenilin2. Dev. Genes Evol. 2002, 212, 486–490. [Google Scholar] [CrossRef]
- Moussavi Nik, S.H.; Wilson, L.; Newman, M.; Croft, K.; Mori, T.A.; Musgrave, I.; Lardelli, M. The BACE1-PSEN-AβPP regulatory axis has an ancient role in response to low oxygen/oxidative stress. J. Alzheimer’s Dis. 2012, 28, 515–530. [Google Scholar] [CrossRef] [PubMed]
- Van Bebber, F.; Hruscha, A.; Willem, M.; Schmid, B.; Haass, C. Loss of Bace2 in zebrafish affects melanocyte migration and is distinct from Bace1 knock out phenotypes. J. Neurochem. 2013, 127, 471–481. [Google Scholar] [CrossRef] [PubMed]
- Francis, R.; McGrath, G.; Zhang, J.; Ruddy, D.A.; Sym, M.; Apfeld, J.; Nicoll, M.; Maxwell, M.; Hai, B.; Ellis, M.C.; et al. aph-1 and pen-2 are required for Notch pathway signaling, γ-secretase cleavage of βAPP, and presenilin protein accumulation. Dev. Cell 2002, 3, 85–97. [Google Scholar] [CrossRef]
- Campbell, W.A.; Yang, H.; Zetterberg, H.; Baulac, S.; Sears, J.A.; Liu, T.; Wong, S.T.C.; Zhong, T.P.; Xia, W. Zebrafish lacking Alzheimer presenilin enhancer 2 (Pen-2) demonstrate excessive p53-dependent apoptosis and neuronal loss. J. Neurochem. 2006, 96, 1423–1440. [Google Scholar] [CrossRef]
- Team MGC (MGC), P. Generation and initial analysis of more than 15,000 full-length human and mouse cDNA sequences. Proc. Natl. Acad. Sci. USA 2002, 99, 16899–16903. [Google Scholar]
- Babin, P.J.; Thisse, C.; Durliat, M.; Andre, M.; Akimenko, M.-A.; Thisse, B. Both apolipoprotein E and A-I genes are present in a nonmammalian vertebrate and are highly expressed during embryonic development. Proc. Natl. Acad. Sci. USA 1997, 94, 8622–8627. [Google Scholar] [CrossRef] [Green Version]
- Woods, I.G.; Wilson, C.; Friedlander, B.; Chang, P.; Reyes, D.K.; Nix, R.; Kelly, P.D.; Chu, F.; Postlethwait, J.H.; Talbot, W.S.; et al. The zebrafish gene map defines ancestral vertebrate chromosomes. Genome Res. 2005, 15, 1307–1314. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Peterson, S.M.; Freeman, J.L. Sex-specific characterization and evaluation of the Alzheimer’s disease genetic risk factor sorl1 in zebrafish during aging and in the adult brain following a 100 ppb embryonic lead exposure. J. Appl. Toxicol. 2017, 37, 400–407. [Google Scholar] [CrossRef]
- Bai, Q.; Mullett, S.J.; Garver, J.A.; Hinkle, D.A.; Burton, E.A. Zebrafish DJ-1 is evolutionarily conserved and expressed in dopaminergic neurons. Brain Res. 2006, 1113, 33–44. [Google Scholar] [CrossRef]
- Flinn, L.; Mortiboys, H.; Volkmann, K.; Kster, R.W.; Ingham, P.W.; Bandmann, O. Complex i deficiency and dopaminergic neuronal cell loss in parkin-deficient zebrafish (Danio rerio). Brain 2009, 132, 1613–1623. [Google Scholar] [CrossRef] [PubMed]
- Anichtchik, O.; Roach, A.; Goldsmith, P.; Diekmann, H.; Rubinsztein, D.C.; Fleming, A. Loss of PINK1 Function Affects Development and Results in Neurodegeneration in Zebrafish. J. Neurosci. 2008, 28, 8199–8207. [Google Scholar] [CrossRef]
- Ren, G.; Xin, S.; Li, S.; Zhong, H.; Lin, S. Disruption of lrrk2 does not cause specific loss of dopaminergic neurons in zebrafish. PLoS ONE 2011, 6, 2–7. [Google Scholar] [CrossRef] [PubMed]
- Karlovich, C.A.; John, R.M.; Ramirez, L.; Stainier, D.Y.; Myers, R.M. Characterization of the Huntington’s disease (HD) gene homolog in the zebrafish Danio rerio. Gene 1998, 217, 117–125. [Google Scholar] [CrossRef]
- Laboissonniere, L.A.; Smith, C.L.; Mesenbrink, J.; Chowdhury, R.; Burney, A.; Lang, M.; Sierra, M.; Stark, A.; Maldonado-Casalduc, G.; Muller, M.; et al. ALS-associated genes display CNS expression in the developing zebrafish. Gene Expr. Patterns 2018, 30, 14–31. [Google Scholar] [CrossRef]
- Schmid, B.; Hruscha, A.; Hogl, S.; Banzhaf-Strathmann, J.; Strecker, K.; van der Zee, J.; Teucke, M.; Eimer, S.; Hegermann, J.; Kittelmann, M.; et al. Loss of ALS-associated TDP-43 in zebrafish causes muscle degeneration, vascular dysfunction, and reduced motor neuron axon outgrowth. Proc. Natl. Acad. Sci. USA 2013, 110, 4986–4991. [Google Scholar] [CrossRef] [Green Version]
- Carlson, K.M.; Melcher, L.; Lai, S.; Zoghbi, H.Y.; Clark, H.B.; Orr, H.T. Characterization of the zebrafish atxn1/axh gene family. J. Neurogenet. 2009, 23, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Wood, J.D.; Bonath, F.; Kumar, S.; Ross, C.A.; Cunliffe, V.T. Disrupted-in-schizophrenia 1 and neuregulin 1 are required for the specification of oligodendrocytes and neurones in the zebrafish brain. Hum. Mol. Genet. 2009, 18, 391–404. [Google Scholar] [CrossRef]
- Kabashi, E.; Brustein, E.; Champagne, N.; Drapeau, P. Zebrafish models for the functional genomics of neurogenetic disorders. Biochim. Biophys. Acta 2011, 1812, 335–3345. [Google Scholar] [CrossRef]
- The State of the Art of Dementia Research: New Frontiers; World Alzheimer Report; Alzheimer’s Disease International: London, UK, 2018; pp. 1–48.
- Voisin, T.; Vellas, B. Diagnosis and Treatment of Patients with Severe Alzheimer’s Disease. Drugs Aging 2009, 26, 135–144. [Google Scholar] [CrossRef]
- Regeur, L.; Badsberg Jensen, G.; Pakkenberg, H.; Evans, S.M.; Pakkenberg, B. No global neocortical nerve cell loss in brains from patients with senile dementia of Alzheimer’s type. Neurobiol. Aging 1994, 15, 347–352. [Google Scholar] [CrossRef]
- Mark, J.; Paul, D.; Dorothy, G.; Juan, C. Differences in the pattern of hippocampal neuronal loss in normal ageing and Alzheimer â€TM s disease. Lancet 1994, 344, 769–772. [Google Scholar]
- Blennow, K.; De Leon, M.J.; Zetterberg, H. SeminarAlzheimer’s disease. Lancet 2006, 368, 1–17. [Google Scholar] [CrossRef]
- Jacobsen, L.; Madsen, P.; Jacobsen, C.; Nielsen, M.S.; Gliemann, J.; Petersen, C.M. Activation and Functional Characterization of the Mosaic Receptor SorLA/LR11. J. Biol. Chem. 2001, 276, 22788–22796. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Martins, R.N.; Lardelli, M. Complex splicing and neural expression of duplicated tau genes in zebrafish embryos. J. Alzheimer’s Dis. 2009, 18, 305–317. [Google Scholar] [CrossRef]
- Abeliovich, A.; Flint Beal, M. Parkinsonism genes: Culprits and clues. J. Neurochem. 2006, 99, 1062–1072. [Google Scholar] [CrossRef]
- Galvin, J.E. Cognitive change in Parkinson disease. Alzheimer Dis. Assoc. Disord. 2006, 20, 302–310. [Google Scholar] [CrossRef] [PubMed]
- Huntington, T.; Macdonald, M.E.; Ambrose, C.M.; Duyao, M.P.; Myers, R.H.; Lin, C.; Srinidhi, L.; Barnes, G.; Taylor, S.A.; James, M.; et al. A Novel Gene Containing a Trinucleotide Repeat That Is Expanded and Unstable on Huntington’s Disease Chromosomes. Cell 1993, 72, 971–983. [Google Scholar]
- Walker, F.O. Huntington’s disease. Lancet 2007, 369, 218–228. [Google Scholar] [CrossRef]
- Lumsden, A.L.; Henshall, T.L.; Dayan, S.; Lardelli, M.T.; Richards, R.I. Huntingtin-deficient zebrafish exhibit defects in iron utilization and development. Hum. Mol. Genet. 2007, 16, 1905–1920. [Google Scholar] [CrossRef] [Green Version]
- Diekmann, H.; Fleming, A.; Rubinsztein, D.C.; Roach, A.; Anichtchik, O.; Goldsmith, P.; Futter, M. Decreased BDNF Levels Are a Major Contributor to the Embryonic Phenotype of Huntingtin Knockdown Zebrafish. J. Neurosci. 2009, 29, 1343–1349. [Google Scholar] [CrossRef] [Green Version]
- Deng, H.X.; Hentati, A.; Tainer, J.A.; Iqbal, Z.; Cayabyab, A.; Hung, W.Y.; Getzoff, E.D.; Hu, P.; Herzfeldt, B.; Roos, R.P. Amyotrophic lateral sclerosis and structural defects in Cu,Zn superoxide dismutase. Science 1993, 261, 1047–1051. [Google Scholar] [CrossRef]
- Rosen, D.R.; Siddiquet, T.; Pattersont, D.; Figlewicz, D.A.; Ii, P.S.; Hentatit, A.; Donaldsont, D.; Goto, J.; Ii, J.P.O.R.; Dengt, H.; et al. Mutations in Cu/Zn superoxide dismutase gene are associated with familial amyotrophic lateral sclerosis. Nature 1993, 362, 59–62. [Google Scholar] [CrossRef] [PubMed]
- Turner, B.J.; Talbot, K. Transgenics, toxicity and therapeutics in rodent models of mutant SOD1-mediated familial ALS. Prog. Neurobiol. 2008, 85, 94–134. [Google Scholar] [CrossRef]
- Weinberger, D. From neuropathology to neurodevelopment. Lancet 1995, 346, 552–557. [Google Scholar] [CrossRef]
- Nuechterlein, K.H.; Barch, M.; Gold, J.M.; Goldberg, T.E.; Green, M.F.; Heaton, R.K. Identification of separable cognitive factors in schizophrenia. Schizophr. Res. 2004, 72, 29–39. [Google Scholar] [CrossRef]
- Swerdlow, N.R.; Butters, N.; Geyer, M.A.; Paulsen, J.; Swenson, M.R.; Braff, D.L. Impaired prepulse inhibition of acoustic and tactile startle response in patients with Huntington’s disease. J. Neurol. Neurosurg. Psychiatry 2008, 58, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Braff, D.L.; Geyer, M.A.; Swerdlow, N.R. Human studies of prepulse inhibition of startle: Normal subjects, patient groups, and pharmacological studies. Psychopharmacology 2001, 156, 234–258. [Google Scholar] [CrossRef]
- Braff, D.; Schork, N.J. Gottesman, I.I. Endophenotyping schizophrenia. Am. J. Psychiatry 2007, 164, 705–707. [Google Scholar] [CrossRef] [PubMed]
- Burgess, H.A.; Granato, M. Sensorimotor Gating in Larval Zebrafish. J. Neurosci. 2007, 27, 4984–4994. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, P.F.; Kendler, K.S.; Neale, M.C. Schizophrenia as a Complex Trait. Arch. Gen. Psychiatry 2003, 60, 1187–1192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Rensburg, E.J.; Xu, B.; Karayiorgou, M.; Levy, S.; Roos, J.L.; Gogos, J.A. Strong association of de novo copy number mutations with sporadic schizophrenia. Nat. Genet. 2008, 40, 880–885. [Google Scholar]
- Tarabeux, J.; Champagne, N.; Brustein, E.; Hamdan, F.F.; Gauthier, J.; Lapointe, M.; Maios, C.; Piton, A.; Spiegelman, D.; Henrion, É.M.; Millet, B.; et al. De Novo Truncating Mutation in Kinesin 17 Associated with Schizophrenia. Biol. Psychiatry 2010, 68, 649–656. [Google Scholar] [CrossRef]
- Girard, S.L.; Dion, P.A.; Rouleau, G.A. Schizophrenia genetics: Putting all the pieces together. Curr. Neurol. Neurosci. Rep. 2012, 12, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Morris, J.A. Zebrafish: A model system to examine the neurodevelopmental basis of schizophrenia. Prog. Brain Res. 2009, 179, 97–106. [Google Scholar] [PubMed]
- Burgess, H.A.; Granato, M. Modulation of locomotor activity in larval zebrafish during light adaptation. J. Exp. Biol. 2007, 210, 2526–2539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millar, J.K.; Christie, S.; Semple, C.A.M.; Porteous, D.J. Chromosomal Location and Genomic Structure of the Human Translin-Associated Factor X Gene (TRAX; TSNAX) Revealed by Intergenic Splicing to DISC1, a Gene Disrupted by a Translocation Segregating with Schizophrenia. Genomics 2000, 67, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Zashikhina, A. Juvenile Chronic Physical Illness in Northern Russia: Studies on Mental Health, Health-Related Quality of Life, and Family Functioning. Ph.D. Thesis, Umeå Universitet, Umeå, Sweden, 2014. [Google Scholar]
- Galanopoulou, A.S.; Buckmaster, P.S.; Staley, K.J.; Moshe, S.L.; Perucca, E.; Engel, J.; Löscher, W.; Noebels, J.L.; Pitkänen, A.; Stables, J.; White, H.S.; et al. Identification of new epilepsy treatments: Issues in preclinical methodology. Epilepsia 2012, 53, 571–582. [Google Scholar] [CrossRef]
- Desmond, D.; Kyzar, E.; Gaikwad, S.; Green, J.; Riehl, R.; Roth, A.; Stewart, A.M.; Kalueffet, A.V. Assessing epilepsy-related behavioral phenotypes in adult zebrafish. In Zebrafish Protocols for Neurobehavioral Research; Springer: Berlin, Germany, 2012; pp. 313–322. [Google Scholar]
- Crawford, A.D.; Liekens, S.; Kamuhabwa, A.R.; Maes, J.; Munck, S.; Busson, R.; Rozenski, J.; Esguerra, C.V.; de Witte, P.A.M. Zebrafish bioassay-guided natural product discovery: Isolation of angiogenesis inhibitors from East African medicinal plants. PLoS ONE 2011, 6, e14694. [Google Scholar] [CrossRef] [PubMed]
- Marcourt, L.; Bock, M.; Maljevic, S.; Dayrit, F.M.; Challal, S.; Queiroz, E.F.; de Witte, P.A.M.; Crawford, A.D.; Harvey, A.L.; Buenafe, O.E.M.; et al. Zebrafish Bioassay-Guided Microfractionation Identifies Anticonvulsant Steroid Glycosides from the Philippine Medicinal Plant Solanum torvum. ACS Chem. Neurosci. 2014, 5, 993–1004. [Google Scholar]
- Pitchai, A.; Nagarajan, N.; Vincent, S.G.P.; Rajaretinam, R.K. Zebrafish bio-assay guided isolation of human acetylcholinesterase inhibitory trans-tephrostachin from Tephrosia purpurea (L.) Pers. Neurosci. Lett. 2018, 687, 268–275. [Google Scholar] [CrossRef] [PubMed]
- Buenafe, O.E.; Orellana-Paucar, A.; Maes, J.; Huang, H.; Ying, X.; De Borggraeve, W.; Crawford, A.D.; Luyten, W.; Esguerra, C.V.; De Witte, P. Tanshinone IIA exhibits anticonvulsant activity in zebrafish and mouse seizure models. ACS Chem. Neurosci. 2013, 4, 1479–1487. [Google Scholar] [CrossRef] [PubMed]
- Esguerra, C.V.; Kamuhabwa, A.R.; Wolfender, J.-L.; Moshi, M.J.; Siverio-Mota, D.; Marcourt, L.; Maes, J.; Bohni, N.; de Witte, P.A.M.; Cordero-Maldonado, M.L.; et al. Integration of Microfractionation, qNMR and Zebrafish Screening for the In Vivo Bioassay-Guided Isolation and Quantitative Bioactivity Analysis of Natural Products. PLoS ONE 2013, 8, e64006. [Google Scholar]
- Jones, R.W.; Huffman, M.N. Fish Embryos as Bio-Assay Material in Testing Chemicals for Effects on Cell Division and Differentiation. Trans. Am. Microsc. Soc. 1957, 76, 177–183. [Google Scholar] [CrossRef]
- Liu, M.; Copmans, D.; Lu, J.G.; Yang, M.R.; Sourbron, J.; Ny, A.; Jiang, Z.H.; de Witte, P.A.M.; Luyten, W. Bioassay-guided isolation of anti-seizure principles from Semen Pharbitidis using a zebrafish pentylenetetrazol seizure model. J. Ethnopharmacol. 2019, 232, 130–134. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.L.; Cheng, L.; Kwok, H.F.; Ko, C.H.; Lau, T.W.; Koon, C.M.; Zhao, M.; Lau, C.P.; Lau, K.M.; Wong, C.W.; et al. Bioassay-guided isolation of norviburtinal from the root of Rehmannia glutinosa, exhibited angiogenesis effect in zebrafish embryo model. J. Ethnopharmacol. 2011, 137, 1323–1327. [Google Scholar] [CrossRef]
- Liang, F.; Han, Y.; Gao, H.; Xin, S.; Chen, S.; Wang, N.; Qin, W.; Zhong, H.; Lin, S.; Yao, X.; et al. Kaempferol Identified by Zebrafish Assay and Fine Fractionations Strategy from Dysosma versipellis Inhibits Angiogenesis through VEGF and FGF Pathways. Sci. Rep. 2015, 5, 1–10. [Google Scholar] [CrossRef]
- Cheng, M.-C.; Lee, T.-H.; Chu, Y.-T.; Syu, L.-L.; Hsu, S.-J.; Cheng, C.-H.; Lee, J.; Wu, C.-K. Melanogenesis Inhibitors from the Rhizoma of Ligusticum Sinense in B16-F10 Melanoma Cells In Vitro and Zebrafish In Vivo. Int. J. Mol. Sci. 2018, 19, 3994. [Google Scholar] [CrossRef]
- Brillatz, T.; Lauritano, C.; Jacmin, M.; Khamma, S.; Marcourt, L.; Righi, D.; Romano, G.; Esposito, F.; Ianora, A.; Queiroz, E.F.; et al. Zebrafish-based identification of the antiseizure nucleoside inosine from the marine diatom Skeletonema marinoi. PLoS ONE 2018, 13, e0196195. [Google Scholar] [CrossRef]
- Thermes, V.; Grabher, C.; Ristoratore, F.; Bourrat, F.; Choulika, A.; Wittbrodt, J.; Joly, J.-S.; Thermes, V.; Grabher, C.; Ristoratore, F.; et al. I-SceI meganuclease mediates highly efficient transgenesis in fish. Mech. Dev. 2002, 118, 91–98. [Google Scholar] [CrossRef]
- Kawakami, K.; Shima, A.; Kawakami, N. Identification of a functional transposase of the Tol2 element, an Ac-like element from the Japanese medaka fish, and its transposition in the zebrafish germ lineage. Proc. Natl. Acad. Sci. USA 2002, 97, 11403–11408. [Google Scholar] [CrossRef]
- Hans, S.; Kaslin, J.; Freudenreich, D.; Brand, M. Temporally-controlled site-specific recombination in zebrafish. PLoS ONE 2009, 4, e4640. [Google Scholar] [CrossRef]
- Halpern, M.E.; Rhee, J.; Goll, M.G.; Akitake, C.M.; Parsons, M.; Leach, S.D. Gal4/UAS Transgenic Tools and Their Application to Zebrafish. Zebrafish 2008, 5, 97–110. [Google Scholar] [CrossRef]
- Schmid, B.; Haass, C. Genomic editing opens new avenues for zebrafish as a model for neurodegeneration. J. Neurochem. 2013, 127, 461–470. [Google Scholar] [CrossRef]
- Hwang, W.Y.; Fu, Y.; Reyon, D.; Maeder, ML.; Tsai, S.Q.; Sander, J.D.; Joung, J.K.; Peterson, R.T.; Yeh, J.R.J. Efficient genome editing in zebrafish using a CRISPR-Cas system. Nat. Biotechnol. 2013, 31, 227–229. [Google Scholar] [CrossRef]
- Burns, C.G.; Milan, D.J.; Grande, E.J.; Rottbauer, W.; MacRae, C.A.; Fishman, M.C. High-throughput assay for small molecules that modulate zebrafish embryonic heart rate. Nat. Chem. Biol. 2005, 1, 263–264. [Google Scholar] [CrossRef] [PubMed]
- Goldsmith, P. Zebrafish as a pharmacological tool: The how, why and when. Curr. Opin. Pharmacol. 2004, 4, 504–512. [Google Scholar] [CrossRef]
- Sun, Z.; Chen, W.; Farrington, S.; Haldi, M.; Hopkins, N.; Amsterdam, A.; Golling, G.; Townsend, K.; Burgess, S. A large-scale insertional mutagenesis screen in zebrafish. Genes Dev. 2002, 13, 2713–2724. [Google Scholar]
- Guo, S.; Wilson, S.W.; Cooke, S.; Chitnis, A.B.; Driever, W.; Rosenthal, A. Mutations in the Zebrafish Unmask Shared Regulatory Pathways Controlling the Development of Catecholaminergic Neurons. Dev. Biol. 1999, 208, 473–487. [Google Scholar] [CrossRef] [Green Version]
- Amatruda, J.F.; Tallarico, J.A.; Weber, G.; Zon, L.I.; Pfaff, K.L.; King, R.W.; Straub, C.T.; Shepard, J.L.; Murphey, R.D.; Stern, H.M. Small molecules that delay S phase suppress a zebrafish bmyb mutant. Nat. Chem. Biol. 2005, 1, 366–370. [Google Scholar]
- MacRae, C.A.; Milan, D.J.; Fishman, M.C.; Schreiber, S.L.; Peterson, R.T.; Peterson, T.A.; Shaw, S.Y.; Zhong, T.P. Chemical suppression of a genetic mutation in a zebrafish model of aortic coarctation. Nat. Biotechnol. 2004, 22, 595–599. [Google Scholar]
- Song, P.; Pimplikar, S.W. Knockdown of amyloid precursor protein in zebrafish causes defects in motor axon outgrowth. PLoS ONE 2012, 7, e34209. [Google Scholar] [CrossRef] [PubMed]
- Prabhudesai, S.; Bensabeur, F.Z.; Abdullah, R.; Basak, I.; Baez, S.; Alves, G.; Holtzman, N.G.; Larsen, J.P.; Møller, S.G. LRRK2 knockdown in zebrafish causes developmental defects, neuronal loss, and synuclein aggregation. J. Neurosci. Res. 2016, 94, 717–735. [Google Scholar] [CrossRef]
- Wang, W.Y.; Fu, Y.; Reyon, D.; Maeder, M.L.; Kaini, P.; Sander, J.D.; Joung, J.K.; Peterson, R.T.; Yeh, J.R.J. Heritable and Precise Zebrafish Genome Editing Using a CRISPR-Cas System. PLoS ONE 2013, 8, e68708. [Google Scholar]
- Armstrong, G.A.B.; Liao, M.; You, Z.; Lissouba, A.; Chen, B.E.; Drapeau, P. Homology directed knockin of point mutations in the zebrafish tardbp and fus genes in ALS using the CRISPR/Cas9 system. PLoS ONE 2016, 11, e0150188. [Google Scholar] [CrossRef] [PubMed]
- Nery, L.R.; Silva, N.E.; Fonseca, R.; Vianna, M.R.M. Presenilin-1 Targeted Morpholino Induces Cognitive Deficits, Increased Brain Aβ1−42 and Decreased Synaptic Marker PSD-95 in Zebrafish Larvae. Neurochem. Res. 2017, 42, 2959–2967. [Google Scholar] [CrossRef] [PubMed]
- Peri, F.; Nüsslein-Volhard, C. Live Imaging of Neuronal Degradation by Microglia Reveals a Role for v0-ATPase a1 in Phagosomal Fusion In Vivo. Cell 2008, 133, 916–927. [Google Scholar] [CrossRef] [Green Version]
- Bretaud, S.; Allen, C.; Ingham, P.W.; Bandmann, O. p53-dependent neuronal cell death in a DJ-1-deficient zebrafish model of Parkinson’s disease. J. Neurochem. 2007, 100, 1626–1635. [Google Scholar] [CrossRef]
- Fett, M.E.; Pilsl, A.; Paquet, D.; van Bebber, F.; Haass, C.; Tatzelt, J.; Schmid, B.; Winklhofer, K.F. Parkin is protective against proteotoxic stress in a transgenic zebrafish model. PLoS ONE 2010, 5, e11783. [Google Scholar] [CrossRef]
- Zhang, Y.; Nguyen, D.T.; Olzomer, E.M.; Poon, G.P.; Cole, N.J.; Puvanendran, A.; Phillips, B.R.; Hesselson, D. Rescue of Pink1 Deficiency by Stress-Dependent Activation of Autophagy. Cell Chem. Biol. 2017, 24, 471–480. [Google Scholar] [CrossRef]
- Tanaka, E.M.; Ferretti, P. Considering the evolution of regeneration in the central nervous system. Nat. Rev. Neurosci. 2009, 10, 713–723. [Google Scholar] [CrossRef]
- Chapouton, P.; Jagasia, R.; Bally-Cuif, L. Adult neurogenesis in non-mammalian vertebrates. BioEssays 2007, 29, 745–757. [Google Scholar] [CrossRef] [Green Version]
- Kizil, C.; Kaslin, J.; Kroehne, V.; Brand, M. Adult neurogenesis and brain regeneration in zebrafish. Dev. Neurobiol. 2012, 72, 429–461. [Google Scholar] [CrossRef] [PubMed]
- Cosacak, M.I.; Papadimitriou, C.; Kizil, C. Regeneration, Plasticity, and Induced Molecular Programs in Adult Zebrafish Brain. Biomed. Res. Int. 2015, 2015, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, S.; Basu, A. Inflammation: A new candidate in modulating adult neurogenesis. J. Neurosci. Res. 2008, 86, 1199–1208. [Google Scholar] [CrossRef] [PubMed]
- Kizil, C.; Kyritsis, N.; Brand, M. Effects of inflammation on stem cells: Together they strive? EMBO Rep. 2015, 16, 416–426. [Google Scholar] [CrossRef] [PubMed]
- Kizil, C.; Brand, M. Cerebroventricular microinjection (CVMI) into adult zebrafish brain is an efficient misexpression method for forebrain ventricular cells. PLoS ONE 2011, 6, e27395. [Google Scholar] [CrossRef] [PubMed]
- Bhattarai, P.; Thomas, A.K.; Cosacak, M.I.; Papadimitriou, C.; Mashkaryan, V.; Froc, C.; Reinhardt, S.; Kurth, T.; Dahl, T.; Zhang, Y.; et al. IL4/STAT6 Signaling Activates Neural Stem Cell Proliferation and Neurogenesis upon Amyloid-β42 Aggregation in Adult Zebrafish Brain. Cell Rep. 2016, 17, 941–948. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.W.; Quaranta, V.; Glenner, G.G. Neuritic plaques and cerebrovascular amyloid in Alzheimer disease are antigenically related. Proc. Natl. Acad. Sci. USA 1985, 82, 8729–8732. [Google Scholar] [CrossRef]
- Masters, C.L.; Simms, G.; Weinman, N.A.; Multhaup, G.; McDonald, B.L.; Beyreuther, K. Amyloid plaque core protein in Alzheimer disease and Down syndrome. Proc. Natl. Acad. Sci. USA 1985, 82, 4245–4249. [Google Scholar] [CrossRef]
- Younkin, S.G. The role of Aβ42 in Alzheimer’s disease. J. Physiol. Paris 1998, 92, 289–292. [Google Scholar] [CrossRef]
- Liu, W.; Guan, Y.; Collodi, P. A Zebrafish Cell Culture Assay for the Identification of MicroRNA Targets. Zebrafish 2010, 7, 343–348. [Google Scholar] [CrossRef] [PubMed]
- Myhre, J.L.; Pilgrim, D.B. Cellular Differentiation in Primary Cell Cultures from Single Zebrafish Embryos as a Model for the Study of Myogenesis. Zebrafish 2010, 7, 255–266. [Google Scholar] [CrossRef]
- Robles, V.; Martí, M.; Belmonte, J.C.I. Study of Pluripotency Markers in Zebrafish Embryos and Transient Embryonic Stem Cell Cultures. Zebrafish 2011, 8, 57–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakowski, S.A.; Lunn, J.S.; Busta, A.S.; Palmer, M.; Dowling, J.J.; Feldman, E.L. A novel approach to study motor neurons from zebrafish embryos and larvae in culture. J. Neurosci. Methods 2012, 205, 277–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciarlo, C.A.; Zon, L. Embryonic cell culture in zebrafish. Methods Cell Biol. 2016, 133, 1–10. [Google Scholar]
- Sassen, W.A.; Lehne, F.; Russo, G.; Wargenau, S.; Dübel, S.; Köster, R.W. Embryonic zebrafish primary cell culture for transfection and live cellular and subcellular imaging. Dev. Biol. 2017, 430, 18–31. [Google Scholar] [CrossRef]
- Giros, B.; Fassier, C.; Nothias, F.; Lumsden, A.; Hazan, J.; Scholpp, S.; Hutt, J.A.; Houart, C.; Schneider-Maunoury, S. Zebrafish atlastin controls motility and spinal motor axon architecture via inhibition of the BMP pathway. Nat. Neurosci. 2010, 13, 1380–1387. [Google Scholar]
- Liu, D.; Westerfield, M. Clustering of muscle acetylcholine receptors requires motoneurons in live embryos, but not in cell culture. J. Neurosci. 2018, 12, 1859–1866. [Google Scholar] [CrossRef]
- Ghosh, C.; Liu, Y.; Ma, C.; Collodi, P. Cell cultures derived from early zebrafish embryos differentiate in vitro into neurons and astrocytes. Cytotechnology 1997, 23, 221–230. [Google Scholar] [CrossRef]
- Andersen, S.S.L. Preparation of dissociated Zebrafish spinal neuron cultures. Methods Cell Sci. 2001, 23, 205–209. [Google Scholar] [CrossRef] [PubMed]
- Beattie, C.E. Control of motor axon guidance in the zebrafish embryo. Brain Res. Bull. 2000, 53, 489–500. [Google Scholar] [CrossRef]
- Eisen, J.S. Motoneuronal development in the embryonic zebrafish. Development 1991, 2, 141–147. [Google Scholar]
- Hendricks, M.; Jesuthasan, S. Electroporation-based methods for in vivo, whole mount and primary culture analysis of zebrafish brain development. Neural Dev. 2007, 2, 1–10. [Google Scholar] [CrossRef]
- Saint-amant, L.; Sprague, S.M.; Hirata, H.; Li, Q.; Cui, W.W.; Zhou, W.; Poudou, O.; Hume, R.I.; Kuwada, J.Y. The Zebrafish ennui Behavioral Mutation Disrupts Acetylcholine Receptor Localization and Motor Axon Stability. Dev. Neurobiol. 2007, 68, 45–61. [Google Scholar] [CrossRef]
- Currie, P.D.; Ferguson, C.; Westerfield, M.; Key, B.; Nixon, S.J.; Méry, P.-F.; Parton, R.G.; Hancock, J.F.; Wegner, J. Zebrafish as a model for caveolin-associated muscle disease; caveolin-3 is required for myofibril organization and muscle cell patterning. Hum. Mol. Genet. 2005, 14, 1727–1743. [Google Scholar] [Green Version]
- Vreede, A.P.; Dowling, J.J.; Low, S.E.; Feldman, E.L.; Bonnemann, C.G.; Gibbs, E.M.; Kuwada, J.Y. Loss of Myotubularin Function Results in T-Tubule Disorganization in Zebrafish and Human Myotubular Myopathy. PLoS Genet. 2009, 5, e1000372. [Google Scholar]
- Don, E.K.; Watchon, M.; Yuan, K.C.; Fifita, J.A.; Blair, I.P.; Nicholson, G.A.; Laird, A.S.; Acosta, J.R.; Goldsbury, C.; Svahn, A.J.; et al. Neuronal cell culture from transgenic zebrafish models of neurodegenerative disease. Biol. Open 2018, 7, bio036475. [Google Scholar] [Green Version]
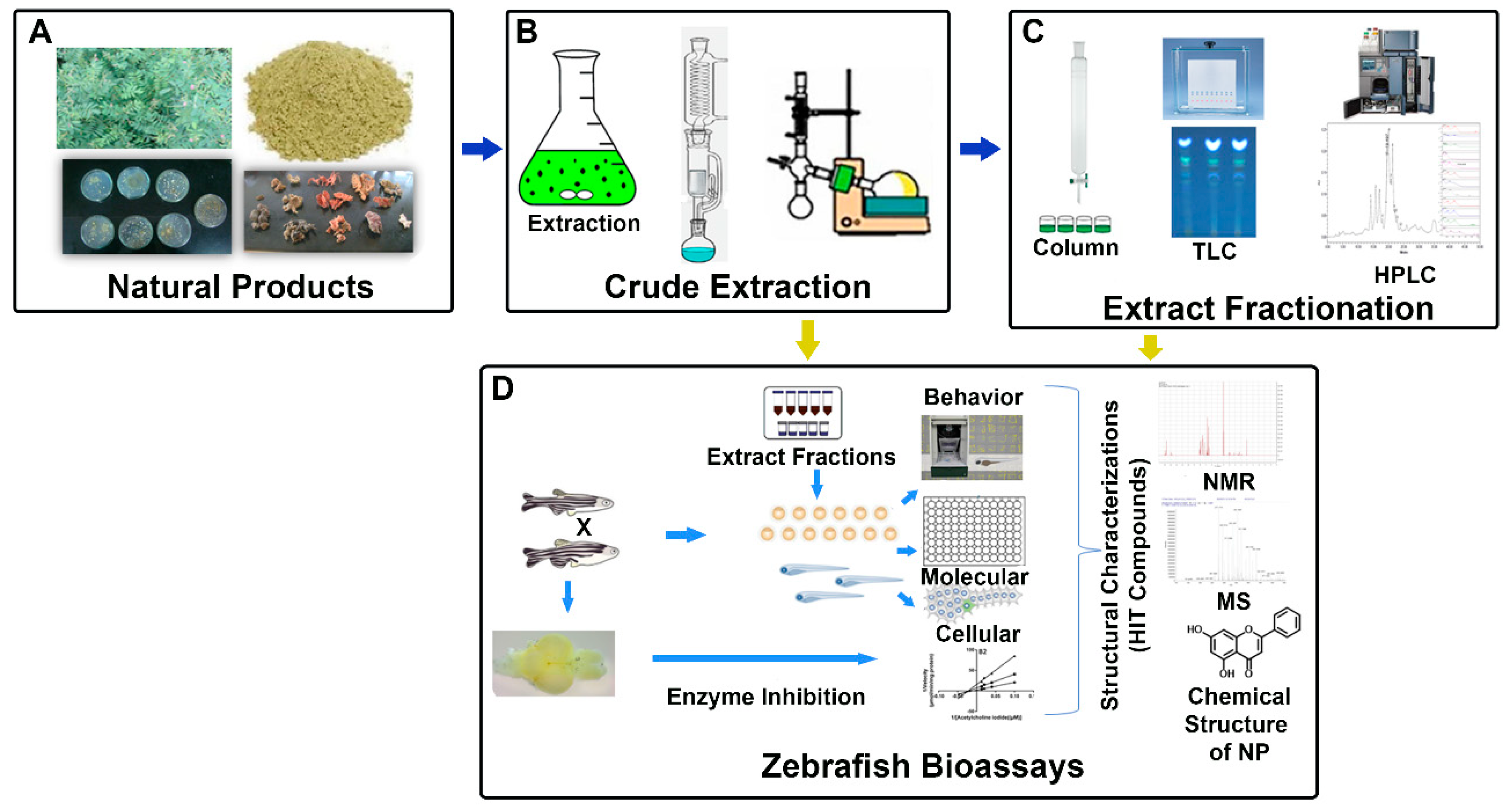
Figure 1.
Schematic representation of zebrafish bioassay-guided isolation of natural products. (A) Different sources of natural products. (B) Crude extraction of natural products. (C) Purification of biologically active compounds from various chromatographic methods. (D) Various zebrafish biological assays and structural characterization of HIT compounds using different spectroscopic techniques. (HPLC: high performance liquid chromatography; MS: mass spectrometry; NMR: nuclear magnetic resonance spectrometry; NPs: natural products; and TLC: thin-layer chromatography).
Figure 1.
Schematic representation of zebrafish bioassay-guided isolation of natural products. (A) Different sources of natural products. (B) Crude extraction of natural products. (C) Purification of biologically active compounds from various chromatographic methods. (D) Various zebrafish biological assays and structural characterization of HIT compounds using different spectroscopic techniques. (HPLC: high performance liquid chromatography; MS: mass spectrometry; NMR: nuclear magnetic resonance spectrometry; NPs: natural products; and TLC: thin-layer chromatography).

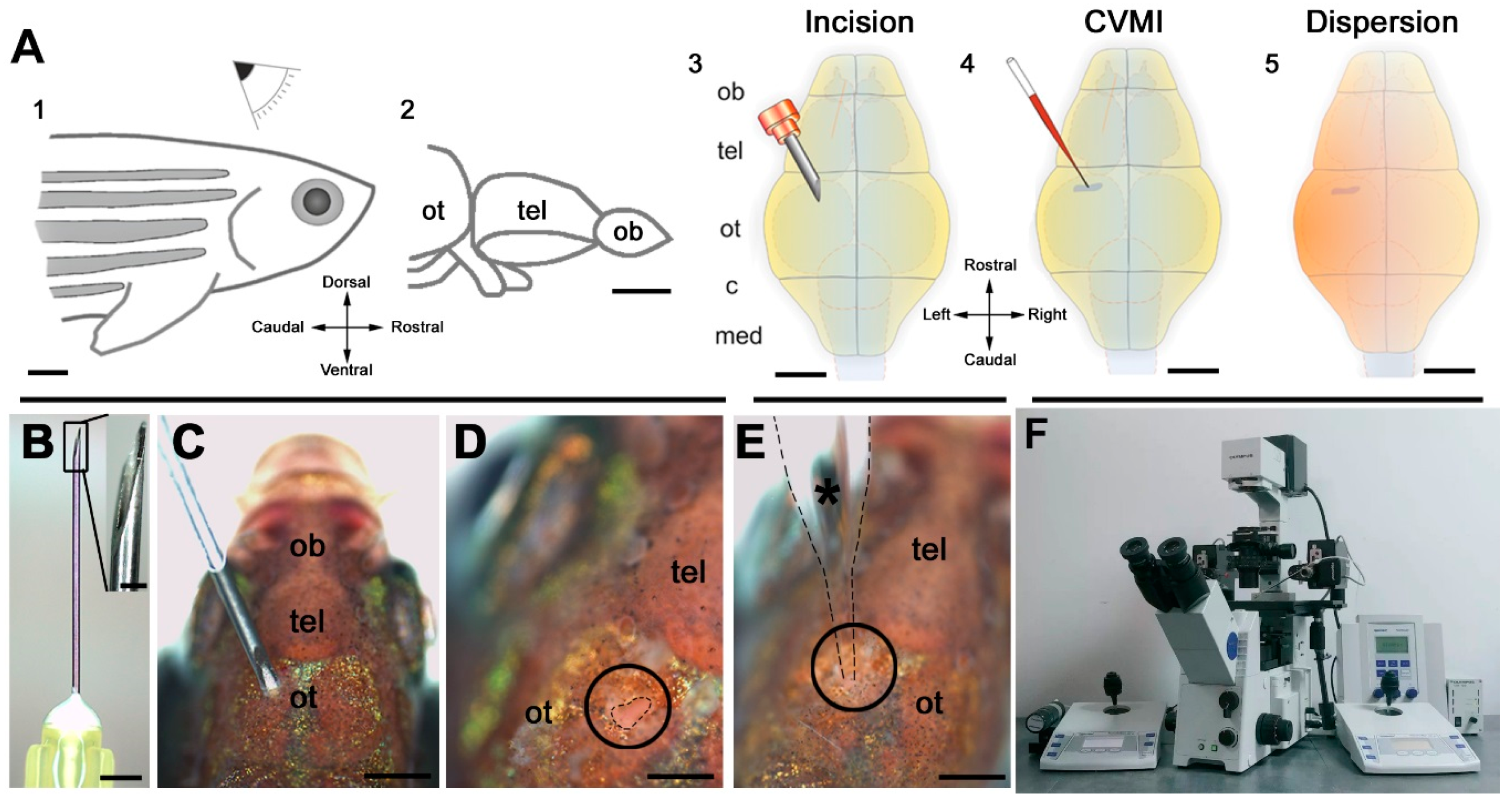
Figure 2.
Outline of the pattern and its target regions of cerebroventricular microinjection (CVMI). (A) CVMI is achieved at the dorsal surface of the head (1) and targets, in this example, the forebrain that is rostral to the optic tectum (2). For injection, an incision is made into the skull over the optic tectum using a barbed-end canula (3). Through this slit, liquid is injected using a glass capillary (4). Injected liquid disperses rostrally (5). (B) The canula used for incision. (C) The incision on an adult fish (dorsal view). (D) The incision site marked by dashed lines. (E) Injection with the glass capillary (*) (dotted lines mark the outline). (F) Injection apparatus. Images (A–E) are adapted from [132]. (c: cerebellum; med: medulla; ob: olfactory bulb; ot: optic tectum; and tel: telencephalon)
Figure 2.
Outline of the pattern and its target regions of cerebroventricular microinjection (CVMI). (A) CVMI is achieved at the dorsal surface of the head (1) and targets, in this example, the forebrain that is rostral to the optic tectum (2). For injection, an incision is made into the skull over the optic tectum using a barbed-end canula (3). Through this slit, liquid is injected using a glass capillary (4). Injected liquid disperses rostrally (5). (B) The canula used for incision. (C) The incision on an adult fish (dorsal view). (D) The incision site marked by dashed lines. (E) Injection with the glass capillary (*) (dotted lines mark the outline). (F) Injection apparatus. Images (A–E) are adapted from [132]. (c: cerebellum; med: medulla; ob: olfactory bulb; ot: optic tectum; and tel: telencephalon)

Figure 3.
Summary of workflow for culturing zebrafish neurons. Embryos (24 hpf or 48 hpf) are collected, dechorionated, and placed in E3 medium and 16 μM tricaine microtubes. Embryos are then washed three times with an ice-cold E3 medium before being placed in 1 trypsin (in PBS) and pipeted for 30 minutes intermittently in a 37 °C water bath. To stop separation, fetal bovine serum (FBS) is then added and the tubes are centrifuged at 1000 rpm for 3 minutes. The supernatant is removed, and cell pellet resuspended in Hanks’ balanced salt solution (HBSS). Cells are counted using a hematocytometer, and ~500,000 cells are placed per well for culturing. It is recommended to change half of the media daily. (hpf: hours post fertilization)
Figure 3.
Summary of workflow for culturing zebrafish neurons. Embryos (24 hpf or 48 hpf) are collected, dechorionated, and placed in E3 medium and 16 μM tricaine microtubes. Embryos are then washed three times with an ice-cold E3 medium before being placed in 1 trypsin (in PBS) and pipeted for 30 minutes intermittently in a 37 °C water bath. To stop separation, fetal bovine serum (FBS) is then added and the tubes are centrifuged at 1000 rpm for 3 minutes. The supernatant is removed, and cell pellet resuspended in Hanks’ balanced salt solution (HBSS). Cells are counted using a hematocytometer, and ~500,000 cells are placed per well for culturing. It is recommended to change half of the media daily. (hpf: hours post fertilization)


{kind=link}
{kind=link}
{kind=link}
Table 1.
Zebrafish orthologs of human genes involved in neurodegenerative disease pathogenesis.
| Disease | Protein | Human Gene | Zebrafish Gene | Amino Acid Similarity (%) | Reference |
|---|---|---|---|---|---|
| Alzheimer’s Disease | Amyloid precursor protein | APP GeneID: 351 Locus: 21q21.2 Protein length: 695 | appa GeneID: 58083 Chromosome: 1 Protein length: 738 | 74 | [41] |
| appb GeneID: 170846 Chromosome: 9 Protein length: 694 | 77 | ||||
| Presenilin-1 | PSEN1 GeneID: 5663 Locus: 14q24.3 Protein length: 467 | psen1 GeneID: 30221 Chromosome: 17 Protein length: 456 | 75 | [42] | |
| Presenilin-2 | PSEN2 GeneID: 5664 Locus: 1q31-q42 Protein length: 448 | psen2 GeneID: 58026 Chromosome: 1 Protein length: 441 | 76 | [43] | |
| β-secretase | BACE1 GeneID: 23621 Locus: 11q23.2-q23.3 Protein length: 501 | bace1 zgc:77409 GeneID: 403005 Chromosome: 15 Protein length: 505 | 82 | [44] | |
| BACE2 GeneID: 25825 Locus: 21q22.2-q22.3 Protein length: 518 | bace2 zgc:103530 GeneID: 449818 chromosome: 15 Protein length: 462 | [45] | |||
| γ-secretase | PSENEN GeneID: 55851 Locus: 19q13.12 Protein length: 101 | psenen GeneID: 402810 chromosome: 15 Protein length: 101 | 91 | [46,47] | |
| NCSTN Gene ID: 23385 Locus: 1q23.2 Protein length: 709 | ncstn GeneID: 494449 chromosome: 2 Protein length: 707 | 56 | [48] | ||
| APH1b Gene ID: 83464 Locus: 15q22.2 Protein length: 257 | aph1p Gene ID: 386808 chromosome: 7 Protein length: 258 | [46] | |||
| Apolipoprotein E (ApoE) | APOE GeneID:348 Locus: 19q13.32 Protein length: 317 | apoea Gene ID: 553587 chromosome: 19 Protein length: 269 | 27.5 | [49] | |
| apoeb Gene ID: 30314 chromosome: 16 Protein length: 281 | [50] | ||||
| Sortilin related receptor 1 (Sorl1) | SORL1 GeneID: 6653, Locus:11q24.1 Protein length: 2214 | sorl1 Gene ID: 497306, chromosome: 15 Protein length: 2213 | 64 | [51] | |
| Parkinson’s Disease | DJ-1 | DJ-1 Gene ID: 11315 Locus: PARK7 1p36.23 Protein length: 189 | dj-1 Gene ID: 449674 Chromosome: 11 Protein length: 189 | 83 | [52] |
| Parkin | PRKN Gene ID: 5071 Locus: PARK2 6q25.2-q27 Protein length: 465 | prkn Gene ID: 550328 Chromosome: 13 Protein length: 458 | 62 | [53] | |
| PTEN-induced kinase 1 (PINK-1) | PINK1 Gene ID: 65018 Locus: PARK6 1p36 Protein length: 581 | pink1 Gene ID: 494085 Chromosome: 6 Protein length: 574 | 54 | [54] | |
| Leucine-rich repeat kinase2 (LRRK2) | LRRK2 GeneID: 120892 Locus: PARK8 12q12 Protein length: 2527 | lrrk2 GeneID: 559366 Chromosome: 25 Protein length: 1985 | 38 | [55] | |
| Huntington’s Disease | Huntingtin | HTT GeneID: 3064 Locus: 4q16.3 Protein length: 3144 | htt GeneID: 30214 Chromosome: 1 Protein length: 3121 | 70 | [56] |
| Amyotrophic Lateral Sclerosis (ALS) | Fused in sarcoma | FUS GeneID: 2521 Locus: 16p11.2 Protein length: 526 | fus Gene ID: 394058 Chromosome: 3 Protein length: 541 | 63 | [57] |
| Tar DNA binding protein of 43 (TDP-43) | TARDBP GeneID: 23435 Locus: 1p36.22 Protein length: 414 | tardpb GeneID: 325052 Chromosome: 6 Protein length: 412 | 71 | [58] | |
| Spinocerebellar Ataxia Type 1 | Ataxin 1 | ATXN1 GeneID: 6310 Locus: 6p23 Protein length: 815 | atxn1a GeneID: 565841 Chromosome: 16 Protein length: 827 | 32 | [59] |
| atxn1b GeneID: 557340 Chromosome: 19 Protein length: 781 | 42 | ||||
| Schizophrenia | Dystrobrevin binding protein | DISC1 Gene ID: 27185 Locus: 1q42.2 Protein length:854 | disc1 GeneID: 407621 Chromosome: 13 Protein length: 994 | 53 | [60] |
| Kinesin family member 17 | KIF17 Gene ID: 57576 Locus: 1p36.12 Protein length:1029 | kif17 GeneID:557863 Chromosome: 11 Protein length: 823 | 83 | [61] |
Table 2.
Zebrafish bioassay-guided isolation and structurally characterized natural products.
| Source | Disease/Targets | Molecules | References |
|---|---|---|---|
| Pharbitis nil (Seeds) | anti-seizure | Pharbitin | [100] |
| Rehmannia glutinosa (Root) | angiogenesis effect | Norviburtinal | [101] |
| Rhynchosia viscosa (Whole Plant) | angiogenesis effect | Rhynchoviscin | [98] |
| Dysosma versipellis | anti-angiogenesis | Kaempferol | [102] |
| Ligusticum sinense (Rhizoma) | anti-melanogenesis | 1. Lignan 2. cis-4-pentylcyclohex-3-ene-1,2-diol | [103] |
| Tephrosia purpurea (Leaves) | anti-acetylcholinesterase | trans-Tephrostachin | [96] |
| Solanum torvum (leaves) | anti-convulsant | Paniculonin A Paniculonin B | [95] |
| Skeletonema marinoi | anti-seizure | Inosine | [104] |
Table 3.
Transgenic zebrafish models for neurodegenerative diseases.
| Disease | Gene | Technique | References |
|---|---|---|---|
| Alzheimer’s Disease | appb | Morpholino injection - knockdown | [117] |
| psen1 | Morpholino injections - knockdown | [121] | |
| bace2 | Zinc-finger nucleases (ZFNs)—knockout | [45] | |
| psenen | morpholino injection—knockdown | [47] | |
| apoea | CRISPR-Cas—knockout | [119] | |
| apoe | morpholino injection (live cell imaging) | [122] | |
| Parkinson’s Disease | djJ-1 | morpholino injection—knockdown | [123] |
| prkn | gripNAsTM-mediated knockdown | [124] | |
| pink1 | transcription activator-like effector nucleases (TALENs)—knockdown | [125] | |
| lrrk2 | morpholino injection—knockdown | [118] | |
| Amyotrophic Lateral Sclerosis | tardbp (bpt1) | CRISPR-Cas—knockin | [40] |
| fus | CRISPR-Cas—knockin | [120] | |
| Huntington’s Disease | htt | morpholino injection—knockdown | [71] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Pitchai, A.; Rajaretinam, R.K.; Freeman, J.L. Zebrafish as an Emerging Model for Bioassay-Guided Natural Product Drug Discovery for Neurological Disorders. Medicines 2019, 6, 61. https://0-doi-org.brum.beds.ac.uk/10.3390/medicines6020061
AMA Style
Pitchai A, Rajaretinam RK, Freeman JL. Zebrafish as an Emerging Model for Bioassay-Guided Natural Product Drug Discovery for Neurological Disorders. Medicines. 2019; 6(2):61. https://0-doi-org.brum.beds.ac.uk/10.3390/medicines6020061
Chicago/Turabian StylePitchai, Arjun, Rajesh Kannan Rajaretinam, and Jennifer L. Freeman. 2019. "Zebrafish as an Emerging Model for Bioassay-Guided Natural Product Drug Discovery for Neurological Disorders" Medicines 6, no. 2: 61. https://0-doi-org.brum.beds.ac.uk/10.3390/medicines6020061
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.