Proteomic and Phosphoproteomic Analysis Reveals that Neurokinin-1 Receptor (NK1R) Blockade with Aprepitant in Human Keratinocytes Activates a Distinct Subdomain of EGFR Signaling: Implications for the Anti-Pruritic Activity of NK1R Antagonists

 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of HaCaT Cells for RPPA Analysis
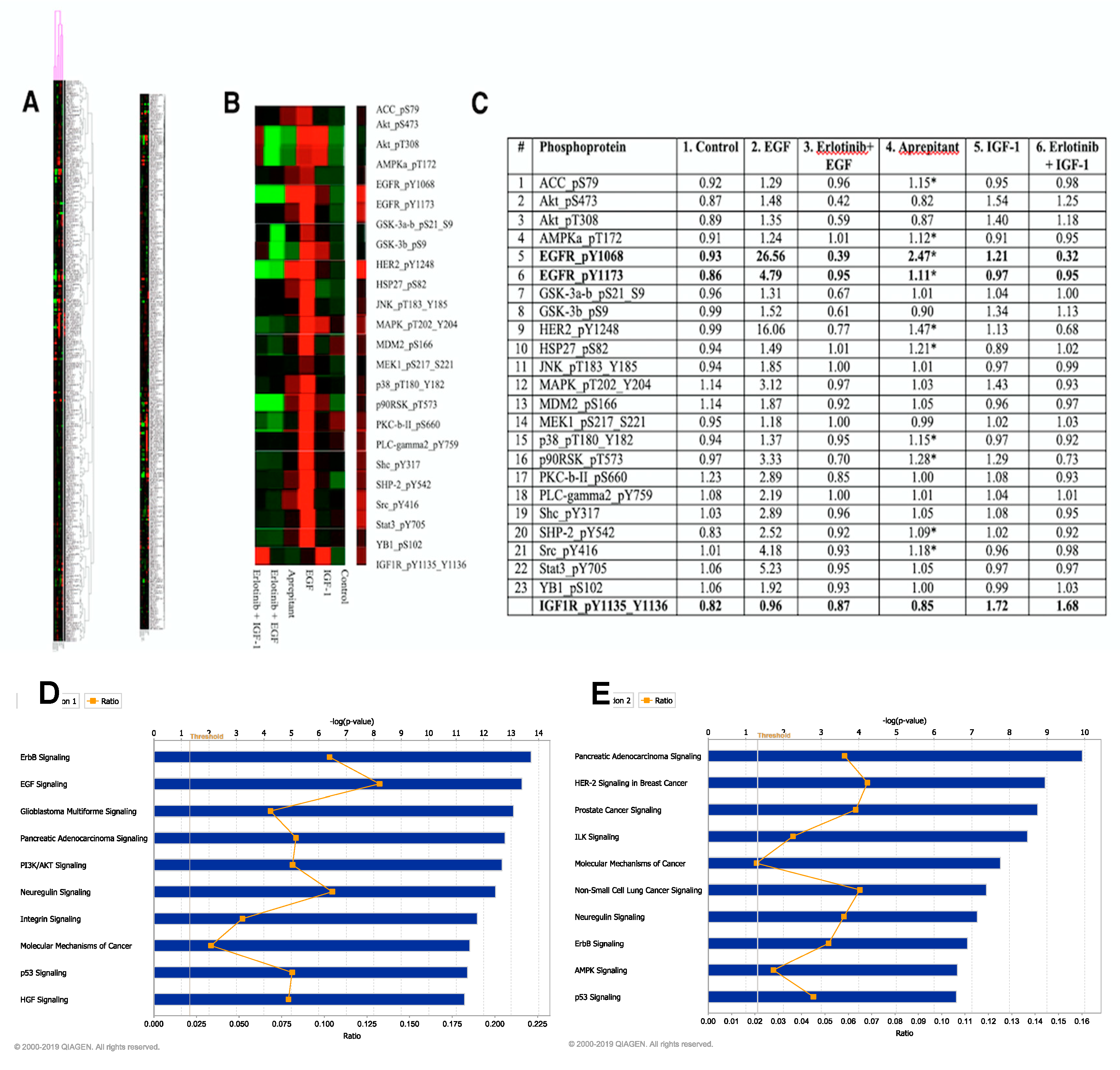
2.2. Ingenuity Pathway Analysis of RPPA Data
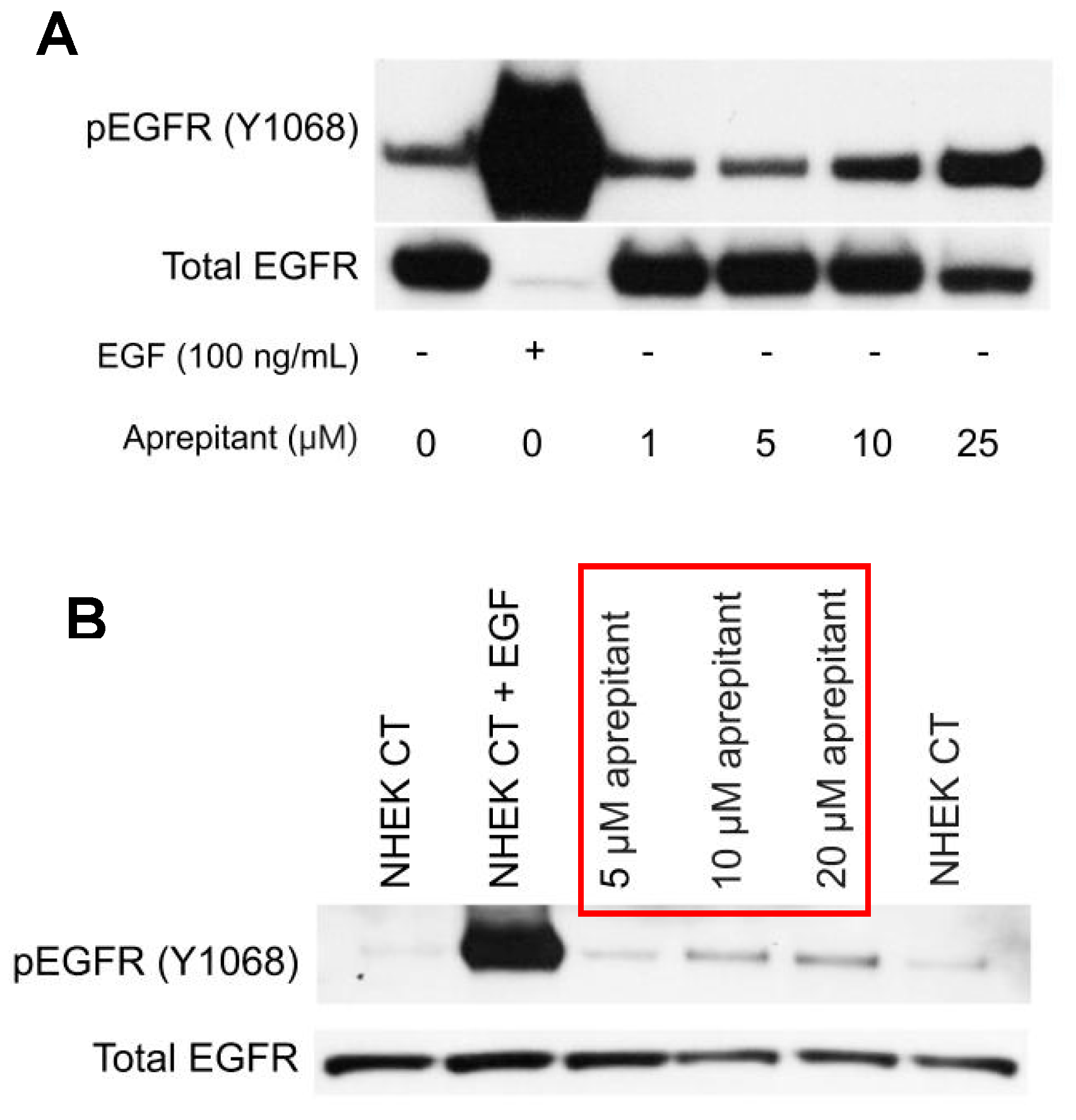
2.3. Western Blotting
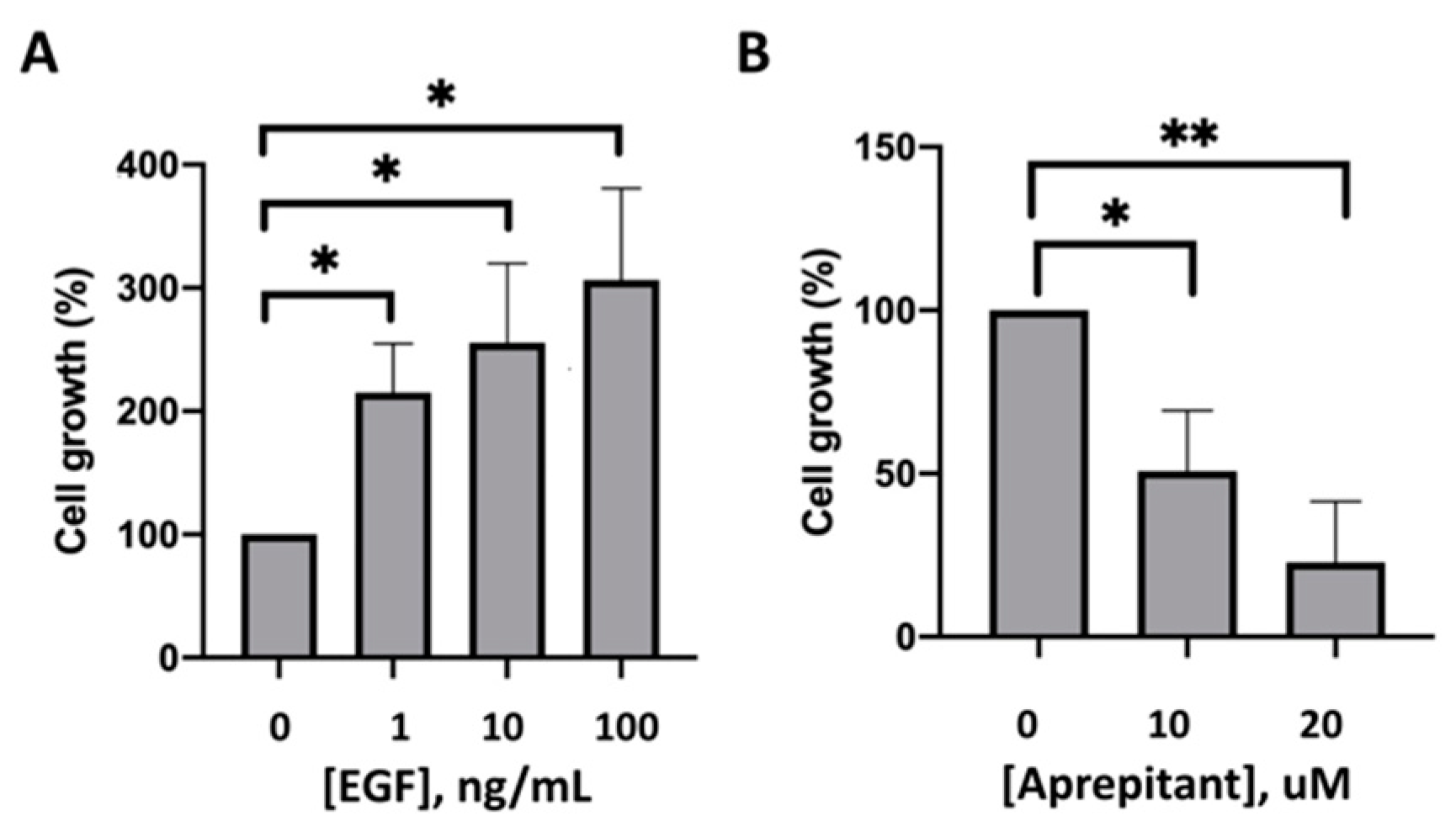
2.4. Effect of EGF and Aprepitant on the Growth of HaCaT Cells
3. Results
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Lacouture, M.E.; Anadkat, M.J.; Bensadoun, R.J.; Bryce, J.; Chan, A.; Epstein, J.B.; Eaby-Sandy, B.; Murphy, B.A. Clinical practice guidelines for the prevention and treatment of EGFR inhibitor-associated dermatologic toxicities. Support. Care Cancer 2011, 19, 1079–1095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, H.J.; Loftus, S.; Taylor, A.; DiCristina, C.; Green, S.; Zwaan, C.M. Aprepitant for the prevention of chemotherapy-induced nausea and vomiting in children: A randomised, double-blind, phase 3 trial. Lancet Oncol. 2015, 16, 385–394. [Google Scholar] [CrossRef]
- Kaul, S.; Kaffenberger, B.H.; Choi, J.N.; Kwatra, S.G. Cutaneous Adverse Reactions of Anticancer Agents. Dermatol. Clin. 2019, 37, 555–568. [Google Scholar] [CrossRef] [PubMed]
- Mascia, F.; Lam, G.; Keith, C.; Garber, C.; Steinberg, S.M.; Kohn, E.; Yuspa, S.H. Genetic Ablation of Epidermal EGFR Reveals the Dynamic Origin of Adverse Effects of Anti-EGFR Therapy. Sci. Transl. Med. 2013, 5, 199ra110. [Google Scholar] [CrossRef]
- Campbell, P.; Morton, P.E.; Takeichi, T.; Salam, A.; Roberts, N.; Proudfoot, L.E.; Mellerio, J.E.; Aminu, K.; Wellington, C.; Patil, S.N.; et al. Epithelial Inflammation Resulting from an Inherited Loss-of-Function Mutation in EGFR. J. Investig. Dermatol. 2014, 134, 2570–2578. [Google Scholar] [CrossRef] [Green Version]
- Lichtenberger, B.M.; Gerber, P.A.; Holcmann, M.; Buhren, B.A.; Amberg, N.; Smolle, V.; Schrumpf, H.; Boelke, E.; Ansari, P.; Mackenzie, C.; et al. Epidermal EGFR Controls Cutaneous Host Defense and Prevents Inflammation. Sci. Transl. Med. 2013, 5, 199ra111. [Google Scholar] [CrossRef]
- Larson, V.A.; Tang, O.; Stander, S.; Kang, S.; Kwatra, S.G. Association between itch and cancer in 16,925 patients with pruritus: Experience at a tertiary care center. J. Am. Acad. Dermatol. 2019, 80, 931–937. [Google Scholar] [CrossRef]
- Hassel, J.C.; Kripp, M.; Al-Batran, S.; Hofheinz, R.-D. Treatment of Epidermal Growth Factor Receptor Antagonist-Induced Skin Rash: Results of a Survey among German Oncologists. Oncol. Res. Treat. 2010, 33, 94–98. [Google Scholar] [CrossRef]
- He, A.; Alhariri, J.M.; Sweren, R.J.; Kwatra, M.M.; Kwatra, S.G. Aprepitant for the Treatment of Chronic Refractory Pruritus. Biomed. Res. Int. 2017, 2017. [Google Scholar] [CrossRef] [Green Version]
- Duval, A.; Dubertret, L. Aprepitant as an Antipruritic Agent? N. Engl. J. Med. 2009, 361, 1415–1416. [Google Scholar] [CrossRef]
- Vincenzi, B.; Tonini, G.; Santini, D. Aprepitant for Erlotinib-Induced Pruritus. N. Engl. J. Med. 2010, 363, 397–398. [Google Scholar] [CrossRef] [PubMed]
- Santini, D.; Vincenzi, B.; Guida, F.M.; Imperatori, M.; Schiavon, G.; Venditti, O.; Frezza, A.M.; Berti, P.; Tonini, G. Aprepitant for management of severe pruritus related to biological cancer treatments: A pilot study. Lancet Oncol. 2012, 13, 1020–1024. [Google Scholar] [CrossRef]
- Gerber, P.A.; Buhren, B.A.; Homey, B. More on Aprepitant for Erlotinib-Induced Pruritus. N. Engl. J. Med. 2011, 364, 486–487. [Google Scholar] [PubMed]
- Marqués, M.M.; Martínez, N.; Rodríguez-García, I.; Alonso, A. EGFR Family-Mediated Signal Transduction in the Human Keratinocyte Cell Line HaCaT. Exp. Cell Res. 1999, 252, 432–438. [Google Scholar] [CrossRef] [PubMed]
- Brown, K.E.; Chagoya, G.; Kwatra, S.G.; Yen, T.; Keir, S.T.; Cooter, M.; Hoadley, K.A.; Rasheed, A.; Lipp, E.S.; Mclendon, R.; et al. Proteomic profiling of patient-derived glioblastoma xenografts identifies a subset with activated EGFR: Implications for drug development. J. Neurochem. 2015, 133, 730–738. [Google Scholar] [CrossRef] [Green Version]
- Gross, K.; Karagiannides, I.; Thomou, T.; Koon, H.W.; Bowe, C.; Kim, H.; Giorgadze, N.; Tchkonia, T.; Pirtskhalava, T.; Kirkland, J.L.; et al. Substance P promotes expansion of human mesenteric preadipocytes through proliferative and antiapoptotic pathways. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 296, G1012–G1019. [Google Scholar] [CrossRef] [Green Version]
- Castagliuolo, I.; Valenick, L.; Liu, J.; Pothoulakis, C. Epidermal growth factor receptor transactivation mediates substance P-induced mitogenic responses in U-373 MG cells. J. Biol. Chem. 2000, 275, 26545–26550. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.G.; Yu, J.; Hu, J.L.; Yang, W.L.; Ren, H.; Ding, D.; Zhang, L.; Liu, X.P. Neurokinin-1 activation affects EGFR-related signal transduction in triple ngetaive breast cancer. Cell Signal. 2015, 27, 1315–1324. [Google Scholar] [CrossRef]
- Ando, Y.; Jensen, P.J. Epidermal Growth Factor and Insulin-Like Growth Factor I Enhance Keratinocyte Migration. J. Investig. Dermatol. 1993, 100, 633–639. [Google Scholar] [CrossRef] [Green Version]
- Munoz, M.; Rosso, M. The NK-1 receptor antagonist aprepitant as a broad spectrum antitumor drug. Investig. New Drugs 2010, 28, 187–193. [Google Scholar] [CrossRef]
- Muñoz, M.; Rosso, M.; Robles-Frias, M.J.; Salinas-Martín, M.V.; Rosso, R.; González-Ortega, A.; Coveñas, R. The NK-1 receptor is expressed in human melanoma and is involved in the antitumor action of the NK-1 receptor antagonist aprepitant on melanoma cell lines. Lab Investig. 2010, 90, 1259–1269. [Google Scholar] [CrossRef] [Green Version]
- Kwatra, S.G.; Boozalis, E.; Kwatra, M.M. Effects of neuroimmune axis modulation by aprepitant on antipruritic and global disease severity in patients with cutaneous T-cell lymphoma. Br. J. Dermatol. 2018, 178, 1221–1222. [Google Scholar] [CrossRef] [PubMed]
- Lacouture, M.E.; Rodeck, U. Skinflammation and Drug Toxicity—A Delicate Balance. Sci. Transl. Med. 2013, 5, 199fs33. [Google Scholar] [CrossRef] [PubMed]
- Chanprapaph, K.; Vachiramon, V.; Rattanakaemakorn, P. Epidermal Growth Factor Receptor Inhibitors: A Review of Cutaneous Adverse Events and Management. Dermatol. Res. Pract. 2014, 2014, 734249. [Google Scholar] [CrossRef] [PubMed] [Green Version]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kwatra, S.G.; Boozalis, E.; Huang, A.H.; Nanni, C.; Khanna, R.; Williams, K.A.; Semenov, Y.R.; Roberts, C.M.; Burns, R.F.; Krischak, M.; et al. Proteomic and Phosphoproteomic Analysis Reveals that Neurokinin-1 Receptor (NK1R) Blockade with Aprepitant in Human Keratinocytes Activates a Distinct Subdomain of EGFR Signaling: Implications for the Anti-Pruritic Activity of NK1R Antagonists. Medicines 2019, 6, 114. https://0-doi-org.brum.beds.ac.uk/10.3390/medicines6040114
Kwatra SG, Boozalis E, Huang AH, Nanni C, Khanna R, Williams KA, Semenov YR, Roberts CM, Burns RF, Krischak M, et al. Proteomic and Phosphoproteomic Analysis Reveals that Neurokinin-1 Receptor (NK1R) Blockade with Aprepitant in Human Keratinocytes Activates a Distinct Subdomain of EGFR Signaling: Implications for the Anti-Pruritic Activity of NK1R Antagonists. Medicines. 2019; 6(4):114. https://0-doi-org.brum.beds.ac.uk/10.3390/medicines6040114
Chicago/Turabian StyleKwatra, Shawn G., Emily Boozalis, Amy H. Huang, Cory Nanni, Raveena Khanna, Kyle A. Williams, Yevgeniy R. Semenov, Callie M. Roberts, Robert F. Burns, Madison Krischak, and et al. 2019. "Proteomic and Phosphoproteomic Analysis Reveals that Neurokinin-1 Receptor (NK1R) Blockade with Aprepitant in Human Keratinocytes Activates a Distinct Subdomain of EGFR Signaling: Implications for the Anti-Pruritic Activity of NK1R Antagonists" Medicines 6, no. 4: 114. https://0-doi-org.brum.beds.ac.uk/10.3390/medicines6040114