
Wine Phenolic Compounds: Antimicrobial Properties against Yeasts, Lactic Acid and Acetic Acid Bacteria

Microbiology & Wine Research, Institute of Molecular Physiology, Johannes Gutenberg-University, Mainz 55099, Germany
*
Author to whom correspondence should be addressed.
Beverages 2017, 3(3), 29; https://0-doi-org.brum.beds.ac.uk/10.3390/beverages3030029
Submission received: 24 April 2017
/
Revised: 2 June 2017
/
Accepted: 23 June 2017
/
Published: 29 June 2017
(This article belongs to the Special Issue Phenolic Compounds in Fruit Beverages)
Abstract
:Microorganisms play an important role in the conversion of grape juice into wine. Yeasts belonging the genus Saccharomyces are mainly responsible for the production of ethanol, but members of other genera are known as producers of off-flavors, e.g., volatile phenols. Lactic acid and acetic acid bacteria also occur regularly in must and wine. They are mostly undesirable due to their capacity to produce wine-spoiling compounds (acetic acid, biogenic amines, N-heterocycles, diacetyl, etc.). In conventional winemaking, additions of sulfite or lysozyme are used to inhibit growth of spoilage microorganisms. However, there is increasing concern about the health risks connected with these enological additives and high interest in finding alternatives. Phenols are naturally occurring compounds in grapes and wine and are well known for their antimicrobial and health-promoting activities. In this study, we tested a selection of phenolic compounds for their effect on growth and viability of wine-associated yeasts and bacteria. Our investigations confirmed the antimicrobial activities of ferulic acid and resveratrol described in previous studies. In addition, we found syringaldehyde highly efficient against wine-spoiling bacteria at concentrations of 250–1000 µg/mL. The promising bioactive activities of this aromatic aldehyde and its potential for winemaking deserves further research.
Keywords:
wine; microorganisms; phenols; sulfite; growth inhibition; syringaldehyde; resveratrol; laccase1. Introduction
Yeasts, lactic acid bacteria and acetic acid bacteria can play a role in winemaking and have an influence on wine quality [1]. Bacterial growth in musts and wines is conventionally controlled by the addition of sulfur dioxide. Unfortunately, the presence of sulfites in alcoholic beverages, particularly in wines, can cause pseudo-allergic responses with symptoms ranging from gastrointestinal problems to anaphylactic shock [2,3]. Other antimicrobials such as sorbic acid and dimethyl dicarbonate are active against yeasts, but have less activity against bacteria [2,3].
Phenolic compounds affect bacterial growth and metabolism [2]. Enological extracts have been found to inhibit clinically important microorganisms like Staphylococcus aureus, Escherichia coli, Bacillus cereus, Campylobacter jejuni, Salmonella infantis and Candida albicans [4,5,6,7]. Phenols kill microorganisms or inhibit the growth of bacteria, fungi or protozoa [8]. Several mechanisms of action in the growth inhibition of bacteria are involved, such as permeabilization and destabilization of the plasma membrane, or inhibition of extracellular microbial enzymes [2,9].
García-Ruiz et al. [10] investigated the inhibitory potential of 18 phenolic compounds (including hydroxybenzoic acids, hydroxycinnamic acids, phenolic alcohols, stilbenes, flavan-3-ols and flavonols) on wine-related lactic acid bacteria of the species Oenococcus oeni, Lactobacillus hilgardii and Pediococcus pentosaceus. In general, flavonols and stilbenes showed the greatest inhibitory effects on the growth of the test strains. Hydroxycinnamic acids and hydroxybenzoic acids and their esters exhibited medium inhibitory effect, and phenolic alcohols as well as flavanol-3-ols showed the lowest effect on the growth of the strains studied. In comparison to the antimicrobial additives used in winemaking, the inhibitory impact of most phenolic compounds was stronger than those of the commonly used potassium metabisulfite and lysozyme.
In a subsequent study, García-Ruiz et al. [11] determined whether phenolic extracts with antimicrobial activity may be considered as an alternative to the use of sulfur dioxide (SO2) for controlling malolactic fermentation in winemaking. Growth inhibition of six enological strains of Lactobacillus hilgardii, Lactobacillus casei, Lactobacillus plantarum, Pediococcus pentosaceus and Oenococcus oeni by phenolic extracts from different origins (spices, flowers, leaves, fruits, legumes, seeds, skins, agricultural by-products and others) was evaluated. A total of 24 extracts were found to significantly inhibit the growth of at least two of the strains studied. Some of these extracts were also active against acetic acid bacteria.
In a similar approach, Pastorkova et al. [12] investigated the antimicrobial potential of 15 grape phenolic compounds of various chemical classes (phenolic acids, stilbenes and flavonoids) against yeasts and acetic acid bacteria. Pterostilbene, resveratrol and luteolin were among six active compounds that possessed the strongest inhibitory effects against all microorganisms tested. In the case of phenolic acids, myricetin, p-coumaric and ferulic acids exhibited selective antimicrobial activity depending upon the species of yeasts and bacteria tested. In comparison with potassium metabisulfite, all microorganisms tested were more susceptible to the phenols.
Recent studies suggested the possible application of plant phenolic extracts to reduce the use of sulfites in winemaking [13]. The use of polyphenols as a SO2 substitute thus presupposes that wine yeasts are not influenced by the phenols in their fermentation activity, and only undesired organisms are inhibited. Hitherto, many studies have been concerned with the antimicrobial action of phenols against lactic acid bacteria but less with acetic acid bacteria and yeasts. The challenge is to find differences in the inhibitory effect of phenols within the various desirable and undesirable species.
2. Materials and Methods
2.1. Microorganisms and Culture Conditions
The yeasts Wickerhamomyces anomalus AS1, Saccharomyces bayanus HL 77, Saccharomyces cerevisiae × Saccharomyces kudriavzevii × Saccharomyces HL 78 and Saccharomyces cerevisiae 16.1 have been isolated from local wineries in Germany and characterized as described previously [14,15,16]. Debaryomyces hansenii 525 has been isolated from garden soil [17]. Strains of lactic acid bacteria (Lactobacillus hilgardii 20166, Lactobacillus plantarum 20174T, Pediococcus parvulus 20332T, Oenococcus oeni 20252) and acetic acid bacteria (Gluconobacter cerinus 9533, Acetobacter acetii 3508) were obtained from the German Collection of Microorganisms and Cell Cultures (DSMZ, Braunschweig).
Yeasts were cultivated in Sabouraud-glucose broth (DSMZ medium 1429) at pH 5.5 or 3.5 depending on the specific experimental conditions. Lactic acid bacteria (were cultivated in MRS medium (pH 5.5) containing tomato juice (DSMZ medium 11). G. cerinus 9533 was grown in YPM broth (DSMZ medium 360, pH 6.1) and A. acetii 3508 in YPS broth (with sorbitol instead of mannitol in YPM, pH 6.6). All cultures were incubated on a shaker at 30 °C.
2.2. Chemicals
All reagents were of analytical grade. Polydatin, caffeic acid, ferulic acid, p-coumaric acid, sinapic acid, 3,4-dihydroxybenzoic acid, gallic acid, ethylgallate, vanillic acid, syringaldehyde, and DMSO were purchased from Sigma-Aldrich (Taufkirchen, Germany). Resveratrol and microbiological culture media or constituents were supplied by Roth (Karlsruhe, Germany). Laccase from Botrytis cinerea strain P16–14 was prepared as described recently [18].
2.3. Influence of Ethanol and DMSO on Yeast Growth
The phenolic compounds were prepared in organic solvents. Their possible impact on yeast growth was tested as follows: Sabouraud-glucose broth media (pH 3.5) were prepared with 0, 1, 5, 10, 15 and 20% v/v ethanol and DMSO, respectively. The nutrient solutions were inoculated at initial cell numbers of 106/mL and incubated at 30 °C on a shaker (75 rpm) for four days. Growth of the cultures was recorded daily by determination of their optical densities (OD590) for 4 days.
2.4. Influence of Phenols on Growth of Microorganisms
The effect of phenols on the growth of microbial liquid cultures was tested at different concentrations by monitoring the increase of cell densities for several days.
2.4.1. Method 1
The first experimental series was conducted using common glass culture tubes. Phenolic compounds were dissolved at concentrations of 5, 10 and 20 mg/mL in DMSO (99.4% v/v) and sterile-filtrated (0.2 µm). From each solution, 210 µL were added separately to 3.99 mL autoclaved Sabouraud-glucose broth (pH 3.5) to achieve final phenol concentrations of 250, 500 and 1000 µg/mL. The control media were prepared with 5% (v/v) DMSO. The inhibition effect of potassium metabisulfite on microbial cultures was compared at the same concentrations as used for the phenolic compounds.
The media were inoculated at initial yeast cell numbers of 106/mL and incubated at 30 °C on a shaker (75 rpm). Growth of the cultures was monitored by determination of their optical densities (OD590) after 3 or 4 days with a Colorimeter model 45 (Fisher Scientific, Cambridge, UK). The optical densities of the DMSO controls were defined as 100% in relation to growth under the influence of phenolic compounds. The results are presented as the means of duplicate assays.
2.4.2. Method 2
The second experimental series was conducted in sterile 96-well plastic microplates (Roth, Karlsruhe, Germany). The phenolic compounds were dissolved at concentrations of 5, 10 and 20 mg/mL in DMSO (50% v/v) and sterile-filtrated (0.2 µm). Ten µL of each concentration and the DMSO control were added to 190 µL precultures of exponentially growing microorganisms (yeasts: 1 × 105 cells/ml; bacteria 1 × 106 cells/mL). The final concentrations of phenols were 250, 500 and 1000 µg/mL. The microplates were incubated in a wet chamber at 30 °C, acetic acid bacteria on a shaker. Growth was monitored by measuring the OD590 spectrophotometrically using a FLUOstar Omega microplate reader (BMG Labtech GmbH, Germany). The time of evaluation was set when the DMSO control reached the middle of the exponential growth phase. The corresponding optical densities were defined as 100% in relation to growth under the influence of phenolic compounds. The results are presented as the means of triplicate assays.
2.5. Influence of Laccase Oxidation on the Inhibition Effect of Phenolic Compounds
Abiotic and/or enzymatic oxidation may alter structure and reactivity of phenols; particularly laccase, a phenoloxidase produced by the wine-relevant fungus Botrytis cinerea plays an important role in this respect. For this reason, we tested the effect of enzymatic oxidations on the antimicrobial activities of selected phenols. The compounds were dissolved at concentrations of 5, 10 and 20 mg/mL in DMSO (50% v/v) and sterile-filtrated (0.2 µm). To the solutions 0.03 units/mL laccase of Botrytis cinerea P16–14 was added and incubation followed for 48 h at 30 °C. The controls contained no phenoloxidase. The next steps were performed as described above for method 2.
3. Results
3.1. Effect of Solvents
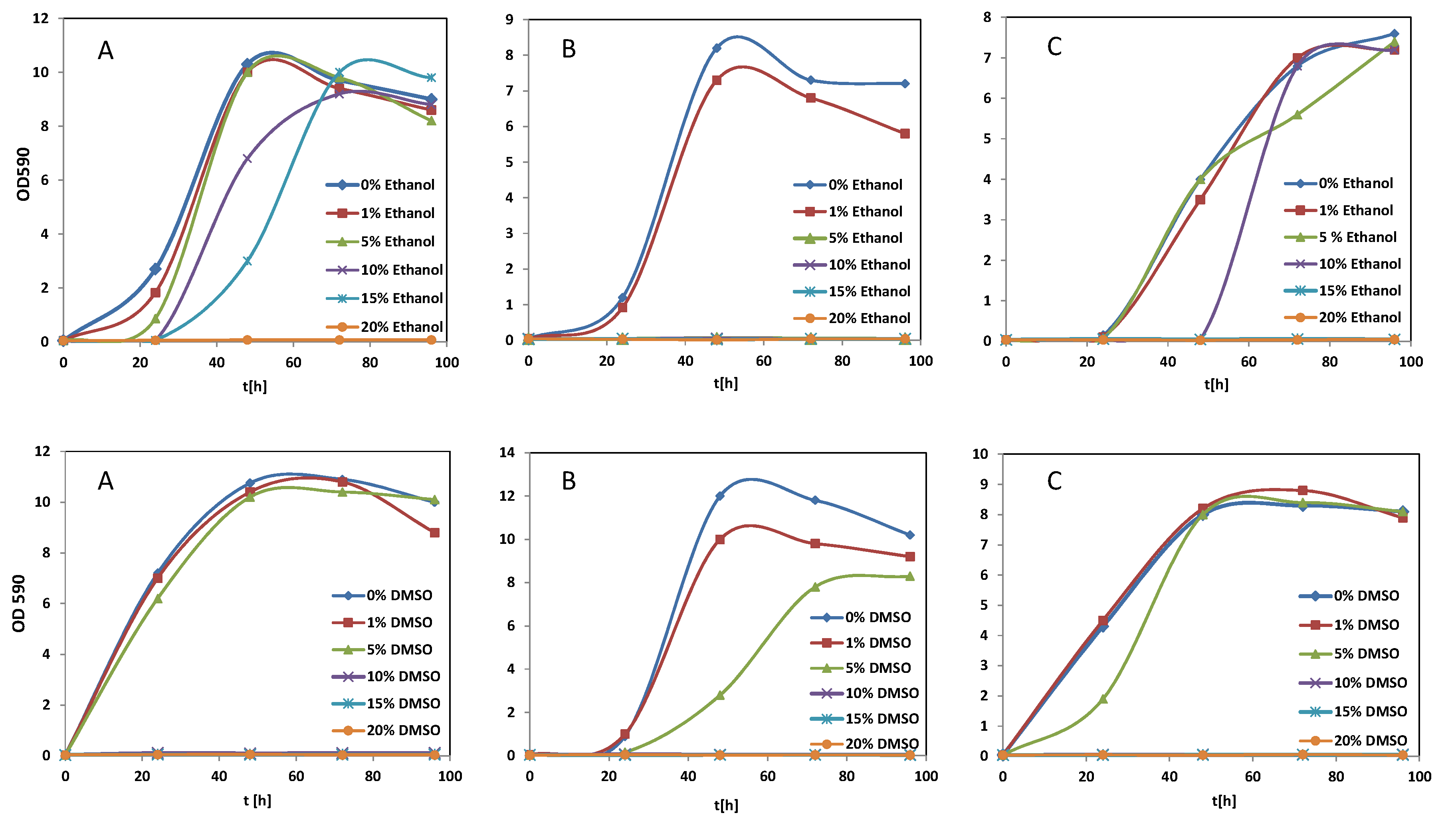
The phenolic compounds tested in this study are poorly soluble in water. For this reason, stock solutions had to be prepared in organic solvents such as dimethoxy sulfoxide (DMSO) or ethanol. As these solvents could have an impact on the inhibition experiments, we investigated the effect of different concentrations on the growth of the test microorganisms (Figure 1).
The Saccharomycetes exhibited quite different tolerances towards ethanol in the culture media. The most sensitive strain was S. bayanus HL 77, already 1% v/v alcohol inhibited growth. The most resistant S. cerevisiae 16.1 showed still—although a delayed—growth at 15% v/v, whereas the triple hybrid HL 78 was completely inhibited by this concentration (Figure 1, top row). A higher ethanol tolerance was expected for the wine yeast S. bayanus HL 77; however, it should be considered that in natural must fermentations yeasts have the chance for gradual adaptation to increasing ethanol concentrations, whereas in our experiments high amounts were already present from the start of cultivation. W. anomalus AS1 showed delayed growth at 10% ethanol in the medium and was completely inhibited by 15% and Debaryomyces hansenii 525 tolerated only 1% (not shown).
Regarding DMSO (Figure 1, bottom row), S. cerevisiae 16.1 showed no restricted growth in presence of 5% v/v, but complete failure at 10% v/v. This finding is in accordance with results of Sadowska-Bartosz et al. [19] reporting an inhibiting concentration of 5–10% v/v DMSO for this species. S. bayanus HL 77 showed delayed growth at 5% v/v, but reached still high cell densities after 4 days. Again, 10% v/v was not tolerated by this strain. This concentration was also the limit for the triple hybrid HL 78, which showed nearly normal growth behavior at 5% v/v. W. anomalus AS1 was not inhibited up to 5% v/v DMSO, whereas cell densities of D. hansenii 525 were drastically reduced at this concentration (not shown).
Although yeast growth, with the exception of strain 525, was generally not significantly reduced at 5% v/v DMSO, a second experimental series was performed with only half of this concentration. At 2.5% v/v, also lactic acid and acetic acid bacteria reached the same cell densities as the controls without DMSO. Only A. acetii 3508T was more susceptible, the maximum OD595 was only 60% of the control (not shown).
3.2. Influence of Phenols on Yeast Growth
Different classes of phenolic compounds were tested for their antimicrobial activities (Table 1).
The first experimental series was performed at 5% v/v DMSO in the culture medium of pH 3.5 (Figure 2).
Growth of S. cerevisiae 16.1 was generally not negatively impaired by phenols. Only high concentrations of ferulic acid or potassium sulfite led to complete inhibition. In contrast, S. bayanus HL 77 and the triple hybrid were depressed by all phenols, even at low concentrations of p-coumaric acid and ferulic acid.
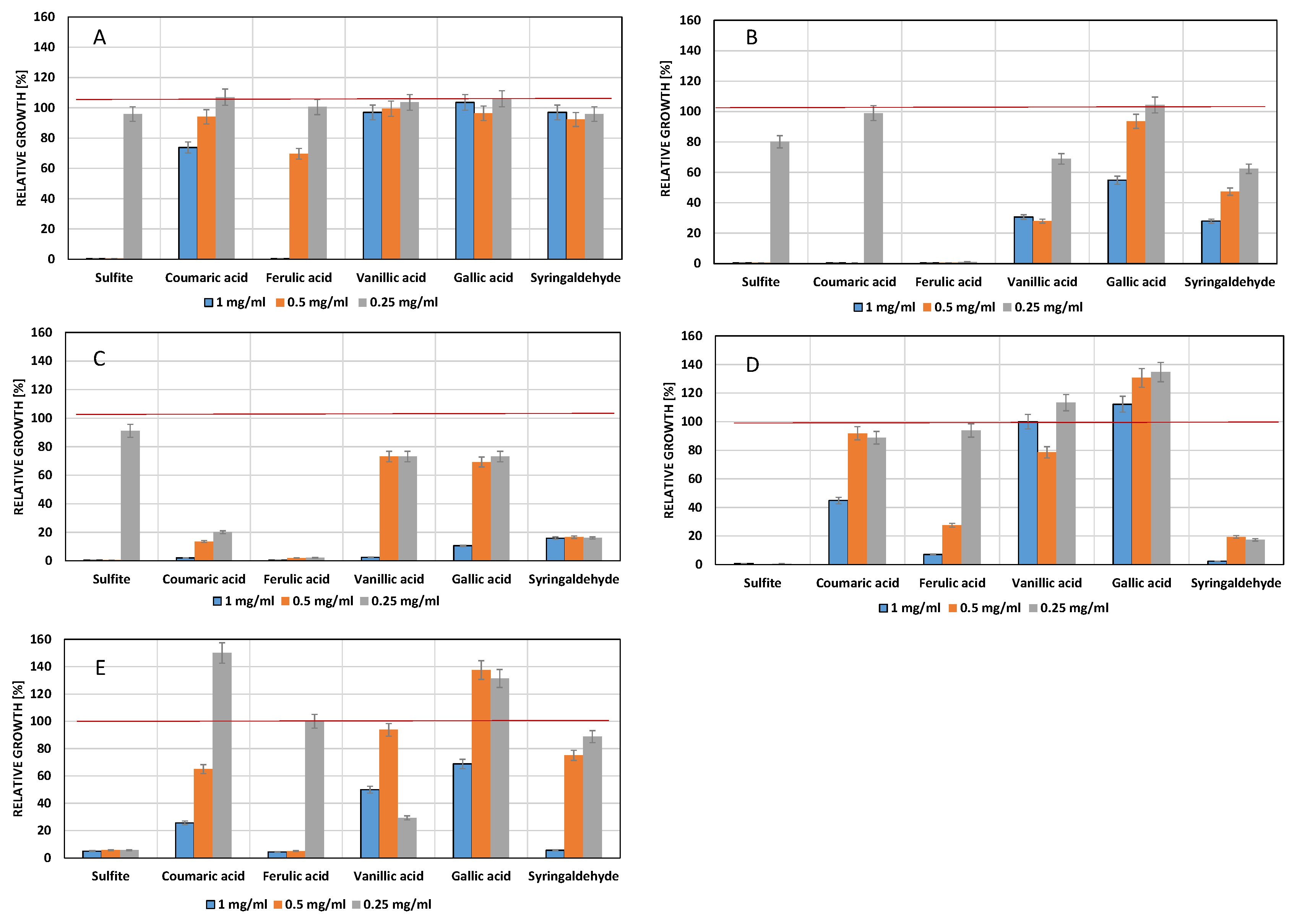
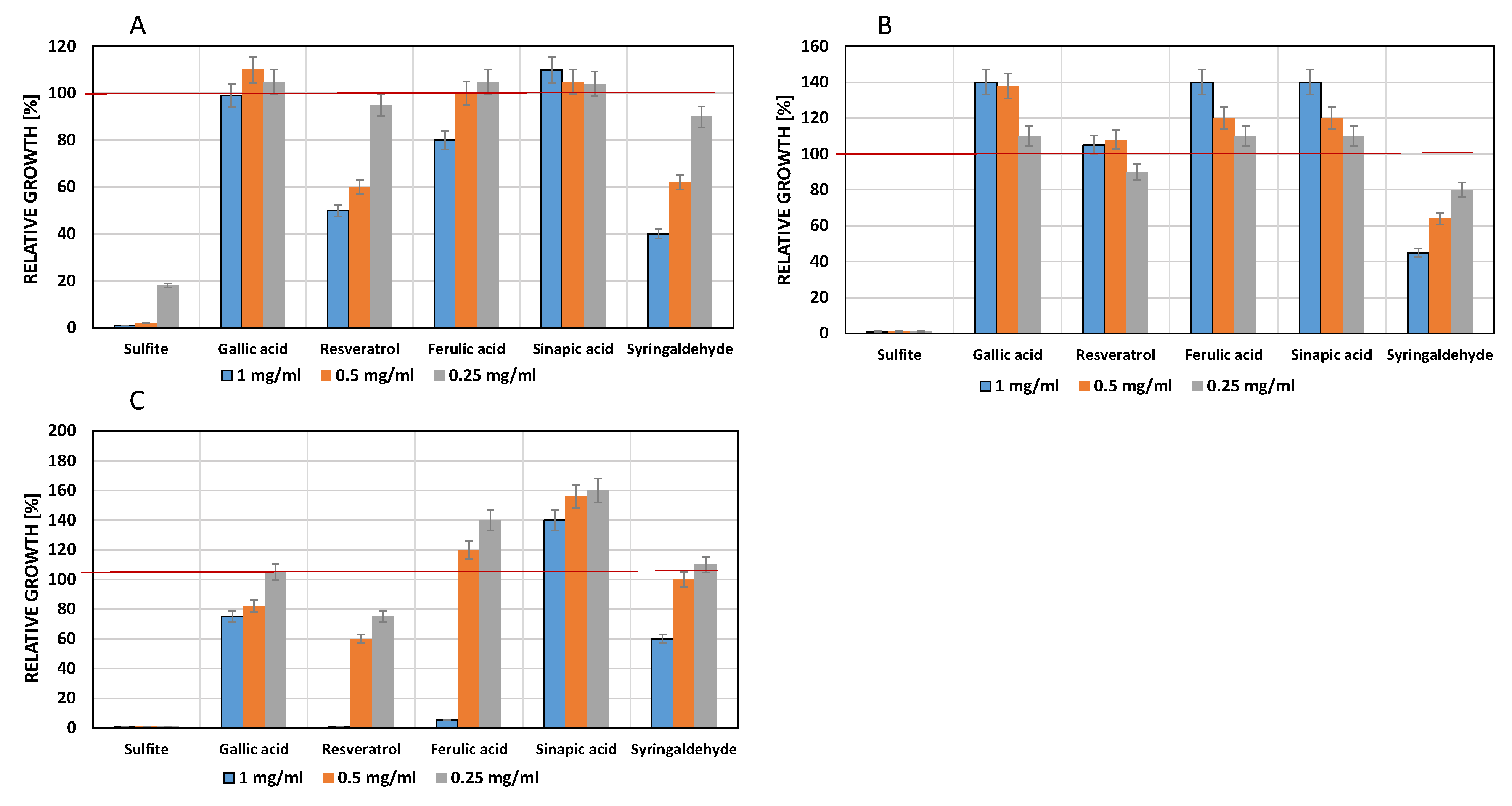
The red line indicates maximum growth (optical density) of the controls which contained DMSO [5.0% v/v].
W. anomalus AS1 and D. hansenii 525 were sensitive to low concentrations of sulfite, ferulic acid and syringaldehyde. On the other hand, gallic acid stimulated growth of both non-Saccharomycetes.
The second experimental series was performed at 2.5% v/v DMSO in the culture medium of pH 5.5 (Figure 3).
Growth of S. cerevisiae 16.1 was completely inhibited by high concentrations of sulfite, resveratrol and ferulic acid and significantly impaired by syringaldehyde. The residual six phenols had no effect. The fact that potassium sulfite was not as effective as in the previous experiments can be explained in a diminished concentration of the effective molecular form (SO2) at pH 5.5 compared to pH 3.5. S. bayanus HL 77 showed a very similar pattern, again resveratrol, ferulic acid and syringaldehyde were the most antimicrobial. Under the given experimental conditions, both non-Saccharomycetes turned out to be rather resistant to most phenols. However, a strong negative effect was exerted by resveratrol, followed by syringaldehyde and ferulic acid.
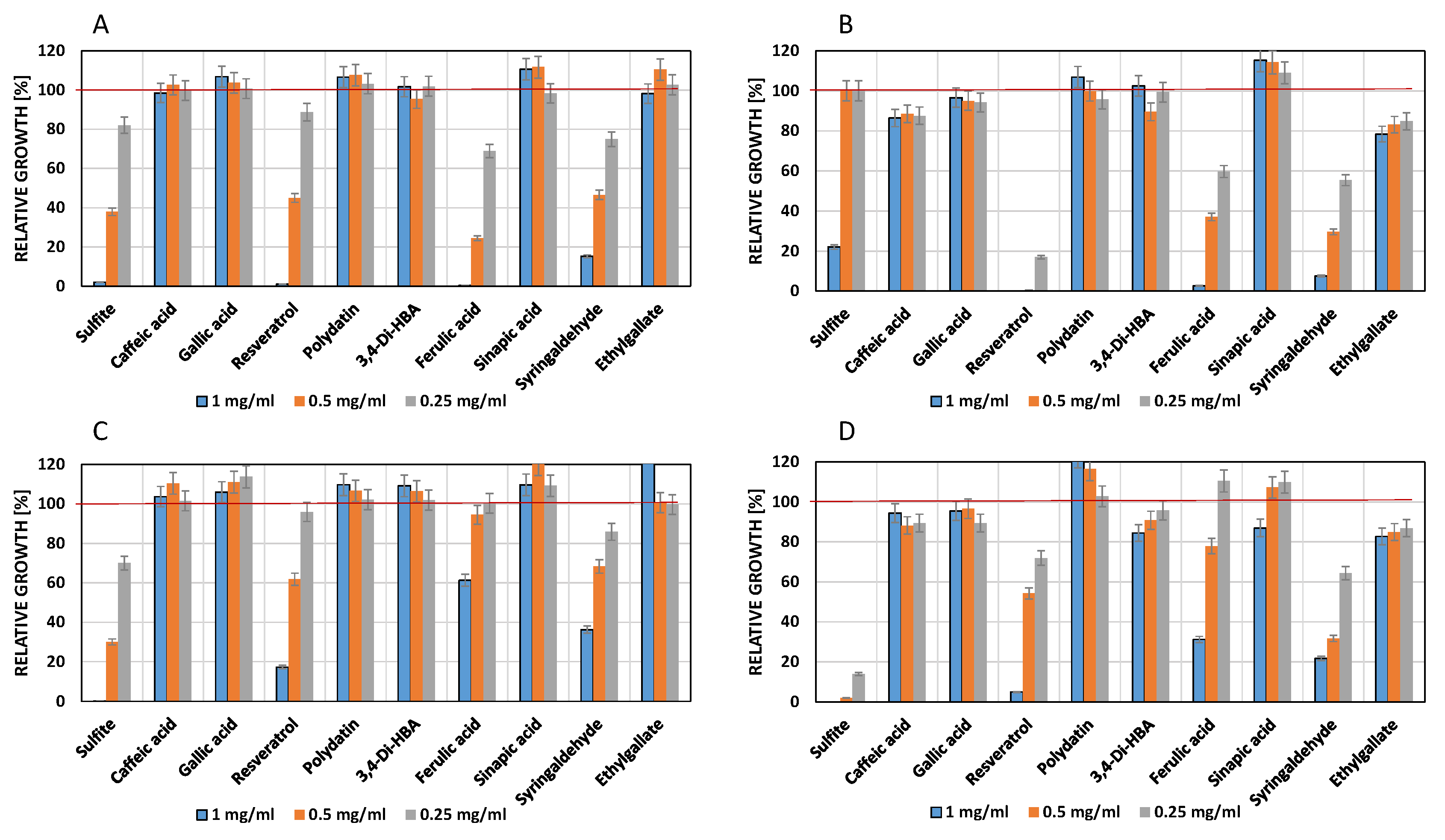
3.3. Influence of Phenols on Growth of Lactic Acid Bacteria
The four lactic acid bacteria were completely inhibited by 0.25 mg/mL sulfite (Figure 4). As in the case of yeasts, resveratrol but not its glycosylated form (polydatin) had a strong antimicrobial effect. The cinnamic acids caffeic acid, ferulic acid and sinapic acid decreased cell densities in a concentration-dependent manner. Syringaldehyde at 1.0 mg/mL completely inhibited growth. On the other hand, gallic acid had no effect or stimulated growth of Pediococcus and Oenococcus.
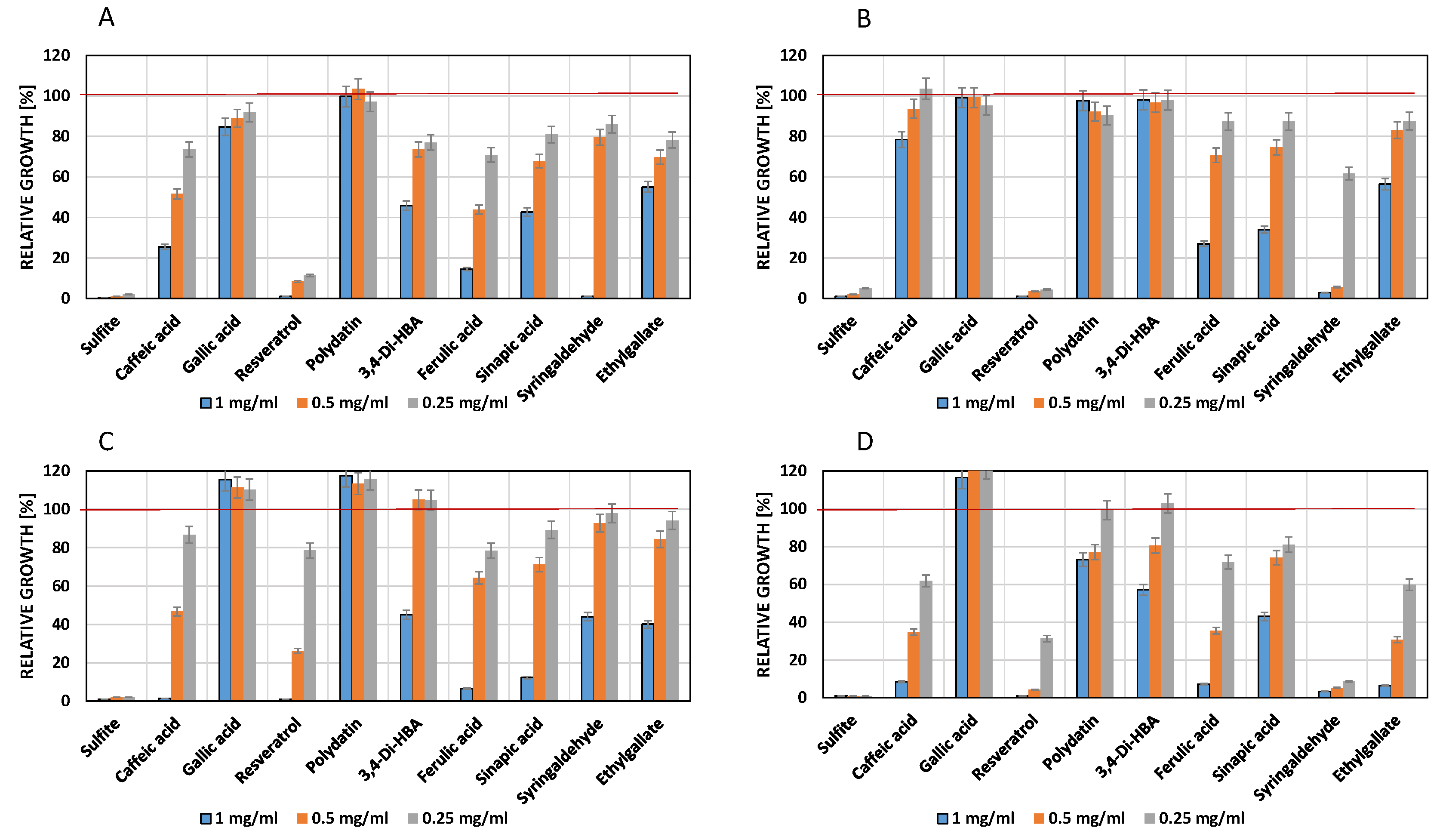
3.4. Influence of Phenols on Growth of Acetic Acid Bacteria
In contrast to previous experiments, browning reactions were observed in the cultures, indicating oxidation of the phenols. Thus, only the hitherto most effective compounds were tested (Figure 5). Acetic acid bacteria did not grow in the presence of sulfite. G. cerinus 9533 was significantly inhibited by syringaldehyde. The cinnamic acids showed no—and resveratrol only a transient—negative effect. Gallic acid favored growth, as already observed with other microorganisms before. A. acetii 358 was inhibited by high concentrations of ferulic acid and resveratrol.
3.5. Effects of Enzymatic Oxidation
As observed with acetic acid bacteria in our experiments, oxidative transformations may alter the molecular structure and reactivity of phenols. In must and wine, phenols can be enzymatically oxidized by the action of phenoloxidases deriving from grapes or fungal sources [18,20]. In this study, we tested the antimicrobial action of phenolic compounds after oxidation with laccase from Botrytis cinerea (Table 2).
After oxidation with fungal laccase the antimicrobial impact of phenolic compounds was diminished or enhanced compared to the untreated controls. In general, oxidized caffeic acid and ferulic acid were less toxic to all test microorganisms. In addition, lactic acid bacteria took benefit by oxidation of sinapic acid, gallic acid and ethylgallate. In contrast, polydatin inhibited yeast growth much stronger after oxidation than before. No general trend can be derived with respect to laccase oxidation for resveratrol, hydroxybenzoic acid and syringaldehyde.
4. Discussion
Small amounts of SO2 have a good inhibitory effect on microorganisms which are undesirable in wine production. The wine yeasts S. cerevisiae, S. bayanus and most of their hybrids have a relatively high insensitivity to this preservative [21]. The use of wine phenols as a SO2 substitute thus presupposes that the yeasts are not influenced by these compounds in their fermentation activity, and only undesired organisms are inhibited.
In the present work, water-insoluble phenols were dissolved in DMSO. EUCAST (European Committee on Antimicrobial Susceptibility Testing) and CLSI (Clinical and Laboratory Standard Institute) recommend 1% v/v DMSO in the medium in inhibition experiments to exclude any solvent effect. On the other hand, Sardowska-Bartozs et al. [19] found that Saccharomycetes are only inhibited >8% v/v DMSO. We found impaired growth of S. bayanus HL77 at 5% v/v DMSO and decided to perform the inhibition experiments at 5% and 2.5% DMSO in the medium. The main results of this work are summarized in Table 3.
4.1. Influence of Phenolic Compounds on Yeast Growth
Examination of the inhibitory effect at culture medium pH 3.5 revealed that non-Saccharomycetes are sensitive to ferulic acid, p-coumaric acid and syringaldehyde. In the case of the Saccharomyces species, differences in the inhibitory effect were detected. Whereas S. cerevisiae 16.1 showed complete inhibition only with 1 mg/mL of ferulic acid or resveratrol, the hybrid HL 78 and S. bayanus HL77 revealed marked sensitivities to phenols. Again, p-coumaric acid, ferulic acid and syringaldehyde acted inhibitory. In the experiments conducted at pH 5.5, the negative effect of the cinnamic acids were usually diminished, probably as they are in their deprotonated state (see Section 4.4).
Ferulic acid turned out as the phenolic acid with the strongest inhibitory effect. With p-coumaric acid, a good inhibitory effect could also be achieved in some cases. This is consistent with the work of Pastorkova et al. [12], who also observed a high sensitivity of wine-damaging yeasts for hydroxycinnamic acids. Harris et al. [22] ruled out the possible application of ferulic acid as an antifungal agent for the control of Dekkera spp.
Syringaldehyde gave surprisingly good inhibition results. The compound is one of the less-explored polyphenols and, so far, there is little information on its antimicrobial activity, as well as its other capacities [23]. Studies with the pathogenic yeast Candida guilliermondii showed a fungicidal effect of syringaldehyde [24]. In general, it can be stated that the so-called wild yeasts were more strongly inhibited than the Saccharomycetes with syringaldehyde. This observation suggests that it is one of the compounds that could have the potential to be used as SO2 substitutes. This should be given greater attention in the future.
Resveratrol had an inhibitory effect on most yeast species under investigation including S. cerevisiae. This seems in some contrast to studies which revealed that the stilbene increases the live expectancy of this most important wine yeast [9].
4.2. Influence of Phenolic Compounds on the Growth of Lactic Acid Bacteria
Phenolic compounds impaired growth of lactic acid bacteria in a concentration-dependent manner. The inhibition effect increased generally in the order sinapic acid < caffeic acid < p-coumaric acid < ferulic acid < syringaldehyde < resveratrol. Less effective were ethylgallate and 3,4-dihydroxy benzoic acid. Polydatin behaved rather neutral, and gallic acid had even a growth stimulating effect on P. parvulus and O. oeni. Comparable results have been reported in previous investigations. Stead [25] found that high concentrations of p-coumaric, ferulic and caffeic acids inhibited the growth of Lactobacillus collinoides and Lactobacillus brevis, whereas low concentrations stimulated their growth. In another study, Rozès and Peres [26] observed that caffeic and ferulic acids negatively affected the growth of L. plantarum and increased the proportion of unsaturated fatty acids of the cell membrane. Salih et al. [27] studied the inhibitory effect of hydroxycinnamic acids towards O. oeni and L. plantarum and found a strong decrease in the growth rate and biomass production in O. oeni but only a growth rate decrease was observed in L. plantarum. Ferulic acid had the strongest effect, followed by p-coumaric and caffeic acids. Campos et al. [28] reported that hydroxycinnamic acids (and particularly p-coumaric acid) were more inhibitory towards O. oeni and L. hilgardii than hydroxybenzoic acids; on the other hand, some phenolic acids showed a beneficial effect on growth of L. hilgardii [28].
Cinnamic acids can also influence malolactic fermentation of lactic acid bacteria: 50–150 µg/mL caffeic acid supported the degradation of malic acid in Merlot wine, whereas ferulic acid was inhibitory; coumaric acid had even a more negative effect [9].
In line with our results, several authors reported a positive effect of gallic acid on the growth of lactic acid bacteria [29,30]. Alberto et al. [31] found that low concentrations (up to 200 µg/mL) of gallic acid stimulated growth of L. hilgardii, whereas at high concentrations (1000 µg/mL) gallic acid was inhibitory to this bacterium. In later works, the same strain of L. hilgardii was shown to be able to metabolise gallic acid, producing some phenolic compounds with oxygen-scavenging capacity [32]. Similarly, Theobald et al. [33] reported that epigallocatechine gallate (EGCG) exerts a concentration-dependent impact on the growth of the wine bacterium Oenococcus oeni strain B2. It was shown that EGCG had a stimulating effect at 400–500 µg mL−1 but an inhibitory effect when present above 540 µg mL−1 in the culture media. In relation to this, other authors observed that flavanols with galloyl moiety exhibited more activity on bacteria growth than those without such residues [34].
Landete et al. [35] reported that L. plantarum was more sensitive to hydroxycinnamic acids (especially p-coumaric acid) than to hydroxybenzoic acids at the same molar concentrations. García-Ruiz et al. [10] observed that stilbenes had a stronger inhibitory effect towards wine lactic acid bacteria than phenolic acids at the same concentration levels.
4.3. Influence of Phenolic Compounds on the Growth of Acetic Acid Bacteria
Acetic acid bacteria are obligate aerobic microorganisms which are able to oxidize numerous sugars, sugar alcohols and alcohols by the aid of various pyrroloquinoline quinone (PQQ)-dependent membrane-bound dehydrogenases. In wine, they are notorious spoilage microorganisms by the conversion of ethanol into acetic acid [1,36].
At 1 mg/mL syringaldehyde, we found a permanent growth reduction of A. acetii 3508 and G. cerinus 9533, whereas resveratrol and ferulic acid only inhibited A. acetii 3508. The same strain was inhibited by ferulic acid and resveratrol in investigations of Pastorkowa et al. [12]. Syringaldehyde was not included their study. There are only a few other publications dealing with the effect of phenolic compounds on the growth of wine-related acetic acid bacteria. García-Ruiz et al. [11] found that some phenolic plant extracts inhibited growth of A. aceti and Gluconobacter oxydans.
In our study, we observed that acetic acid bacteria transformed phenols into brownish products. This is in line with a study of Buchert and Niemelä [37], who investigated the effect of some aromatic aldehydes on the growth of G. oxydans. The organism detoxified all possible inhibitors except syringaldehyde by simultaneously oxidizing and reducing them to the corresponding acids and alcohols.
4.4. Mode of Antimicrobial Action
Besides differing in structure and complexity, phenolic compounds also exhibit different properties regarding their antimicrobial activities. Benzoic acid was one of the first authorized preservatives for use in the food industry to attain Generally Regarded As Safe (GRAS) status [2]. Although the precise antimicrobial mechanism of phenolic acids is not well known, there is growing evidence that the primary effect is to interfere with the cytoplasmic membrane, increasing its permeability and causing leakage of intracellular constituents such as proteins, nucleic acids, and inorganic ions. They thereby act as decoupling agents leading to a disruption of the transmembrane pH and charge gradient [38]. Phenolic acids are weak organic acids (pKa ≈ 4.2) and their antimicrobial activity is considered to be dependent on the concentration of the nondissociated carboxyl group. Due to their lipophilic nature, they can cross the microbial cell membrane in their non-ionic form and acidify the cytosol, causing protein denaturation and interfering with cellular activity [2]. Destruction of the cell wall structure is another bactericidal effect of phenolic compounds as observed for lactic acid bacteria [39]. Gram-negative bacteria are more resistant to phenolic compounds than Gram-positive bacteria due to the presence of their protective outer membrane [2].
Benzaldehydes act primarily by combining with sulfhydryl groups of proteins on the external surface of microorganisms. Other works have shown that these compounds can also cause cellular leakage [2].
The hydroxystilbene resveratrol is one of the best analyzed compounds in wine, especially because of its beneficial influence on human health [40,41]. Being a phytoalexin, the compound is produced by plants in response to biotic and abiotic stress factors and has antifungal and antimicrobial activities. Its glycosylated derivate (polydatin or piceid) occurs in red wines as much as tenfold higher. The antibiotic action mechanism of stilbenes is not well understood. It has been suggested that resveratrol inhibits the respiration of fungal cells, probably by acting as an uncoupling agent or by membrane lipid peroxidation [20]. Pterostilbene is a methoxylated derivate of resveratrol originating from natural sources like wine grapes or blue berries. Its potent capability against methicillin-resistant Staphylococcus aureus (MRSA) has been related to bacterial membrane leakage, chaperone protein downregulation, and ribosomal protein upregulation [42].
4.5. Enyzmatic Oxidations
Phenolic compounds of grapes can be rapidly transformed by phenoloxidases, thereby altering their biological properties [18]. Here we found that the antimicrobial action of phenols mostly decreased after laccase oxidation, but also the opposite situation. This may be explained by a different biochemical reactivity of the oxidation products. It is generally believed that fungal laccases as such from Botrytis cinerea detoxify phenolic compounds (e.g., the phytoalexin resveratrol) by oxidative transformation to di-, oligo- or polymers which are less bioavailable and bioactive [20]. On the other hand, the first enzymatic oxidation products from polyphenolics are high-reactive quinones which can rapidly react with amino acids, peptides and proteins by Michael-type additions [43]. In the case of enzymes this will lead to their inhibition [9,44].
5. Conclusions
The main subject of this work was to assess the different inhibition behaviors of phenols against wine-associated microorganisms and factors influencing the inhibitory effect, e.g., enzymatic oxidations, pH and organic solvents. The focus was on the evaluation of the individual phenols as a potential sulfite substitute. The most promising compounds were ferulic acid, resveratrol and syringaldehyde. Low concentrations of ferulic acids (250 µg/mL) inhibited potential wine-spoiling bacteria but not growth of wine yeasts. The biotechnological potential of ferulic acid for medical and food applications has already been highlighted [45].
Although antifungal activities of resveratrol are well known, its strong inhibition effect on bacteria and yeasts is somewhat surprising but confirms previous studies [10]. Considering the technological application of resveratrol in the process of wine stabilization, its concentration in wine is too low to effectively inhibit spoilage microorganisms (Table 1). Therefore, wine fortification by addition of pure stilbenes or plant extracts might be considered as prospective strategy [12]. Glycosylated resveratrol (polydatin) was ineffective in our study, whereas another derivate, pterostilbene, displayed up to four times stronger antimicrobial efficacy than resveratrol [12]. Another possibility to increase the trans-resveratrol content in wines is the enzymatic hydrolysis of glycosylated precursors by application of β-glucosidases [46].
A main conclusion which can be drawn from our experiments is that among the phenols tested, syringaldehyde seems to have the highest biotechnological potential, since it acted at relatively low concentrations, especially on the wild yeasts and bacteria. Syringaldehyde as antimicrobial agent has been given little attention so far. This gap should be closed in the future, especially since this phenol also has health-promoting and antioxidant properties [23].
Another point that should be taken into account in the assessment of the inhibitory capacity of phenols are their interactions with each other and with other wine constituents. For example, the combination of different phenols may have a positive or negative effect on the inhibitory effect. Also, the influence of phenoloxidases is not insignificant, as demonstrated in this work. The investigation of these interactions is indispensable in order to be able to make a precise statement about the inhibitory behavior of the phenols. Finally, the influence of antimicrobial plant extracts on the composition and sensorial quality of wines needs to be considered [47]. Particularly, in the case of syringaldehyde the sensory threshold in wine for this compound has been reported at 50 µg/mL, thus at a significant lower level than the lowest concentration (250 µg/mL) tested in this study. The actual minimum antimicrobial concentrations of syringaldehyde will be evaluated in consecutive research.
Acknowledgments
This research project was supported by the Stiftung Rheinland-Pfalz für Innovation (Germany) Project number 961-386261/1051. The authors want to dedicate this article to Prof. Dr. Helmut König.
Author Contributions
Andrea Sabel, Simone Bredefeld and Martina Schlander performed the experiments and analyzed the data. Harald Claus conceived the study and wrote the paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- König, H.; Unden, F.; Fröhlich, J. Biology of Microorganisms on Grapes, in Must and in Wine; Springer: Berlin/Heidelberg, Germany, 2009. [Google Scholar]
- Campos, F.M.; Couto, J.A.; Hogg, T. Utilisation of natural and by-products to improve wine safety. In Wine Safety, Consumer Preference, and Human Health; Moreno-Arribas, M.V., Bartolomé Sualdea, B., Eds.; Springer Int. Publ.: Cham, Switzerland, 2016; pp. 27–49. [Google Scholar]
- Pozo-Bayón, M.A.; Monagas, M.; Bartolomé, B.; Moreno-Arribas, M.V. Wine features related to safety and consumer health: An integrated perspective. Crit. Rev. Food Sci. Nutr. 2012, 52, 31–54. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulou, C.; Soulti, K.; Roussis, I.G. Potential antimicrobial activity of red and white wine phenolic extracts against strains of Staphylococcus aureus, Escherichia coli and Candida albicans. Food Technol. Biotechnol. 2005, 43, 41–46. [Google Scholar]
- Cueva, C.; Mingo, S.; Muňoz-González, I.; Bustos, I.; Requena, T.; Del Campo, R.; Martín-Álvarez, P.J.; Bartolomé, B.; Moreno-Arribas, M.V. Antibacterial activity of wine phenolic compounds and oenological extracts against potential respiratory pathogens. Lett. Appl. Microbiol. 2012, 54, 557–563. [Google Scholar] [CrossRef] [PubMed]
- Katalinic, V.; Smole Mozina, S.; Generalic, I.; Skroza, D.; Ljubenkov, I.; Klancik, A. Phenolic profile, antioxidant capacity, and antimicrobial activity of leaf extracts from six Vitis. vinifera L. varieties. Int. J. Food Prop. 2013, 16, 45–60. [Google Scholar] [CrossRef]
- Xu, C.; Yagiz, Y.; Zhao, L.; Simonne, A.; Lu, J.; Marshall, M.R. Fruit quality, nutraceutical and antimicrobial properties of 58 muscadine grape varieties (Vitis. rotindifolia Michx.) grown in United States. Food Chem. 2017, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Landete, J.M. Updated knowledge about polyphenols: Functions, bioavailability, metabolism, and health. Crit. Rev. Food Sci. Nutr. 2012, 52, 936–948. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, H.; Pour-Nikfardjam, M.S. Influence of phenolic compounds and tannins on wine-related microorganisms. In Biology of Microorganisms on Grapes, in Must and in Wine; König, H., Unden, F., Fröhlich, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 307–334. [Google Scholar]
- García-Ruiz, A.; Moreno-Arribas, M.V.; Martín-Álvarez, P.J.; Bartolomé, B. Comparative study of the inhibitory effects of wine polyphenols on the growth of enological lactic acid bacteria. Int. J. Food Microbiol. 2011, 145, 426–431. [Google Scholar] [CrossRef] [PubMed]
- García-Ruiz, A.; Cueva, C.; Gonzales-Rampinelli, E.M.; Yuste, M.; Torres, M.; Martin-Alvarez, P.J.; Bartolomé, B.; Moreno-Arribas, M.V. Antimicrobial phenolic extracts able to inhibit lactic acid bacteria growth. Food Control 2012, 28, 212–219. [Google Scholar] [CrossRef]
- Pastorkowa, E.; Zakova, T.; Landa, P.; Navakova, J.; Vadlejch, J.; Kokoska, L. Growth inhibitory effect of grape phenolics against wine spoilage yeast and acetic acid bacteria. Int. J. Food Microb. 2013, 161, 209–213. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Rompinelli, E.M.; Rodriguez-Bencomo, J.J.; Garcia-Ruiz, A.; Sanchez-Patan, F.; Martin-Alvarez, P.J.; Bartolome, B.; Morena-Arribas, M. A winery-scale trial of the use of antimicrobial plant phenolic extracts as preservatives during wine ageing in barrels. Food Control 2013, 33, 440–447. [Google Scholar] [CrossRef]
- Sabel, A.; Martens, S.; Petri, A.; König, H.; Claus, H. Wickerhamomyces anomalus AS1: A new strain with potential to improve wine aroma. Ann. Microbiol. 2014, 64, 483–491. [Google Scholar] [CrossRef]
- Christ, E.; Kowalczyk, M.; Zuchowska, M.; Claus, H.; Löwenstein, R.; Szopinska-Morawska, A.; Renaut, J.; König, H. An exemplary model study for overcoming stuck fermentation during spontaneous fermentation with the aid of a Saccharomyces triple hybrid. J. Agric. Sci. 2015, 7, 18–34. [Google Scholar] [CrossRef]
- Zuchowska, M.; Jaenicke, E.; König, H.; Claus, H. Allelic variants of hexose transporter Hxt3p and hexokinases Hxk1p/Hxk2p in strains of Saccharomyces cerevisiae and interspecies hybrids. Yeast 2015, 32, 657–669. [Google Scholar] [CrossRef] [PubMed]
- Bäumlisberger, M.; Moellecken, U.; König, H.; Claus, H. The potential of the yeast Debaryomyces hansenii H525 to degrade biogenic amines in food. Microorganisms 2015, 3, 839–850. [Google Scholar] [CrossRef] [PubMed]
- Riebel, M.; Sabel, A.; Claus, H.; Xia, N.; Li, H.; König, H.; Decker, H.; Fronk, P. Antioxidant capacity of phenolic compounds on human cell lines as affected by grape-tyrosinase and Botrytis-laccase oxidation. Food Chem. 2017, 229, 779–789. [Google Scholar] [CrossRef] [PubMed]
- Sadowska-Bartosz, I.; Pączka, A.; Molon, M.; Bartosz, G. Dimethyl sulfoxid induces oxidative stress in the yeast Saccharomyces cerevisiae. FEMS Yeast Res. 2013, 13, 820–830. [Google Scholar] [CrossRef] [PubMed]
- Claus, H.; Sabel, A.; König, H. Wine Phenols and Laccase: An ambivalent relationship. In Wine Phenolic Composition, Classification and Health Benefits; El Rayess, E.Y., Ed.; Nova publishers: New York, NY, USA, 2014; pp. 155–185. [Google Scholar]
- Dittrich, H.H.; Großmann, M. Mikrobiologie des Weines 4. Auflage; Verlag Eugen Ulmer: Stuttgart, Germany, 2011. [Google Scholar]
- Harris, V.; Jiranek, V.; Ford, C.M.; Grbin, P.R. Inhibitory effect of hydroxycinnamic acids on Dekkera spp. Appl. Microbiol. Biotechnol. 2010, 86, 721–729. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, M.N.M.; Sriprasanthi, R.B.; Shamsudeen, S.; Adam, F.; Bhawani, S.A. A concise review of the natural existence, synthesis, properties and applications of syringaldehyde. BioResources 2012, 7, 4377–4399. [Google Scholar]
- Kelly, C.; Jones, O.; Barnhart, C.; Lajoie, C. Effect of furfural, vanillin and syringaldehyde on Candida guilliermondii: Growth and xylitol biosynthesis. Appl. Biochem. Biotechnol. 2008, 148, 97–108. [Google Scholar] [CrossRef] [PubMed]
- Stead, D. The effect of hydroxycinnamic acids on the growth of wine-spoilage lactic acid bacteria. J. Appl. Bact. 1993, 75, 135–141. [Google Scholar] [CrossRef]
- Rozès, N.; Peres, C. Effects of phenolic compounds on the growth and fatty acid composition of Lactobacillus plantarum. Appl. Microbiol. Biotechnol. 1998, 49, 108–111. [Google Scholar] [CrossRef]
- Salih, A.G.; Le Quéré, J.M.; Drilleau, J.F. Effect of hydroxycinnamic acids on the growth of lactic bacteria. Sci. Aliment. 2000, 20, 537–560. [Google Scholar] [CrossRef]
- Campos, F.M.; Couto, J.A.; Hogg, T. Influence of phenolic acids on growth and inactivation of Oenococcus oeni and Lactobacillus hilgardii. J. Appl. Microbiol. 2003, 94, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Stead, D. The effect of chlorogenic, gallic and quinic acids on the growth of spoilage strains of Lactobacillus collinoides and Lactobacillus brevis. Lett. Appl. Microbiol. 1994, 112–114. [Google Scholar] [CrossRef]
- Vivas, N.; Lonvaud-Funel, A.; Glories, Y. Effect of phenolic acids and anthoyanins on growth, viability and malolactic activity of a lactic acid bacterium. Food Microbiol. 1997, 291–299. [Google Scholar] [CrossRef]
- Alberto, M.R.; Farías, M.E.; Manca de Nadra, M.C. Effect of gallic acid and catechin on Lactobacillus hilgardii 5w growth and metabolism of organic compounds. J. Agric. Food Chem. 2001, 49, 4359–4363. [Google Scholar] [CrossRef] [PubMed]
- Alberto, M.R.; Gómez-Cordovés, C.; Manca de Nadra, M.C. Metabolism of gallic acid and catechin on Lactobacillus hilgardii from wine. J. Agric. Food Chem. 2004, 52, 6465–6469. [Google Scholar] [CrossRef] [PubMed]
- Theobald, S.; Pfeiffer, P.; Zuber, U.; König, H. Influence of epigallocatchin gallate and phenolic compounds from green tea on the growth of Oenococcus oeni. J. Appl. Microbiol. 2008, 104, 566–672. [Google Scholar] [CrossRef] [PubMed]
- González de Llano, D.; Gil-Sánchez, I.; Esteban-Fernández, A.; Ramos, A.M.; Cueva, C.; Moreno-Arribas, M.V.; Bartolomé, B. Some contributions to the study of oenological lactic acid bacteria through their interaction with polyphenols. Beverages 2016, 2, 27. [Google Scholar] [CrossRef]
- Landete, J.M.; Rodríguez, H.; de las Rivas, B.; Munoz, R. High-added-value antioxidants obtained from the degradation of wine phenolic by Lactobacillus plantarum. J. Food. Prot. 2007, 70, 2670–2675. [Google Scholar] [CrossRef] [PubMed]
- Guillamón, J.M.; Mas, A. Acetic acid bacteria. In Molecular Wine Microbiology; Carrascosa, A.V., Muñoz, R., González, R., Eds.; Elsevier: London, UK, 2011; pp. 227–255. [Google Scholar]
- Buchert, J.; Niemelä, K. Oxidative detoxification of wood-derived inhibitors by Gluconobacter oxydans. J. Biotechnol. 1991, 18, 1–12. [Google Scholar] [CrossRef]
- Amborabé, B.E.; Fleurat-Lessard, P.; Chollet, J.F.; Roblin, G. Antifungal effects of salicylic acid and other benzoic derivates towards Eutypa lata: Structure-activity relationship. Plant Phys. Biochem. 2002, 40, 1051–1060. [Google Scholar] [CrossRef]
- Rodríguez, H.; Curiel, J.A.; Landete, J.M.; de las Rivas, B.; de Filipe, F.L.; Gómez-Cordovés, C.; Mancheño, J.M.; Muñoz, R. Food phenolics and lactic acid bacteria. Int. J. Food. Microbiol. 2009, 132, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Suart, J.A.; Robb, E.A. Bioactive Polyphenols from Wine Grapes; Springer: New York, NY, USA, 2013. [Google Scholar]
- El Rayess, Y. Wine–Phenolic Composition, Classification and Health Benefits; Nova Publishers: New York, NY, USA, 2014. [Google Scholar]
- Yang, S.C.; Tseng, C.H.; Wang, P.W.; Lu, P.L.; Weng, Y.H.; Yen, F.L.; Fang, J.Y. Pterostilbene, a methoxylated resveratrol derivate, efficiently eradicates planctonic, biofilm, and intracellular MRSA by topical application. Front. Microbiol. 2017, 8, 1103. [Google Scholar] [CrossRef]
- Fernandes, M.S.; Kerkar, S. Microorganisms as a source of tyrosinase inhibitors: A review. Ann. Microbiol. 2017, 67, 343–358. [Google Scholar] [CrossRef]
- Pourcel, L.; Routaboul, J.M.; Cheynier, V.; Lepiniec, L.; Debeaujon, I. Flavanoid oxidation in plants: From biochemical properties to physiological functions. Trends Plant Sci. 2006, 12, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Pruthi, V. Potential applications of ferulic acid from natural sources. Biotechnol. Rep. 2014, 4, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Schwentke, J.; Sabel, A.; Petri, A.; König, H.; Claus, H. The yeast Wickerhamomyces. anomalus AS1 secretes a multifunctional exo-β-1,3-glucanase with implications for winemaking. Yeast 2014, 31, 349–359. [Google Scholar] [CrossRef] [PubMed]
- García-Ruiz, A.; Rodríguez-Bencomo, J.J.; Garrido, I.; Martín-Álvarez, M.; Moreno-Arribas, M.V.; Bartolomé, B. Assessment of the impact of the additions of antimicrobial plant extracts to wine: Volatile and phenolic composition. J. Sci. Food Agric. 2013, 93, 2507–2516. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Influence of ethanol and DMSO on the growth of (A): S. cerevisiae 16.1; (B): S. bayanus HL 77; (C): S. cerevisiae × S. kudriavzevii × S. bayanus HL 78. Culture medium: Sabouraud-glucose broth (pH 3.5).
Figure 1.
Influence of ethanol and DMSO on the growth of (A): S. cerevisiae 16.1; (B): S. bayanus HL 77; (C): S. cerevisiae × S. kudriavzevii × S. bayanus HL 78. Culture medium: Sabouraud-glucose broth (pH 3.5).

Figure 2.
Influence of phenolic compounds on the growth of Saccharomyces cerevisiae 16.1 (A), Saccharomyes bayanus HL 77 (B); Saccharomyces cerevisiae × S. kudriavzevii × S. bayanus HL 78 (C), Wickerhamomyces anomalus AS1 (D) and Debaryomyces hansenii 525 (E). The red line indicates maximum growth (optical density) of the controls which contained DMSO (5.0 % v/v) but no phenols. Culture medium: Sabouraud-2% Glucose both (pH 3.5).
Figure 2.
Influence of phenolic compounds on the growth of Saccharomyces cerevisiae 16.1 (A), Saccharomyes bayanus HL 77 (B); Saccharomyces cerevisiae × S. kudriavzevii × S. bayanus HL 78 (C), Wickerhamomyces anomalus AS1 (D) and Debaryomyces hansenii 525 (E). The red line indicates maximum growth (optical density) of the controls which contained DMSO (5.0 % v/v) but no phenols. Culture medium: Sabouraud-2% Glucose both (pH 3.5).

Figure 3.
Influence of phenolic compounds on the growth of S. cerevisiae 16.1 (A) 16.1, S. bayanus HL77 (B), W. anomalus AS1 (C) and D. hansenii 525 (D). The red line indicates maximum growth (optical density) of the controls which contained DMSO (2.5% v/v) but no phenols. Culture medium: Sabouraud-glucose broth (pH 5.5).
Figure 3.
Influence of phenolic compounds on the growth of S. cerevisiae 16.1 (A) 16.1, S. bayanus HL77 (B), W. anomalus AS1 (C) and D. hansenii 525 (D). The red line indicates maximum growth (optical density) of the controls which contained DMSO (2.5% v/v) but no phenols. Culture medium: Sabouraud-glucose broth (pH 5.5).

Figure 4.
Influence of phenolic compounds on the growth of Lactobacillus hilgardii 20176 (A), Lactobacillus plantarum 20174T (B), Pediococcus parvulus 20332T (C) and Oenococcus oeni 20252 (D). The red line indicates maximum growth (optical density) of the controls which contained DMSO (2.5% v/v) but no phenols. Culture medium: MRS (pH 5.5).
Figure 4.
Influence of phenolic compounds on the growth of Lactobacillus hilgardii 20176 (A), Lactobacillus plantarum 20174T (B), Pediococcus parvulus 20332T (C) and Oenococcus oeni 20252 (D). The red line indicates maximum growth (optical density) of the controls which contained DMSO (2.5% v/v) but no phenols. Culture medium: MRS (pH 5.5).

Figure 5.
Influence of phenolic compounds on the growth of Gluconobacter cerinus 9533 after 24 h (A) or 48 h (B) and on Acetobacter acetii 3508 after 48 h (C). The red line indicates maximum growth (optical density) of the controls which contained DMSO (2.5% v/v) but no phenols. Culture medium YPM (pH 6.1) for (A) and (B) and YPS (pH 6.6) for (C).
Figure 5.
Influence of phenolic compounds on the growth of Gluconobacter cerinus 9533 after 24 h (A) or 48 h (B) and on Acetobacter acetii 3508 after 48 h (C). The red line indicates maximum growth (optical density) of the controls which contained DMSO (2.5% v/v) but no phenols. Culture medium YPM (pH 6.1) for (A) and (B) and YPS (pH 6.6) for (C).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Phenolic compounds included in this study and their natural concentrations in wine.
| Chemical Class | Representative Structures | Examples | Mean Contents [µg/mL] 1 in | |
|---|---|---|---|---|
| Red Wine | White Wine | |||
| Hydroxybenzoic acids |  | Gallic acid | 35.9 | 2.2 |
| Vanillic acid | 3.2 | 0.4 | ||
| 4-Hydroxybenzoic acid | 5.5 | 0.2 | ||
| Syringic acid | 2.7 | <0.01 | ||
| Ethylgallate | 15.3 | nd | ||
| Cinnamic acids |  | p-Coumaric acid | 5.5 | 1.5 |
| Caffeic acid | 18.8 | 2.4 | ||
| Ferulic acid | 0.8 | 0.9 | ||
| Sinapic acid | 0.7 | 0.6 | ||
| Stilbenes |  | cis-Resveratrol | 1.3 | 0.2 |
| Trans-Resveratrol | 1.8 | 0.4 | ||
| trans-Resveratrol-3-O-glucoside | 4.1 | 1.7 | ||
| Hydroxybenz-aldehydes |  | Syringaldehyde | 6.6 | <0.01 |
1 Source: http://phenol-explorer.eu; nd: no data.
Table 2.
Effect of B. cinerea laccase on the growth effect of phenols 1.
| Species | Caffeic Acid | Gallic Acid | Resveratrol | Polydatin | 3,4-di-HB | Ferulic Acid | Sinapic Acid | Syring-Aldehyde | Ethyl-gallate |
|---|---|---|---|---|---|---|---|---|---|
| S. cerevisiae | +4.29 | −3.98 | +2.30 | −103.00 | +3.94 | +5.51 | −9.37 | +9.63 | +4.36 |
| S. bayanus | +24.23 | +7.77 | ±0 | −106.00 | −3.72 | +3.98 | −7.00 | +2.07 | −5.77 |
| D. hansenii | +7.11 | −15.58 | −4.74 | −14.29 | −19.94 | +1.04 | −4.12 | −8.75 | −32.30 |
| W. anomalus | +9.19 | −0.89 | +6.17 | −88.13 | −9.36 | −4.98 | −7.12 | −1.58 | −5.37 |
| L. hilgardii | +5.01 | +3.71 | −8.37 | +15.57 | +7.99 | +1.49 | +21.93 | +0.49 | +3.29 |
| L. plantarum | +10.00 | +2.36 | −4.41 | −11.17 | −7.94 | +8.77 | +23.12 | −2.14 | +5.06 |
| P. parvulus | +1.51 | +8.16 | ± 0 | +0.68 | +16.01 | +8.34 | +37.18 | −31.65 | +29.22 |
| O. oeni | +19.21 | +115.74 | +9.79 | +40.63 | +47.38 | +12.20 | +25.37 | +3.01 | +30.15 |
1 data are given in percentage positive (+) or negative (−) growth compared to the samples without laccase additions; initial concentrations of phenols were 1 mg/mL.
Table 3.
Minimum inhibitory concentration [µg/mL] of phenolic compounds on wine-related microorganisms.
Table 3.
Minimum inhibitory concentration [µg/mL] of phenolic compounds on wine-related microorganisms.
| (A) Yeasts 1 | |||||
| Compound | S. cerevisiae | S. bayanus | S. c. × S. k. × S. b. | W. anomalus | D. hansenii |
| Caffeic acid | >1000 | >1000 | nd | >1000 | >1000 |
| p-Coumaric acid | >1000 | 500 | 1000 | 1000 | >1000 |
| Ferulic acid | 1000 | 250–500 | 1000 | 1000 | 500 |
| Sinapic acid | >1000 | >1000 | nd | >1000 | 1000 |
| Gallic acid | >1000 | >1000 | 1000 | >1000 | >1000 |
| Vanillic acid | >1000 | 1000 | 1000 | >1000 | >1000 |
| Hydroxybenzoic acid | >1000 | >1000 | nd | >1000 | 1000 |
| Syringaldehyde | 250–1000 | 250–500 | 250 | 250 | 250–1000 |
| Resveratrol | 250 | 250 | 250 | 250–500 | 250 |
| Polydatin | >1000 | >1000 | nd | >1000 | >1000 |
| Potassium sulfite | 250–500 | 500–1000 | 500 | 250 | 250 |
| (B) Lactic Acid Bacteria | |||||
| Compound | L. hilgardii | L. plantarum | P. parvulus | O. oeni | |
| Caffeic acid | 250 | >1000 | 250 | 250 | |
| Ferulic acid | 250 | 250 | 250 | 250 | |
| Sinapic acid | 250 | 250 | 250 | 250 | |
| Gallic acid | >1000 | >1000 | >1000 | >1000 | |
| Hydroxybenzoic acid | 250 | >1000 | 1000 | 500 | |
| Syringaldehyde | 250 | 250 | 1000 | 250 | |
| Resveratrol | 250 | 250 | 250 | 250 | |
| Polydatin | >1000 | >1000 | >1000 | >1000 | |
| Potassium sulfite | 250 | 250 | 250 | 250 | |
| (C) Acetic Acid Bacteria | |||||
| Compound | G. cerinus | A. acetii | |||
| Ferulic acid | >1000 | 1000 | |||
| Sinapic acid | >1000 | >1000 | |||
| Gallic acid | >1000 | 1000 | |||
| Syringaldehyde | 250 | 1000 | |||
| Resveratrol | >1000 | 250 | |||
| Potassium sulfite | 250 | 250 | |||
1 results from experimental series 1 and 2; nd: not determined S. c. × S. k. × S. b. = S. cerevisiae × S. kudriavzevii × S. bayanus.
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Sabel, A.; Bredefeld, S.; Schlander, M.; Claus, H. Wine Phenolic Compounds: Antimicrobial Properties against Yeasts, Lactic Acid and Acetic Acid Bacteria. Beverages 2017, 3, 29. https://0-doi-org.brum.beds.ac.uk/10.3390/beverages3030029
AMA Style
Sabel A, Bredefeld S, Schlander M, Claus H. Wine Phenolic Compounds: Antimicrobial Properties against Yeasts, Lactic Acid and Acetic Acid Bacteria. Beverages. 2017; 3(3):29. https://0-doi-org.brum.beds.ac.uk/10.3390/beverages3030029
Chicago/Turabian StyleSabel, Andrea, Simone Bredefeld, Martina Schlander, and Harald Claus. 2017. "Wine Phenolic Compounds: Antimicrobial Properties against Yeasts, Lactic Acid and Acetic Acid Bacteria" Beverages 3, no. 3: 29. https://0-doi-org.brum.beds.ac.uk/10.3390/beverages3030029
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.