
Human iPSC-Derived Vascular Smooth Muscle Cells in a Fibronectin Functionalized Collagen Hydrogel Augment Endothelial Cell Morphogenesis

 , ,
, ,
Abstract
:1. Introduction
2. Methods
2.1. Cell Culture
2.2. HUVEC Culture
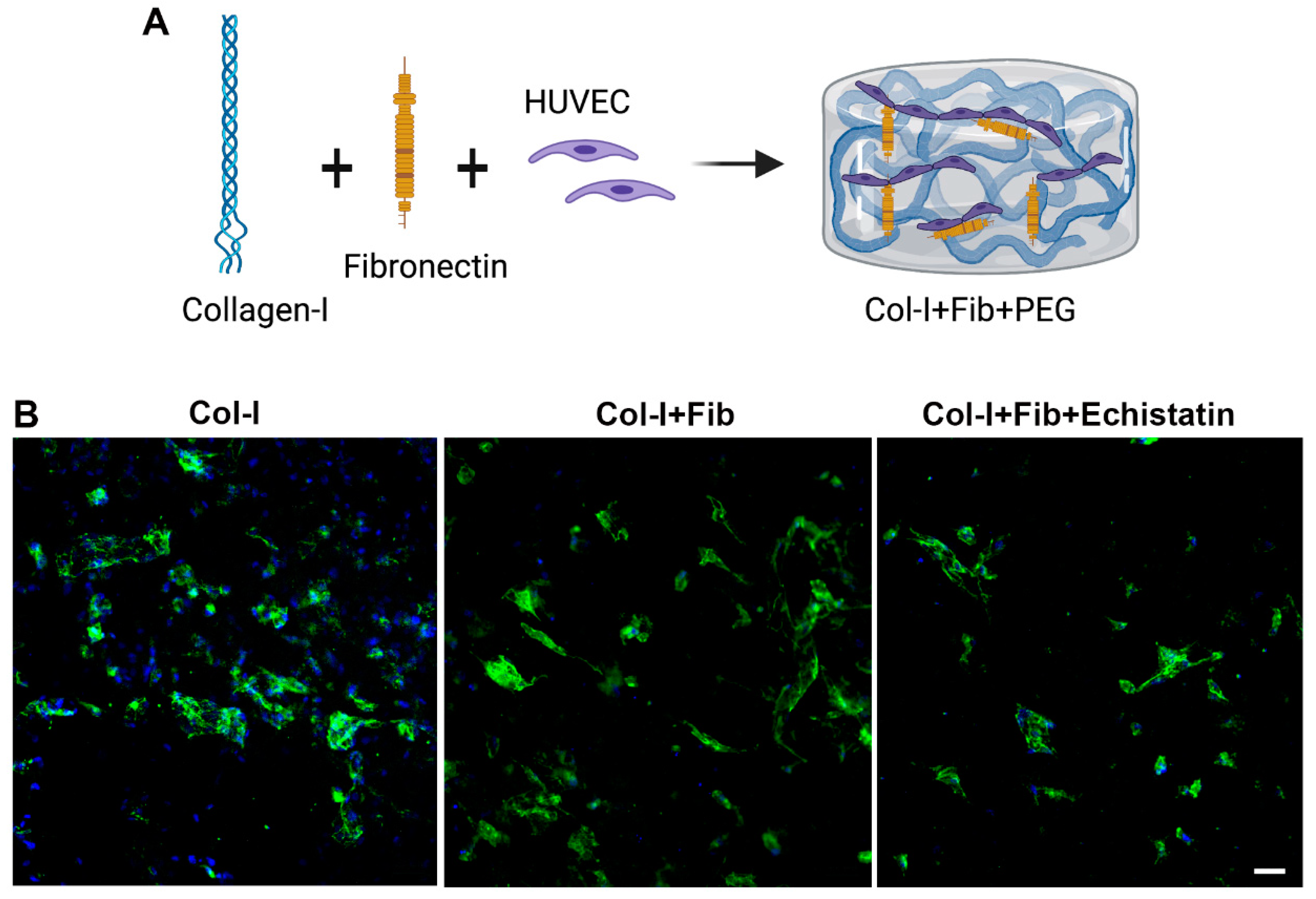
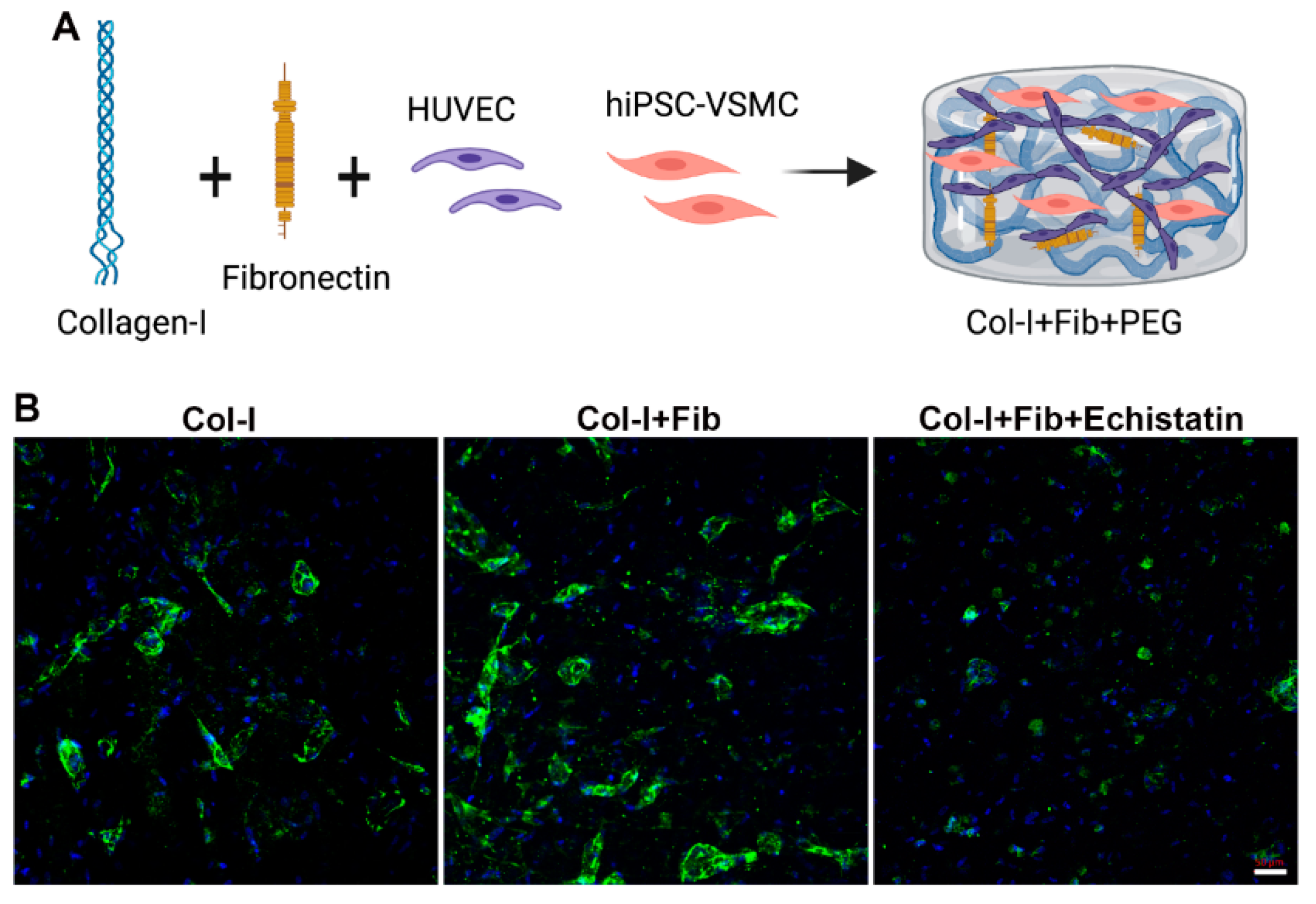
2.3. Fibronectin Collagen Hydrogel Production
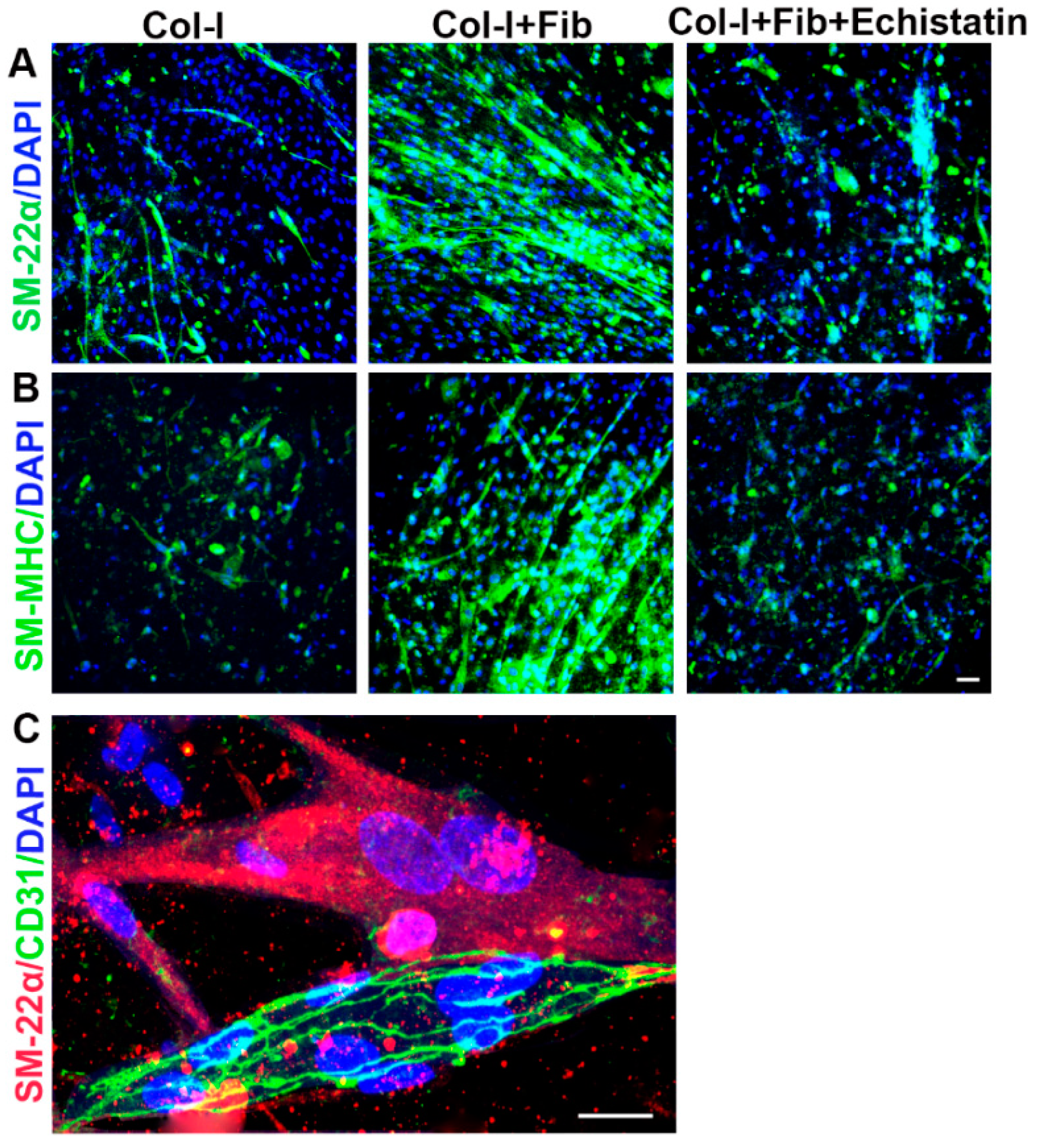
2.4. αvβ3 Integrin Signaling Inhibition Experiment
2.5. Immunofluorescence Staining
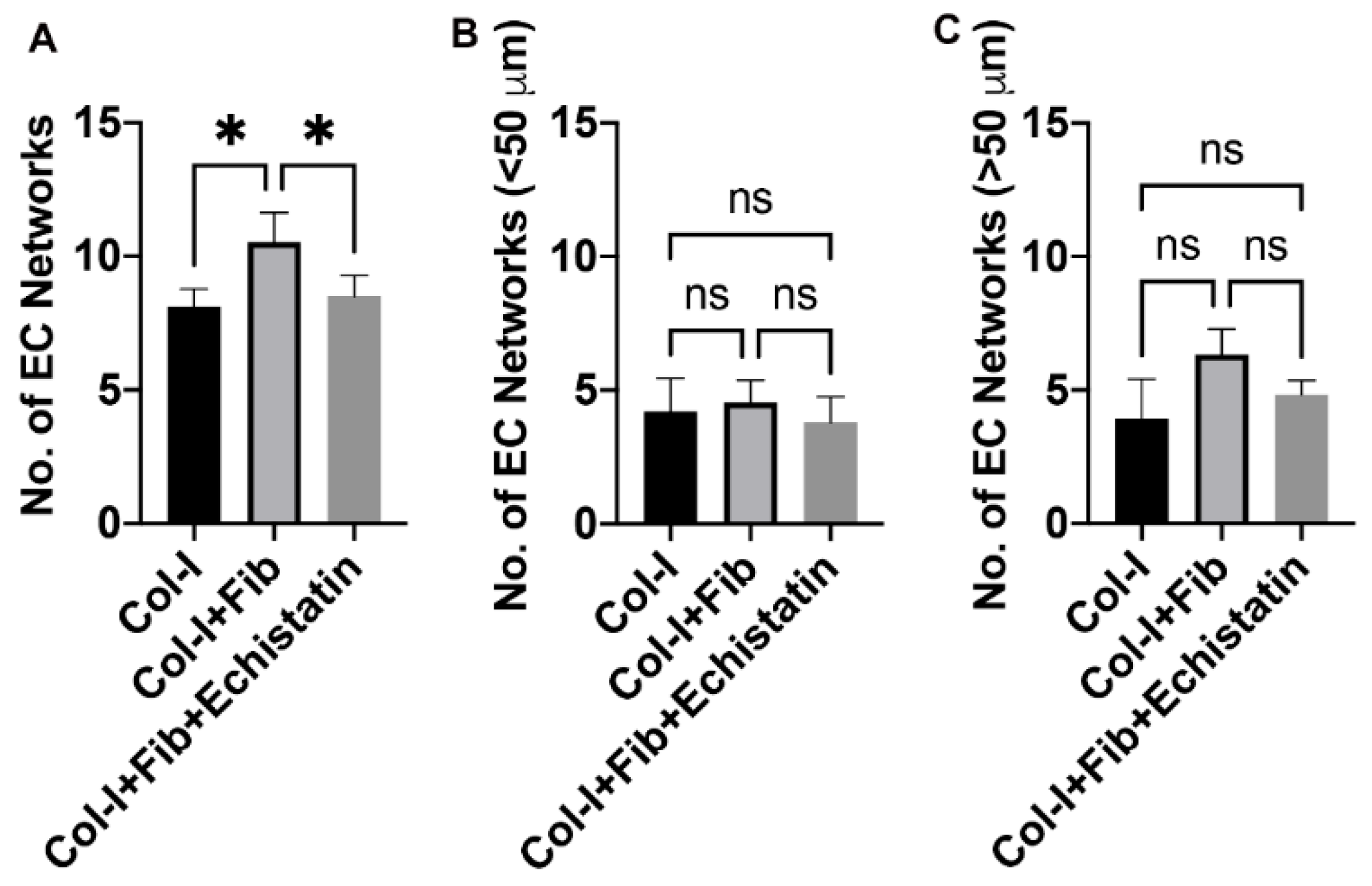
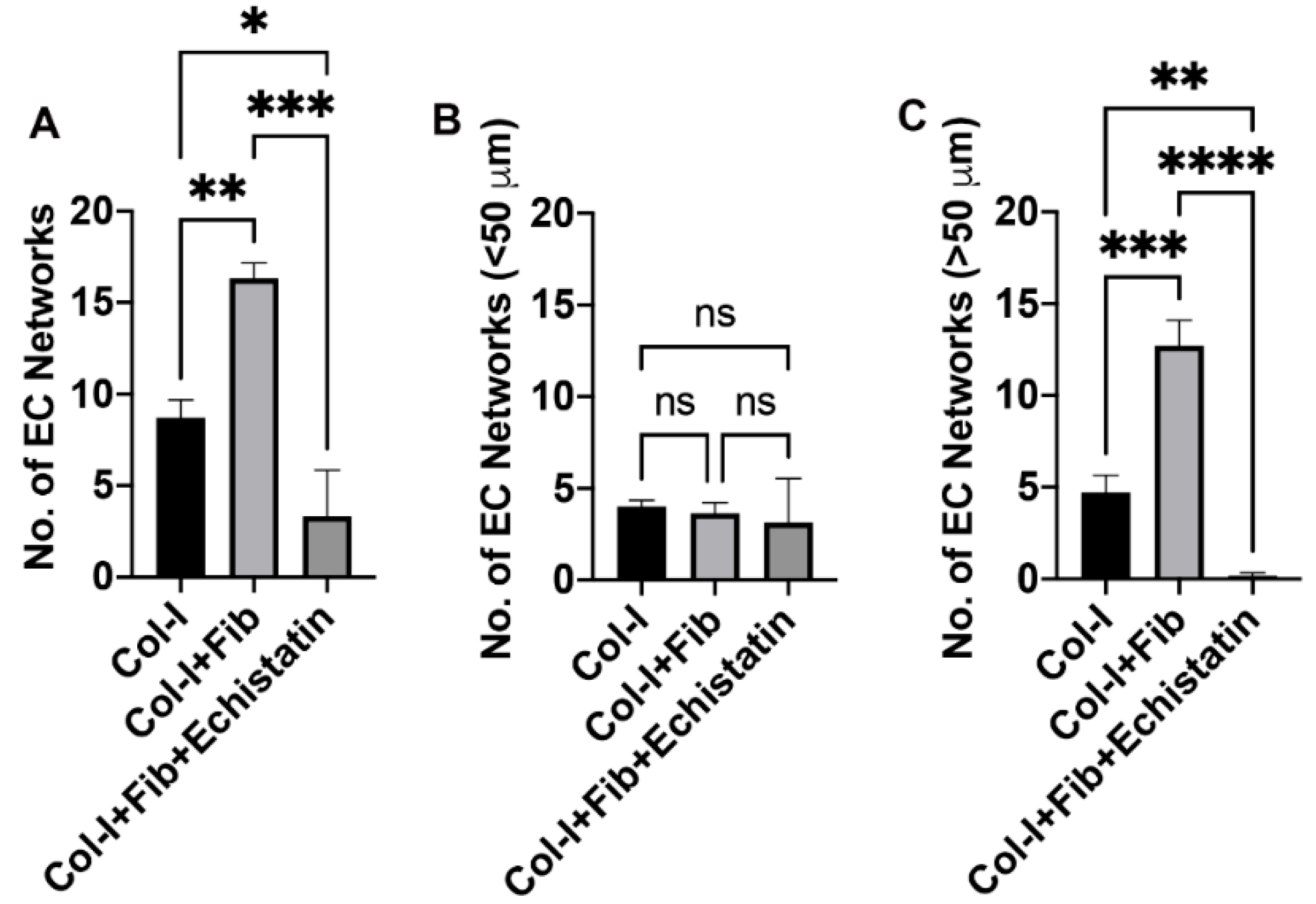
2.6. EC Network Quantification
2.7. AlamarBlue Assay
2.8. ELISA
3. Results
3.1. Fibronectin Increased the Number of EC Networks
3.2. hiPSC-VSMC in the Presence of Fibronectin Enhanced the Number and Size of EC Networks
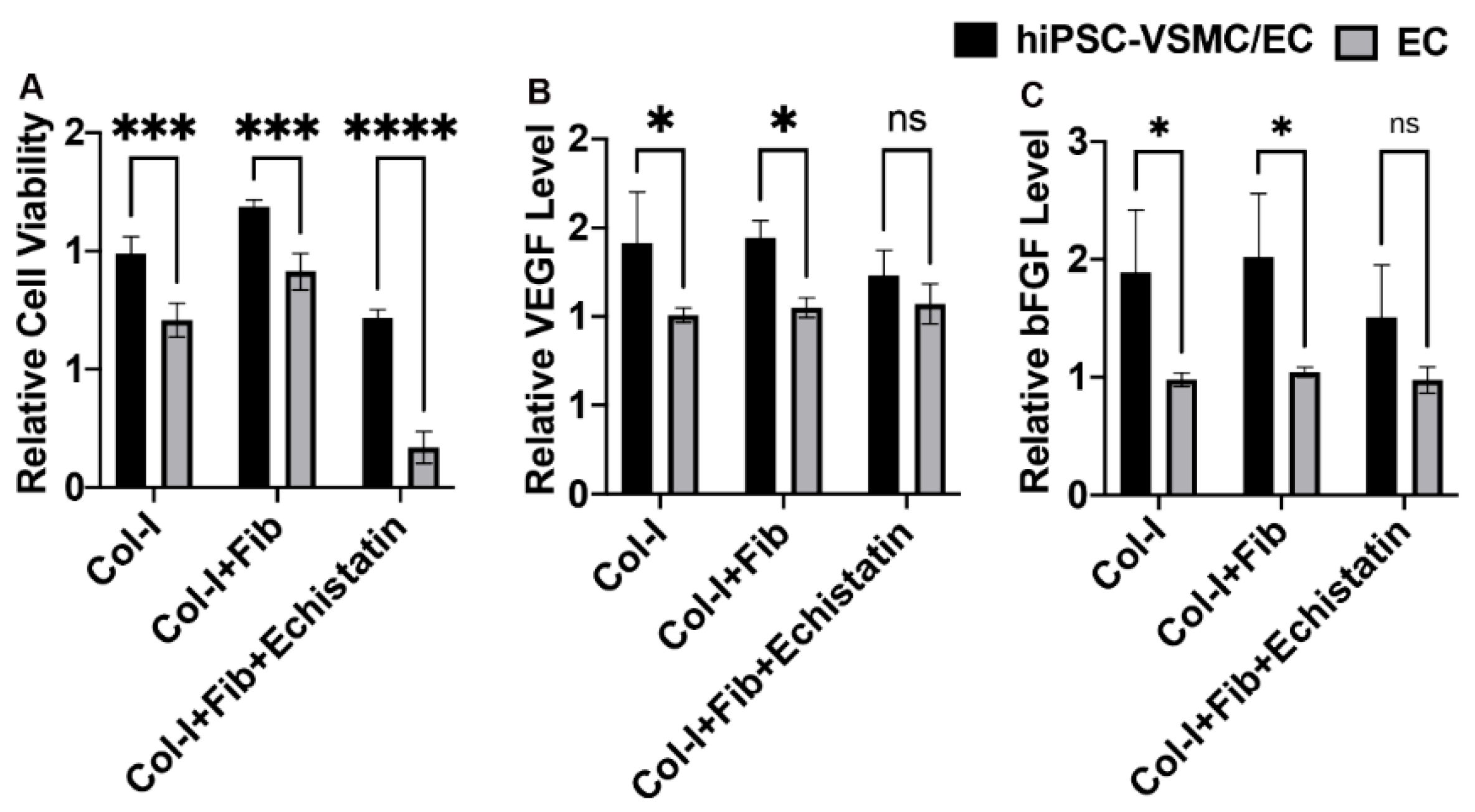
3.3. hiPSC-VSMC Irrespective of Fibronectin Enhanced the Viability and Proangiogenic Growth Factor Secretion
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dickinson, L.E.; Gerecht, S. Engineered Biopolymeric Scaffolds for Chronic Wound Healing. Front. Physiol. 2016, 7, 341. [Google Scholar] [CrossRef] [PubMed]
- Ho, J.; Walsh, C.; Yue, D.; Dardik, A.; Cheema, U. Current Advancements and Strategies in Tissue Engineering for Wound Healing: A Comprehensive Review. Adv. Wound Care 2017, 6, 191–209. [Google Scholar] [CrossRef] [PubMed]
- Bagheri, M.; Validi, M.; Gholipour, A.; Makvandi, P.; Sharifi, E. Chitosan nanofiber biocomposites for potential wound healing applications: Antioxidant activity with synergic antibacterial effect. Bioeng. Transl. Med. 2021, e10254. [Google Scholar] [CrossRef]
- Sharifi, E.; Chehelgerdi, M.; Fatahian-Kelishadrokhi, A.; Yazdani-Nafchi, F.; Ashrafi-Dehkordi, K. Comparison of therapeutic effects of encapsulated Mesenchymal stem cells in Aloe vera gel and Chitosan-based gel in healing of grade-II burn injuries. Regen. Ther. 2021, 18, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Dash, B.C.; Xu, Z.; Lin, L.; Koo, A.; Ndon, S.; Berthiaume, F.; Dardik, A.; Hsia, H. Stem Cells and Engineered Scaffolds for Regenerative Wound Healing. Bioengineering 2018, 5, 23. [Google Scholar] [CrossRef]
- Vijayan, A.; Sabareeswaran, A.; Kumar, G.S.V. PEG grafted chitosan scaffold for dual growth factor delivery for enhanced wound healing. Sci. Rep. 2019, 9, 19165. [Google Scholar] [CrossRef]
- Tra Thanh, N.; Ho Hieu, M.; Tran Minh Phuong, N.; Do Bui Thuan, T.; Nguyen Thi Thu, H.; Thai, V.P.; Do Minh, T.; Nguyen Dai, H.; Vo, V.T.; Nguyen Thi, H. Optimization and characterization of electrospun polycaprolactone coated with gelatin-silver nanoparticles for wound healing application. Mater. Sci. Eng. C Mater. Biol. Appl. 2018, 91, 318–329. [Google Scholar] [CrossRef] [PubMed]
- Shepherd, B.R.; Enis, D.R.; Wang, F.; Suarez, Y.; Pober, J.S.; Schechner, J.S. Vascularization and engraftment of a human skin substitute using circulating progenitor cell-derived endothelial cells. FASEB J 2006, 20, 1739–1741. [Google Scholar] [CrossRef]
- Yao, T.; Wieringa, P.A.; Chen, H.; Amit, C.; Samal, P.; Giselbrecht, S.; Baker, M.B.; Moroni, L. Fabrication of a self-assembled honeycomb nanofibrous scaffold to guide endothelial morphogenesis. Biofabrication 2020, 12, 045001. [Google Scholar] [CrossRef]
- Zhang, S.; Wan, Z.; Kamm, R.D. Vascularized organoids on a chip: Strategies for engineering organoids with functional vasculature. Lab Chip 2021, 21, 473–488. [Google Scholar] [CrossRef] [PubMed]
- Simon, M.C. Vascular morphogenesis and the formation of vascular networks. Dev. Cell 2004, 6, 479–482. [Google Scholar] [CrossRef]
- Tsuji-Tamura, K.; Ogawa, M. Morphology regulation in vascular endothelial cells. Inflamm. Regen. 2018, 38, 25. [Google Scholar] [CrossRef] [PubMed]
- Hielscher, A.; Ellis, K.; Qiu, C.; Porterfield, J.; Gerecht, S. Fibronectin Deposition Participates in Extracellular Matrix Assembly and Vascular Morphogenesis. PLoS ONE 2016, 11, e0147600. [Google Scholar] [CrossRef] [PubMed]
- Kusuma, S.; Macklin, B.; Gerecht, S. Derivation and network formation of vascular cells from human pluripotent stem cells. Methods Mol. Biol. 2014, 1202, 1–9. [Google Scholar] [CrossRef]
- Ren, J.; Zhou, T.; Pilli, V.S.S.; Phan, N.; Wang, Q.; Gupta, K.; Liu, Z.; Sheibani, N.; Liu, B. Novel Paracrine Functions of Smooth Muscle Cells in Supporting Endothelial Regeneration Following Arterial Injury. Circ. Res. 2019, 124, 1253–1265. [Google Scholar] [CrossRef]
- Bargehr, J.; Low, L.; Cheung, C.; Bernard, W.G.; Iyer, D.; Bennett, M.R.; Gambardella, L.; Sinha, S. Embryological Origin of Human Smooth Muscle Cells Influences Their Ability to Support Endothelial Network Formation. Stem Cells Transl. Med. 2016, 5, 946–959. [Google Scholar] [CrossRef]
- Dash, B.C.; Setia, O.; Gorecka, J.; Peyvandi, H.; Duan, K.; Lopes, L.; Nie, J.; Berthiaume, F.; Dardik, A.; Hsia, H.C. A Dense Fibrillar Collagen Scaffold Differentially Modulates Secretory Function of iPSC-Derived Vascular Smooth Muscle Cells to Promote Wound Healing. Cells 2020, 9, 966. [Google Scholar] [CrossRef] [PubMed]
- Drews, K.; Jozefczuk, J.; Prigione, A.; Adjaye, J. Human induced pluripotent stem cells—from mechanisms to clinical applications. J. Mol. Med. 2012, 90, 735–745. [Google Scholar] [CrossRef] [PubMed]
- Atchison, L.; Abutaleb, N.O.; Snyder-Mounts, E.; Gete, Y.; Ladha, A.; Ribar, T.; Cao, K.; Truskey, G.A. iPSC-Derived Endothelial Cells Affect Vascular Function in a Tissue-Engineered Blood Vessel Model of Hutchinson-Gilford Progeria Syndrome. Stem Cell Rep. 2020, 14, 325–337. [Google Scholar] [CrossRef] [PubMed]
- Atchison, L.; Zhang, H.; Cao, K.; Truskey, G.A. A Tissue Engineered Blood Vessel Model of Hutchinson-Gilford Progeria Syndrome Using Human iPSC-derived Smooth Muscle Cells. Sci. Rep. 2017, 7, 8168. [Google Scholar] [CrossRef] [PubMed]
- Dash, B.C.; Levi, K.; Schwan, J.; Luo, J.; Bartulos, O.; Wu, H.; Qiu, C.; Yi, T.; Ren, Y.; Campbell, S.; et al. Tissue-Engineered Vascular Rings from Human iPSC-Derived Smooth Muscle Cells. Stem Cell Rep. 2016, 7, 19–28. [Google Scholar] [CrossRef]
- Dash, B.C.; Hsia, H.C. Generation and Encapsulation of Human iPSC-Derived Vascular Smooth Muscle Cells for Proangiogenic Therapy. Methods Mol. Biol. 2021, 1–11. [Google Scholar] [CrossRef]
- Gorecka, J.; Gao, X.; Fereydooni, A.; Dash, B.C.; Luo, J.; Lee, S.R.; Taniguchi, R.; Hsia, H.C.; Qyang, Y.; Dardik, A. Induced pluripotent stem cell-derived smooth muscle cells increase angiogenesis and accelerate diabetic wound healing. Regen. Med. 2020, 15, 1277–1293. [Google Scholar] [CrossRef]
- Ahmed, M.; Ffrench-Constant, C. Extracellular Matrix Regulation of Stem Cell Behavior. Curr. Stem Cell Rep. 2016, 2, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Burdick, J.A.; Prestwich, G.D. Hyaluronic acid hydrogels for biomedical applications. Adv. Mater. 2011, 23, H41–H56. [Google Scholar] [CrossRef] [PubMed]
- Park, K.M.; Gerecht, S. Hypoxia-inducible hydrogels. Nat. Commun. 2014, 5, 4075. [Google Scholar] [CrossRef]
- Ceccarelli, J.; Putnam, A.J. Sculpting the blank slate: How fibrin’s support of vascularization can inspire biomaterial design. Acta Biomater. 2014, 10, 1515–1523. [Google Scholar] [CrossRef] [PubMed]
- Dash, B.C.; Duan, K.; Xing, H.; Kyriakides, T.R.; Hsia, H.C. An in situ collagen-HA hydrogel system promotes survival and preserves the proangiogenic secretion of hiPSC-derived vascular smooth muscle cells. Biotechnol. Bioeng. 2020, 117, 3912–3923. [Google Scholar] [CrossRef] [PubMed]
- Grinnell, F. Fibronectin and wound healing. J. Cell. Biochem. 1984, 26, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Nakakura, T.; Suzuki, T.; Tanaka, H.; Arisawa, K.; Miyashita, T.; Nekooki-Machida, Y.; Kurosawa, T.; Tega, Y.; Deguchi, Y.; Hagiwara, H. Fibronectin is essential for formation of fenestrae in endothelial cells of the fenestrated capillary. Cell Tissue Res. 2021, 383, 823–833. [Google Scholar] [CrossRef] [PubMed]
- Dash, B.C.; Duan, K.; Kyriakides, T.R.; Hsia, H.C. Integrin β3 Targeting Biomaterial Preferentially Promotes Secretion of bFGF and Proliferation of iPSC-Derived Vascular Smooth Muscle Cells. Biomater. Sci. 2021, 9, 5319–5329. [Google Scholar] [CrossRef] [PubMed]
- Dai, W.M.; Lai, K.W.; Wu, A.; Hamaguchi, W.; Lee, M.Y.; Zhou, L.; Ishii, A.; Nishimoto, S. DNA cleavage potency, cytotoxicity, and mechanism of action of a novel class of enediyne prodrugs. J. Med. Chem. 2002, 45, 758–761. [Google Scholar] [CrossRef]
- Lee, C.S.; Kim, S.; Fan, J.; Hwang, H.S.; Aghaloo, T.; Lee, M. Smoothened agonist sterosome immobilized hybrid scaffold for bone regeneration. Sci. Adv. 2020, 6, eaaz7822. [Google Scholar] [CrossRef] [PubMed]
- Pourshahrestani, S.; Zeimaran, E.; Kadri, N.A.; Gargiulo, N.; Jindal, H.M.; Naveen, S.V.; Sekaran, S.D.; Kamarul, T.; Towler, M.R. Potency and Cytotoxicity of a Novel Gallium-Containing Mesoporous Bioactive Glass/Chitosan Composite Scaffold as Hemostatic Agents. ACS Appl. Mater. Interfaces 2017, 9, 31381–31392. [Google Scholar] [CrossRef]
- Wani, M.Y.; Bhat, A.R.; Azam, A.; Choi, I.; Athar, F. Probing the antiamoebic and cytotoxicity potency of novel tetrazole and triazine derivatives. Eur. J. Med. Chem. 2012, 48, 313–320. [Google Scholar] [CrossRef]
- Xing, Q.; Parvizi, M.; Lopera Higuita, M.; Griffiths, L.G. Basement membrane proteins modulate cell migration on bovine pericardium extracellular matrix scaffold. Sci. Rep. 2021, 11, 4607. [Google Scholar] [CrossRef] [PubMed]
- Caldwell, A.S.; Rao, V.V.; Golden, A.C.; Bell, D.J.; Grim, J.C.; Anseth, K.S. Mesenchymal stem cell-inspired microgel scaffolds to control macrophage polarization. Bioeng. Transl. Med. 2021, 6, e10217. [Google Scholar] [CrossRef]
- Chen, W.; Sun, Y.; Gu, X.; Cai, J.; Liu, X.; Zhang, X.; Chen, J.; Hao, Y.; Chen, S. Conditioned medium of human bone marrow-derived stem cells promotes tendon-bone healing of the rotator cuff in a rat model. Biomaterials 2021, 271, 120714. [Google Scholar] [CrossRef]
- Thomas, D.; Fontana, G.; Chen, X.; Sanz-Nogues, C.; Zeugolis, D.I.; Dockery, P.; O’Brien, T.; Pandit, A. A shape-controlled tuneable microgel platform to modulate angiogenic paracrine responses in stem cells. Biomaterials 2014, 35, 8757–8766. [Google Scholar] [CrossRef]
- Wan, X.; Xie, M.K.; Xu, H.; Wei, Z.W.; Yao, H.J.; Wang, Z.; Zheng, D.C. Hypoxia-preconditioned adipose-derived stem cells combined with scaffold promote urethral reconstruction by upregulation of angiogenesis and glycolysis. Stem Cell Res. Ther. 2020, 11, 535. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Density (mg/mL) | Collagen (µL) | 10× MEM (µL) | 10× PBS (µL) | 1 M NaOH (µL) | Fibronectin 1 mg/mL (µL) | hiPSC VSMC 8 × 103/µL | HUVEC 16 × 103/µL |
|---|---|---|---|---|---|---|---|---|
| Col-I+EC | 4 | 400 | 50 | 50 | 8.4 | 0 | 0 | 62.5 |
| Col-I+Fib+EC | 4 | 400 | 50 | 0 | 8.9 | 50 | 0 | 62.5 |
| Col-I+iPSC-VSMC+EC | 4 | 400 | 50 | 50 | 8.4 | 0 | 25 | 50 |
| Col-I+Fib+iPSC-VSMC+EC | 4 | 400 | 50 | 0 | 8.9 | 50 | 25 | 50 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duan, K.; Dash, B.C.; Sasson, D.C.; Islam, S.; Parker, J.; Hsia, H.C. Human iPSC-Derived Vascular Smooth Muscle Cells in a Fibronectin Functionalized Collagen Hydrogel Augment Endothelial Cell Morphogenesis. Bioengineering 2021, 8, 223. https://0-doi-org.brum.beds.ac.uk/10.3390/bioengineering8120223
Duan K, Dash BC, Sasson DC, Islam S, Parker J, Hsia HC. Human iPSC-Derived Vascular Smooth Muscle Cells in a Fibronectin Functionalized Collagen Hydrogel Augment Endothelial Cell Morphogenesis. Bioengineering. 2021; 8(12):223. https://0-doi-org.brum.beds.ac.uk/10.3390/bioengineering8120223
Chicago/Turabian StyleDuan, Kaiti, Biraja C. Dash, Daniel C. Sasson, Sara Islam, Jackson Parker, and Henry C. Hsia. 2021. "Human iPSC-Derived Vascular Smooth Muscle Cells in a Fibronectin Functionalized Collagen Hydrogel Augment Endothelial Cell Morphogenesis" Bioengineering 8, no. 12: 223. https://0-doi-org.brum.beds.ac.uk/10.3390/bioengineering8120223