
Biosynthesis of Poly(3-hydroxybutyrate-co-3-hydroxyhexanoate) from CO2 by a Recombinant Cupriavidusnecator


Abstract
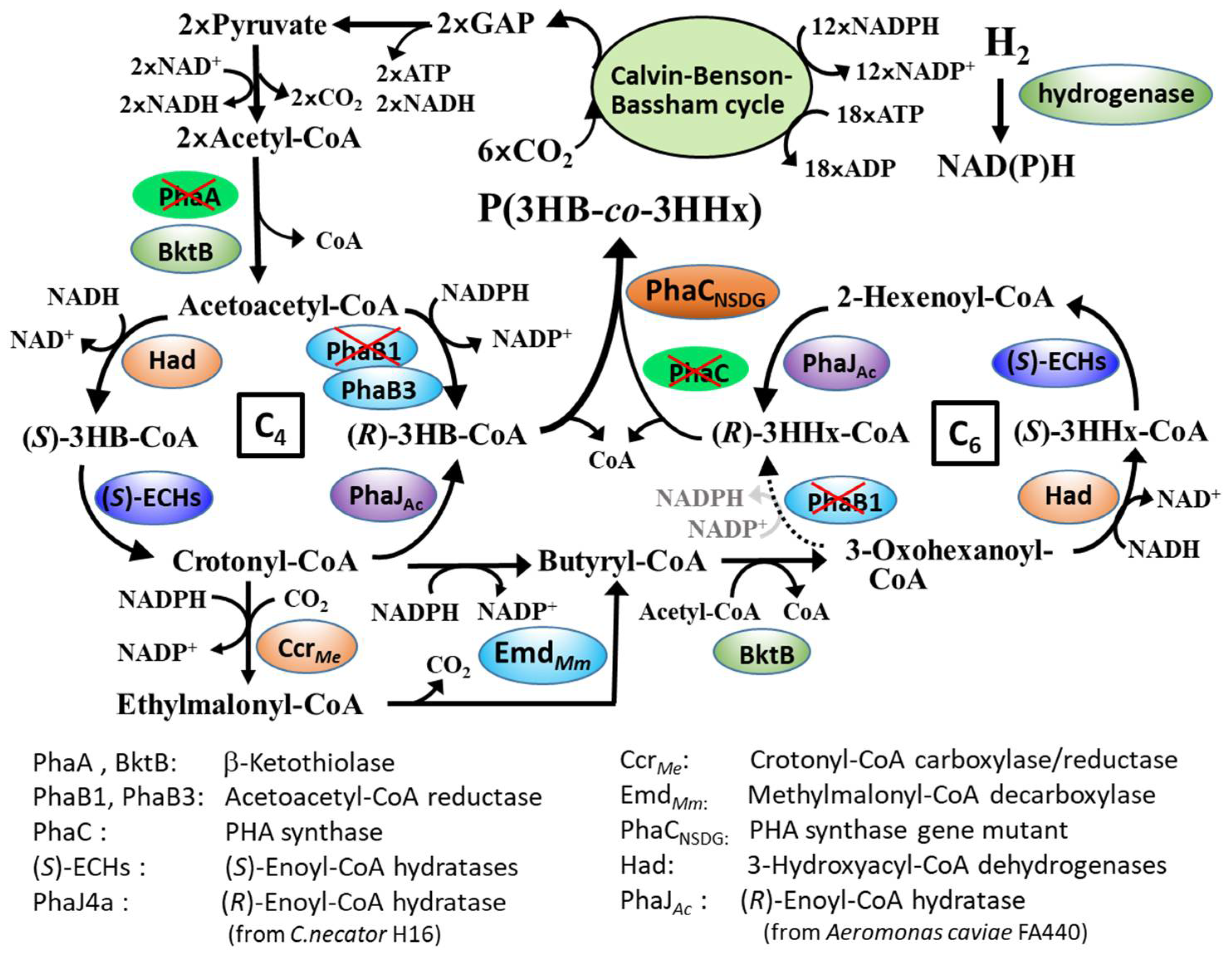
:1. Introduction
2. Materials and Methods
2.1. Construction of Plasmids and Strains
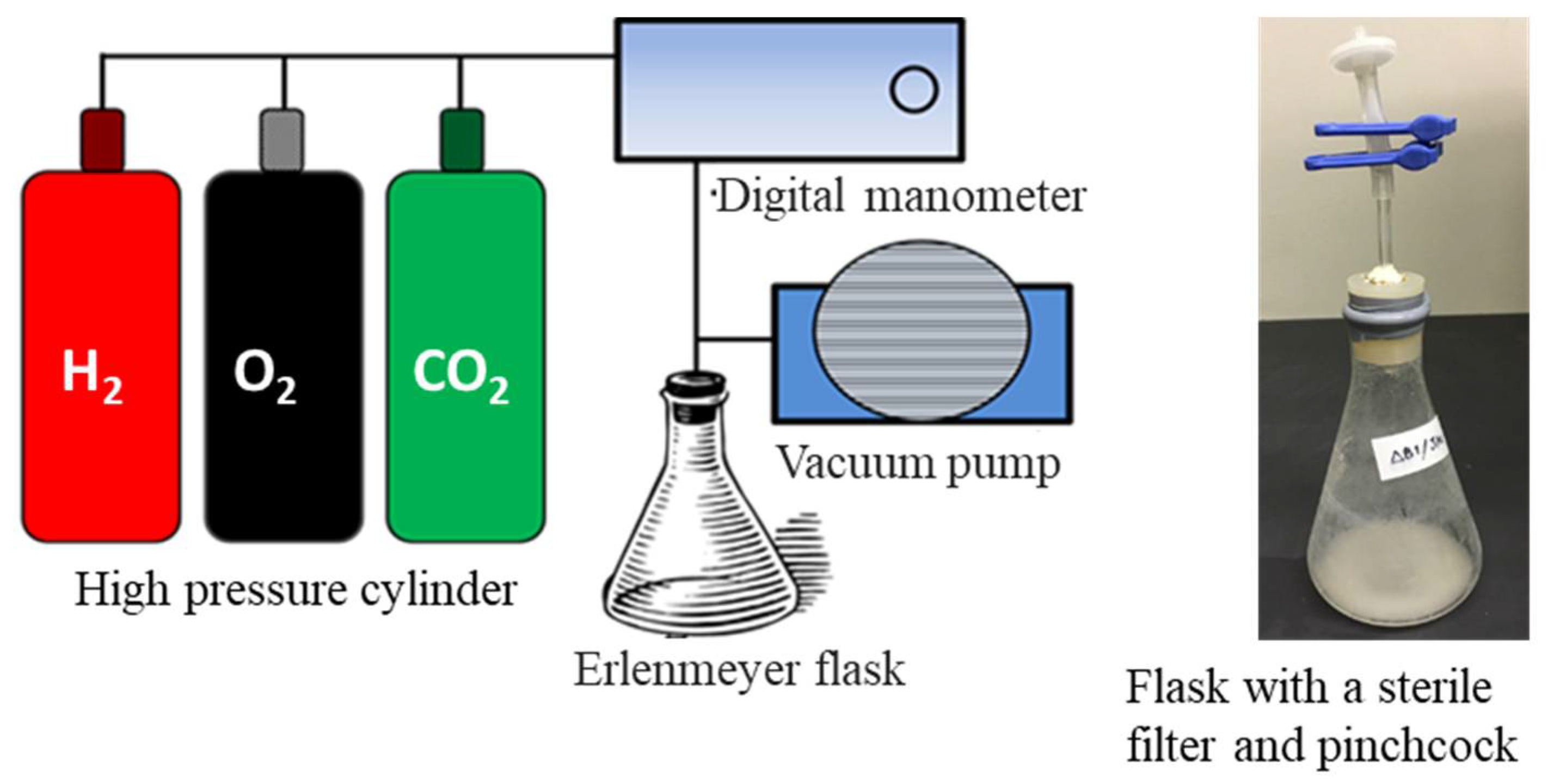
2.2. Culture Medium and Condition
2.3. Analyses
3. Result
3.1. Autotrophic PHA Synthesis by C. necator MF01/pBPP-ccrMeJ4a-emd and MF01ΔB1/pBPP-ccrMeJ4a-emd
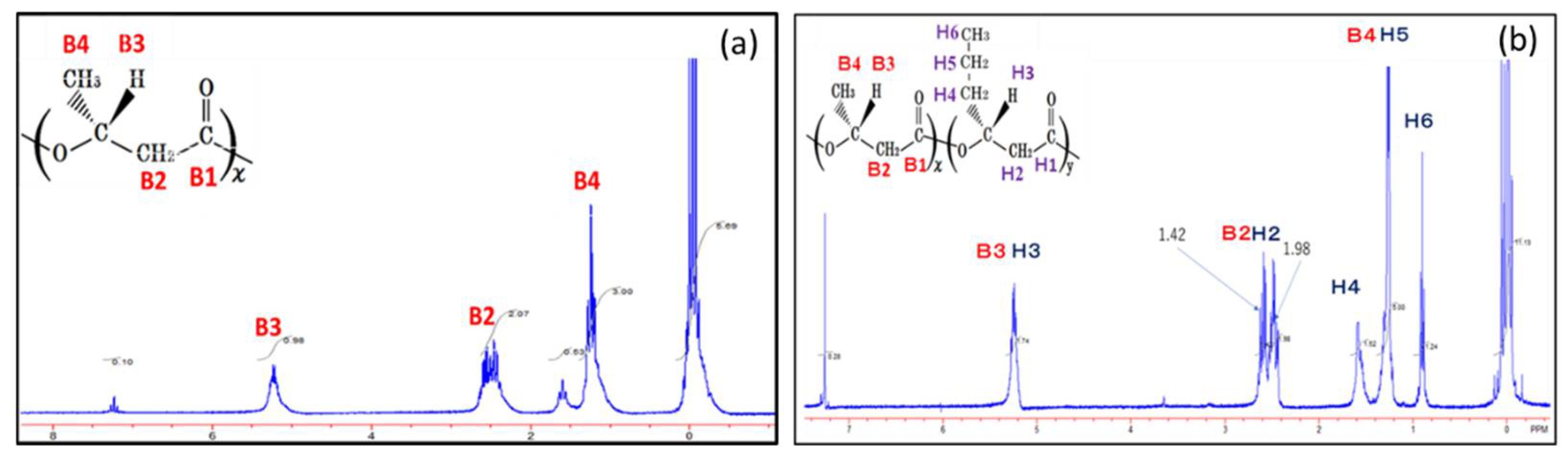
3.2. NMR Analysis of PHBHHx Synthesized by the Autotrophic Condition
3.3. Autotrophic PHA Synthesis by C. necator MF01/pBPP-ccrMeJAc-emd and MF01ΔB1/pBPP-ccrMeJAc-emd
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sen, K.Y.; Baidura, S. Renewable biomass feedstocks for production of sustainable biodegradable polymer. Curr. Opin. Green Sustain. Chem. 2021, 27, 100412. [Google Scholar] [CrossRef]
- Barron, A.; Taylor, D.S. Commercial Marine-Degradable Polymers for Flexible Packaging. Iscience 2020, 23, 101353. [Google Scholar] [CrossRef] [PubMed]
- Hazer, B.; Steinbüchel, A. Increased diversification of polyhydroxyalkanoates by modification reactions for industrial and medical applications. Appl. Microbiol. Biotechnol. 2007, 74, 1–12. Available online: https://0-link-springer-com.brum.beds.ac.uk/article/10.1007%2Fs00253-006-0732-8 (accessed on 12 June 2021). [CrossRef]
- Lu, J.; Tappel, R.C.; Nomura, C.T. Mini-Review: Biosynthesis of Poly(hydroxyalkanoates). Polym. Rev. 2009, 49, 226–248. [Google Scholar] [CrossRef]
- Turco, R.; Santagata, G.; Corrado, I.; Pezzella, C.; Serio, M.D. In Vivo and Post-Synthesis Strategies to Enhance the Properties of PHB-Based Materials: A Review. Front. Bioeng. Biotechnol. 2021, 8, 619266. [Google Scholar] [CrossRef] [PubMed]
- Sashiwa, H.; Fukuda, R.; Okura, T.; Sato, S.; Nakayama, A. Microbial Degradation Behavior in Seawater of Polyester Blends Containing Poly(3-hydroxybutyrate-co-3-hydroxyhexanoate) (PHBHHx). Mar. Drugs 2018, 16, 34. [Google Scholar] [CrossRef] [Green Version]
- Kaneka Corporation News Release. Completion of Kaneka Biodegradable Polymer PHBH™ Plant with Annual Production of 5000 Tons. 19 December 2019. Available online: https://www.kaneka.co.jp/en/topics/news/nr20191219/ (accessed on 20 August 2021).
- Shimamura, E.; Kasuya, K.; Kobayashi, G.; Shiotani, T.; Shima, Y.; Doi, Y. Physical Properties and Biodegradability of Microbial Poly(3-hydroxybutyrate-co-3-hydroxyhexanoate). Macromolecules 1994, 27, 878–880. [Google Scholar] [CrossRef]
- Doi, Y.; Kitamura, S.; Abe, H. Microbial Synthesis and Characteriza-tion of Poly(3-hydroxybutyrate-co-3-hydroxyhexanoate). Macromolecules 1995, 28, 4822–4828. [Google Scholar] [CrossRef]
- Boey, J.Y.; Mohamad, L.; Khok, Y.S.; Tay, G.S.; Baidurah, S. A Review of the Applications and Biodegradation of Polyhydroxyalkanoates and Poly(lactic acid) and Its Composites. Polymers 2021, 13, 1544. [Google Scholar] [CrossRef]
- Insomphun, C.; Xie, H.; Mifune, J.; Kawashima, Y.; Orita, I.; Nakamura, S.; Fukui, T. Improved artificial pathway for biosynthesis of poly(3-hydroxybutyrate-co-3-hydroxyhexanoate) with high C6-monomer composition from fructose in Ralstonia eutropha. Metab. Eng. 2015, 27, 38–45. [Google Scholar] [CrossRef]
- Matassa, S.; Boon, N.; Verstraete, W. Resource recovery from used water: The manufacturing abilities of hydrogen-oxidizing bacteria. Water Res. 2015, 68, 467–478. [Google Scholar] [CrossRef] [PubMed]
- Volova, T.G.; Barashkov, V.A. Characteristics of proteins synthesized by hydrogen-oxidizing microorganisms. Appl. Biochem. Microbiol. 2010, 46, 574–579. Available online: https://www.frontiersin.org/articles/10.3389/fbioe.2020.619266/full (accessed on 12 June 2021). [CrossRef]
- Ishizaki, A.; Tanaka, K. Production of poly-β-hydroxybutyric acid from carbon dioxide by Alcaligenes eutrophus ATCC 17697T. J. Ferment. Bioeng. 1991, 71, 254–257. [Google Scholar] [CrossRef]
- Tanaka, K.; Ishizaki, A.; Kanamaru, T.; Kawano, T. Production of poly(D-3-hydroxybutyrate) from CO2, H2, and CO2 by high cell density autotrophic cultivation of Alcaligenes eutrophus. Biotechnol. Bioeng. 1995, 45, 268–275. Available online: https://0-onlinelibrary-wiley-com.brum.beds.ac.uk/doi/10.1002/bit.260450312 (accessed on 12 June 2021). [CrossRef]
- Taga, N.; Tanaka, K.; Ishizaki, A. Effects of rheological change by addition of carboxymethylcellulose in culture media of an air-lift fermentor on poly-D-3-hydroxybutyric acid productivity in autotrophic culture of hydrogen-oxidizing bacterium, Alcaligenes eutrophus. Biotechnol. Bioeng. 1997, 53, 529–533. [Google Scholar] [CrossRef]
- Sugimoto, T.; Tsuge, T.; Tanaka, K.; Ishizaki, A. Control of acetic acid concentration by pH-stat continuous substrate feeding in heterotrophic culture phase of two-stage cultivation of Alcaligenes eutrophus for production of P(3HB) from CO2, H2 and O2 under non-explosive condition. Biotechnol. Bioeng. 1999, 62, 625–631. Available online: https://0-onlinelibrary-wiley-com.brum.beds.ac.uk/doi/10.1002/(SICI)1097-0290(19990320)62:6%3C625::AID-BIT1%3E3.0.CO;2-D (accessed on 15 June 2021). [CrossRef]
- Ishizaki, A.; Tanaka, K.; Taga, N. Microbial production of poly-D-3-hydroxybutyrate from CO2. Appl. Microbiol. Biotechnol. 2001, 57, 6–12. Available online: https://0-link-springer-com.brum.beds.ac.uk/article/10.1007%2Fs002530100775 (accessed on 15 June 2021). [PubMed]
- Mifune, J.; Nakamura, S.; Fukui, T. Engineering of pha operon on Cupriavidus necator chromosome for efficient biosynthesis of poly(3-hydroxybutyrate-co-3-hydroxyhexanoate) from vegetable oil. Polym. Degrad. Stab. 2010, 95, 1305–1312. [Google Scholar] [CrossRef]
- Zhang, M.; Kurita, S.; Orita, I.; Nakamura, S.; Fukui, T. Modification of acetoacetyl-CoA reduction step in Ralstonia eutropha for biosynthesis of poly(3-hydroxybutyrate-co-3-hydroxyhexanoate) from structurally unrelated compounds. Microb. Cell Fact. 2019, 18, 147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuge, T.; Watanabe, S.; Shimada, D.; Abe, H.; Doi, Y.; Taguchi, S. Combination of N149S and D171G mutations in Aeromonas caviae polyhydroxyalkanoate synthase and impact on polyhydroxyalkanoate biosynthesis. FEMS Microbiol. Lett. 2007, 277, 217–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawashima, Y.; Cheng, W.; Mifune, J.; Orita, I.; Nakamura, S.; Fukui, T. Characterization and functional analyses of R-specific enoyl coenzyme A hydratases in polyhydroxyalkanoate-producing Ralstonia eutropha. Appl. Environ. Microbiol. 2012, 78, 493–502. Available online: http://0-refhub-elsevier-com.brum.beds.ac.uk/S1096-7176(14)00128-1/sbref20 (accessed on 15 June 2021). [CrossRef] [PubMed] [Green Version]
- Fukui, T.; Ohsawa, K.; Mifune, J.; Orita, I.; Nakamura, S. Evaluation of promoters for gene expression in polyhydroxyalkanoate-producing Cupriavidus necator H16. Appl. Microbiol. Biotechnol. 2011, 89, 1527–1536. Available online: http://0-refhub-elsevier-com.brum.beds.ac.uk/S1096-7176(14)00128-1/sbref13 (accessed on 15 June 2021). [CrossRef] [PubMed]
- Kato, M.; Bao, H.J.; Kang, C.-K.; Fukui, T.; Doi, Y. Production of a novel copolyester of 3-hydroxybutyric acid and medium-chain-length 3-hydroxyalkanoic acids by Pseudomonas sp. 61-3 from sugars. Appl. Microbiol. Biotechnol. 1996, 45, 363–370. Available online: https://0-link-springer-com.brum.beds.ac.uk/article/10.1007/s002530050697 (accessed on 10 August 2021). [CrossRef]
- Volova, T.G.; Kiselev, E.G.; Shishatskaya, E.I.; Zhila, E.I.; Boyandin, A.N.; Syrvacheva, D.A.; Vinogradova, O.N.; Kalacheva, G.S.; Vasiliev, A.D.; Peterson, I.V. Cell growth and accumulation of polyhydroxyalkanoates from CO2 and H2 of a hydrogen-oxidizing bacterium, Cupriavidus eutrophus B-10646. Bioresour. Technol. 2013, 146, 215–222. [Google Scholar] [CrossRef]
- Heinrich, D.; Raberg, M.; Steinbüchel, A. Synthesis of poly(3-hydroxybutyrate-co-3-hydroxyvalerate) from unrelated carbon sources in engineered Rhodospirillum rubrum. FEMS Microbiol. Lett. 2015, 362, fnv038. [Google Scholar] [CrossRef] [Green Version]
- Nangle, S.N.; Ziesack, M.; Buckley, S.; Trivedi, D.; Loh, D.M.; Nocera, D.G.; Silvera, P.A. Valorization of CO2 through lithoautotrophic production of sustainable chemicals in Cupriavidus necator. Metab. Eng. 2020, 62, 207–220. [Google Scholar] [CrossRef] [PubMed]
- Miyahara, Y.; Yamamoto, M.; Thorbecke, R.; Mizuno, S.; Tsuge, T. Autotrophic biosynthesis of polyhydroxyalkanoate by Ralstonia eutropha from non-combustible gas mixture with low hydrogen content. Biotechnol. Lett. 2020, 42, 1655–1662. Available online: https://0-link-springer-com.brum.beds.ac.uk/article/10.1007/s10529-020-02876-3 (accessed on 10 August 2021). [CrossRef]
- Tarawat, S.; Incharoensakdi, A.; Monshupanee, T. Cyanobacterial production of poly (3-hydroxybutyrate-co-3-hydroxyvalerate) from carbon dioxide or a single organic substrate: Improved polymer elongation with an extremely high 3-hydroxyvalerate mole proportion. J. Appl. Phycol. 2020, 32, 1095–1102. [Google Scholar] [CrossRef]
- Taepucharoen, K.; Tarawat, S.; Puangcharoen, M.; Incharoensakdi, A.; Monshupanee, T. Production of poly(3-hydroxybutyrate-co-3-hydroxyvalerate) under photoautotrophy and heterotrophy by non-heterocystous N2-fixing cyanobacterium. Bioresour. Technol. 2017, 239, 523–527. [Google Scholar] [CrossRef]
- Bowien, B.; Kusian, B. Genetics and control of CO2 assimilation in the chemoautotroph Ralstonia eutropha. Arch. Microbiol. 2002, 178, 85–93. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Bacterial Strain or Plasmids | Genotype/Characteristic | References or Sources |
|---|---|---|
| C. necator | ||
| H16 | Wild type | DSM 428 |
| MF01 | H16 derivative, ∆phaC::phaCNSDG, ∆phaA::bktB | [19] |
| MF01ΔB1 | MF01 derivative; ΔphaB1 | [11] |
| Plasmids | ||
| pBPP | pBBR1-MCS2 derivative, PphaP1, TrrnB, | [23] |
| pBPP-ccrMeJ4a-emd | pBPP derivative, ccrMe, phaJ4a, emdMm | [11] |
| pBPP-ccrMeJAc-emd | pBPP derivative, ccrMe, phaJAc, emdMm | [20] |
| Recombinant (Strains/Plasmid Vector) | (NH4)2SO4 (g/L) | DCM (g/L) | PHBHHx Content (wt%) | Monomer Composition (mol%) | |
|---|---|---|---|---|---|
| 3HB | 3HHx | ||||
| MF01/pBPP-ccrMeJ4a-emd | 0.5 | 8.52 ± 1.92 | 85.8 ± 13.2 | 96.7 ± 1.4 | 3.3 ± 1.4 |
| MF01/pBPP-ccrMeJ4a-emd | 1.0 | 12.18 ± 0.40 | 64.0 ± 3.4 | 94.8 ± 1.1 | 5.3 ± 1.1 |
| MF01/pBPP-ccrMeJ4a-emd | 2.0 | 8.01 ± 1.65 | 37.4 ± 2.4 | 97.7 ± 0.8 | 2.4 ± 0.8 |
| MF01ΔB1/pBPP-ccrMeJ4a-emd | 0.5 | 4.35 ± 0.78 | 59.0 ± 16.2 | 56.5 ± 1.7 | 43.5 ± 1.7 |
| MF01ΔB1/pBPP-ccrMeJ4a-emd | 1.0 | 10.65 ± 1.35 | 61.7 ± 4.6 | 52.3 ± 6.2 | 47.7 ± 6.2 |
| MF01ΔB1/pBPP-ccrMeJ4a-emd | 2.0 | 6.99 ± 1.14 | 21.1 ± 0.5 | 71.6 ± 1.9 | 28.5 ± 1.9 |
| Recombinant (Strains/Plasmid) | DCM (g/L) | PHBHHx Content (wt%) | 3HHx (mol%) |
|---|---|---|---|
| MF01/pBPP-ccrMeJ4a-emd a | 1.76 ± 0.02 | 48.5 ± 0.4 | 6.4 ± 0.13 |
| MF01/pBPP-ccrMeJAc-emd | 1.81 ± 0.03 | 54.0 ± 3.1 | 10.4 ± 0.2 |
| MF01ΔB1/pBPP-ccrMeJ4a-emd a | 1.57 ± 0.02 | 47.9 ± 2.0 | 22.2 ± 1.2 |
| MF01ΔB1/pBPP-ccrMeJAc-emd | 1.72 ± 0.01 | 54.1 ± 2.1 | 14.0 ± 0.6 |
| Recombinant (Strains/Plasmid Vector) | (NH4)2 SO4 (g/L) | DCM (g/L) | PHBHHx Content (wt%) | Monomer (mol%) | |
|---|---|---|---|---|---|
| 3HB | 3HHx | ||||
| MF01/pBPP-ccrMeJAc-emd | 0.5 | 7.25 ± 0.57 | 76.2 ± 0.0 | 94.0 ± 1.3 | 6.0 ± 1.3 |
| MF01/pBPP-ccrMeJAc-emd | 1.0 | 11.22 ± 2.67 | 64.6 ± 8.1 | 88.7 ± 6.4 | 11.3 ± 6.4 |
| MF01/pBPP-ccrMeJAc-emd | 2.0 | 8.46 ± 0.42 | 19.9 ± 1.7 | 88.6 ± 0.9 | 11.5 ± 0.9 |
| MF01ΔB1/pBPP-ccrMeJAc-emd | 0.5 | 6.93 ± 0.36 | 74.6 ± 2.2 | 86.6 ± 1.0 | 14.0 ± 1.3 |
| MF01ΔB1/pBPP-ccrMeJAc-emd | 1.0 | 8.52 ± 1.00 | 67.8 ± 1.8 | 87.1 ± 2.3 | 11.1 ± 1.3 |
| MF01ΔB1/pBPP-ccrMeJAc-emd | 2.0 | 6.03 ± 0.78 | 39.1 ± 1.3 | 90.4 ± 0.7 | 9.6 ± 0.7 |
| Scheme. | Nutritional Condition | PHA Production | PHA Composition (mol%) | Ref. |
|---|---|---|---|---|
| C. necator H 16 a | H2/CO2/air = 30:15:balance | - | Various type of PHAs with C4-C14 monomers | [27] |
| C. necator H 16 b | H2/O2/CO2/N2 = 3.6: 7.6: 12.3: 76.5 (non-combustible gas) | DCM0.14 ± 0.05 g/L (PHA content 57 ± 10 wt%) | 3HB-based copolymer with 1.2 % 3HV c and 1.2% 3H4MV d | [28] |
| Anabaena spiroides TISTR 8075 | CO2 + light | PHBV productivity 0.5 mg gdw−1day−1 | PHBV (3HV 43.2%) | [29] |
| Oscillatoria okeni TISTR 8549 | CO2 + light | PHBV concentration 108 mg/L | PHBV (3HV 5.5%) | [30] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tanaka, K.; Yoshida, K.; Orita, I.; Fukui, T. Biosynthesis of Poly(3-hydroxybutyrate-co-3-hydroxyhexanoate) from CO2 by a Recombinant Cupriavidusnecator. Bioengineering 2021, 8, 179. https://0-doi-org.brum.beds.ac.uk/10.3390/bioengineering8110179
Tanaka K, Yoshida K, Orita I, Fukui T. Biosynthesis of Poly(3-hydroxybutyrate-co-3-hydroxyhexanoate) from CO2 by a Recombinant Cupriavidusnecator. Bioengineering. 2021; 8(11):179. https://0-doi-org.brum.beds.ac.uk/10.3390/bioengineering8110179
Chicago/Turabian StyleTanaka, Kenji, Kazumasa Yoshida, Izumi Orita, and Toshiaki Fukui. 2021. "Biosynthesis of Poly(3-hydroxybutyrate-co-3-hydroxyhexanoate) from CO2 by a Recombinant Cupriavidusnecator" Bioengineering 8, no. 11: 179. https://0-doi-org.brum.beds.ac.uk/10.3390/bioengineering8110179