Targeting Aberrant Sialylation to Treat Cancer



Institute of Genetic Medicine, Newcastle University, Newcastle upon Tyne NE1 3BZ, UK
*
Author to whom correspondence should be addressed.
Medicines 2019, 6(4), 102; https://0-doi-org.brum.beds.ac.uk/10.3390/medicines6040102
Submission received: 20 September 2019
/
Accepted: 10 October 2019
/
Published: 13 October 2019
(This article belongs to the Special Issue Application of Glycobiology in the Treatment of Diseases)
Abstract
:Cell surface carbohydrates (known as glycans) are often aberrantly expressed or found at atypical levels in cancer. Glycans can impact all steps in tumour progression, from malignant transformation to metastasis, and have roles in all the cancer hallmarks. An increased understanding of glycans in the metastatic cascade offers exciting new therapeutic opportunities. Glycan-based targeting strategies are currently being tested in clinical trials and are a rich and untapped frontier for development. As we learn more about cancer glycobiology, new targets will continue to emerge for drug design. One key change in tumour glycosylation is the upregulation of cancer-associated sialylated glycans. Abnormal sialylation is integral to tumour growth, metastasis and immune evasion; therefore, targeting sialic acid moieties in cancer could be of high therapeutic value. Here, we summarise the changes to sialic acid biology in cancer and discuss recent advances and technologies bringing sialic-acid targeting treatments to the forefront of cancer therapeutics.
1. Introduction
All cells are coated with carbohydrates, known as glycans, which form a layer on the cell surface known as the glycocalyx [1]. Glycans are present in many different forms (including glycoproteins, proteoglycans and glycolipids) and build the basis for a universal language (the glycome) that is used for communication between cells [2]. Aberrant glycosylation is a universal feature of cancer cells, and it is well established that even small changes to the glycome can severely affect tumour cell biology [3,4]. Common cancer-associated glycome changes include aberrant sialylation, fucosylation, truncated O-glycans, and changes to N- and O-glycan branching [5]. Aberrant glycosylation is linked to all of the cancer hallmarks, and glycans can impact all steps in tumour progression, from malignant transformation to metastasis [4,6]. This makes cancer-associated glycans attractive therapeutic targets, and new technologies to study glycans have spurred a renewed interest in this area [7].
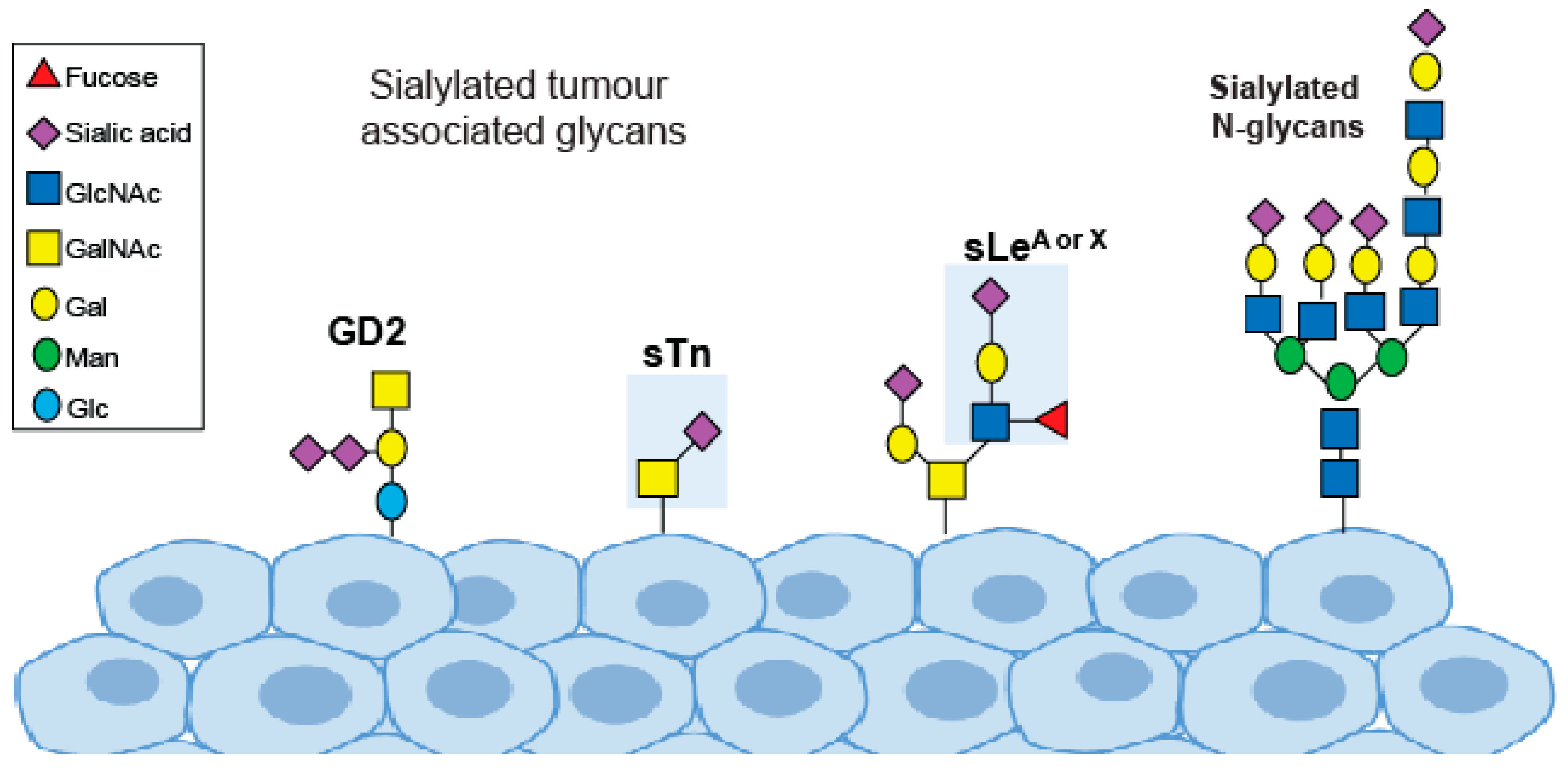
One key change in tumour glycosylation is the upregulation of cancer-associated sialylated glycans, known as tumour sialoglycans [8]. Sialic acids are negatively charged sugar residues that often terminate the glycans of glycoproteins and glycolipids. The cell surface of cancer cells is covered with a dense layer of sialoglycans including SLeA, SLeX, STn and GM2 [9,10] (Figure 1). Aberrant sialylation plays a fundamental role in cancer cell growth, metastasis and immune evasion, and targeting sialic acid in cancer is an attractive therapeutic option. Here, we summarise the changes to sialic acid glycans in cancer and highlight recent advances in sialic acid therapeutics.
2. Tumour Growth and Metastasis
Aberrant sialylation contributes to tumour growth and metastasis at multiple levels and has been described as a key player in cancer progression [11,12,13]. Sialic acids can promote cancer progression by driving tumour growth, protecting cells from apoptosis, facilitating cancer cell detachment, enhancing invasion, promoting immune evasion, and enabling extravasation from the bloodstream to form metastases [8,10,14,15,16,17]. Sialylation of specific proteins has been linked to numerous signalling molecules and pathways important in cancer. For example, an increase in α2-3 sialylation in gastric carcinoma can lead to activation of the receptor tyrosine kinases MET and RON to promote a more invasive phenotype [18,19]. The terminal sialylation of N-glycans can also confer tumour cell resistance to hypoxia and have a major impact on malignant cell phenotypes [20,21]. N-glycans containing terminal α2-6 sialic acid contribute to the aberrant regulation of E-cadherin in cancer to impair cancer cell adhesion and promote cell invasion and metastasis [22,23,24].
New tools to target sialic acids are being developed and have shown promise in preventing/inhibiting cancer metastasis. The sialic acid glycomimetic (P-3Fax-Neu5Ac) delays tumour growth in vivo and can be delivered to cancer cells using nanoparticles to prevent the metastatic spread of melanoma cells to the lung in a murine model [25,26]. Intra-tumoral injections with Ac53FaxNeu5Ac can block sialic acid expression and suppress tumour growth in multiple in vivo tumour models [10]. These findings demonstrate the important role of sialic acids in tumour growth and metastasis and indicate that a sialic acid blockade could be of high therapeutic value.
3. Glycosyltransferase and Glycosidase Enzymes
The tumour glyco-code is generated through the cooperative action of multiple glycosylation enzymes that catalyse the addition or removal of specific glycans onto proteins and lipids. These changes are often driven by the altered expression of genes involved in glycan synthesis [27]. Abnormal sialylation in cancer cells has been linked to the altered expression of both sialyltransferase and sialidase enzymes [28]. Sialyltransferases are enzymes that transfer sialic acid residues to the terminal position of glycoconjugates. The sialyltransferases are a family of 20 enzymes that catalyse the attachment of sialic acid to the underlying glycan chain via different glycosidic linkages (α2-3, α2-6, or α2-8). Sialyltransferases are expressed in a tissue-specific manner and can be further divided into four sub-families (ST3Gal, ST6Gal. ST6GalNAc, and ST8SIA) [29]. Sialyltransferases are often misregulated in cancer and this is linked to the expression of cancer-associated antigens that contain sialic acid [5,29,30].
Key examples of sialyltransferases important in cancer include ST6GAL1; ST3GAL4 ST3GAL6 and ST6GalNAc1/2 (summarised in Table 1). ST6GAL1 is upregulated in numerous cancer types and has recently been linked to all of the cancer hallmarks [11,31,32,33]. Therapeutically, ST6GAL1 can be used to predict response to EGFR/HER2 inhibitors in ovarian cancer cells [34]. Similarly, ST3GAL4 is also upregulated in cancer, and is associated with poor prognosis; metastasis and the synthesis of Sialyl Lewix X (sLeX) in gastric carcinoma [18,35]. ST3GAL6 is also linked to the synthesis of sLeX and the generation of E-selectin ligands [36,37]. Recently, ST3GAL6 was found to be critical to bone marrow homing and resistance to therapy in multiple myeloma [36], and a follow-up study demonstrated this can be inhibited with the E-selectin antagonist GMI-1271 [37]. Other cancer-associated sialyltrasferases include ST6GalNAc1 (which catalyses the cancer-associated sTn antigen) and is associated with metastasis [8,27,38,39,40,41,42,43], and ST6GALNAC2, which has been identified as a metastasis suppressor in breast cancer patients and could be used to stratify patients for treatment with galectin-3 inhibitors [44,45].
The removal of sialic acids from glycoconjugates is catalysed by sialidase enzymes. Sialidases can modulate the binding sites of functional molecules and are important in various biological processes [46]. There are four sialidase enzymes, NEU1, NEU2, NEU3 and NEU4 and each of these can be altered in cancer [47,48,49,50,51]. Sialidase enzymes are emerging as novel therapeutic targets in cancer. Of particular interest, NEU1 can be targeted with oseltavimir phosphate in pancreatic, breast and ovarian tumours, and this may improve the drug sensitivity of chemoresistant cells [48].
4. Cancer-Associated Sialyloglycans
Changes to sialylated glycans in cancer include an upregulation of the Sialyl Lewis antigens (sLeA and sLeX), an increase in the truncated O-glycan sialyl-Tn (STn) and an increase in the sialylated ganglioside GM2 [52] (Figure 1). The sialyl Lewis antigens are part of the Lewis family of blood group antigens, named after the discoverer of a series of antigens found on red blood cells, and are the minimal recognition motif for ligands of selectins (a family of lectins with roles in leukocyte trafficking and cancer metastasis). Tumour cells coated with sLeA and sLex are recognised as migrating leucocytes, enabling them to escape the bloodstream and colonise other organs and tissues [53]. Many solid tumours and adenocarcinomas express high levels of sLeA and sLeX, and thus targeting selectins and sLeA/X is attractive therapeutically [53]. Potential strategies include the use of glycomimetic drugs, such as the selectin inhibitors Uproleselan (GMI-1271) and Rivipansel (GMI-1070), which have been tested in clinical trials [52,54] (Table 2). Uproleselan (GMI-1271) is in phase 3 clinical trials, in combination with chemotherapy, to treat relapsed acute myeloid leukemia (NCT03616470), and has also shown promise in pre-clinical models of breast cancer, where it can prevent bone metastasis and improve survival [55]. Rivipansel (GMI-1070) reached phase 3 clinical trials for sickle cell disease but reported negative results (NCT02187003).
One of the best characterised cancer-associated glycans is the truncated O-glycan STn, which is upregulated in virtually all epithelial cancers and linked to metastasis and poor patient outcome [8]. STn is carried by a variety of glycoproteins and glycolipids and has important role in tumour development and invasiveness [8]. The STn antigen has been investigated widely as a circulating biomarker for numerous cancer types [17], and a vaccine against STn has been tested in clinical trials and can increase survival in a subset of breast cancer patients receiving hormonal therapy [56,57,58].
Gangliosides (sialic-acid-containing glycosphingolipids) are important regulators of cell signalling in cancer [59]. The complex ganglioside GD2 is expressed on tumours of neuroectodermal origin and has a key role in the aggressiveness of some cancers including neuroblastoma and melanoma. GD2 can be inhibited with monoclonal antibodies and holds major potential as a target for cancer therapy [52,60]. The anti-GD2 monoclonal antibody dinutuximab can help improve survival in patients with high-risk neuroblastoma. Dinutuximab is currently in phase 3 clinical trials for neuroblastoma, and may be relevant to other cancer types [61].
5. Siglecs and Cancer Immunotherapy
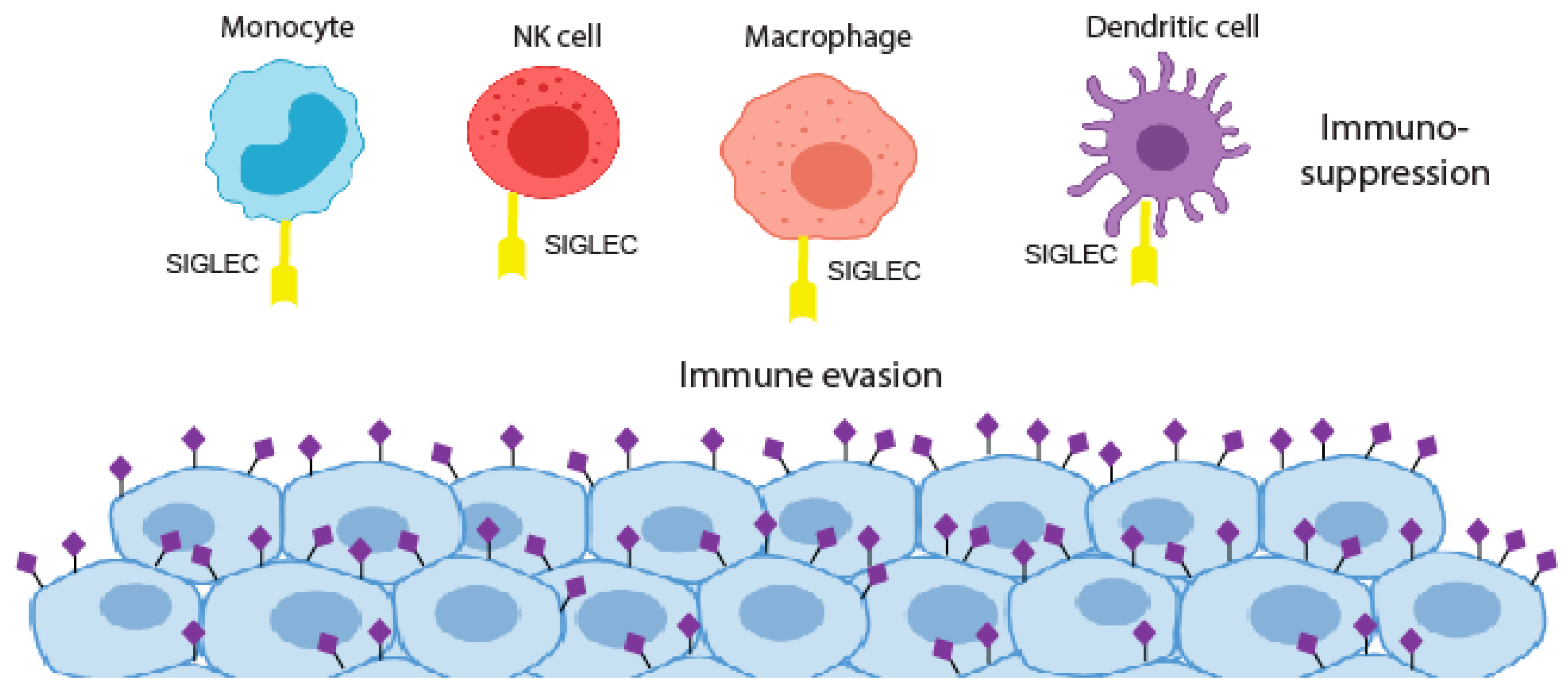
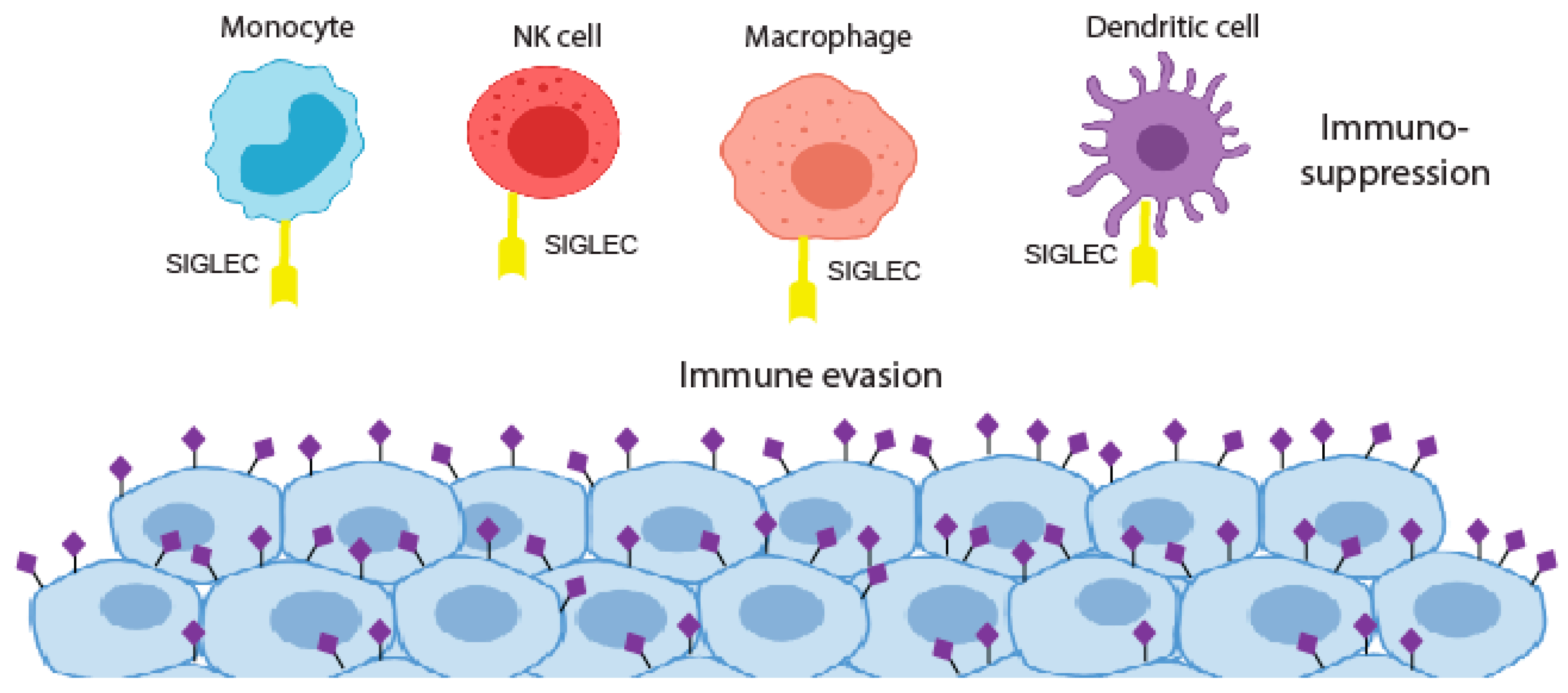
The aberrant glycosylation of tumour cells can lead to new connections with immune cells resulting in an immunosuppressive phenotype. This can be through the induction of self-glycan structures to limit immune self-reactive responses or by the expression of glycans that can reduce the function of effector T cells [71]. The dense layer of sialic acids on the surface of cancer cells has long been implicated in protecting tumours from eradication by the immune system. In the 1960s and 1970s, immunotherapy clinical trials using irradiated cancer cells treated with bacterial sialidase were carried out but were not taken forward [72,73,74]. Recent developments in glyco-tools have driven a renewed interest in the role of sialic acids in the formation of an immuno-suppressive environment, with sialic acids on the surface of cancer cells believed to play a crucial role in immune modulation and tumour immune evasion [10]. Sialylated glycans found on both glycoproteins and glycolipids are recognised by Siglecs, a family of lectins that are expressed on the surface of many immune cell subtypes in the tumour microenvironment [75,76,77,78,79]. The interaction of cancer cell bound sialic acid with Siglecs can thus modulate immune cell phenotype and allow tumours to escape the immune system [64] (Figure 2). Cancer-associated glycans, such as STn and sialyl T, are well characterised examples of this, being linked to the impaired maturation and activation of macrophages and dendritic cells, and are also implicated in the deactivation of natural killer (NK) cells and the formation of regulatory T cells [63,78,80,81,82]. Hence, determining the specific glycan signature of cancer cells (known as the ‘glyco-code’) will be crucial to understand how glyans promote immune evasion [71,83].
Targeting sialic acid moieties has become an upcoming strategy for cancer immunotherapy [10,52]. Studies have shown that a sialic acid blockade has a profound effect on the tumour immune microenvironment, with pro-inflammatory effects, including increased numbers and activation state of CD8+ T cells, reduced percentages of myeloid and regulatory T (Tregs) cells, and increased tumour cell killing by cytotoxic t cells [10]. Bull et al. (2018) also demonstrated that a sialic acid blockade induces a protection upon rechallenge, suggestive of a curative immune response and conversion to a more immune-permissive microenvironment [10]. How sialic acid inhibition promotes these changes in tumour microenvironment is poorly understood; however, emerging studies suggest loss of tumour sialic acid can block the action of immune modulatory Siglecs on immune cells. To date, Siglecs-9, -7, -10 and -15 and their ligands have shown promise as targets to dampen anti-tumor immunity [62,63,64,65,66,67,68,69]. The interaction of Siglec-9 with sialylated MUC1 can induce the differentiation of monocytes to tumour-associated macrophages and increase levels of the checkpoint ligand PD-L1 [78,79]. Siglec-9 is upregulated on tumour-infiltrating T cells from patients with non-small lung cancer (NSCLC), colorectal and ovarian cancer and targeting of the sialoglycan-Siglec-9 pathway could be used to enhance T cell activation [63]. A high-profile 2019 study found that sialoglycoprotein CD24 acts an anti-phagocytic ‘don’t eat me’ signal that can protect cancer cells from attack by Siglec-10-expressing macrophages. Blockade of CD24-Siglec10 enhances clearance of CD24+ tumours and is a potrntial immunotherapy target [62]. Siglec-15 is upregulated in cancer cells and tumour-infiltrating myeloid cells, and is a critical immune suppressor. Siglec-15 suppresses antigen-specific T cell responses at the tumour site. An anti-Siglec-15 antibody has been developed and can reverse T cell suppression to promote tumour immunity and is potentially useful for cancer patients who are resistant to current therapies [69].
Precision glycocalyx editing with antibody-sialidase conjugates has also been reported and is a promising avenue for cancer immune therapy. Here, an antibody directs sialidase to selectively remove sialic acid from tumour cells and enable immune cells to kill the desialylated cancer cells [84]. The EAGLE platform (that delivers a targeted sialidase enzyme to the tumours) has shown great promise in pre-clinical models and is about to enter clinical trials for breast cancer [70]. Other potential strategies to target sialic acid glycans in cancer immunotherapies include anti-glycan vaccines, blocking cancer-associated glycan lectin interactions, and dendritic cell targeting [71].
6. Conclusions and Future Perspectives
In the era of personalised medicine, there is a huge potential to develop new glycan-based therapies to treat cancer. A key change in cancer glycosylation is an upregulation in the levels of sialylation, as well as the expression of cancer-associated sialoglycans. Aberrant sialylation is integral to tumour growth, survival, metastasis and immune evasion, and targeting abnormal sialylation will likely be of high therapeutic value. New technologies to study glycosylation are bringing glycan targeting strategies to the forefront of cancer therapeutics, particularly in the area of cancer immunology. Moving forward, drugs targeting tumour glycans will likely be used synergistically with existing chemotherapy and/or radiotherapy approaches to impact disease outcomes. As well as themselves being drug targets, changes to glycans can likely also be exploited to predict sensitivity and resistance to other treatment strategies and, ultimately, improve clinical outcome.
Author Contributions
J.M. and E.S. jointly wrote and edited the manuscript.
Funding
This work was supported by Prostate Cancer UK through a Research Innovation Award (RIA16-ST2-011).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Tarbell, J.M.; Cancel, L.M. The glycocalyx and its significance in human medicine. J. Int. Med. 2016, 280, 97–113. [Google Scholar] [CrossRef] [Green Version]
- Varki, A. Biological roles of glycans. Glycobiology 2017, 27, 3–49. [Google Scholar] [CrossRef] [PubMed]
- Pinho, S.S.; Reis, C.A. Glycosylation in cancer: Mechanisms and clinical implications. Nat. Rev. Cancer 2015, 15, 540–555. [Google Scholar] [CrossRef] [PubMed]
- Munkley, J.; Elliott, D.J. Hallmarks of glycosylation in cancer. Oncotarget 2016, 7, 35478–35489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munkley, J.; Mills, I.G.; Elliott, D.J. The role of glycans in the development and progression of prostate cancer. Nat. Rev. Urol. 2016, 13, 324–333. [Google Scholar] [CrossRef] [PubMed]
- Vajaria, B.N.; Patel, P.S. Glycosylation: A hallmark of cancer? Glycoconj. J. 2017, 34, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Hudak, J.E.; Bertozzi, C.R. Glycotherapy: New advances inspire a reemergence of glycans in medicine. Chem. Biol. 2014, 21, 16–37. [Google Scholar] [CrossRef]
- Munkley, J. The Role of Sialyl-Tn in Cancer. Int. J. Mol. Sci. 2016, 17, 275. [Google Scholar] [CrossRef]
- Munkley, J. The glycosylation landscape of pancreatic cancer. Oncol. Lett. 2019, 17, 2569–2575. [Google Scholar] [CrossRef]
- Bull, C.; Boltje, T.J.; Balneger, N.; Weischer, S.M.; Wassink, M.; van Gemst, J.J.; den Brok, M.H. Sialic Acid Blockade Suppresses Tumor Growth by Enhancing T-cell-Mediated Tumor Immunity. Cancer Res. 2018, 78, 3574–3588. [Google Scholar] [CrossRef]
- Garnham, R.; Scott, E.; Livermore, K.; Munkley, J. ST6GAL1: A key player in cancer (Review). Oncol. Lett. 2019, 18, 983–989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schultz, M.J.; Swindall, A.F.; Bellis, S.L. Regulation of the metastatic cell phenotype by sialylated glycans. Cancer Metastas. Rev. 2012, 31, 501–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bull, C.; Stoel, M.A.; den Brok, M.H.; Adema, G.J. Sialic acids sweeten a tumor’s life. Cancer Res. 2014, 74, 3199–3204. [Google Scholar] [CrossRef] [PubMed]
- Hauselmann, I.; Borsig, L. Altered tumor-cell glycosylation promotes metastasis. Front. Oncol. 2014, 4, 28. [Google Scholar] [CrossRef] [PubMed]
- Amano, M.; Eriksson, H.; Manning, J.C.; Detjen, K.M.; Andre, S.; Nishimura, S.; Gabous, H.J. Tumour suppressor p16(INK4a)-anoikis-favouring decrease in N/O-glycan/cell surface sialylation by down-regulation of enzymes in sialic acid biosynthesis in tandem in a pancreatic carcinoma model. FEBS J. 2012, 279, 4062–4080. [Google Scholar] [CrossRef]
- Bos, P.D.; Zhang, X.H.; Nadal, C.; Shu, W.; Gomis, R.R.; Nguyen, D.X.; Massagué, J. Genes that mediate breast cancer metastasis to the brain. Nature 2009, 459, 1005–1009. [Google Scholar] [CrossRef] [PubMed]
- Scott, E.; Munkley, J. Glycans as Biomarkers in Prostate Cancer. Int. J. Mol. Sci. 2019, 20, 1389. [Google Scholar] [CrossRef]
- Gomes, C.; Osorio, H.; Pinto, M.T.; Campos, D.; Oliveira, M.J.; Reis, C.A. Expression of ST3GAL4 leads to SLe(x) expression and induces c-Met activation and an invasive phenotype in gastric carcinoma cells. PLoS ONE 2013, 8, e66737. [Google Scholar] [CrossRef]
- Mereiter, S.; Magalhaes, A.; Adamczyk, B.; Jin, C.; Almeida, A.; Drici, L.; Santos, L.L. Glycomic analysis of gastric carcinoma cells discloses glycans as modulators of RON receptor tyrosine kinase activation in cancer. Biochim. Biophys. Acta 2016, 1860, 1795–1808. [Google Scholar] [CrossRef]
- Jones, R.B.; Dorsett, K.A.; Hjelmeland, A.B.; Bellis, S.L. The ST6Gal-I sialyltransferase protects tumor cells against hypoxia by enhancing HIF-1alpha signaling. J. Biol. Chem. 2018, 293, 5659–5667. [Google Scholar] [CrossRef]
- Isaji, T.; Im, S.; Kameyama, A.; Wang, Y.Q.; Fukuda, T.; Gu, J.G. A complex between phosphatidylinositol 4-kinase II and integrin 31 is required for N-glycan sialylation in cancer cells. J. Biol. Chem. 2019, 294, 4425–4436. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.B.; Lee, I.; Kamar, M.; Pierce, M. N-acetylglucosaminyltransferase V expression levels regulate cadherin-associated homotypic cell-cell adhesion and intracellular signaling pathways. J. Biol. Chem. 2003, 278, 52412–52424. [Google Scholar] [CrossRef] [PubMed]
- Ihara, S.; Miyoshi, E.; Ko, J.H.; Murata, K.; Nakahara, S.; Honke, K.; Taniguchi, N. Prometastatic effect of N-acetylglucosaminyltransferase V is due to modification and stabilization of active matriptase by adding beta 1-6 GlcNAc branching. J. Biol. Chem. 2002, 277, 16960–16967. [Google Scholar] [CrossRef] [PubMed]
- Pinho, S.S.; Figueiredo, J.; Cabral, J.; Carvalho, S.; Dourado, J.; Magalhaes, A.; Gärtner, F.; Mendonfa, A.M.; Isaji, T.; Gu, J.; et al. E-cadherin and adherens-junctions stability in gastric carcinoma: Functional implications of glycosyltransferases involving N-glycan branching biosynthesis, N-acetylglucosaminyltransferases III and V. Biochim. Biophys. Acta 2013, 1830, 2690–2700. [Google Scholar] [CrossRef]
- Bull, C.; Boltje, T.J.; van Dinther, E.A.; Peters, T.; de Graaf, A.M.; Leusen, J.H.; Adema, G.J. Targeted delivery of a sialic acid-blocking glycomimetic to cancer cells inhibits metastatic spread. ACS Nano 2015, 9, 733–745. [Google Scholar] [CrossRef] [PubMed]
- Bull, C.; Boltje, T.J.; Wassink, M.; de Graaf, A.M.; van Delft, F.L.; den Brok, M.H.; Adema, G.J. Targeting aberrant sialylation in cancer cells using a fluorinated sialic acid analog impairs adhesion, migration, and in vivo tumor growth. Mol. Cancer Ther. 2013, 12, 1935–1946. [Google Scholar] [CrossRef]
- Munkley, J.; Vodak, D.; Livermore, K.E.; James, K.; Wilson, B.T.; Knight, B.; Leung, H.Y. Glycosylation is an Androgen-Regulated Process Essential for Prostate Cancer Cell Viability. EBioMedicine 2016, 8, 103–116. [Google Scholar] [CrossRef] [Green Version]
- Vajaria, B.N.; Patel, K.R.; Begum, R.; Patel, P.S. Sialylation: An Avenue to Target Cancer Cells. Pathol. Oncol. Res. 2016, 22, 443–447. [Google Scholar] [CrossRef]
- Dall’Olio, F.; Malagolini, N.; Trinchera, M.; Chiricolo, M. Sialosignaling: Sialyltransferases as engines of self-fueling loops in cancer progression. Biochim. Biophys. Acta 2014, 1840, 2752–2764. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, A.S.; Harduin-Lepers, A.; Magalhaes, A.; Machado, E.; Mendes, N.; Costa, L.T.; Reis, C.A. Differential expression of alpha-2,3-sialyltransferases and alpha-1,3/4-fucosyltransferases regulates the levels of sialyl Lewis a and sialyl Lewis x in gastrointestinal carcinoma cells. Int. J. Biochem. Cell Biol. 2010, 42, 80–89. [Google Scholar] [CrossRef]
- Hsieh, C.C.; Shyr, Y.M.; Liao, W.Y.; Chen, T.H.; Wang, S.E.; Lu, P.C.; Hsiao, M. Elevation of β-galactoside alpha2,6-sialyltransferase 1 in a fructoseresponsive manner promotes pancreatic cancer metastasis. Oncotarget 2017, 8, 7691–7709. [Google Scholar] [CrossRef] [PubMed]
- Wei, A.; Fan, B.; Zhao, Y.; Zhang, H.; Wang, L.; Yu, X.; Wang, S. ST6Gal-I overexpression facilitates prostate cancer progression via the PI3K/Akt/GSK-3beta/beta-catenin signaling pathway. Oncotarget 2016, 7, 65374–65388. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Isaji, T.; Im, S.; Fukuda, T.; Hashii, N.; Takakura, D.; Kawasaki, N.; Gu, J. beta-Galactoside alpha2,6-sialyltranferase 1 promotes transforming growth factor-beta-mediated epithelial-mesenchymal transition. J. Biol. Chem. 2014, 289, 34627–34641. [Google Scholar] [CrossRef] [PubMed]
- Britain, C.M.; Holdbrooks, A.T.; Anderson, J.C.; Willey, C.D.; Bellis, S.L. Sialylation of EGFR by the ST6Gal-I sialyltransferase promotes EGFR activation and resistance to gefitinib-mediated cell death. J. Ovarian Res. 2018, 11, 12. [Google Scholar] [CrossRef] [PubMed]
- Mereiter, S.; Magalhaes, A.; Adamczyk, B.; Jin, C.; Almeida, A.; Drici, L.; Reis, C.A. Glycomic and sialoproteomic data of gastric carcinoma cells overexpressing ST3GAL4. Data Brief 2016, 7, 814–833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glavey, S.V.; Manier, S.; Natoni, A.; Sacco, A.; Moschetta, M.; Reagan, M.R.; Zhang, Y. The sialyltransferase ST3GAL6 influences homing and survival in multiple myeloma. Blood 2014, 124, 1765–1776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Natoni, A.; Smith, T.A.G.; Keane, N.; McEllistrim, C.; Connolly, C.; Jha, A.; Kirkham-McCarthy, L. E-selectin ligands recognised by HECA452 induce drug resistance in myeloma, which is overcome by the E-selectin antagonist, GMI-1271. Leukemia 2017, 31, 2642–2651. [Google Scholar] [CrossRef]
- Munkley, J.; Oltean, S.; Vodak, D.; Wilson, B.T.; Livermore, K.E.; Zhou, Y.; McCullagh, P. The androgen receptor controls expression of the cancer-associated sTn antigen and cell adhesion through induction of ST6GalNAc1 in prostate cancer. Oncotarget 2015, 6, 34358–34374. [Google Scholar] [CrossRef]
- Munkley, J.; Elliott, D.J. Sugars and cell adhesion: The role of ST6GalNAc1 in prostate cancer progression. Cancer Cell Microenviron. 2016, 3, e1174. [Google Scholar]
- Genega, E.M.; Hutchinson, B.; Reuter, V.E.; Gaudin, P.B. Immunophenotype of high-grade prostatic adenocarcinoma and urothelial carcinoma. Modern Pathol. 2000, 13, 1186–1191. [Google Scholar] [CrossRef]
- Julien, S.; Adriaenssens, E.; Ottenberg, K.; Furlan, A.; Courtand, G.; Vercoutter-Edouart, A.S.; Le Bourhis, X. ST6GalNAc I expression in MDA-MB-231 breast cancer cells greatly modifies their O-glycosylation pattern and enhances their tumourigenicity. Glycobiology 2006, 16, 54–64. [Google Scholar] [CrossRef] [PubMed]
- Munkley, J. Glycosylation is a global target for androgen control in prostate cancer cells. Endocr. Relat. Cancer 2017, 24, R49–R64. [Google Scholar] [CrossRef] [PubMed]
- Tamura, F.; Sato, Y.; Hirakawa, M.; Yoshida, M.; Ono, M.; Osuga, T.; Kawano, Y. RNAi-mediated gene silencing of ST6GalNAc I suppresses the metastatic potential in gastric cancer cells. Gastric Cancer 2014, 19, 85–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrer, C.M.; Reginato, M.J. Sticking to sugars at the metastatic site: Sialyltransferase ST6GalNAc2 acts as a breast cancer metastasis suppressor. Cancer Discov. 2014, 4, 275–277. [Google Scholar] [CrossRef] [PubMed]
- Murugaesu, N.; Iravani, M.; van Weverwijk, A.; Ivetic, A.; Johnson, D.A.; Antonopoulos, A.; Mitsopoulos, C. An in vivo functional screen identifies ST6GalNAc2 sialyltransferase as a breast cancer metastasis suppressor. Cancer Discov. 2014, 4, 304–317. [Google Scholar] [CrossRef] [PubMed]
- Miyagi, T.; Yamaguchi, K. Mammalian sialidases: Physiological and pathological roles in cellular functions. Glycobiology 2012, 22, 880–896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forcella, M.; Mozzi, A.; Stefanini, F.M.; Riva, A.; Epistolio, S.; Molinari, F.; Frattini, M. Deregulation of sialidases in human normal and tumor tissues. Cancer Biomark. 2018, 21, 591–601. [Google Scholar] [CrossRef]
- Haxho, F.; Neufeld, R.J.; Szewczuk, M.R. Neuraminidase-1: A novel therapeutic target in multistage tumorigenesis. Oncotarget 2016, 7, 40860–40881. [Google Scholar] [CrossRef] [Green Version]
- Uemura, T.; Shiozaki, K.; Yamaguchi, K.; Miyazaki, S.; Satomi, S.; Kato, K.; Miyagi, T. Contribution of sialidase NEU1 to suppression of metastasis of human colon cancer cells through desialylation of integrin beta4. Oncogene 2009, 28, 1218–1229. [Google Scholar] [CrossRef]
- Glanz, V.Y.; Myasoedova, V.A.; Grechko, A.V.; Orekhov, A.N. Sialidase activity in human pathologies. Eur. J. Pharmacol. 2019, 842, 345–350. [Google Scholar] [CrossRef]
- Nath, S.; Mandal, C.; Chatterjee, U.; Mandal, C. Association of cytosolic sialidase Neu2 with plasma membrane enhances Fas-mediated apoptosis by impairing PI3K-Akt/mTOR-mediated pathway in pancreatic cancer cells. Cell Death Dis. 2018, 9, 210. [Google Scholar] [CrossRef] [PubMed]
- Mereiter, S.; Balmana, M.; Campos, D.; Gomes, J.; Reis, C.A. Glycosylation in the Era of Cancer-Targeted Therapy: Where Are We Heading? Cancer Cell 2019, 36, 6–16. [Google Scholar] [CrossRef] [PubMed]
- Natoni, A.; Macauley, M.S.; O’Dwyer, M.E. Targeting Selectins and Their Ligands in Cancer. Front. Oncol. 2016, 6, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ernst, B.; Magnani, J.L. From carbohydrate leads to glycomimetic drugs. Nat. Rev. Drug Discov. 2009, 8, 661–677. [Google Scholar] [CrossRef] [PubMed]
- Esposito, M.; Mondal, N.; Greco, T.M.; Wei, Y.; Spadazzi, C.; Lin, S.C.; Cristea, I.M. Bone vascular niche E-selectin induces mesenchymal-epithelial transition and Wnt activation in cancer cells to promote bone metastasis. Nat. Cell Biol. 2019, 21, 627–639. [Google Scholar] [CrossRef] [PubMed]
- Holmberg, L.A.; Sandmaier, B.M. Vaccination with Theratope (STn-KLH) as treatment for breast cancer. Expert Rev. Vaccin. 2004, 3, 655–663. [Google Scholar] [CrossRef] [PubMed]
- Mayordomo, J.; Tres, A.; Miles, D.; Finke, L.; Jenkins, H. Long-term follow-up of patients concomitantly treated with hormone therapy in a prospective controlled randomized multicenter clinical study comparing STn-KLH vaccine with KLH control in stage IV breast cancer following first-line chemotherapy. J. Clin. Oncol. 2004, 22, 2603. [Google Scholar] [CrossRef]
- Reddish, M.A.; MacLean, G.D.; Poppema, S.; Berg, A.; Longenecker, B.M. Pre-immunotherapy serum CA27.29 (MUC-1) mucin level and CD69+ lymphocytes correlate with effects of Theratope sialyl-Tn-KLH cancer vaccine in active specific immunotherapy. Cancer Immunol. Immunother. 1996, 42, 303–309. [Google Scholar] [CrossRef]
- Groux-Degroote, S.; Rodriguez-Walker, M.; Dewald, J.H.; Daniotti, J.L.; Delannoy, P. Gangliosides in Cancer Cell Signaling. Prog. Mol. Biol. Trans. Sci. 2018, 156, 197–227. [Google Scholar]
- Cavdarli, S.; Dewald, J.H.; Yamakawa, N.; Guerardel, Y.; Terme, M.; Le Doussal, J.M.; Groux-Degroote, S. Identification of 9-O-acetyl-N-acetylneuraminic acid (Neu5,9Ac2) as main O-acetylated sialic acid species of GD2 in breast cancer cells. Glycoconj. J. 2019, 36, 79–90. [Google Scholar] [CrossRef]
- Ladenstein, R.; Potschger, U.; Valteau-Couanet, D.; Luksch, R.; Castel, V.; Yaniv, I.; Trahair, T. Interleukin 2 with anti-GD2 antibody ch14.18/CHO (dinutuximab beta) in patients with high-risk neuroblastoma (HR-NBL1/SIOPEN): A multicentre, randomised, phase 3 trial. Lancet Oncol. 2018, 19, 1617–1629. [Google Scholar] [CrossRef]
- Barkal, A.A.; Brewer, R.E.; Markovic, M.; Kowarsky, M.; Barkal, S.A.; Zaro, B.W.; Weissman, I.L. CD24 signalling through macrophage Siglec-10 is a target for cancer immunotherapy. Nature 2019, 572, 392–396. [Google Scholar] [CrossRef] [PubMed]
- Stanczak, M.A.; Siddiqui, S.S.; Trefny, M.P.; Thommen, D.S.; Boligan, K.F.; von Gunten, S.; von Bergwelt-Baildon, M. Self-associated molecular patterns mediate cancer immune evasion by engaging Siglecs on T cells. J. Clin. Investig. 2018, 128, 4912–4923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bull, C.; den Brok, M.H.; Adema, G.J. Sweet escape: Sialic acids in tumor immune evasion. Biochim. Biophys. Acta 2014, 1846, 238–246. [Google Scholar] [CrossRef] [PubMed]
- Pearce, O.M.; Laubli, H. Sialic acids in cancer biology and immunity. Glycobiology 2016, 26, 111–128. [Google Scholar] [CrossRef] [PubMed]
- Boligan, K.F.; Mesa, C.; Fernandez, L.E.; von Gunten, S. Cancer intelligence acquired (CIA): Tumor glycosylation and sialylation codes dismantling antitumor defense. Cell. Mol. Life Sci. CMLS 2015, 72, 1231–1248. [Google Scholar] [CrossRef] [PubMed]
- Fraschilla, I.; Pillai, S. Viewing Siglecs through the lens of tumor immunology. Immunol. Rev. 2017, 276, 178–191. [Google Scholar] [CrossRef] [Green Version]
- Adams, O.J.; Stanczak, M.A.; von Gunten, S.; Laubli, H. Targeting sialic acid-Siglec interactions to reverse immune suppression in cancer. Glycobiology 2018, 28, 640–647. [Google Scholar] [CrossRef]
- Wang, J.; Sun, J.; Liu, L.N.; Flies, D.B.; Nie, X.; Toki, M.; Han, X. Siglec-15 as an immune suppressor and potential target for normalization cancer immunotherapy. Nat. Med. 2019, 25, 656–666. [Google Scholar] [CrossRef]
- Peng, L. A novel immunomodulatory strategy of targeting glyco-immune checkpoints with EAGLE technology. Eur. J. Cancer 2018, 103, E77. [Google Scholar]
- RodrIguez, E.; Schetters, S.T.T.; van Kooyk, Y. The tumour glyco-code as a novel immune checkpoint for immunotherapy. Nat. Rev. Immunol. 2018, 18, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Sanford, B.H. An alteration in tumor histocompatibility induced by neuraminidase. Transplantation 1967, 5, 1273–1279. [Google Scholar] [CrossRef] [PubMed]
- Sedlacek, H.H.; Seiler, F.R. Immunotherapy of Neoplastic Diseases with Neuraminidase–Contradictions, New Aspects, and Revised Concepts. Cancer Immunol. Immun. 1978, 5, 153–163. [Google Scholar] [CrossRef]
- Bagshawe, K.D.; Currie, G.A. Immunogenicity of L 1210 murine leukaemia cells after treatment with neuraminidase. Nature 1968, 218, 1254–1255. [Google Scholar] [CrossRef] [PubMed]
- Crocker, P.R.; Paulson, J.C.; Varki, A. Siglecs and their roles in the immune system. Nat. Rev. Immunol. 2007, 7, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Pillai, S.; Netravali, I.A.; Cariappa, A.; Mattoo, H. Siglecs and Immune Regulation. Annu. Rev. Immunol. 2012, 30, 357–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hudak, J.E.; Canham, S.M.; Bertozzi, C.R. Glycocalyx engineering reveals a Siglec-based mechanism for NK cell immunoevasion. Nat. Chem. Biol. 2014, 10, 69. [Google Scholar] [CrossRef]
- Beatson, R.; Tajadura-Ortega, V.; Achkova, D.; Picco, G.; Tsourouktsoglou, T.D.; Klausing, S.; Taylor-Papadimitriou, J. The mucin MUC1 modulates the tumor immunological microenvironment through engagement of the lectin Siglec-9. Nat. Immunol. 2016, 17, 1273–1281. [Google Scholar] [CrossRef] [Green Version]
- Laubli, H.; Pearce, O.M.T.; Schwarz, F.; Siddiqui, S.S.; Deng, L.Q.; Stanczak, M.A.; Schwartz, A.G. Engagement of myelomonocytic Siglecs by tumor-associated ligands modulates the innate immune response to cancer. Proc. Natl. Acad. Sci. USA 2014, 111, 14211–14216. [Google Scholar] [CrossRef] [Green Version]
- Bhatia, R.; Gautam, S.K.; Cannon, A.; Thompson, C.; Hall, B.R.; Aithal, A.; Batra, S.K. Cancer-associated mucins: Role in immune modulation and metastasis. Cancer Metastas. Rev. 2019, 38, 223–236. [Google Scholar] [CrossRef]
- Jandus, C.; Boligan, K.F.; Chijioke, O.; Liu, H.; Dahlhaus, M.; Demoulins, T.; Simon, H.U. Interactions between Siglec-7/9 receptors and ligands influence NK cell-dependent tumor immunosurveillance. J. Clin. Investig. 2014, 124, 1810–1820. [Google Scholar] [CrossRef] [PubMed]
- Cagnoni, A.J.; Perez Saez, J.M.; Rabinovich, G.A.; Marino, K.V. Turning-Off Signaling by Siglecs, Selectins, and Galectins: Chemical Inhibition of Glycan-Dependent Interactions in Cancer. Front. Oncol. 2016, 6, 109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, J.M.; Li, C.W.; Lai, Y.J.; Hung, M.C. Posttranslational Modifications of PD-L1 and Their Applications in Cancer Therapy. Cancer Res. 2018, 78, 6349–6353. [Google Scholar] [PubMed] [Green Version]
- Xiao, H.; Woods, E.C.; Vukojicic, P.; Bertozzi, C.R. Precision glycocalyx editing as a strategy for cancer immunotherapy. Proc. Natl. Acad. Sci. USA 2016, 113, 10304–10309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Sialylated glycans important in cancer.

Figure 2.
The siglec and sialoglycan axis in cancer. Siglec binding to hypersialylated tumour glycans blocks immune cell activation to promote immunosuppression.
Figure 2.
The siglec and sialoglycan axis in cancer. Siglec binding to hypersialylated tumour glycans blocks immune cell activation to promote immunosuppression.

{kind=link}
{kind=link}
Table 1.
Key examples of sialyltransferases important in cancer.
| Enzyme | Link to Cancer | Reference |
|---|---|---|
| ST6GAL1 | Upregulated in numerous cancer types (including pancreatic, prostate, breast and ovarian cancer). Role in tumour growth and metastasis. Linked to several pathways intrinsic to tumour cell biology. | [11,31,32,33] |
| ST3GAL4 | Upregulated in gastric carcinoma. Linked to poor prognosis, metastasis and the synthesis of sLeX. | [18,35] |
| ST3GAL6 | High expression correlates with reduced survival in multiple myeloma. Influences homing and engraftment to the bone marrow niche in vivo. Plays a key role in selectin ligand synthesis through generation of sLeX. | [36,37] |
| ST6GALNAC1 | Catalyses the sTn antigen and is associated with metastasis. | [8,27,38,39,40,41,42,43] |
| ST6GALNAC2 | Metastasis suppressor in breast cancer. Could be used to stratify patients for treatment with galectin-3 inhibitors. | [44,45] |
Table 2.
Overview of pre-clinical models and clinical trials targeting aberrant sialylation in cancer.
Table 2.
Overview of pre-clinical models and clinical trials targeting aberrant sialylation in cancer.
| Target | Approach | Reference or Identifier |
|---|---|---|
| Selectins | Selectin antagonist Uproleselan (GMI-1271) mimics SLeX. Uproleselan tested in pre-clinical models for breast cancer bone metastasis. | NCT03616470 phase 3 study in combination with chemotherapy for relapsed acute myeloid leukaemia. [55] |
| Sialylation | A sialic acid-blocking glycomimetic delivered using nanoparticles can inhibit metastasis and has been shown to be safe in pre-clinical models. | [26] |
| Siglecs | Siglecs-9, -7, -10 and -15 and their ligands have shown promise as targets to dampen anti-tumor immunity. | [62,63,64,65,66,67,68,69] |
| The EAGLE platform (that delivers a targeted sialidase enzyme to the tumours) is about to enter clinical trials for breast cancer | [70] | |
| sTn glycan | THERATOPE STn-KLH vaccine | NCT00003638 phase 3 clinical trial for metastatic breast cancer. [56,57,58] |
| Glycolipid GD2 | anti-GD2 antibody ch14.18/CHO (dinutuximab) | Neuroblastoma phase 3 trial [61] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Munkley, J.; Scott, E. Targeting Aberrant Sialylation to Treat Cancer. Medicines 2019, 6, 102. https://0-doi-org.brum.beds.ac.uk/10.3390/medicines6040102
AMA Style
Munkley J, Scott E. Targeting Aberrant Sialylation to Treat Cancer. Medicines. 2019; 6(4):102. https://0-doi-org.brum.beds.ac.uk/10.3390/medicines6040102
Chicago/Turabian StyleMunkley, Jennifer, and Emma Scott. 2019. "Targeting Aberrant Sialylation to Treat Cancer" Medicines 6, no. 4: 102. https://0-doi-org.brum.beds.ac.uk/10.3390/medicines6040102
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.