
Extracellular Vesicles Derived from Primary Adipose Stromal Cells Induce Elastin and Collagen Deposition by Smooth Muscle Cells within 3D Fibrin Gel Culture

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. ASC Culture Conditions and Conditioned Media Collection
2.2. EV Isolation
2.3. EV Characterization
2.3.1. Transmission Electron Microscopy
2.3.2. Dynamic Light Scattering
2.3.3. Total Protein Concentration
2.4. SMC Cell Culture Conditions
2.5. Proliferation Assays
2.6. In Vitro Wound Closure Assays
2.7. 3D SMC-Fibrin Gel Construct Fabrication
2.8. Media Treatment Conditions for Fibrin Gel Constructs
2.9. qPCR Analysis
2.10. Ninhydrin (Insoluble Elastin) and Hydroxyproline (Collagen) Assays
2.11. Tensile Testing of Soft Substrate Fibrin Gel Constructs
2.12. Statistical Analysis
3. Results
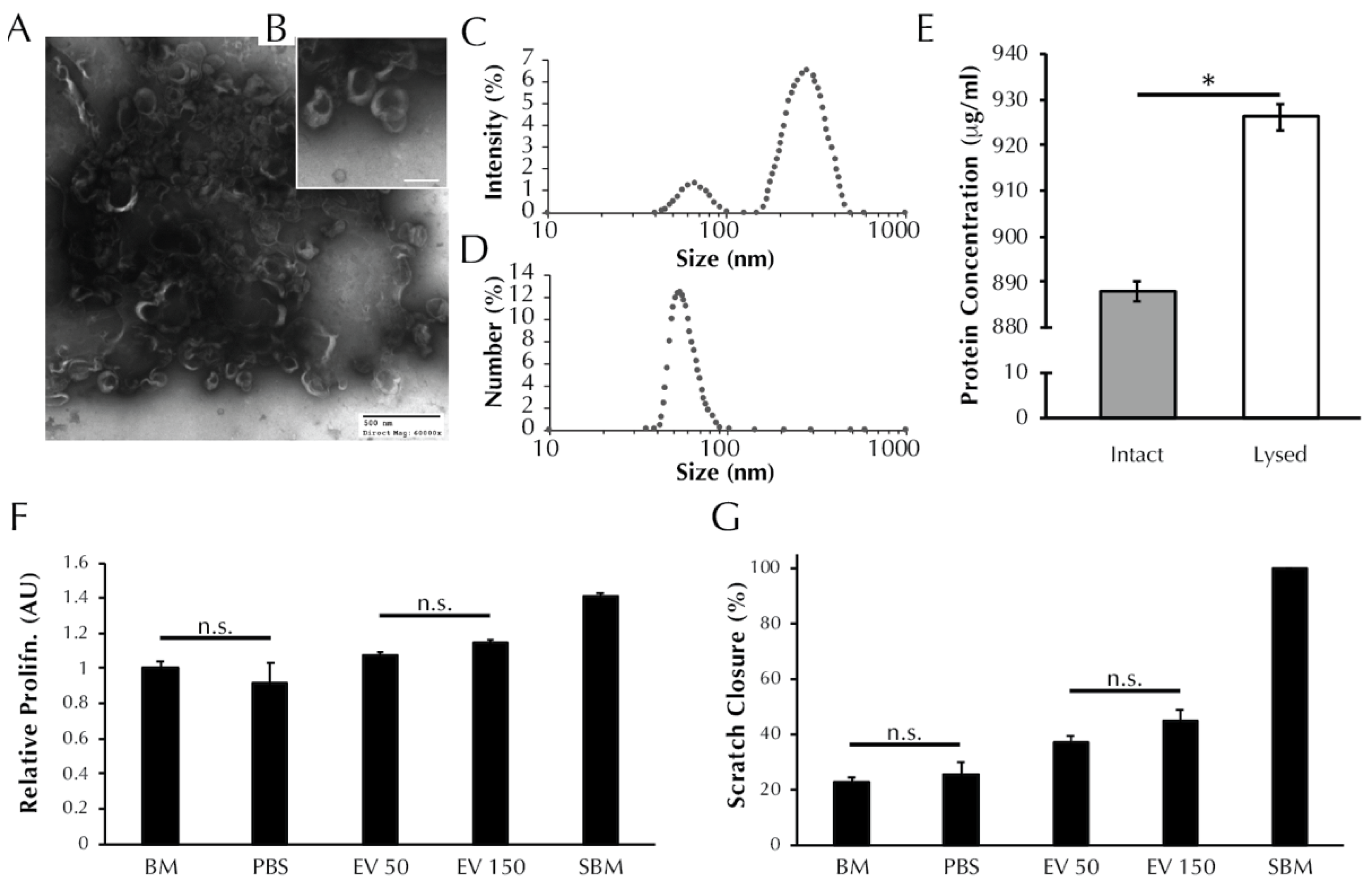
3.1. EVs Secreted from ASCs Are Characteristic of Exosomes and Microvesicles
3.2. ASC-Derived EVs Promote SMC Proliferation
3.3. ASC-Derived EVs Promote SMC Migration in an In Vitro Scratch Wound Assay
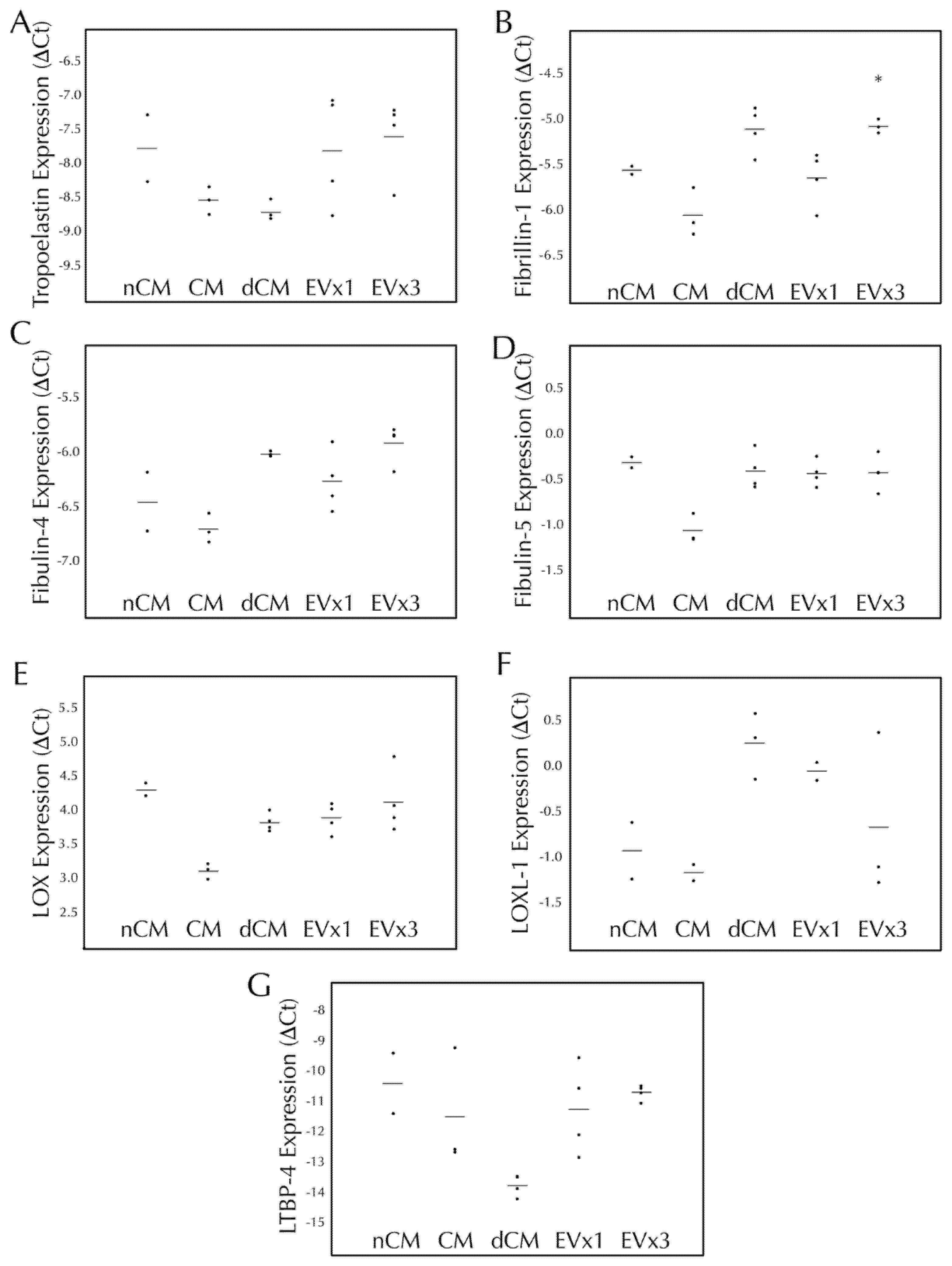
3.4. ASC-Derived EVs Had a Minimal Effect on Elastic Fiber Gene Transcription within SMC Disc Constructs after 30 Days of Culture
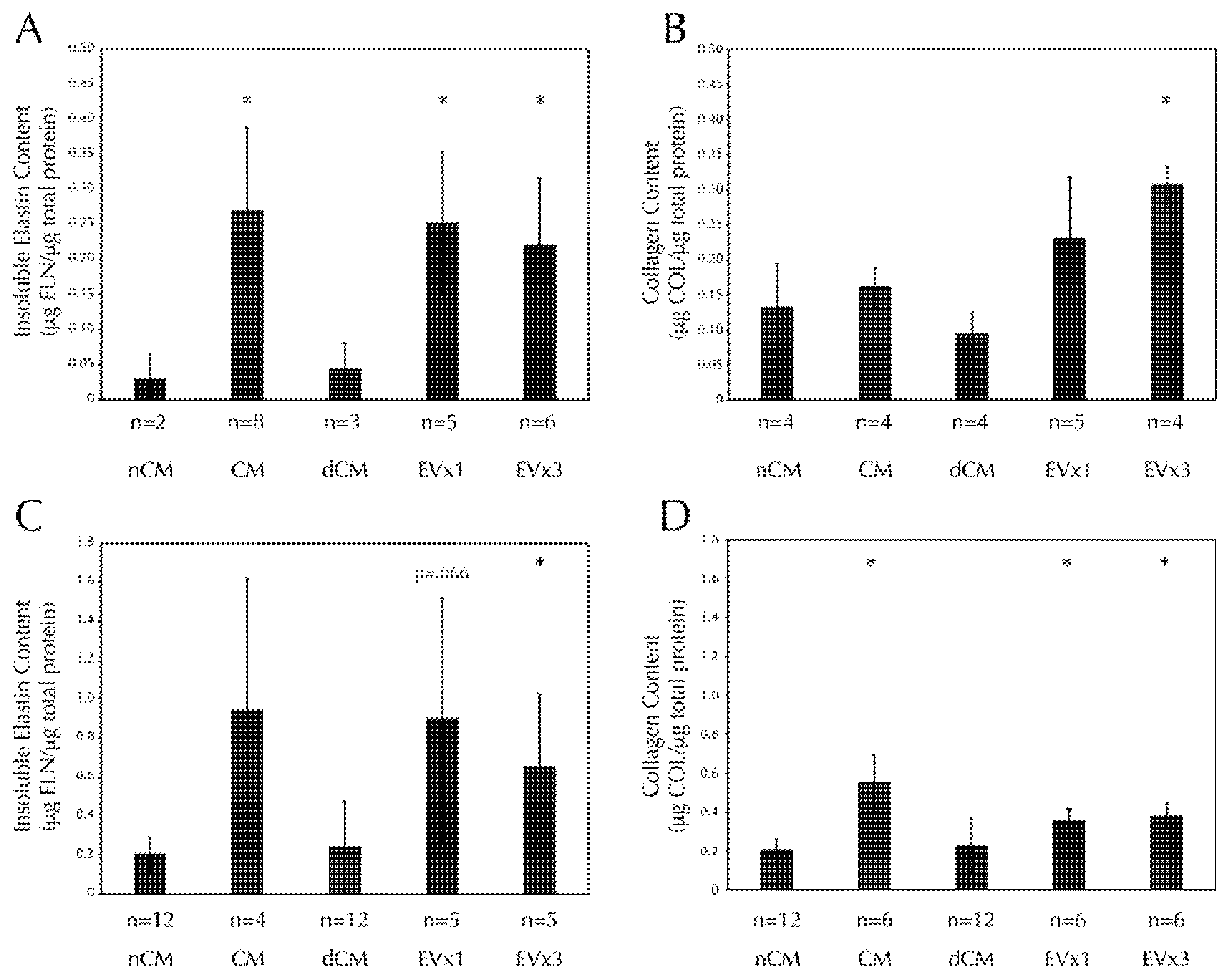
3.5. ASC-Derived EVs Promote Insoluble Elastin and Collagen Deposition within SMC Disc Constructs after 30 Days of Culture
3.6. ASC-Derived EVs Promote Insoluble Elastin and Collagen Deposition within SMC Strand Constructs after 30 Days of Culture
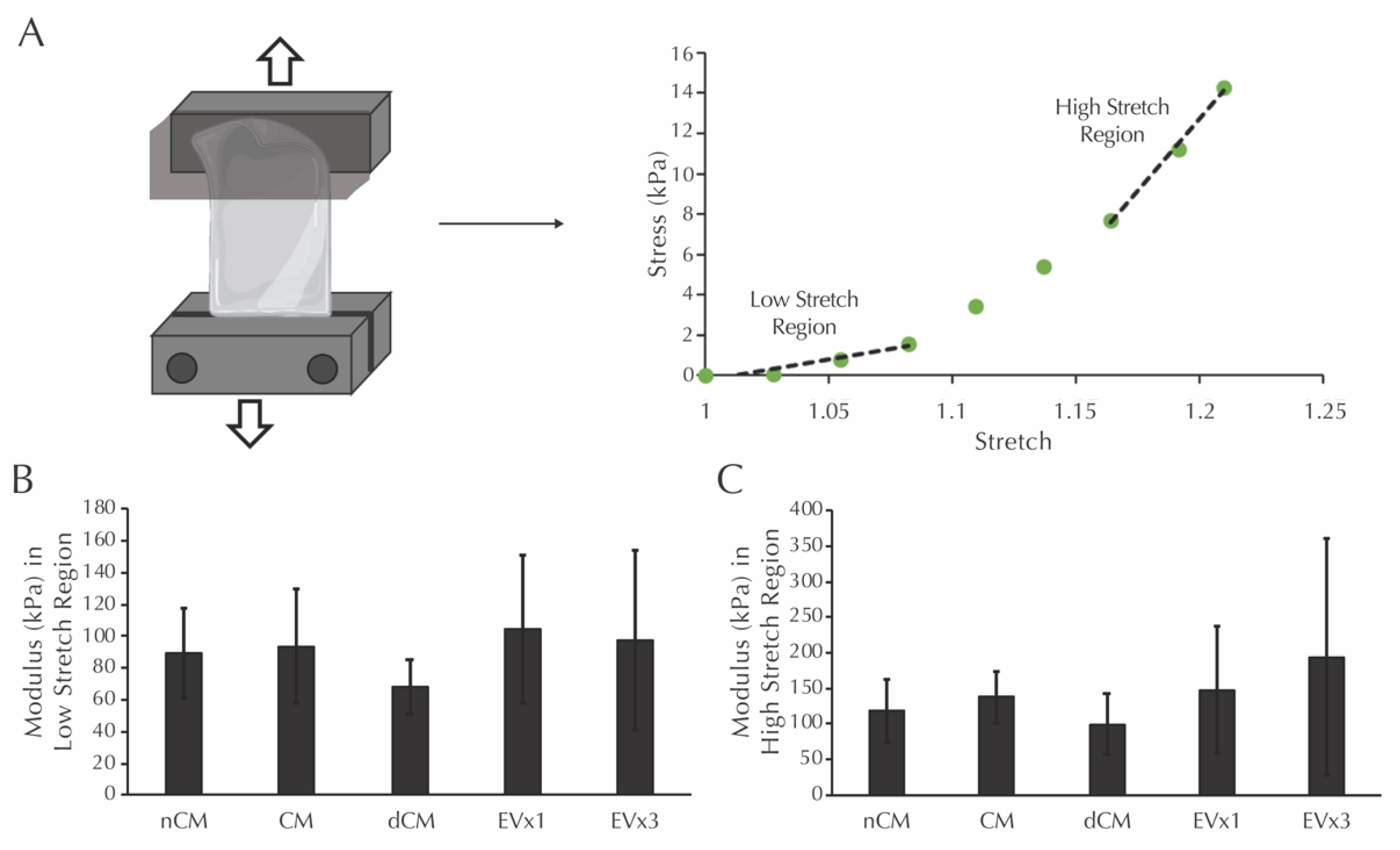
3.7. ASC-Derived EVs Did Not Affect the Modulus in the Low or High Stretch Regions of Strands after 30 Days of Culture
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Abbreviation | Definition |
| AA | Aortic aneurysm |
| ACA | ε-amino caproic acid |
| ASC | Adipose-derived stromal cell |
| BCA | Bicinchoninic acid |
| BM | Basal SMC media |
| BSA | Bovine serum albumin |
| CM | Conditioned ASC culture media |
| CVD | Cardiovascular disease |
| dCM | CM depleted of extracellular vesicles |
| DLS | Dynamic light scattering |
| ECM | Extracellular matrix |
| EV | Extracellular vesicle |
| FBS | Fetal bovine serum |
| nCM | Non-conditioned ASC culture media |
| PBS | Phosphate buffered saline |
| SBM | Supplemented SMC growth media |
| SDS | Sodium dodecyl sulfate |
| SMC | Aortic smooth muscle cell |
| TEM | Transmission electron microscopy |
References
- Virani, S.S.; Alonso, A.; Benjamin, E.J.; Bittencourt, M.S.; Callaway, C.W.; Carson, A.P.; Chamberlain, A.M.; Chang, A.R.; Cheng, S.; Delling, F.N.; et al. American Heart Association Council on E, Prevention Statistics C, Stroke Statistics S. Heart Disease and Stroke Statistics-2020 Update: A Report from the American Heart Association. Circulation 2020, 141, e139–e596. [Google Scholar] [CrossRef] [PubMed]
- Goodney, P.P.; Beck, A.W.; Nagle, J.; Welch, H.G.; Zwolak, R.M. National trends in lower extremity bypass surgery, endovascular interventions, and major amputations. J. Vasc. Surg. 2009, 50, 54–60. [Google Scholar] [CrossRef] [Green Version]
- Alexander, J.H.; Smith, P.K. Coronary-Artery Bypass Grafting. N. Engl. J. Med. 2016, 374, 1954–1964. [Google Scholar] [CrossRef] [Green Version]
- Chun, A.S.; Elefteriades, J.A.; Mukherjee, S.K. Do beta-Blockers Really Work for Prevention of Aortic Aneurysms?: Time for Reassessment. Aorta (Stamford) 2013, 1, 45–51. [Google Scholar] [CrossRef] [Green Version]
- Baxter, B.T.; Terrin, M.C.; Dalman, R.L. Medical management of small abdominal aortic aneurysms. Circulation 2008, 117, 1883–1889. [Google Scholar] [CrossRef]
- Nicholls, S.C.; Gardner, J.B.; Meissner, M.H.; Johansen, H.K. Rupture in small abdominal aortic aneurysms. J. Vasc. Surg. 1998, 28, 884–888. [Google Scholar] [CrossRef] [Green Version]
- Isenberg, B.C.; Williams, C.; Tranquillo, R.T. Small-diameter artificial arteries engineered in vitro. Circ. Res. 2006, 98, 25–35. [Google Scholar] [CrossRef]
- Harskamp, R.E.; Alexander, J.H.; Ferguson, T.B., Jr.; Hager, R.; Mack, M.J.; Englum, B.; Wojdyla, D.; Schulte, P.J.; Kouchoukos, N.T.; de Winter, R.J.; et al. Frequency and Predictors of Internal Mammary Artery Graft Failure and Subsequent Clinical Outcomes: Insights From the Project of Ex-vivo Vein Graft Engineering via Transfection (PREVENT) IV Trial. Circulation 2016, 133, 131–138. [Google Scholar] [CrossRef] [Green Version]
- Desai, M.; Seifalian, A.M.; Hamilton, G. Role of prosthetic conduits in coronary artery bypass grafting. Eur. J. Cardiothorac. Surg. 2011, 40, 394–398. [Google Scholar] [CrossRef] [PubMed]
- Klinkert, P.; Post, P.N.; Breslau, P.J.; van Bockel, J.H. Saphenous vein versus PTFE for above-knee femoropopliteal bypass. A review of the literature. Eur. J. Vasc. Endovasc. Surg. 2004, 27, 357–362. [Google Scholar] [CrossRef] [Green Version]
- Ramaswamy, A.K.; Vorp, D.A.; Weinbaum, J.S. Functional Vascular Tissue Engineering Inspired by Matricellular Proteins. Front. Cardiovasc. Med. 2019, 6, 74. [Google Scholar] [CrossRef] [Green Version]
- Cunnane, E.M.; Weinbaum, J.S.; O’Brien, F.J.; Vorp, D.A. Future Perspectives on the Role of Stem Cells and Extracellular Vesicles in Vascular Tissue Regeneration. Front. Cardiovasc. Med. 2018, 5, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saitow, C.B.; Wise, S.G.; Weiss, A.S.; Castellot, J.J.; Kaplan, D.L. Elastin biology and tissue engineering with adult cells. Biomol. Concepts 2013, 4, 173–185. [Google Scholar] [CrossRef] [Green Version]
- Kelleher, C.M.; McLean, S.E.; Mecham, R.P. Vascular extracellular matrix and aortic development. Curr. Top. Dev. Biol. 2004, 62, 153–188. [Google Scholar] [CrossRef]
- Ramaswamy, A.K.; Sides, R.E.; Cunnane, E.M.; Lorentz, K.L.; Reines, L.M.; Vorp, D.A.; Weinbaum, J.S. Adipose-derived stromal cell secreted factors induce the elastogenesis cascade within 3D aortic smooth muscle cell constructs. Matrix Biol. Plus. 2019, 4. [Google Scholar] [CrossRef]
- Sharma, A.K.; Salmon, M.D.; Lu, G.; Su, G.; Pope, N.H.; Smith, J.R.; Weiss, M.L.; Upchurch, G.R., Jr. Mesenchymal Stem Cells Attenuate NADPH Oxidase-Dependent High Mobility Group Box 1 Production and Inhibit Abdominal Aortic Aneurysms. Arterioscler. Thromb. Vasc. Biol. 2016, 36, 908–918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spinosa, M.; Lu, G.; Su, G.; Bontha, S.V.; Gehrau, R.; Salmon, M.D.; Smith, J.R.; Weiss, M.L.; Mas, V.R.; Upchurch, G.R., Jr.; et al. Human mesenchymal stromal cell-derived extracellular vesicles attenuate aortic aneurysm formation and macrophage activation via microRNA-147. FASEB J. 2018, fj201701138RR. [Google Scholar] [CrossRef] [PubMed]
- Eirin, A.; Riester, S.M.; Zhu, X.Y.; Tang, H.; Evans, J.M.; O’Brien, D.; van Wijnen, A.J.; Lerman, L.O. MicroRNA and mRNA cargo of extracellular vesicles from porcine adipose tissue-derived mesenchymal stem cells. Gene 2014, 551, 55–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eirin, A.; Zhu, X.Y.; Puranik, A.S.; Woollard, J.R.; Tang, H.; Dasari, S.; Lerman, A.; van Wijnen, A.J.; Lerman, L.O. Integrated transcriptomic and proteomic analysis of the molecular cargo of extracellular vesicles derived from porcine adipose tissue-derived mesenchymal stem cells. PLoS ONE 2017, 12, e0174303. [Google Scholar] [CrossRef] [PubMed]
- Eirin, A.; Zhu, X.Y.; Puranik, A.S.; Woollard, J.R.; Tang, H.; Dasari, S.; Lerman, A.; van Wijnen, A.J.; Lerman, L.O. Comparative proteomic analysis of extracellular vesicles isolated from porcine adipose tissue-derived mesenchymal stem/stromal cells. Sci. Rep. 2016, 6, 36120. [Google Scholar] [CrossRef]
- Haskett, D.G.; Saleh, K.S.; Lorentz, K.L.; Josowitz, A.D.; Luketich, S.K.; Weinbaum, J.S.; Kokai, L.E.; D’Amore, A.; Marra, K.G.; Rubin, J.P.; et al. An exploratory study on the preparation and evaluation of a "same-day" adipose stem cell-based tissue-engineered vascular graft. J. Thorac. Cardiovasc. Surg. 2018, 156, 1814–1822 e3. [Google Scholar] [CrossRef]
- Krawiec, J.T.; Liao, H.T.; Kwan, L.L.; D’Amore, A.; Weinbaum, J.S.; Rubin, J.P.; Wagner, W.R.; Vorp, D.A. Evaluation of the stromal vascular fraction of adipose tissue as the basis for a stem cell-based tissue-engineered vascular graft. J. Vasc. Surg. 2017, 66, 883–890 e1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmann, K.A.; Weinbaum, J.S.; Johnson, S.L.; Tranquillo, R.T. Fibrin degradation enhances vascular smooth muscle cell proliferation and matrix deposition in fibrin-based tissue constructs fabricated in vitro. Tissue Eng. Part A 2010, 16, 3261–3270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mulvihill, J.J.; Walsh, M.T. On the mechanical behaviour of carotid artery plaques: The influence of curve-fitting experimental data on numerical model results. Biomech. Model. Mechanobiol. 2013, 12, 975–985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blose, K.J.; Pichamuthu, J.E.; Weinbaum, J.S.; Vorp, D.A. Design and Validation of a Vacuum Assisted Anchorage for the Uniaxial Tensile Testing of Soft Materials. Soft Mater. 2016, 14, 72–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsamis, A.; Phillippi, J.A.; Koch, R.G.; Pasta, S.; D’Amore, A.; Watkins, S.C.; Wagner, W.R.; Gleason, T.G.; Vorp, D.A. Fiber micro-architecture in the longitudinal-radial and circumferential-radial planes of ascending thoracic aortic aneurysm media. J. Biomech. 2013, 46, 2787–2794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holzapfel, G.A.; Sommer, G.; Regitnig, P. Anisotropic mechanical properties of tissue components in human atherosclerotic plaques. J. Biomech. Eng. 2004, 126, 657–665. [Google Scholar] [CrossRef] [PubMed]
- Walsh, M.T.; Cunnane, E.M.; Mulvihill, J.J.; Akyildiz, A.C.; Gijsen, F.J.; Holzapfel, G.A. Uniaxial tensile testing approaches for characterisation of atherosclerotic plaques. J. Biomech. 2014, 47, 793–804. [Google Scholar] [CrossRef] [PubMed]
- Holzapfel, G.A. Determination of material models for arterial walls from uniaxial extension tests and histological structure. J. Theor. Biol. 2006, 238, 290–302. [Google Scholar] [CrossRef] [Green Version]
- Cunnane, E.M.; Mulvihill, J.J.E.; Barrett, H.E.; Hennessy, M.M.; Kavanagh, E.G.; Walsh, M.T. Mechanical properties and composition of carotid and femoral atherosclerotic plaques: A comparative study. J. Biomech. 2016, 49, 3697–3704. [Google Scholar] [CrossRef] [PubMed]
- Thery, C.; Ostrowski, M.; Segura, E. Membrane vesicles as conveyors of immune responses. Nat. Rev. Immunol. 2009, 9, 581–593. [Google Scholar] [CrossRef]
- Blose, K.J.; Ennis, T.L.; Arif, B.; Weinbaum, J.S.; Curci, J.A.; Vorp, D.A. Periadventitial adipose-derived stem cell treatment halts elastase-induced abdominal aortic aneurysm progression. Regen. Med. 2014, 9, 733–741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cunnane, E.M.; Lorentz, K.L.; Ramaswamy, A.K.; Gupta, P.; Mandal, B.B.; O’Brien, F.J.; Weinbaum, J.S.; Vorp, D.A. Extracellular Vesicles Enhance the Remodeling of Cell-Free Silk Vascular Scaffolds in Rat Aortae. ACS Appl. Mater. Interfaces 2020, 12, 26955–26965. [Google Scholar] [CrossRef]
- Li, P.; Kaslan, M.; Lee, S.H.; Yao, J.; Gao, Z. Progress in Exosome Isolation Techniques. Theranostics 2017, 7, 789–804. [Google Scholar] [CrossRef]
- Lotvall, J.; Hill, A.F.; Hochberg, F.; Buzas, E.I.; Di Vizio, D.; Gardiner, C.; Gho, Y.S.; Kurochkin, I.V.; Mathivanan, S.; Quesenberry, P.; et al. Minimal experimental requirements for definition of extracellular vesicles and their functions: A position statement from the International Society for Extracellular Vesicles. J. Extracell. Vesicles 2014, 3, 26913. [Google Scholar] [CrossRef] [PubMed]
- Webber, J.; Clayton, A. How pure are your vesicles? J. Extracell. Vesicles 2013, 2. [Google Scholar] [CrossRef] [PubMed]
- Collins, M.J.; Eberth, J.F.; Wilson, E.; Humphrey, J.D. Acute mechanical effects of elastase on the infrarenal mouse aorta: Implications for models of aneurysms. J. Biomech. 2012, 45, 660–665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, A.J.; O’Brien, F.J. Insoluble elastin reduces collagen scaffold stiffness, improves viscoelastic properties, and induces a contractile phenotype in smooth muscle cells. Biomaterials 2015, 73, 296–307. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cunnane, E.M.; Ramaswamy, A.K.; Lorentz, K.L.; Vorp, D.A.; Weinbaum, J.S. Extracellular Vesicles Derived from Primary Adipose Stromal Cells Induce Elastin and Collagen Deposition by Smooth Muscle Cells within 3D Fibrin Gel Culture. Bioengineering 2021, 8, 51. https://0-doi-org.brum.beds.ac.uk/10.3390/bioengineering8050051
Cunnane EM, Ramaswamy AK, Lorentz KL, Vorp DA, Weinbaum JS. Extracellular Vesicles Derived from Primary Adipose Stromal Cells Induce Elastin and Collagen Deposition by Smooth Muscle Cells within 3D Fibrin Gel Culture. Bioengineering. 2021; 8(5):51. https://0-doi-org.brum.beds.ac.uk/10.3390/bioengineering8050051
Chicago/Turabian StyleCunnane, Eoghan M., Aneesh K. Ramaswamy, Katherine L. Lorentz, David A. Vorp, and Justin S. Weinbaum. 2021. "Extracellular Vesicles Derived from Primary Adipose Stromal Cells Induce Elastin and Collagen Deposition by Smooth Muscle Cells within 3D Fibrin Gel Culture" Bioengineering 8, no. 5: 51. https://0-doi-org.brum.beds.ac.uk/10.3390/bioengineering8050051