
Early Time-Restricted Feeding Amends Circadian Clock Function and Improves Metabolic Health in Male and Female Nile Grass Rats

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals and Feeding Protocol
2.2. Blood Parameters
2.3. Tissue Collection and Histological Analysis
2.4. Transcriptional Changes
2.5. Quantification of Hepatic Metabolites
2.6. Statistical Analysis
3. Results
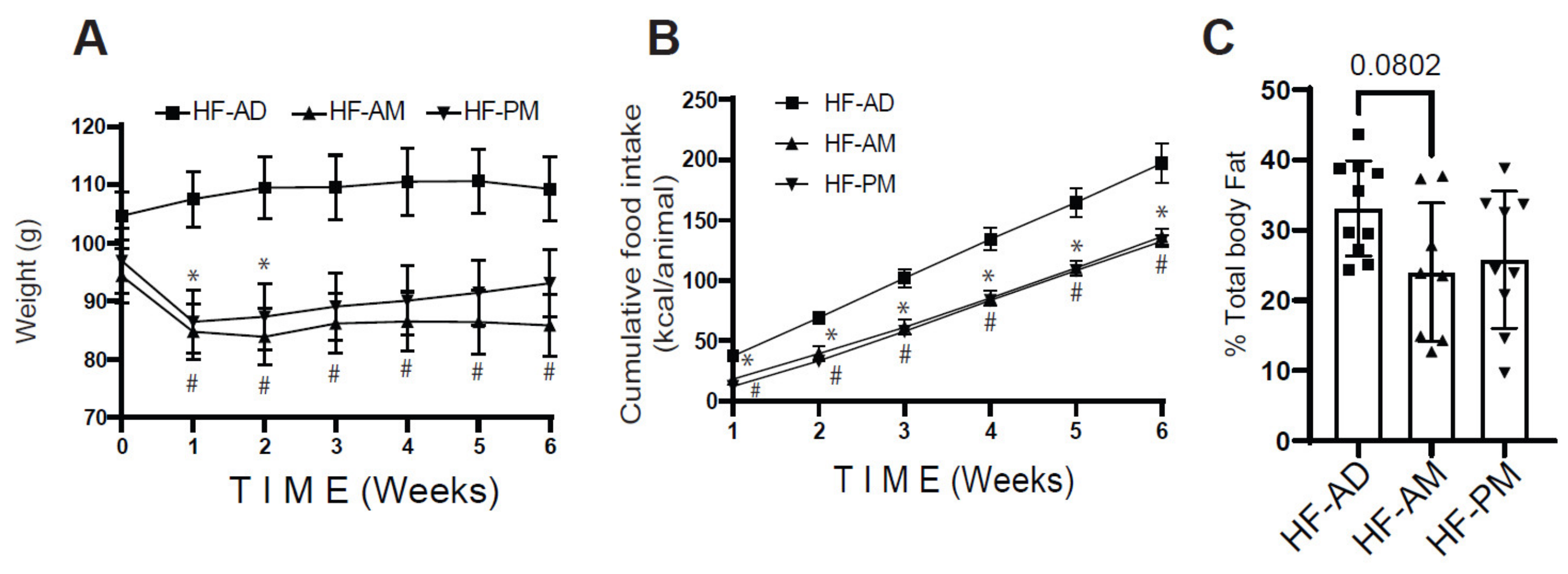
3.1. Early Time-Restricted High-Fat Feeding Reduced Weight Gain
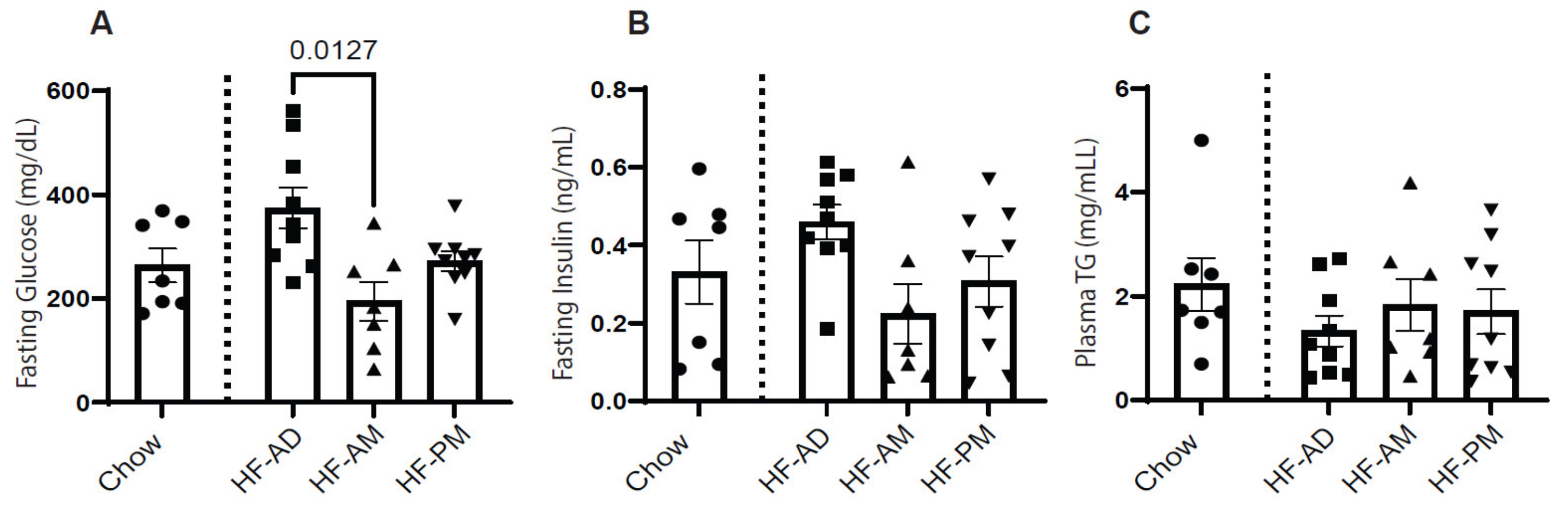
3.2. Time-Restricted High Fat Feeding Altered Fasting Blood Glucose Level
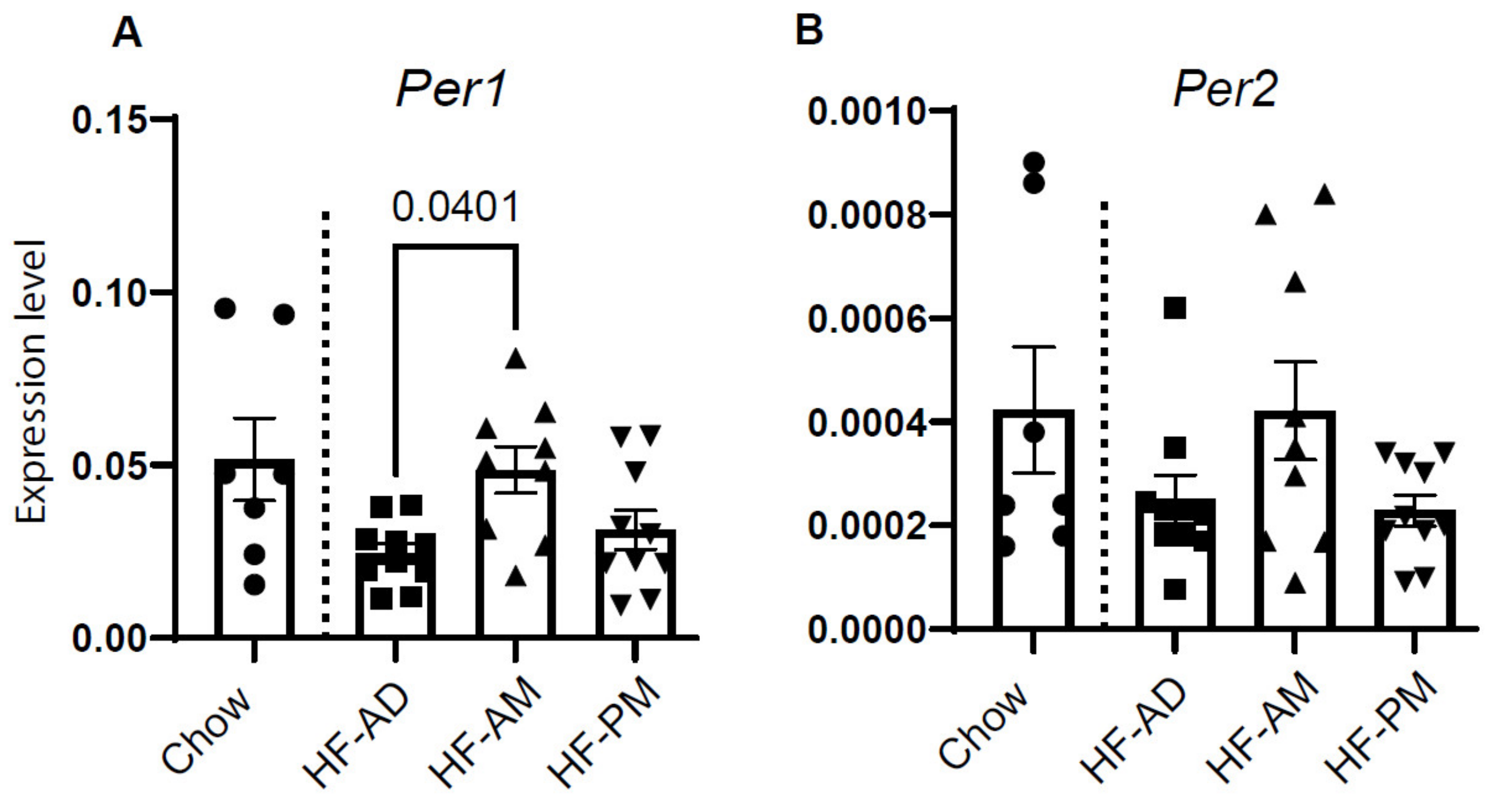
3.3. Early TRF Improves Liver Clock Gene Expression
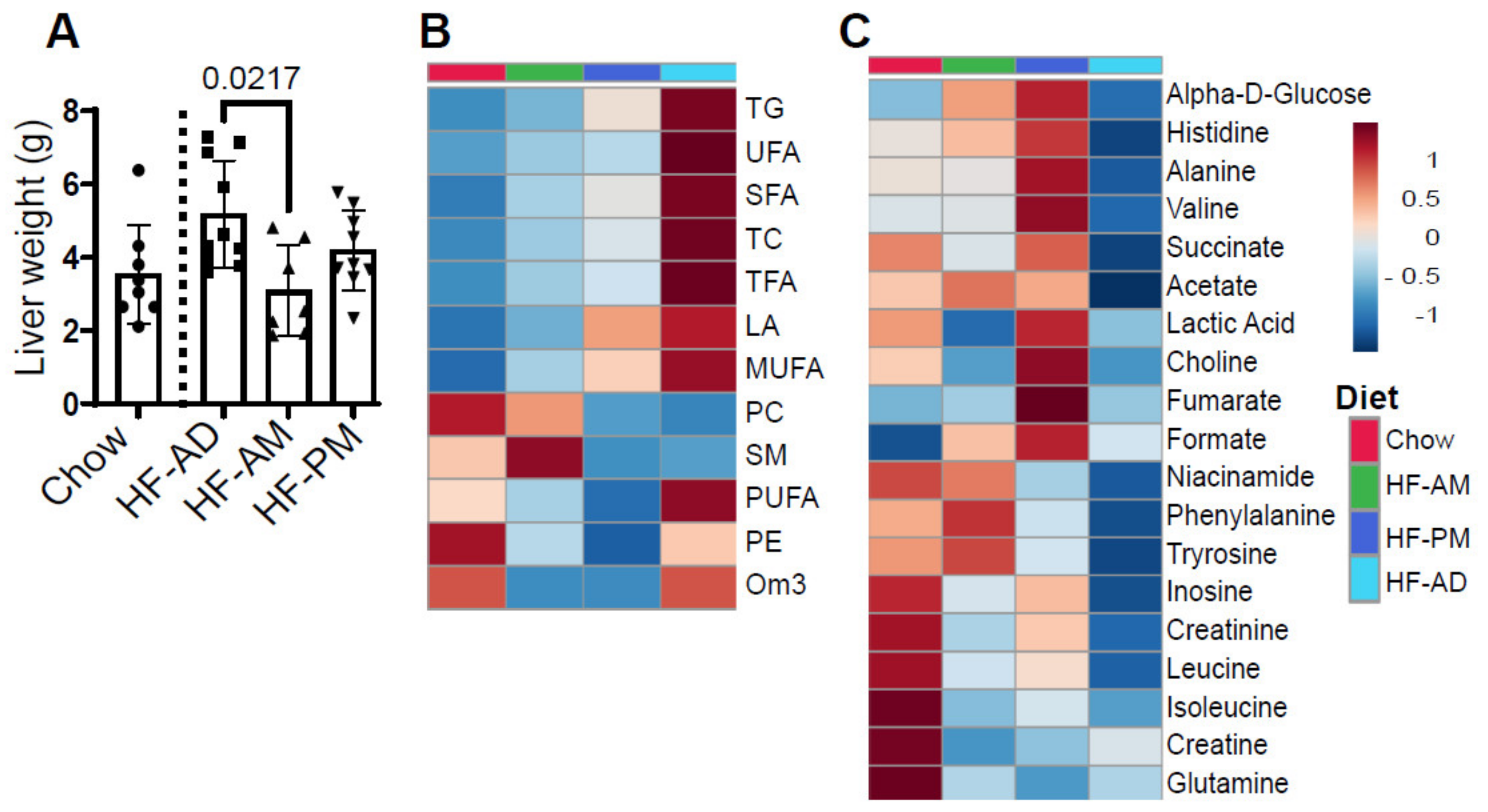
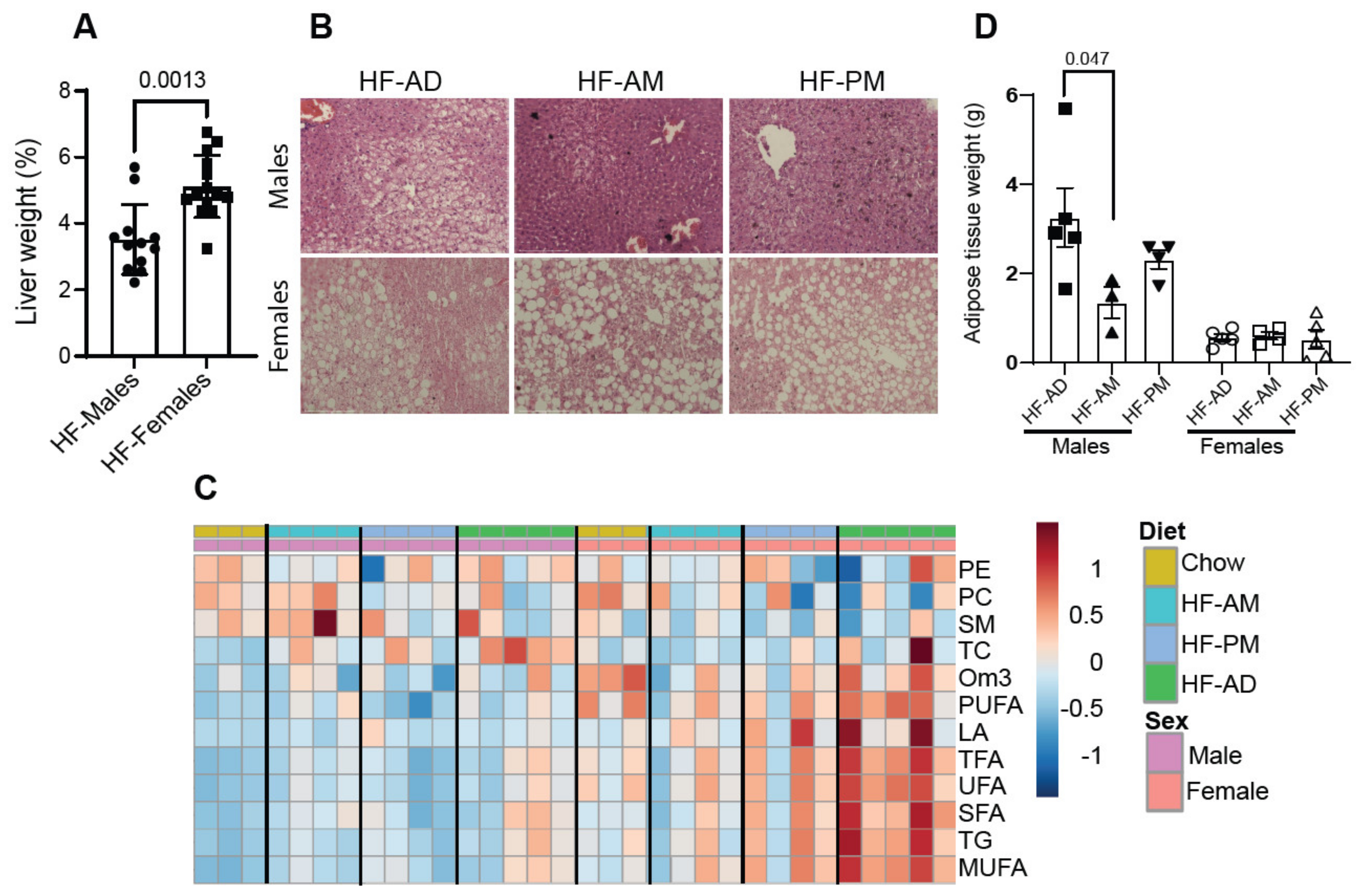
3.4. Early TRF Decreases Liver Weight and Alters Hepatic Metabolic Processes
3.5. Lipid Distribution Differs between Male and Female NGRs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Hirode, G.; Wong, R.J. Trends in the Prevalence of Metabolic Syndrome in the United States, 2011–2016. J. Am. Med. Assoc. 2020, 323, 2526–2528. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.; Panda, S. A Smartphone App Reveals Erratic Diurnal Eating Patterns in Humans that Can Be Modulated for Health Benefits. Cell Metab. 2015, 22, 789–798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suliga, E.; Kozieł, D.; Cieśla, E.; Rębak, D.; Głuszek, S. Dietary patterns in relation to metabolic syndrome among adults in Poland: A cross-sectional study. Nutrients 2017, 9, 1366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaix, A.; Lin, T.; Le, H.D.; Chang, M.W.; Panda, S. Time-Restricted Feeding Prevents Obesity and Metabolic Syndrome in Mice Lacking a Circadian Clock. Cell Metab. 2019, 29, 303–319.e4. [Google Scholar] [CrossRef]
- Manoogian, E.N.C.; Panda, S. Circadian rhythms, time-restricted feeding, and healthy aging. Ageing Res. Rev. 2017, 39, 59–67. [Google Scholar] [CrossRef]
- Rothschild, J.; Hoddy, K.K.; Jambazian, P.; Varady, K.A. Time-restricted feeding and risk of metabolic disease: A review of human and animal studies. Nutr. Rev. 2014, 72, 308–318. [Google Scholar] [CrossRef]
- Sutton, E.F.; Beyl, R.; Early, K.S.; Cefalu, W.T.; Ravussin, E.; Peterson, C.M. Early Time-Restricted Feeding Improves Insulin Sensitivity, Blood Pressure, and Oxidative Stress Even without Weight Loss in Men with Prediabetes. Cell Metab. 2018, 27, 1212–1221.e3. [Google Scholar] [CrossRef] [Green Version]
- Stephan, F.K. The “other” circadian system: Food as a zeitgeber. J. Biol. Rhythm. 2002, 17, 284–292. [Google Scholar] [CrossRef]
- Reppert, S.M.; Weaver, D.R. Coordination of circadian timing in mammals. Nature 2002, 418, 935–941. [Google Scholar] [CrossRef]
- Takahashi, J.S. Transcriptional architecture of the mammalian circadian clock. Nat. Rev. Genet. 2017, 18, 164–179. [Google Scholar] [CrossRef] [Green Version]
- Rijo-Ferreira, F.; Takahashi, J.S. Genomics of circadian rhythms in health and disease. Genome Med. 2019, 11, 82. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Lahens, N.F.; Ballance, H.I.; Hughes, M.E.; Hogenesch, J.B. A circadian gene expression atlas in mammals: Implications for biology and medicine. Proc. Natl. Acad. Sci. USA 2014, 111, 16219–16224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delahaye, L.B.; Bloomer, R.J.; Butawan, M.B.; Wyman, J.M.; Hill, J.L.; Lee, H.W.; Liu, A.C.; McAllan, L.; Han, J.C.; van der Merwe, M. Time-restricted feeding of a high-fat diet in male C57BL/6 mice reduces adiposity but does not protect against increased systemic inflammation. Appl. Physiol. Nutr. Metab. 2018, 43, 1033–1042. [Google Scholar] [CrossRef] [PubMed]
- Hatori, M.; Vollmers, C.; Zarrinpar, A.; DiTacchio, L.; Bushong, E.A.; Gill, S.; Leblanc, M.; Chaix, A.; Joens, M.; Fitzpatrick, J.A.J.; et al. Time-restricted feeding without reducing caloric intake prevents metabolic diseases in mice fed a high-fat diet. Cell Metab. 2012, 15, 848–860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Refinetti, R. The nile grass rat as a laboratory animal. Lab Anim. 2004, 33, 54–57. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Smale, L.; Nunez, A.A. Circadian and photic modulation of daily rhythms in diurnal mammals. Eur. J. Neurosci. 2020, 51, 551–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noda, K.; Melhorn, M.I.; Zandi, S.; Frimmel, S.; Tayyari, F.; Hisatomi, T.; Almulki, L.; Pronczuk, A.; Hayes, K.C.; Hafezi-Moghadam, A. An animal model of spontaneous metabolic syndrome: Nile grass rat. FASEB J. 2010, 24, 2443–2453. [Google Scholar] [CrossRef] [Green Version]
- Chaabo, F.; Pronczuk, A.; Maslova, E.; Hayes, K.C. Nutritional correlates and dynamics of diabetes in the Nile rat (Arvicanthis niloticus): A novel model for diet-induced type 2 diabetes and the metabolic syndrome. Nutr. Metab. 2010, 7, 29. [Google Scholar] [CrossRef] [Green Version]
- Ran, D.H.; Lin, W.J.; Zhen, R.H.; Lei, S.X. Lipometabolism and glycometabolism in liver diseases. BioMed Res. Int. 2018, 2018, 1287127. [Google Scholar]
- Simona D’Amore Giuseppe Palasciano, A.M. The Liver in Metabolic Syndrome. In A Systems Biology Approach to Study Metabolic Syndrome; Orešič, M., Vidal-Puig, A., Eds.; Springer: Cham, Switzerland, 2014. [Google Scholar] [CrossRef]
- Raja, G.; Gupta, H.; Gebru, Y.A.; Youn, G.S.; Choi, Y.R.; Kim, H.S.; Yoon, S.J.; Kim, D.J.; Kim, T.J.; Suk, K.T. Recent advances of microbiome-associated metabolomics profiling in liver disease: Principles, mechanisms, and applications. Int. J. Mol. Sci. 2021, 22, 1160. [Google Scholar] [CrossRef]
- Sostare, J.; di Guida, R.; Kirwan, J.; Chalal, K.; Palmer, E.; Dunn, W.B.; Viant, M.R. Comparison of modified Matyash method to conventional solvent systems for polar metabolite and lipid extractions. Anal. Chim. Acta. 2018, 1037, 301–315. [Google Scholar] [CrossRef] [PubMed]
- Johnson, H.; Puppa, M.; van der Merwe, M.; Tipirneni-Sajja, A. CRAFT for NMR lipidomics: Targeting lipid metabolism in leucine-supplemented tumor-bearing mice. Magn. Reson. Chem. 2021, 59, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Krishnamurthy, K. CRAFT (complete reduction to amplitude frequency table)–Robust and time-efficient Bayesian approach for quantitative mixture analysis by NMR. Magn. Reson. Chem. 2013, 51, 821–829. [Google Scholar] [CrossRef] [PubMed]
- Wishart, D.S.; Feunang, Y.D.; Marcu, A.; Guo, A.C.; Liang, K.; Vázquez-Fresno, R.; Sajed, T.; Johnson, D.; Li, C.; Karu, N.; et al. HMDB 4.0: The human metabolome database for 2018. Nucleic Acids Res. 2018, 46, D608–D617. [Google Scholar] [CrossRef]
- Wishart, D.S.; Jewison, T.; Guo, A.C.; Wilson, M.; Knox, C.; Liu, Y.; Djoumbou, Y.; Mandal, R.; Aziat, F.; Dong, E.; et al. HMDB 3.0-The Human Metabolome Database in 2013. Nucleic Acids Res. 2013, 41, D801–D807. [Google Scholar] [CrossRef]
- Amiel, A.; Tremblay-Franco, M.; Gautier, R.; Ducheix, S.; Montagner, A.; Polizzi, A.; Debrauwer, L.; Guillou, H.; Bertrand-Michel, J.; Canlet, C. Proton NMR enables the absolute quantification of aqueous metabolites and lipid classes in unique mouse liver samples. Metabolites 2020, 10, 9. [Google Scholar] [CrossRef] [Green Version]
- Chi, Y.; Gupta, R.K. Alterations in membrane fatty acid unsaturation and chain length in hypertension as observed by 1H NMR spectroscopy. Am. J. Hypertens. 1998, 11, 340–348. [Google Scholar] [CrossRef] [Green Version]
- Światkiewicz, I.; Woźniak, A.; Taub, P.R. Time-restricted eating and metabolic syndrome: Current status and future perspectives. Nutrients 2021, 13, 221. [Google Scholar] [CrossRef]
- Jamshed, H.; Beyl, R.A.; della Manna, D.L.; Yang, E.S.; Ravussin, E.; Peterson, C.M. Early Time-Restricted Feeding Improves 24-Hour. Nutrients 2019, 11, 1234. [Google Scholar] [CrossRef] [Green Version]
- Chaix, A.; Zarrinpar, A.; Miu, P.; Panda, S. Time-restricted feeding is a preventative and therapeutic intervention against diverse nutritional challenges. Cell Metab. 2014, 20, 991–1005. [Google Scholar] [CrossRef] [Green Version]
- Turek, F.W.; Joshu, C.; Kohsaka, A.; Lin, E.; Ivanova, G.; McDearmon, E.; Laposky, A.; Losee-Olson, S.; Easton, A.; Jensen, D.R.; et al. Obesity and metabolic syndrome in circadian Clock mutant nice. Science 2005, 308, 1043–1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohsaka, A.; Laposky, A.D.; Ramsey, K.M.; Estrada, C.; Joshu, C.; Kobayashi, Y.; Turek, F.W.; Bass, J. High-Fat Diet Disrupts Behavioral and Molecular Circadian Rhythms in Mice. Cell Metab. 2007, 6, 414–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vollmers, C.; Gill, S.; DiTacchio, L.; Pulivarthy, S.R.; Le, H.D.; Panda, S. Time of feeding and the intrinsic circadian clock drive rhythms in hepatic gene expression. Proc. Natl. Acad. Sci. USA 2009, 106, 21453–21458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wishart, D.S. NMR metabolomics: A look ahead. J. Magn. Reson. 2019, 306, 155–161. [Google Scholar] [CrossRef]
- Ryan, E.P.; Heuberger, A.L.; Broeckling, C.D.; Borresen, E.C.; Tillotson, C.; Prenni, J.E. Advances in Nutritional Metabolomics. Curr. Metab. 2013, 1, 109–120. [Google Scholar] [CrossRef] [Green Version]
- Guasch-Ferre, M.; Bhupathiraju, S.N.; Hu, F.B. Use of metabolomics in improving assessment of dietary intake. Clin. Chem. 2018, 64, 82–98. [Google Scholar] [CrossRef] [Green Version]
- An, Y.; Xu, W.; Li, H.; Lei, H.; Zhang, L.; Hao, F.; Duan, Y.; Yan, X.; Zhao, Y.; Wu, J.; et al. High-fat diet induces dynamic metabolic alterations in multiple biological matrices of rats. J. Proteome. Res. 2013, 12, 3755–3768. [Google Scholar] [CrossRef]
- Xie, Z.; Li, H.; Wang, K.; Lin, J.; Wang, Q.; Zhao, G.; Jia, W.; Zhang, Q. Analysis of transcriptome and metabolome profiles alterations in fatty liver induced by high-fat diet in rat. Metabolism 2010, 59, 554–560. [Google Scholar] [CrossRef]
- Green, C.J.; Hodson, L. The influence of dietary fat on liver fat accumulation. Nutrients 2014, 6, 5018–5033. [Google Scholar] [CrossRef]
- Tsuru, H.; Osaka, M.; Hiraoka, Y.; Yoshida, M. HFD-induced hepatic lipid accumulation and inflammation are decreased in Factor D deficient mouse. Sci. Rep. 2020, 10, 17593. [Google Scholar] [CrossRef]
- Yaligar, J.; Gopalan, V.; Kiat, O.W.; Sugii, S.; Shui, G.; Lam, B.D.; Henry, C.J.; Wenk, M.R.; Tai, E.S.; Velan, S.S. Evaluation of dietary effects on hepatic lipids in high fat and placebo diet fed rats by in vivo MRS and LC-MS techniques. PLoS ONE 2014, 9, e91436. [Google Scholar] [CrossRef]
- Sundaram, S.; Yan, L. Time-restricted feeding reduces adiposity in mice fed a high-fat diet. Nutr. Res. 2016, 36, 603–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, Y.; Xu, H.; Xie, Z.; Wang, L.; Sun, Y.; Yang, H.; Hu, D.; Mao, Y. Time-Restricted Feeding Reduces the Detrimental Effects of a High-Fat Diet, Possibly by Modulating the Circadian Rhythm of Hepatic Lipid Metabolism and Gut Microbiota. Front. Nutr. 2020, 7, 596285. [Google Scholar] [CrossRef]
- Schneider, J.; Han, W.H.; Matthew, R.; Sauvé, Y.; Lemieux, H. Age and sex as confounding factors in the relationship between cardiac mitochondrial function and type 2 diabetes in the Nile Grass rat. PLoS ONE 2020, 15, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Chaix, A.; Deota, S.; Bhardwaj, R.; Lin, T.; Panda, S. Sex- and age-dependent outcomes of 9-hour time-restricted feeding of a Western high-fat high-sucrose diet in C57BL/6J mice. Cell Rep. 2021, 36, 109543. [Google Scholar] [CrossRef] [PubMed]
- Jakubowicz, D.; Wainstein, J.; Tsameret, S.; Landau, Z. Role of high energy breakfast “big breakfast diet” in clock gene regulation of postprandial hyperglycemia and weight loss in type 2 diabetes. Nutrients 2021, 13, 1558. [Google Scholar] [CrossRef]
- Charlot, A.; Hutt, F.; Sabatier, E.; Zoll, J. Beneficial effects of early time-restricted feeding on metabolic diseases: Importance of aligning food habits with the circadian clock. Nutrients 2021, 13, 1405. [Google Scholar] [CrossRef]
- Dashti, H.S.; Gómez-Abellán, P.; Qian, J.; Esteban, A.; Morales, E.; Scheer, F.A.J.L.; Garaulet, M. Late eating is associated with cardiometabolic risk traits, obesogenic behaviors, and impaired weight loss. Am. J. Clin. Nutr. 2021, 113, 154–161. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolite | Chemical Shift & Multiplicity | HF-AD | HF-AM | HF-EV | p-Value |
|---|---|---|---|---|---|
| PC | 3.14 (s) | 2.7 ± 0.6 | 3.1 ± 0.4 | 2.7 ± 0.5 | 0.14 |
| PE | 3.08 (t) | 1.6 ± 0.6 | 1.5 ± 0.1 | 1.4 ± 0.6 | 0.70 |
| SM | 3.13 (s) | 0.2 ± 0.1 | 0.3 ± 0.1 | 0.2 ± 0.1 | 0.25 |
| Om3 | 0.63 (s) | 6.9 ± 2.4 | 5.1 ±2.1 | 5.1 ± 2.0 | 0.15 |
| TC | 4.27 (dd) | 4.7 ± 2.2 | 2.8 ± 1.1 | 3.2 ± 1.2 | 0.048 |
| Tg | 0.92 (t) | 49.6 ± 26.9 | 22.9 ± 13.8 | 31.6 ± 21.2 | 0.046 * |
| TFA | 2.73 (t) | 154.6 ± 65.7 | 98.5 ± 37.0 | 105.7 ± 60.7 | 0.09 |
| LA | UFA minus MUFA | 32.9 ± 33.0 | 16.2 ± 10.6 | 27.1 ± 24.7 | 0.39 |
| UFA | 0.83 (m) | 96.9 ± 41.1 | 64.5 ± 25.9 | 67.1 ± 39.0 | 0.13 |
| SFA | SFA-% times TFA | 57.7 ± 26.5 | 34.0 ± 12.0 | 38.6 ± 22.1 | 0.07 |
| MUFA | 1.97 (m) | 46.1 ± 22.6 | 25.6 ± 13.3 | 33.1 ± 19.1 | 0.09 |
| PUFA | TFA minus UFA | 50.8 ± 19.9 | 38.9 ± 14.0 | 34.0 ± 21.2 | 0.17 |
| UFA-% | From equation | 63% ± 6% | 65% ± 2% | 64% ± 4% | 0.66 |
| SFA-% | 100 minus UFA-% | 37% ± 6% | 35% ± 2% | 36% ± 4% | 0.66 |
| MUFA-% | MUFA divided by TFA | 29% ± 5% | 25% ± 3% | 33% ± 12% | 0.20 |
| PUFA-% | UFA-% minus MUFA-% | 34% ± 9% | 40% ± 4% | 31% ± 11% | 0.17 |
| Metabolite | Chemical Shift & Multiplicity | HF-AD | HF-AM | HF-EV | p-Value |
|---|---|---|---|---|---|
| Acetate | 1.79 (s) | 0.17 ± 0.04 | 0.25 ± 0.05 | 0.24 ± 0.06 | 0.005 *‡ |
| Alanine | 1.35 (d) | 0.42 ± 0.12 | 0.52 ± 0.10 | 0.63 ± 0.20 | 0.022 ‡ |
| Choline | 3.08 (s) | 0.11 ± 0.05 | 0.11 ± 0.06 | 0.13 ± 0.04 | 0.66 |
| Creatine | 2.91 (s) | 0.14 ± 0.11 | 0.09 ± 0.03 | 0.11 ± 0.11 | 0.55 |
| Creatinine | 2.92(s) | 0.03 ± 0.02 | 0.03 ± 0.01 | 0.04 ± 0.02 | 0.47 |
| Formate | 8.33 (s) | 0.07 ± 0.05 | 0.07 ± 0.05 | 0.09 ± 0.07 | 0.70 |
| Fumarate | 6.39 (s) | 0.06 ± 0.03 | 0.06 ± 0.02 | 0.11 ± 0.03 | 0.002 ‡‡ |
| Glucose | 5.11 (d) | 7.89 ± 3.30 | 10.58 ± 3.86 | 11.58 ± 3.37 | 0.09 |
| Glutamine | 2.32 (m) | 1.10 ± 0.46 | 1.10 ± 0.36 | 0.97 ± 0.30 | 0.751 |
| Histidine | 7.75 (s) | 0.07 ± 0.02 | 0.10 ± 0.03 | 0.11 ± 0.04 | 0.09 |
| Inosine | 8.22 (s) | 0.62 ± 0.22 | 0.80 ± 0.27 | 0.88 ± 0.18 | 0.06 |
| Isoleucine | 0.89 (t) | 0.08 ± 0.09 | 0.09 ± 0.09 | 0.12 ± 0.12 | 0.67 |
| Lactate | 1.20 (d) | 1.56 ± 0.69 | 1.39 ± 0.52 | 2.00 ± 0.94 | 0.25 |
| Leucine | 0.83 (m) | 0.10 ± 0.04 | 0.12 ± 0.07 | 0.13 ± 0.05 | 0.47 |
| Niacinamide | 8.81 (d) & 7.48 (m) | 0.06 ± 0.02 | 0.08 ± 0.03 | 0.07 ± 0.02 | 0.08 |
| Phenylalanine | 7.20 (d) & 7.29 (d) | 0.02 ± 0.02 | 0.04 ± 0.02 | 0.03 ± 0.02 | 0.14 |
| Succinate | 2.27 (s) | 0.44 ± 0.28 | 0.55 ± 0.21 | 0.62 ± 0.16 | 0.28 |
| Tyrosine | 6.78 (d) & 7.06 (d) | 0.02 ± 0.01 | 0.04 ± 0.02 | 0.03 ± 0.02 | 0.14 |
| Valine | 0.91 (d) | 0.10 ± 0.02 | 0.14 ± 0.08 | 0.20 ± 0.16 | 0.13 |
| Metabolite | Male | Female | Sex | Diet | Interaction | ||||
|---|---|---|---|---|---|---|---|---|---|
| HF-AD | HF-AM | HF-EV | HF-AD | HF-AM | HF-EV | ||||
| PC | 2.8 ± 0.5 | 3.3 ± 0.3 | 2.8 ± 0.2 | 2.5 ± 0.7 | 3.0 ± 0.4 | 2.6 ± 0.8 | 0.26 | 0.16 | 0.94 |
| PE | 1.7 ± 0.3 | 1.5 ± 0.2 | 1.3 ± 0.7 | 1.4 ± 0.8 | 1.5 ± 0.1 | 1.4 ± 0.6 | 0.69 | 0.73 | 0.82 |
| SM | 0.3 ± 0.1 | 0.4 ± 0.1 | 0.3 ± 0.1 | 0.2 ± 0.1 | 0.2 ± 0.05 | 0.2 ± 0.1 | 0.018 m | 0.20 | 0.53 |
| Om3 | 5.7 ± 2.0 | 4.8 ± 2.0 | 4.2 ± 1.9 | 8.2 ± 2.3 | 5.5 ± 2.5 | 6.1 ± 1.7 | 0.049 f | 0.13 | 0.64 |
| TC | 5.1 ± 1.5 | 3.6 ± 1.0 | 3.5 ± 1.5 | 4.3 ± 2.9 | 2.1 ± 0.5 | 2.8 ± 0.9 | 0.14 | 0.05 | 0.89 |
| Tg | 30.6 ± 15.8 | 17.9 ± 5.9 | 17.7 ± 9.9 | 68.6 ± 21.8 | 27.9 ± 18.5 | 45.6 ± 20.8 | 0.001 f | 0.009 # | 0.59 |
| TFA | 103.3 ± 37.5 | 90.0 ± 19.9 | 62.9 ± 28.1 | 205.8 ± 41.7 | 107.1 ± 51.1 | 148.5 ± 54.1 | <0.001 f | 0.015 #* | 0.09 |
| LA | 14.3 ± 5.7 | 10.3 ± 6.6 | 15.6 ± 12.1 | 51.5 ± 39.5 | 22.1 ± 11.3 | 38.5 ± 30.4 | 0.014 f | 0.31 | 0.50 |
| UFA | 63.7 ± 20.1 | 55.5 ± 13.6 | 38.9 ± 13.9 | 130.1 ± 25.4 | 73.5 ± 34.2 | 95.3 ± 35.2 | <0.001 f | 0.020 #* | 0.14 |
| SFA | 39.6 ± 18.5 | 34.4 ± 7.1 | 24.0 ± 14.3 | 75.7 ± 20.6 | 33.6 ± 16.9 | 53.1 ± 19.1 | 0.004 f | 0.017 # | 0.08 |
| MUFA | 28.4 ± 14.4 | 19.6 ± 6.2 | 20.7 ± 10.2 | 63.8 ± 12.7 | 31.6 ± 16.7 | 45.6 ± 18.3 | <0.001 f | 0.015 # | 0.22 |
| PUFA | 35.3 ± 9.0 | 35.9 ± 11.1 | 18.2 ± 10.1 | 66.3 ± 14.5 | 41.9 ± 17.6 | 49.7 ± 17.0 | <0.001 f | 0.041 * | 0.11 |
| UFA-% | 63% ± 7% | 62% ± 3% | 64% ± 6% | 63% ± 5% | 69% ± 2% | 64% ± 2% | 0.18 | 0.64 | 0.23 |
| SFA-% | 37% ± 7% | 38% ± 3% | 36% ± 6% | 37% ± 5% | 31% ± 2% | 36% ± 2% | 0.18 | 0.64 | 0.23 |
| MUFA-% | 26% ± 6% | 22% ± 6% | 35% ± 17% | 31% ± 1% | 29% ± 3% | 30% ± 3% | 0.48 | 0.20 | 0.30 |
| PUFA-% | 37% ± 12% | 39% ± 4% | 29% ± 17% | 32% ± 5% | 40% ± 4% | 34% ± 2% | 0.91 | 0.20 | 0.58 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramanathan, C.; Johnson, H.; Sharma, S.; Son, W.; Puppa, M.; Rohani, S.N.; Tipirneni-Sajja, A.; Bloomer, R.J.; van der Merwe, M. Early Time-Restricted Feeding Amends Circadian Clock Function and Improves Metabolic Health in Male and Female Nile Grass Rats. Medicines 2022, 9, 15. https://0-doi-org.brum.beds.ac.uk/10.3390/medicines9020015
Ramanathan C, Johnson H, Sharma S, Son W, Puppa M, Rohani SN, Tipirneni-Sajja A, Bloomer RJ, van der Merwe M. Early Time-Restricted Feeding Amends Circadian Clock Function and Improves Metabolic Health in Male and Female Nile Grass Rats. Medicines. 2022; 9(2):15. https://0-doi-org.brum.beds.ac.uk/10.3390/medicines9020015
Chicago/Turabian StyleRamanathan, Chidambaram, Hayden Johnson, Suman Sharma, Wangkuk Son, Melissa Puppa, Saba Neyson Rohani, Aaryani Tipirneni-Sajja, Richard J. Bloomer, and Marie van der Merwe. 2022. "Early Time-Restricted Feeding Amends Circadian Clock Function and Improves Metabolic Health in Male and Female Nile Grass Rats" Medicines 9, no. 2: 15. https://0-doi-org.brum.beds.ac.uk/10.3390/medicines9020015