Natural Nrf2 Activators from Juices, Wines, Coffee, and Cocoa


1
Department of Pharmaceutical Sciences, Daniel K. Inouye College of Pharmacy, University of Hawaii at Hilo, 200 West Kawili Street, Hilo, HI 96720, USA
2
Jing Brand Research Institute, Jing Brand Co., Ltd., Daye 435100, China
*
Author to whom correspondence should be addressed.
Beverages 2020, 6(4), 68; https://0-doi-org.brum.beds.ac.uk/10.3390/beverages6040068
Submission received: 17 September 2020
/
Revised: 1 November 2020
/
Accepted: 9 November 2020
/
Published: 1 December 2020
(This article belongs to the Special Issue Phenolic Compounds and Functional Beverages)
Abstract
:Juices, wine, coffee, and cocoa are rich sources of natural polyphenolic compounds that have potent antioxidant activities proven by in vitro and in vivo studies. These polyphenolic compounds quench reactive oxygen and nitrogen species (RONS) or reactive free radicals and act as natural antioxidants which are also able to protect against reactive oxygen species (ROS)-mediated oxidative damage, which elevates cellular antioxidant capacity to induce antioxidant defense mechanisms by modulating transcription factors. Nuclear factor (erythroid-derived 2)-like 2 (Nrf2) is a transcription factor encoded in humans. It is activated as a result of oxidative stress and induces the expression of its target genes. This is one of the most important cellular defense mechanisms against oxidative stress. However, the oxidative stress alone is not enough to activate Nrf2. Hence phytochemicals, especially polyphenolics, act as natural Nrf2 activators. Herein, this review discusses the natural products identified in juices, coffee, cocoa and wines that modulate Nrf2 activity in cellular systems.
1. Introduction
In biological systems, the balance between the generation and removal of oxidants is required to restore oxidative balance. If there is an overproduction or unbalanced formation of these oxidants, this triggers oxidative stress. These oxidants are free radicals of atoms or molecules with an unpaired single electron which is highly reactive [1]. These highly reactive species could be categorized into reactive oxygen species (ROS) and reactive nitrogen species (RNS). These oxygen or nitrogen containing molecules have one or more unpaired electrons in their molecular or atomic orbitals [2]. The hydroxyl radical (•OH) is the main ROS in biological systems, whose reactivity is much higher than other ROS like superoxide (O2•−) and hydrogen peroxide (H2O2), thus reacting quickly at the site or very close to the site where it forms. RNS is generated when nitric oxide (NO) interacts with reactive oxygen species like superoxide (O2•−) and hydrogen peroxide (H2O2). The nitrogen containing reactive species, nitric oxide (NO•), is relatively unreactive, while peroxy-nitrite (ONOO−) is a powerful oxidant able to damage many biological molecules [3,4,5]. Nitric oxide can also act as an electrophile to react with nucleophiles like thiols and amines in the human body. Not only free radicals, but non-radical species (for example, hydrogen peroxide (H2O2), hypochlorous acid (HOCl), and hypo-bromous acid (HOBr) [6]) are also powerful oxidizing agents in biological systems, actively participating in free radical reactions. Possible ROS or RNS found in biological systems are listed below in Table 1.
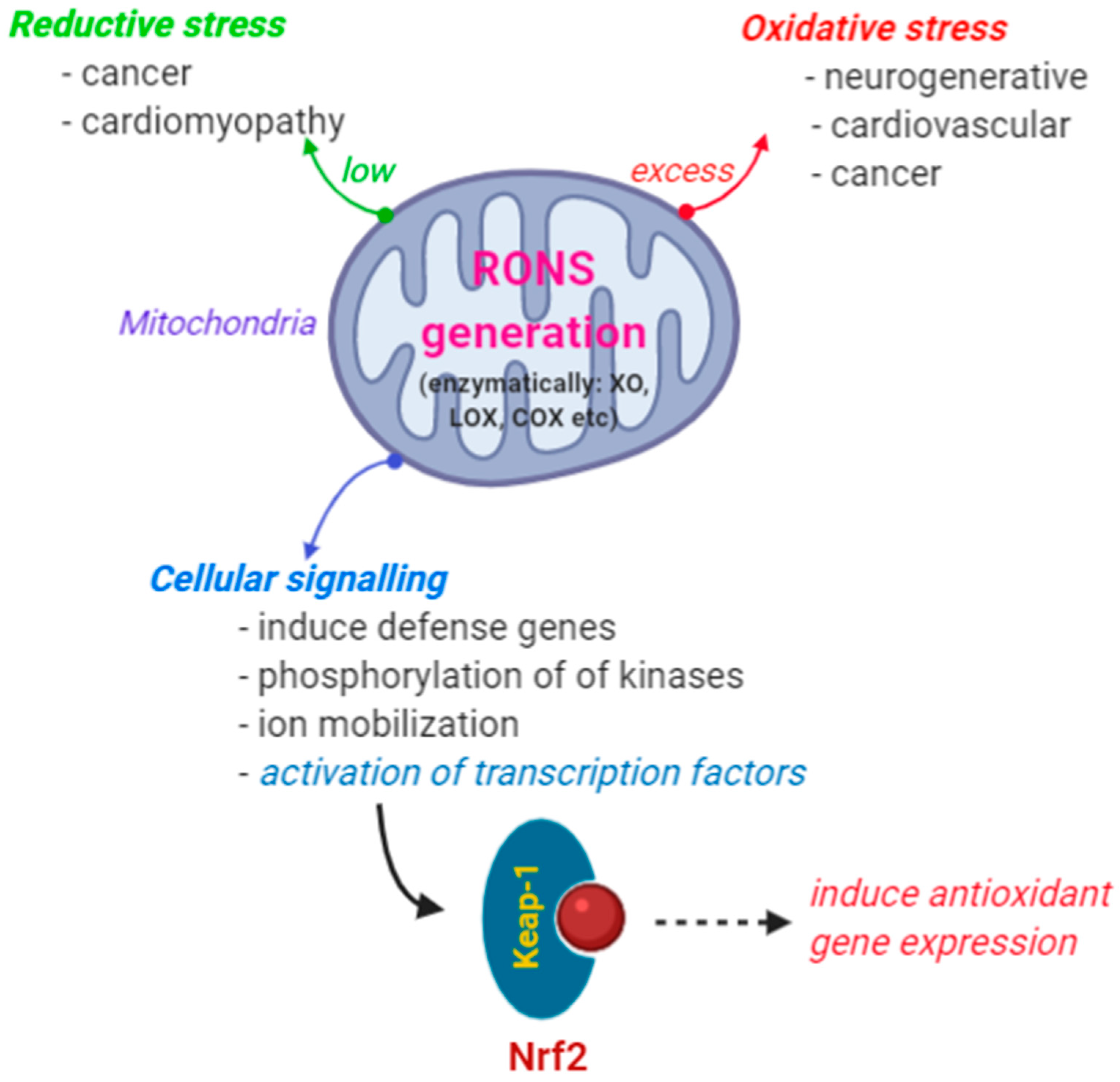
Generation of these reactive species (RONS, both ROS and RNS) in the cells is due to many enzymatic actions, e.g., enzymes that utilize oxygen as a substrate: nicotinamide adenine dinucleotide phosphate oxidase (NADPH-oxidases), xanthine oxidoreductase (XO), lipoxygenase (LOX, lipid-peroxidizing enzymes), myeloperoxidase (MPO), nitric oxide synthase (NOS), and cyclooxygenase (COX) [7,8]. Among these, more than 90% of RONS are generated during the mitochondrial oxidative phosphorylation in the mitochondrial respiratory complex. In this process oxidation of NADH or FADH (the reduced form of flavin adenine dinucleotide) generates energy for protons. These produced electrons react directly with oxygen and electron acceptors and generate RONS [9]. The generation of RONS in the extracellular enzymes is developed as a defense mechanism to kill bacteria. However, the production of excess amounts of RONS may also induce deleterious effects in the biological systems. Another set of enzymes releases RONS intracellularly as a byproduct of metabolic processes. For instance, superoxide (O2•−) is released as a byproduct of mitochondrial respiration and the monooxygenase activity of cytochrome p450 [8,10]. Production of a high amount of RONS in the cells will cause severe damage to different kinds of important biomacromolecules such as carbohydrates, proteins, nucleic acid (DNA, RNA), and lipids [11,12]. This process is referred to as oxidative stress. Thus, altering the normal redox balance in cellular systems leads to increased oxidative stress. RONS induced oxidative stress has been reported to cause several leading diseases, such as diabetes mellitus, neurodegenerative disorders (Parkinson’s disease, Alzheimer’s disease, Huntington’s disease, multiple sclerosis, Cerebral ischemia), cardiovascular diseases (atherosclerosis and hypertension), respiratory diseases (asthma), cataract development, rheumatoid arthritis, and cancers (colorectal, prostate, breast, lung, bladder cancers) [8,13]. On the other hand, if RONS production is too low, reductive stress occurs, which can cause pathogenesis including cancer and cardiomyopathy [14] (Figure 1).
1.1. RONS in Cellular Signaling
Amidst these deleterious effects of RONS, they are now recognized as important in cell signaling and biological processes for induction of defense genes, activation of transcription factors, phosphorylation of kinases, and mobilization of ions in transport systems. It is found that cellular signaling is mainly due to the presence of O2•− and H2O2 as ROS and NO• and ONOO− as RNS [8,15]. These RONS are mainly responsible for the promotion of cell proliferation and apoptotic pathways for the proper regulation of the cell cycle and programmed cell death [16,17]. In cancer, cancer cells drive redox signaling reactions by elevating RONS levels to continue proliferation [18]. As an immune response also, a high level of RONS activates the inflammasomes: the surveillance receptors which are required to release proinflammatory cyto-kinases as an appropriate immune response [19]. Low level of RONS prevents the activation of immune response and leads to immunosuppression [20]. In cardiovascular systems, the generation of RONS together with NADPH-oxidase is important to maintain the functions of endothelial and vascular smooth muscle cells, inflammation, cell growth and proliferation, modulation of extracellular matrix production, apoptosis and angiogenesis [21,22].
Biological systems are made up of systems for balancing the negative effects caused by reactive species and related oxidative stress. For this, the biological system has its own defense mechanism to quench harmful radicals before they attack the cells. These defense mechanisms include scavenging of active radicals enzymatically (superoxide dismutase (SOD), catalase (CAT), ascorbate peroxidase (APX), guaiacol peroxidase (GPX), glutathione reductase (GR), monodehydroascorbate reductase (MDHAR), and dehydro-ascorbate reductase (DHAR) or non-enzymatically (ascorbic acid, reduced glutathione, α-tocopherol, carotenoids, flavonoids etc., which are defined as antioxidants), chelating with metals, and activation of genes that encodes defensive enzymes, transcription factors, and structural proteins [16,23]. Specifically, RONS play a crucial role in activating transcription factors and their encoded defensive enzymes. Since RONS molecules are small, they can easily penetrate the plasma membrane and can directly act on the catalytic receptors or cellular signal inducers leading to the activation of transcription factors. Therefore, as a cellular defense mechanism against oxidative stress, the elevated RONS stimulate the gene expression which produces antioxidant enzymes [24]. In this review we mainly focus on cellular defense against oxidative stress by gene expression and the Nrf2 pathway.
The Nrf2/ARE (Nuclear factor (erythroid-derived 2)-like 2/antioxidant response element) pathway is one of the most important cellular defense mechanisms against oxidative stress. It controls gene expression in which produced proteins are involved in detoxification and elimination of reactive species [25]. Nrf2 is a family member of the transcription factors that activate the battery of cytoprotective genes through ARE, participating in biotransformation, antioxidant reactions, and inflammation [26].
1.2. Regulation of Nrf2 as a Cellular Signal for Oxidative Stress
Nrf2 is a short-lived protein that acts as a transcription factor for the systemic antioxidant defense system. It is localized and expressed in the cytosol. Nrf2 up-regulates the expression of cytoprotective genes involved in antioxidant and anti-inflammatory responses, including heme oxygenase-1 (HO-1), SOD, NADPH Quinone oxydoreductase enzyme (NQO1), catalase, and enzymes involved in glutathione metabolism such as glutathione S-transferase (GST), glutathione cysteine ligase modifier (GCLM) subunit, and glutathione cysteine ligase catalytic (GCLC) subunit [26].
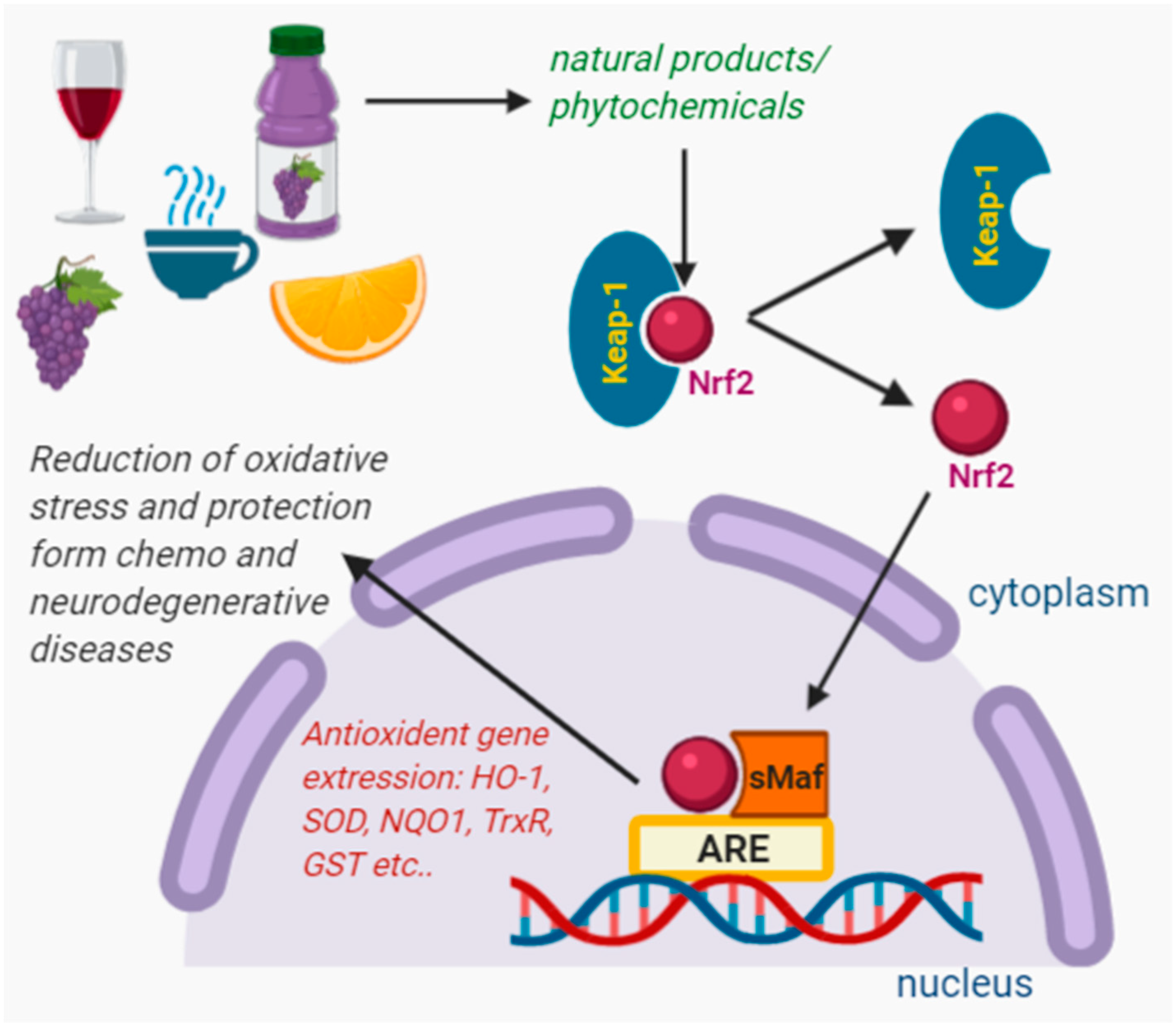
Nrf2 binds to protein Keap1 (Kelch-like erythroid-cell-derived protein with Cap’n’Collar (CNC) homology-(ECH-) associated protein 1) as a complex in the cytoplasm, where it undergoes ubiquitination and proteasomal degradation [27]. Under stressed conditions (increased RONS), Nrf2 separates from Keap1 (a primary Nrf2 inhibitor) due to the conformational changes and translocates into the nucleus. There it induces the production of smaller proteins like sMaf proteins and binds to phase 2 of the antioxidant response element (ARE) in the DNA promoter region [28]. It initiates the protein expressions involved in response to oxidative stress i.e., reduces oxidative stress. Some antioxidative enzymes are induced by Nrf2, such as SOD, catalase, GST, NQO1, HO-1, thioredoxin reductase (TrxR), glutathione reductase (GR), and GST, which detoxify ROS through glutathione regulation [25,26,28,29,30]. There are many comprehensive review articles dealing with Nrf2 activation, regulation and action, which provide good insight into Nrf2. Here we have cited some review articles that could be beneficial for the reader [29,31,32,33].
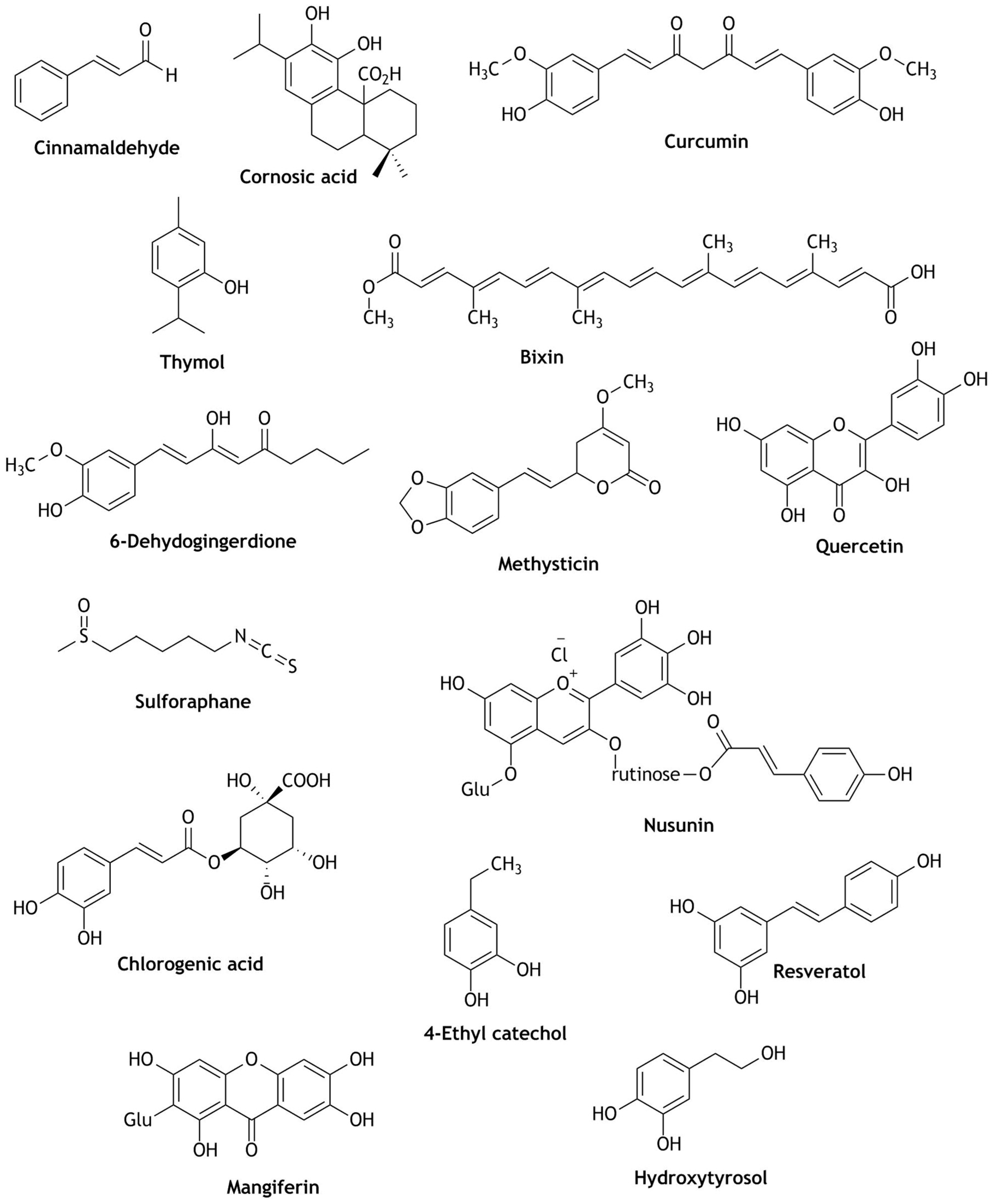
Many plant derived natural compounds or extracts have been claimed to have extremely good antioxidant activity by stimulating the nuclear translocation of Nrf2 [34]. Famous household culinary ingredients like spices: cinnamon (cinnamaldehyde) [35], rosemary (carnosic acid) [36], turmeric (curcumin) [37], thyme (thymol) [38], achiote (bixin) [39], ginger (6-dehydrogingerdione) [40], pepper (methysticin) [41]; vegetables: red onion (quercetin) [42], broccoli, cabbage, kale (sulforaphane), eggplant (nasunin) [42,43,44]; beverages: coffee (chlorogenic acid) [42], tea (quercetin, chologenic acid) [45], wine (quercetin, chologenic acid, resveratol) [46], fruit juices (quercetin, chologenic acid, 4-ethyl catechol) [47]; fruits: grapes (quercetin, chologenic acid, resveratol) [48], mango (mangiferin), citrus fruits, pomegranate [49], olive (hydroxytyrosol) [46] are potential candidates that activate the Nrf2 pathway and have proven ability to prevent chemo and neuro degenerative diseases and in therapy for diseases induced by oxidative stress [50]. These natural compounds can: (i) modify the Keap-1 proteins by oxidation or alkylation, (ii) inhibit the binding of Nrf2 to Keap-1, and (iii) cause phosphorylation of Nrf2 by GSK-3 and subsequently proteasomal degradation [51]. Thus, natural compounds are important and have shown proven ability to activate Nrf2 as a cellular defensive mechanism. Hence, searching for Nrf2 activators could be a promising strategy for drug discovery. Figure 2 depicts some natural compounds (phytochemicals) responsible for the activation of Nrf2.
It was found that wine, juices, and other beverages contain large amounts of antioxidants; their consumption could prevent the activity of RONS, leading to the prevention of oxidative stress-related diseases [52]. Meanwhile, scavenging or neutralizing these RONS, antioxidants activate the Nrf2 signaling system (Figure 3).
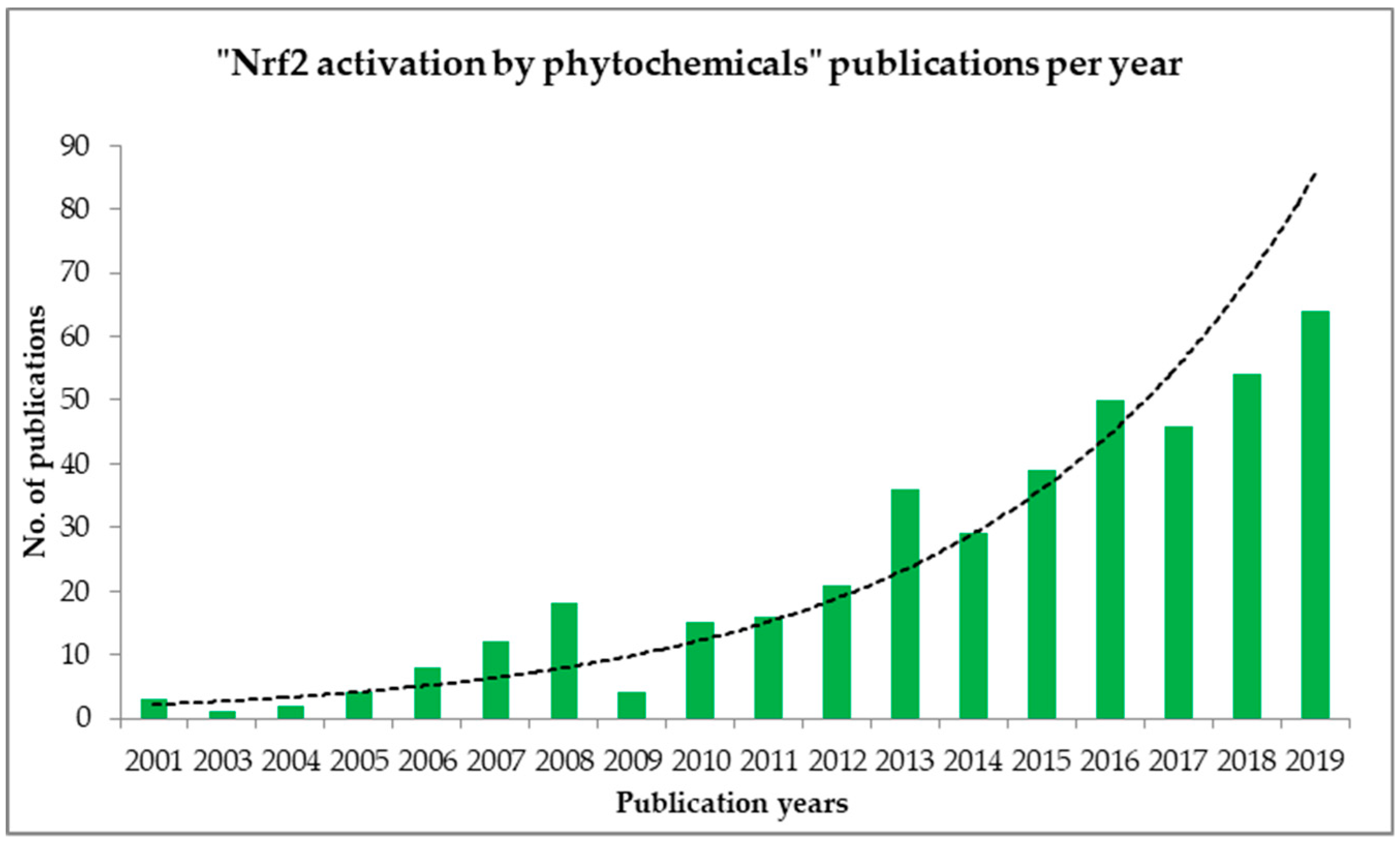
The present review deals with the activation of Nrf2 by the natural components present in juices, coffee, cocoa, and wines. To the best of our knowledge, this is the first review of Nrf2 activation by natural chemical entities present in juices, wines, coffee, and cocoa beverages. According to PubMed, studies on Nrf2 activators from natural products (phytochemicals) are increasing (Figure 4). This could be the reason why scientists are working tremendously to discover new Nrf2 activators and develop new drugs or drug leads to combat oxidative stress-related diseases. According to the recently published report by the US Department of Health and Human Services together with Centers for Disease Control and Prevention (CDC), oxidative stress-related diseases like diabetes mellitus, Alzheimer’s disease, cardiovascular diseases, and cancer are listed among the leading causes of death in the United States [53]. This review compiles the available data regarding the activation of Nrf2 from popular beverages. The information provided here will be valuable to beverage industries and stakeholders, as well as for consumers. It will present some scientific validation of the beverages that we consume without knowing of their importance. Since these data show that the beverages have a positive impact on oxidative stress-related diseases, it is worth disseminating these findings among interested parties. Hence, this review will provide a better picture for consumers of the importance of the consumption of beverages and their impact against oxidative stress-related diseases, as well as for industries and stakeholders about how to improve their product quality and to know more about their products with proven scientific evidence.
2. Nrf2 Modulation of Natural Compounds Present in Fruit Juices
It is proved that consumption of a high amount of fruits, fruit juices, and other forms of plant based foods (such as vegetables and nuts) will reduce the risks of developing many life threatening chemo- and neuro-degenerative diseases such as cancer and cardiovascular diseases [54]. The absence of undesirable side effects has promoted consumption significantly. The presence of natural bioactive phytochemicals such as phenolic, vitamins (e.g., vitamin E), and carotenoids is attributed to their potential antioxidant activity. Apart from scavenging ability against RONS, these antioxidants can protect against ROS-mediated oxidative damage by elevating cellular antioxidant capacity [55]. Though there are many reports of fruit juices that were found to be potential candidates for the prevention of chemo- and neuro-degenerative diseases, the underlying mechanism of action is still unclear. As a result, studies have been conducted to unveil the mechanism of the Nrf2 pathway. A number of phenolic compounds abundantly present in fruit juices have been identified and these could modulate the Nrf2 pathway through translocation of Nrf2 into the nucleus, up-regulation of antioxidant enzymes, and reduction of RONS.
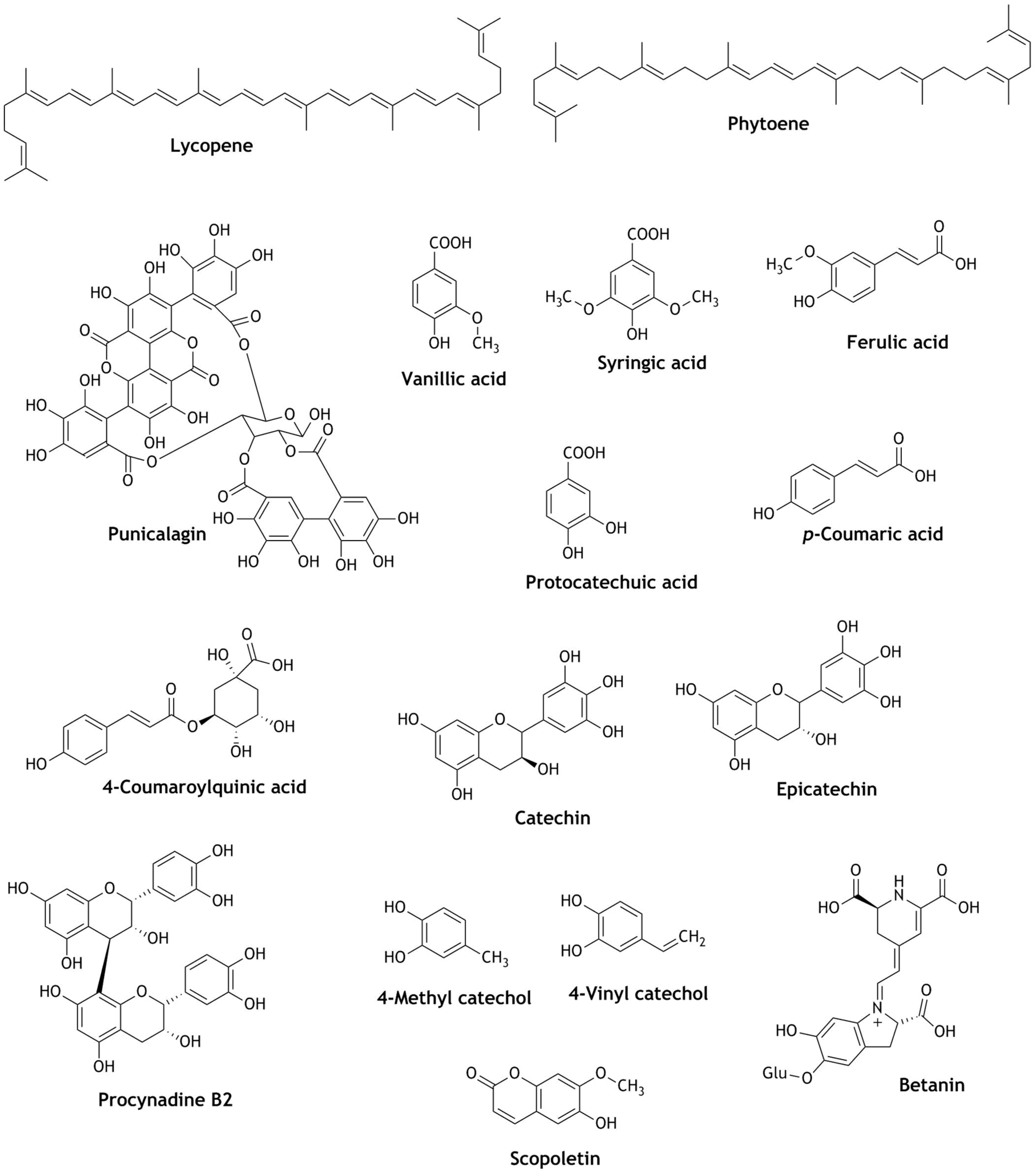
Citrus sinensis, also known as Washington Navel orange, is chemically rich in carotenoids with known antioxidant and anti-inflammatory activity, and cellular protective activity against oxidative stress [56]. Carotenoids like lycopene, phytoene, and β-carotenes are abundantly found in their peel and pulp. A study carried out on pasteurized C. sinensis orange juices extracted from pulp showed significantly increased reduction of ROS, up-regulation of antioxidant enzyme genes such as GST-4, SOD-4, GCS-1, and HSP-16-2, and resistivity against oxidative stress as a consequence of high carotenoid content [56]. Anthocyanins are another class of natural phytochemicals called flavonoids, abundantly found in nature. Anthocyanins in pomegranate are responsible for the color of the fruit and its biological function is still not fully known [57], although anthocyanins and hydrolysable tannins isolated from the juices of pomegranate significantly increased Nrf2 expression in nitrosodiethylamine-induced fibrotic rats [58]. Meanwhile, another study showed that punicalagin isolated from pomegranate fruit juice increased Nrf2 nuclear translocation, up-regulated HO-1 gene expression and decreased the generation of RONS. As a result, stress-induced cell death and apoptosis of IEC-6 cells were significantly reduced [59]. Chlorogenic acid, vanillic acid, syringic acid, trans-ferulic acid, protocatechuic acid, 4-coumaroylquinic acid, and p-coumaric acid are some of the common phenolic acids abundantly found in fruits [60]. These natural acids together with flavonoids catechins, epicatechins, and the epicatechin dimer procyanidin B2 showed strong antioxidant activity, because of their ability to activate Nrf2 translocation and strong up-regulation of HO-1, GPX2GSR, CAT, and NQO-1 antioxidant enzymes. Furthermore, it was reported that these phytochemicals identified in fruit juices like apple and blueberry provided protection against RONS-induced toxicity and were beneficial to endothelial cell activation and vascular functions [61,62]. Noni is a tropical tree native to Southeast Asia, Australia, and the South Pacific including Hawaii. Noni is used in traditional medicine for a diverse range of ailments. Its anti-cancer, anti-inflammatory, and antidiabetic properties were well known in folklore, but there was a lack of scientific evidence to support the claims [63]. As a result, Chen et al. in 2018 tested different organic extracts of Noni juice against Nrf2 activity. They found that aqueous and chloroform extracts protect the cells from tert-butyl hydroperoxide (TBHP)-induced cell damage and significantly decrease TBHP cytotoxicity, apoptosis, and RONS generation. Meanwhile, nuclear accumulation of Nrf2 and up-regulation of HO-1, CAT, and SOD-1 were also observed [64]. This antioxidant and neuroprotection activity may be due to the presence of different types of phytochemicals in both fractions, such as high phenolic and flavonoid content in the chloroform fraction and rich polysaccharides content in the aqueous fraction [64]. This study showed the neuroprotective activity of Noni extracts and their molecular mechanisms. In two studies carried out by Senger and Cao on Noni fruit juice, three alkyl catechols, namely 4-ethyl catechol, 4-methyl catechol, and 4-vinyl catechol [47], and a coumarin compound scopoletin [65] were identified as Nrf2 co-factors. These compounds significantly increased the nuclear translocation of Nrf2 and the up-regulation of the expression of HO-1 and NQO-1 genes [47,65]. Similarly, vegetable juices like broccoli, sprout, garlic, cabbage juices and betanin from beetroot juice activated Nrf2 translocation and up-regulation of a panel of antioxidant genes such as GSTP, GSTM, GSTT, NQO-1, HO-1, thioredoxin, thioredoxin reductase, and NAD(P)H:quinones [66,67,68]. Phytochemicals present in fruit juices as Nrf2 activators (Figure 5) are listed in Table 2.
3. Nrf2 Modulation of Natural Compounds Present in Wines
The health benefits associated with wines have been documented and its popularity noted since early Roman days [71]. In vitro and in vivo studies have proven the scientific rationale. These studies claimed that a certain amount of wine consumption on a daily basis could reduce the risk of cardiovascular and chronic diseases [72]. Red wine from grapes is one of the most famous beverages in the world. It is composed of carbohydrates, minerals, water, alcohol, and a notable number of polyphenols. These polyphenols are the main natural products that account for the reported potential health benefits [72]. The most important polyphenols present in wine are resveratrol, anthocyanins, catechins, pro-anthocyanidins, and ellagitannins [73]. These phenolic compounds are mainly found in pulp/flesh, skins, and seeds that are extracted during the winemaking process [74]. Depending on the cultivars, climate, harvesting and post-harvesting periods, and winemaking techniques will greatly affect the polyphenol contents of the grapes [74,75,76]. Guaita and Bosso [74] recently studied four red grape varieties and their polyphenolic content. Their results showed that Barbera and Albarossa grape varieties had a significantly higher number of polyphenols compared to Nebbiolo and Uvalino grapes. Thus, it is important to mention that, depending on the above factors, the polyphenolic content of grapes varies, and so does their bioactivity.
In Nrf2 pathway studies, resveratrol was found to be the predominant therapeutically active candidate for chemoprevention, neuroprotection, improvement of cardiac functions and regeneration, and protection of brain cells against cerebral ischemia by suppressing oxidative and inflammatory stress [77,78,79,80]. Via molecular mechanisms, resveratrol also enhances the expression of Nrf2 and up-regulation of NQO-1, HO-1, GST-P1, and GSH [77,78,79]. Moreover, in a preclinical study conducted by Bishayee et al. (2010) [81] at a dose of 100 mg/kg, resveratrol reduced liver tumorigenesis and increased nuclear translocation of Nrf2 in mice. Furthermore, some studies have shown that, apart from the increased Nrf2 expression, increased redox effector factor-1 (REF-1) which is important in DNA repairing and redox signaling make resveratrol an excellent target for chemotherapy, stem cell survival, proliferation, and cell regeneration [82]. Shan et al. also showed that a dose of 20 mM of resveratrol increased Nrf2 translocation, protected oxygen-glucose deprivation/reoxygenation through up-regulation of NQO-1and HO-1 genes and increased activity of SOD and GSH [80].
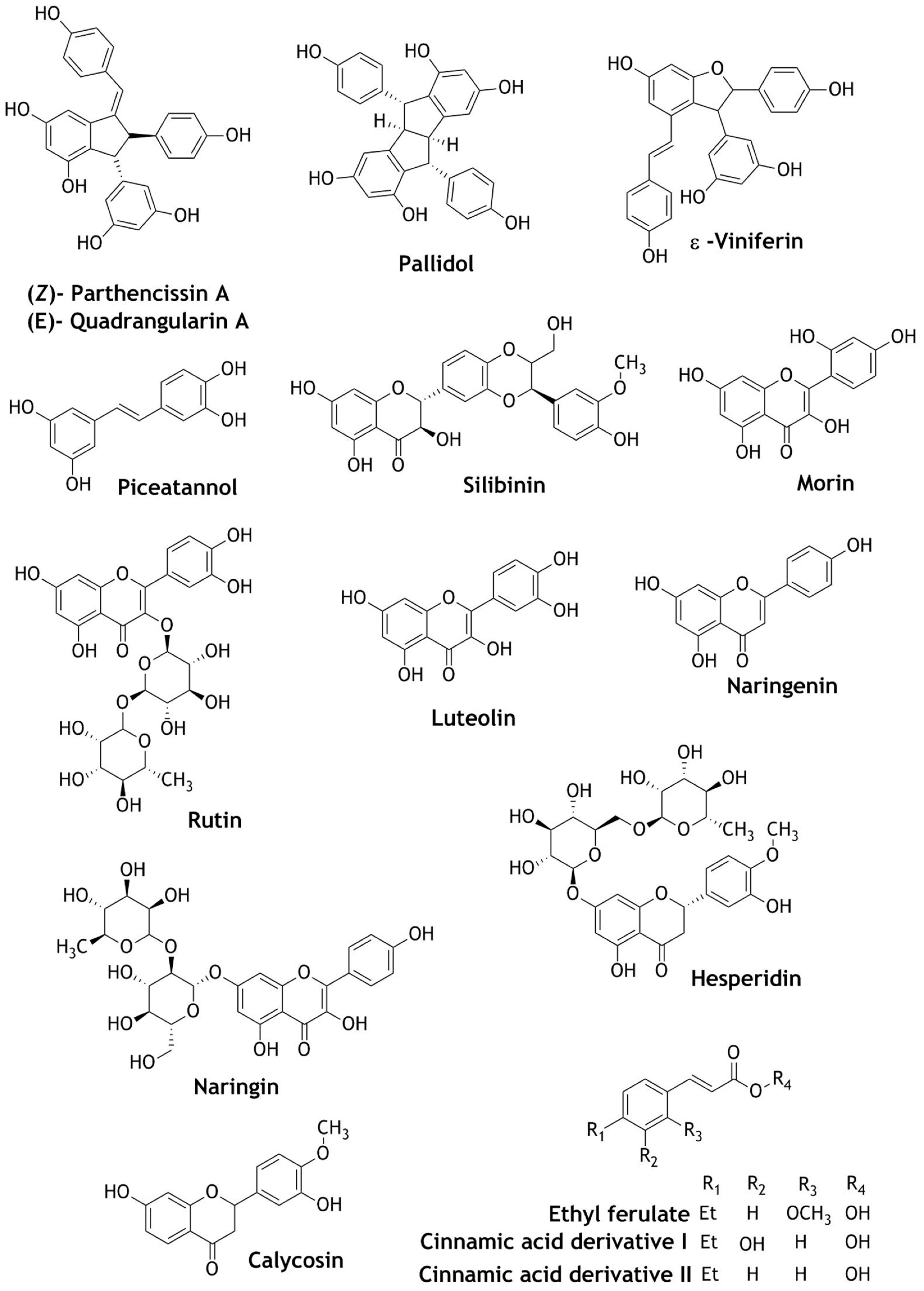
Naturally occurring resveratrol dimers parthenocissin A, quadrangularin A, pallidol, and dehydro-dimer ε-viniferin present in wine have been studied for activation of Nrf2 transcription. The dimers were capable of selectively quenching only the singlet oxygen (1O2) species, but not hydroxyl or superoxide anions. Only pallidol at a dose of 30 μM activated the Nrf2 activity [83]. Similarly, ε-viniferin more effectively inhibited the platelet-derived growth factor-induced cell proliferation and migration than resveratrol and less effectively inhibited ROS production and nitric oxide generation. ε-viniferin also increased the expression of the antioxidant enzyme HO-1 via the Nrf2 pathway. Therefore, ε-viniferin is more effective than resveratrol, and collectively both compounds could prevent atherosclerosis [84]. Further, polyphenolic compounds abundantly found in red wine such as quercetin, catechin, epicatechin, procyanidin B2, piceatannol, and silibinin, have been reported to protect against oxidative stress via the activation of the Nrf2 pathway and up-regulation of a series of antioxidant enzymes HO-1, GSH, NADPH, GST, NQO-1 [85,86,87], SOD [88], GSTP1 [89], and metallothioneins 1 and 2 (MT-1/2) [90]. The protective mechanisms of pancreatic β-cells in silibinin were studied in detail. A dose of 100 mg/kg of silibinin administrated to the Wistar rats for four weeks decreased blood glucose levels as well as increased the expression of estrogen receptor-α (ER-α), Nrf2, and HO-1 in pancreatic β-cells. Silibinin at 10 μM increased insulin biosynthesis and secretion and decreased the production of ROS. Therefore silibinin used therapeutically could improve the glucose homeostasis in diabetic patients [91]. Similarly, a dose of 10 µM of piceatannol was used to study neuroprotective potential. The expression of HO-1 via Nrf2 activation was increased and phenolic piceatannol was found to be a good candidate for the protection of neurons [92].
Weng et al. [90] have studied structurally diverse flavonoids such as flavanols: quercetin, morin, rutin; flavone: luteolin; flavanone: naringenin, naringin, hesperidin; and flavanol: catechin for their chemo-preventive mechanism. Of these flavonoids quercetin, morin, and rutin showed significant in-vitro antioxidant activity, which correlated with the flavonoid induced metallothionein expression in HepG2 cells. Furthermore, the observed correlation may be due to structural variations, such as the presence of phenolic hydroxyl groups that stabilize the scavenging activity [90].
Apart from red wine, other wines such as raffia palm (Raphia hookeri) wine, Makgeolli lees (ML) fermented raw rice wine, yellow rice wine, and Chinese Jing liquor also showed protective effects against oxidative stress through Nrf2 activation [93,94]. They enhanced the translocation of Nrf2 into nuclei, which up-regulated the expression of NQO-1, HO-1, GPX, SOD, CAT, and peroxiredoxin 3 and 4 enzymes [93,94]. Chinese yellow rice wine is one of the oldest wines in the world [95]. The polyphenolics present in yellow rice wine were beneficial against cardiovascular diseases. A dose of 30 mg/kg was effective in preventing doxorubicin-induced cardiotoxicity through the activation of Nrf2 signaling pathway [96]. A study of Chinese Jing liquor, one of the most popular health beverages in China, revealed the presence of a broad spectrum of bioactive metabolites including 189 natural phytochemicals. All the compounds were assayed for their Nrf2 luciferase activity. The crude extract increased the Nrf2/ARE luciferase activity. Eighteen compounds activated Nrf2, among which calycosin, ethyl ferulate, and two cinnamic acid derivatives (I and II) strongly activate the Nrf2 reporter activity at 40 μg/mL [97]. Phytochemicals present in wine (Figure 6) as Nrf2 activators are listed in Table 3.
4. Nrf2 Modulation of Natural Compounds Present in Coffee and Cocoa
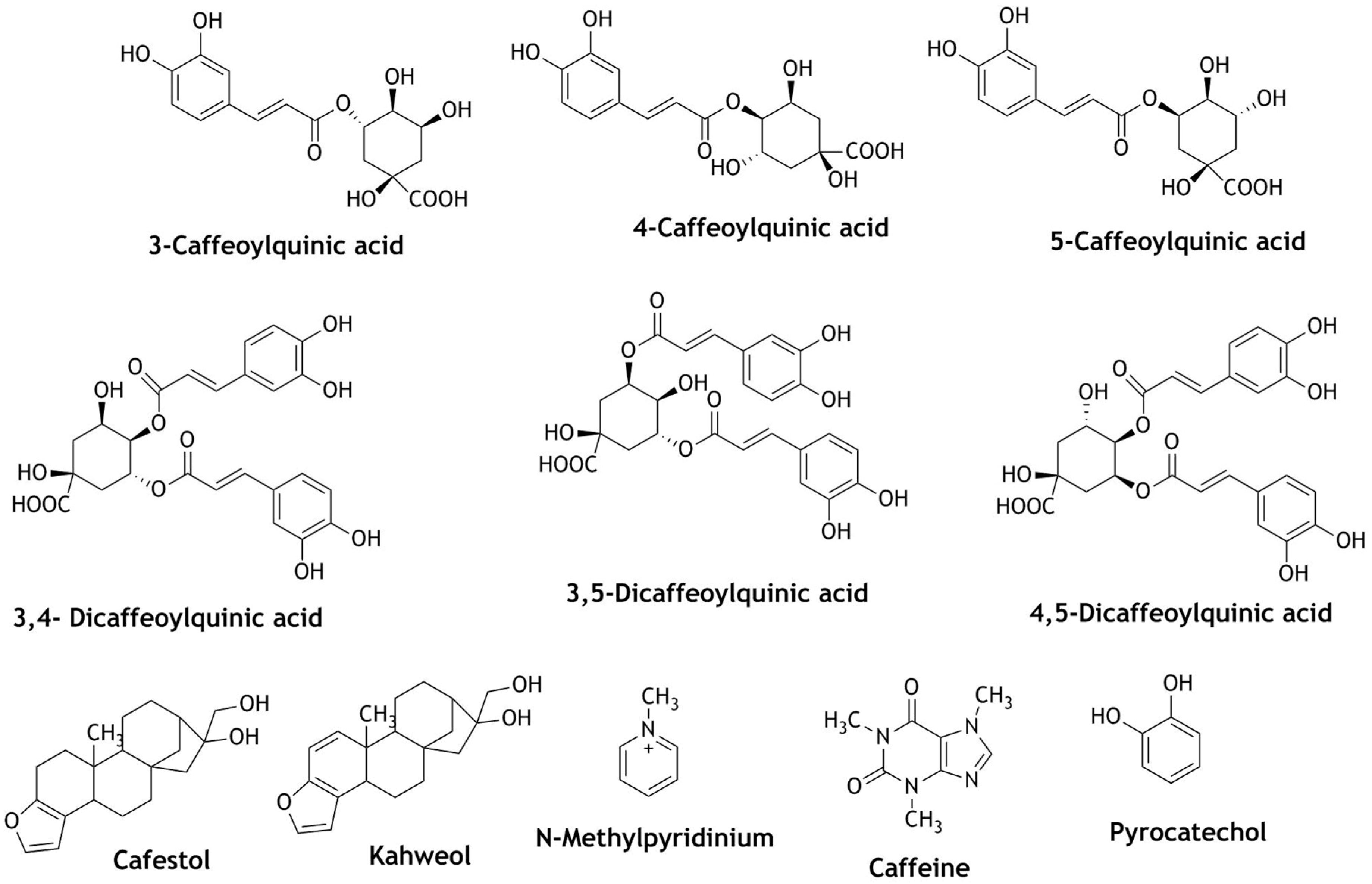
Coffee, tea, and cocoa-containing beverages are known to have potential antioxidant activity due to the presence of rich flavonoid and polyphenolic compounds. A cup of coffee contains about 200–500 mg of polyphenols, 150–200 mg in a cup of tea, and 200–400 mg in 100 g of cocoa powder [98]. Therefore, these beverages make a significant contribution in terms of phenolic intake for the human diet [98,99]. Coffee contains a large number of polyphenolic compounds such as chlorogenic acid, ferulic acid, p-coumaric acid, and caffeic acid. Ten grams of coffee per cup of brew contains about 325 mg of chlorogenic acid [100]. Similarly, the polyphenols present in cocoa are catechins, procyanidins, and anthocyanidins which account for its antioxidant activity [100]. Similar to other foods and beverages, the mechanism of these natural polyphenols in the prevention of chemo- and neuro-degenerative diseases have not been fully revealed although many research studies have been carried out. Since the consumption of coffee, tea, wines, and cocoa-containing products such as chocolates and chocolate drinks are very popular globally, it is worthwhile to review the epidemiological studies carried out so far. The polyphenolic compounds present in these beverages vary, but they are mainly epicatechins in green teas; epicatechins and tannins in black teas; catechins, procyanidins, and anthocyanins in cocoa; and chlorogenic acids, caffeic acid, and melanoidins (high molecular weight nitrogenous polymeric and brown-colored compounds) in roasted coffee [100,101]. In biological systems, the polyphenols show dual actions to maintain the oxidative balance. As a direct antioxidant, they quench the RONS and make them less harmful. On the other hand, they enhance the production of hydroxyl radicals to stimulate cellular signaling. As a response, the biological system will react to the cellular injury [2]. Table 4 summarizes the major polyphenols isolated from coffee, tea, and cocoa (Figure 7) and how they behave in the Nrf2 pathway.
Chlorogenic acid and its derivatives 3-CQA (caffeoylquinic acid), 4-CQA, 5-CQA, 3,4-diCQA, 3,5-diCQA, and 4,5-diCQ activate antioxidant enzyme genes such as HO-1, NQO-1, glutathione, thioredoxin reductase-1 (TrxR), and thioredoxin-1 [102,103,104]. In cocoa, catechins, epicatechins and procyanidin B2 have been studied for their Nrf2 activity. While activating the antioxidant enzymes in the Nrf2 pathway, these compounds reduced the production of RONS, improved cellular redox state, and modulated the mitochondrial function. Moreover, there are different preparations of coffee, cocoa, or tea in different countries. A study conducted by Richelle et al. (2001) reported that coffee, cocoa, or tea do not have any effect on antioxidant activity when consumed with milk [100]. Light roasting will degrade the polyphenolics and, as a result, antioxidant activity decreases [100]. Similarly, caffeinated and decaffeinated coffee have the same effect on cellular signaling and antioxidant activity [100]. Trinh et al. (2010) identified cafestol from caffeinated and decaffeinated coffee, which is one of the natural products triggering Nrf2 translocation into the nucleus [105]. Cafestol is a diterpenoid made up of four C5 isoprenyl units. Terpenoids represent a structurally diverse and biologically highly active class of compounds in natural products. The only structural difference between cafestol and kahweol is the presence of one conjugated double bond (C=C) on the furan ring in kahweol [106]. Both compounds have the ability to activate Nrf2 translocation by activating antioxidant enzymes and inhibiting RONS production in cellular systems [107,108,109,110]. Moreover, kahweol increased the expression and up-regulation of phase II antioxidant enzymes more significantly than cafestol [111]. A study of green coffee and roasted coffee identified 5-CQA and N-methylpyridinium respectively as the major metabolites. These compounds showed increased nuclear translocation of Nrf2 and ARE dependent antioxidant genes NQO-1 and GSTA-1 in HT29 cells [112]. Chlorogenic acid, caffeine, and caffeic acid are some of the key bioactive metabolites in coffee. Studies conducted on these metabolites from coffee show that these compounds have potential therapeutic applications as anti-inflammatory, antidiabetic, and in cancer therapy. Supplementation of these compounds activated Nrf2 translocation and expression of HO-1 and CAT proteins, decreased the generation of intercellular RONS, and modulated the interaction between Keap1 and Nrf2 via p62 expression [113,114,115].
5. Nrf2 Inhibitors from Beverages
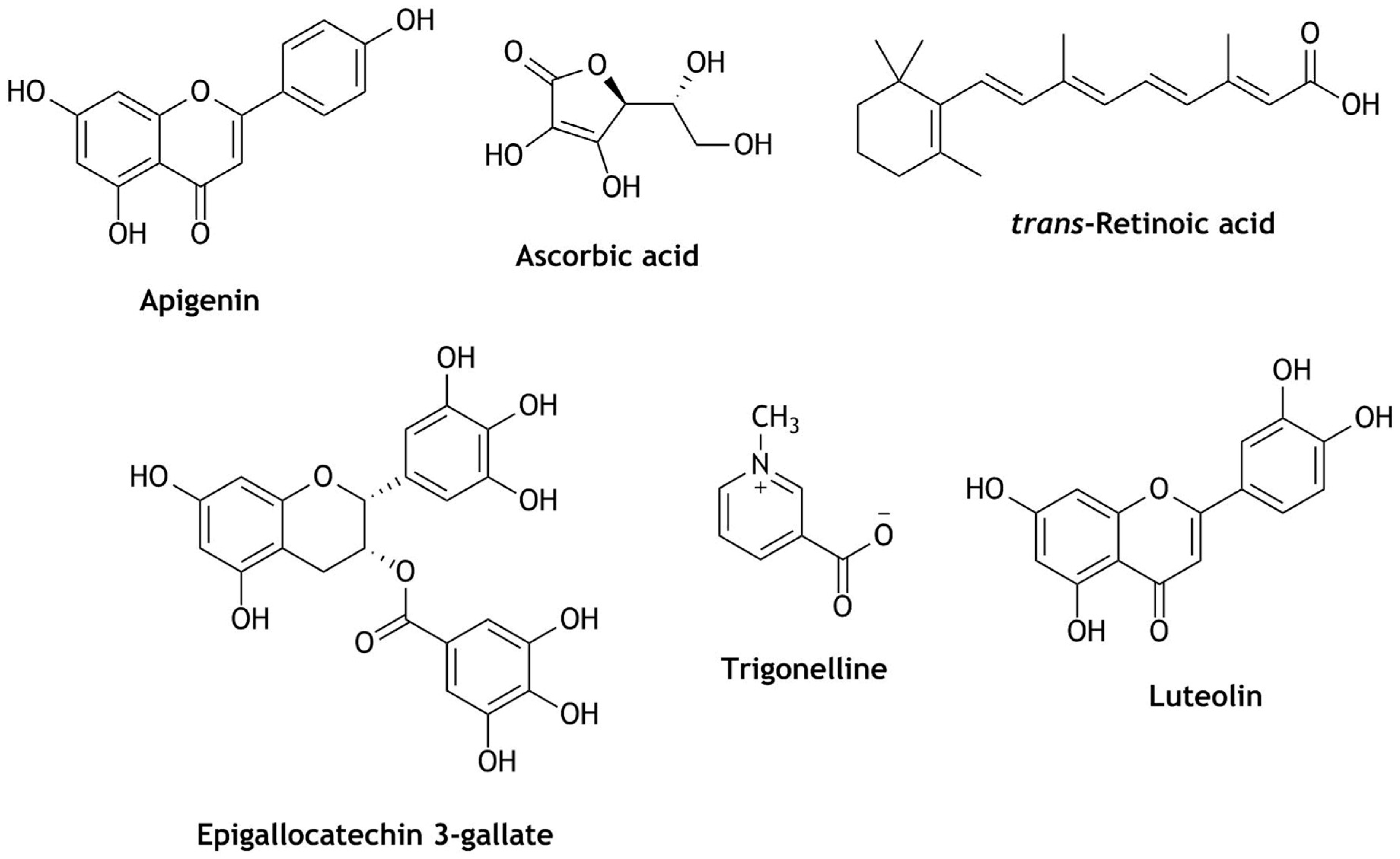
Though the activation of Nrf2 has been studied extensively and found to be important in chemo- and neuro-preventive diseases, the dark side of Nrf2 has been identified—substantial overexpression of Nrf2 resulted in enhanced resistance of cancer cells to chemotherapeutic drugs. The prolonged expression of Nrf2 inhibits nuclear accumulation, which prevents Nrf2 dependent gene expression. This increased expression of Nrf2 might cause chemo-resistivity, hyperkeratosis of the upper digestive tract, metastasis, atherosclerosis, and cancer progression [126,127,128]. Hence, in cancer biology Nrf2 is considered as a double-edged sword [127]. It was found that high Nrf2 levels were produced as a result of mutations of Keap1. This is frequent in lung, head, neck, and bladder cancer tissues and, therefore, develops chemo-resistance and cancer proliferation [51,126,128,129]. These studies concluded that Nrf2 is an important transcription factor in the early stages of some cancers and when it progresses over-expression of Nrf2 in cancer cells will become resistant to cancer treatments (chemo- and radiotherapy). Thus, inhibiting Nrf2 could have potential applications in cancer therapy [51,129]. Particularly in beverages, very few molecules have been identified as Nrf2 inhibitors. Ascorbic acid (fruit juices) [130], apigenin (coffee, tea, fruit juices) [131], epigallocatechin 3-gallate (tea) [132], trans-retinoic acid (juices of fruits, vegetables) [133,134], luteolin (tea, juices of fruits, vegetables) [135,136] and trigonelline (coffee) [137] are reported to have Nrf2 inhibitory activity (Figure 8). These phytochemicals reduced Nrf2 activation, selectively downregulated the mRNA and protein levels in biological systems, degraded Nrf2, suppressed the expression of Nrf2-HO-1 genes, decreased the binding ability of Nrf2-ARE and HO-1-ARE luciferase activity, and reduced the expression of ARE-driven genes, leading to sensitization to therapeutics [51,130,131,132,133,135,136,137]. Although studies showed that the above-mentioned polyphenols inhibited Nrf2 activity, it is important to understand the mechanism of action and to further study their selectivity for Nrf2 inhibition.
6. Conclusion and Future Perspectives
Natural products play a central role in the drug discovery process regardless of their source. Plant phytochemicals have been extensively studied for centuries due to their beneficial biological properties and abundance in nature. The production of reactive species as a result of oxidative stress is the main cause of many chronic diseases. Phytochemicals play a major role in scavenging these reactive species. The availability and position of phenolic hydroxyl groups stabilize free radical scavenging activity. Since these phenolic hydroxyl groups are electron donor groups, they enhance antioxidant activity. In cellular systems, the Nrf2/ARE pathway is important to maintain the oxidative balance as well as to express the phase II antioxidant enzymes. Therefore, as a result of oxidative stress, transcriptional activation of Nrf2 antioxidant target genes will take place as a protective action. Hence, the Nrf2 pathway is considered a cellular survival pathway. Antioxidant natural products could break the specific interaction between Keap1 and Nrf2 and translocate the Nrf2 into the nucleus. As a result, this induces the production of smaller proteins and binds to ARE in the DNA promoter region, which initiates phase II protein expressions as a response to oxidative stress, i.e., reducing oxidative stress. Studies have shown the importance of phytochemicals and their impact on Nrf2 as a therapeutic approach for chemo-, inflammatory, and neurodegenerative diseases. On the other hand, the abnormal activation or accumulation of Nrf2 can cause adverse effects. This is common in many tumors, which gives cancer cells a selective advantage and is associated with malignant progression, chemo-resistance and poor prognosis. Hence, it is worth mentioning that Nrf2 activators could be used for the prevention of chemical carcinogenesis, whereas Nrf2 inhibitors could be used for cancer treatment. Therefore, it is important to study both Nrf2 activators as well as inhibitors in detail.
Fruit juices, wine, and beverages like coffee, tea and cocoa are the most popular drinks in the world. Many studies have shown their potential antioxidant activity because of the presence of high flavanoid/phenolics content. Fruit juices, wine, and other above-mentioned beverages are also a main source of phenolics in our diet. Therefore, the consumption of these beverages as a dietary supplement is an important part of keeping a healthy lifestyle, which could positively influence our health and lifespans. Meanwhile, manufacturers should pay attention to which products are rich in Nrf2 activators and which contain Nrf2 inhibitors. Consumers should choose beverages cautiously according to their own health conditions because different beverages contain different Nrf2 modulators.
Author Contributions
M.Q. drafted and corrected the manuscript; S.C. conceived, guided, and revised this article. J.X., Y.Y., and Y.L. had funding secured. All authors have read and agreed to the published version of the manuscript.
Funding
The authors are grateful to Jing Brand Research Institute, Jing Brand Co., Ltd. for financial support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Tramutola, A.; Lanzillotta, C.; Perluigi, M.; Butterfield, D.A. Oxidative stress, protein modification and Alzheimer disease. Brain Res. Bull. 2017, 133, 88–96. [Google Scholar] [CrossRef]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.D.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef] [PubMed]
- Radi, R. Peroxynitrite, a stealthy biological oxidant. J. Biol. Chem. 2013, 288, 26464–26472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durairajanayagam, D. Physiological Role of Reactive Oxygen Species in Male Reproduction. In Oxidants, Antioxidants and Impact of the Oxidative Status in Male Reproduction; Elsevier Academic Press: New York, NY, USA, 2018; pp. 65–78. ISBN 9780128125014. [Google Scholar]
- Hardy, M.; Zielonka, J.; Karoui, H.; Sikora, A.; Michalski, R.; Podsiadły, R.; Lopez, M.; Vasquez-Vivar, J.; Kalyanaraman, B.; Ouari, O. Detection and Characterization of Reactive Oxygen and Nitrogen Species in Biological Systems by Monitoring Species-Specific Products. Antioxid. Redox Signal. 2018, 28, 1416–1432. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B. Current status review: Free radicals, reactive oxygen species and human disease: A critical evaluation with special reference to atherosclerosis. Br. J. Exp. Pathol. 1989, 70, 737–757. [Google Scholar] [PubMed]
- Thuy, T.T.; Hai, H.; Kawada, N. Role of oxidative and nitrosative stress in hepatic fibrosis. In Liver Pathophysiology: Therapies and Antioxidants; Elsevier Academic Press: New York, NY, USA, 2017; pp. 213–224. [Google Scholar]
- Weidinger, A.; Kozlov, A.V. Biological activities of reactive oxygen and nitrogen species: Oxidative stress versus signal transduction. Biomolecules 2015, 5, 472–484. [Google Scholar] [CrossRef] [Green Version]
- Balaban, R.S.; Nemoto, S.; Finkel, T. Mitochondria, oxidants, and aging. Cell 2005, 120, 483–495. [Google Scholar] [CrossRef] [Green Version]
- Snezhkina, A.V.; Kudryavtseva, A.V.; Kardymon, O.L.; Savvateeva, M.V.; Melnikova, N.V.; Krasnov, G.S.; Dmitriev, A.A. ROS Generation and Antioxidant Defense Systems in Normal and Malignant Cells. Oxid. Med. Cell. Longev. 2019, 2019, 6175804. [Google Scholar] [CrossRef]
- Halliwell, B.; Aruoma, O.I. DNA damage by oxygen-derived species Its mechanism and measurement in mammalian systems. FEBS Lett. 1991, 281, 9–19. [Google Scholar] [CrossRef] [Green Version]
- Lyras, L.; Perry, R.H.; Perry, E.K.; Ince, P.G.; Jenner, A.; Jenner, P.; Halliwell, B. Oxidative damage to proteins, lipids, and DNA in cortical brain regions from patients with dementia with Lewy bodies. J. Neurochem. 1998, 71, 302–312. [Google Scholar] [CrossRef] [Green Version]
- Phaniendra, A.; Jestadi, D.B.; Periyasamy, L. Free radicals: Properties, sources, targets, and their implication in various diseases. Indian J. Clin. Biochem. 2015, 30, 11–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liou, G.Y.; Storz, P. Reactive oxygen species in cancer. Free Radic. Res. 2010, 44, 479–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, K.; Thomas, S.R.; Keaney, J.F. Beyond LDL oxidation: ROS in vascular signal transduction. Free Radic. Biol. Med. 2003, 35, 117–132. [Google Scholar] [CrossRef]
- Mittler, R. ROS Are Good. Trends Plant Sci. 2017, 22, 11–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, R.; Rinker, L.; Peng, J.; Chilian, W.M. Reactive oxygen species: The good and the bad. In Reactive Oxygen Species (ROS) in Living Cells; Filip, C., Albu, E., Eds.; IntechOpen: London, UK, 2018; pp. 7–20. ISBN 9781789231342. [Google Scholar]
- Cairns, R.A.; Harris, I.S.; Mak, T.W. Regulation of cancer cell metabolism. Nat. Rev. Cancer 2011, 11, 85–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, R.; Yazdi, A.S.; Menu, P.; Tschopp, J. A role for mitochondria in NLRP3 inflammasome activation. Nature 2011, 469, 221–226. [Google Scholar] [CrossRef] [PubMed]
- West, A.P.; Shadel, G.S.; Ghosh, S. Mitochondria in innate immune responses. Nat. Rev. Immunol. 2011, 11, 389–402. [Google Scholar] [CrossRef] [Green Version]
- Crestani, B.; Besnard, V.; Boczkowski, J. Signalling pathways from NADPH oxidase-4 to idiopathic pulmonary fibrosis. Int. J. Biochem. Cell Biol. 2011, 43, 1086–1089. [Google Scholar] [CrossRef]
- Da Costa, R.M.; Rodrigues, D.; Pereira, C.A.; Silva, J.F.; Alves, J.V.; Lobato, N.S.; Tostes, R.C. Nrf2 as a potential mediator of cardiovascular risk in metabolic diseases. Front. Pharmacol. 2019, 10, 382. [Google Scholar] [CrossRef] [Green Version]
- Kurutas, E.B. The importance of antioxidants which play the role in cellular response against oxidative/nitrosative stress: Current state. Nutr. J. 2016, 15, 71. [Google Scholar] [CrossRef] [Green Version]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, T.; Nioi, P.; Pickett, C.B. The Nrf2-antioxidant response element signaling pathway and its activation by oxidative stress. J. Biol. Chem. 2009, 284, 13291–13295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niture, S.K.; Khatri, R.; Jaiswal, A.K. Regulation of Nrf2—An update. Free Radic. Biol. Med. 2014, 66, 36–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jadeja, R.N.; Upadhyay, K.K.; Devkar, R.V.; Khurana, S. Naturally occurring Nrf2 activators: Potential in treatment of liver injury. Oxid. Med. Cell. Longev. 2016, 2016, 3453926. [Google Scholar] [CrossRef] [Green Version]
- Wu, K.C.; McDonald, P.R.; Liu, J.; Klaassen, C.D. Screening of natural compounds as activators of the keap1-nrf2 pathway. Planta Med. 2014, 80, 97–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Q. Role of Nrf2 in oxidative stress and toxicity. Annu. Rev. Pharmacol. Toxicol. 2013, 53, 401–426. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.F.; Zhou, D.D.; Xie, T.; Malik, T.H.; Lu, C.B.; Li, H.J.; Wang, F.; Shu, C.; Liu, C.; Lu, C.W.; et al. Nrf2, a potential therapeutic target against oxidative stress in corneal diseases. Oxid. Med. Cell. Longev. 2017, 2017, 2326178. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Lu, H.; Bai, Y. Nrf2 in cancers: A double-edged sword. Cancer Med. 2019, 8, 2252–2267. [Google Scholar] [CrossRef]
- Ahmed, S.M.U.; Luo, L.; Namani, A.; Wang, X.J.; Tang, X. Nrf2 signaling pathway: Pivotal roles in inflammation. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 585–597. [Google Scholar] [CrossRef]
- Tonelli, C.; Chio, I.I.C.; Tuveson, D.A. Transcriptional regulation by Nrf2. Antioxid. Redox Signal. 2018, 29, 1727–1745. [Google Scholar] [CrossRef] [Green Version]
- Reisman, S.A.; Aleksunes, L.M.; Klaassen, C.D. Oleanolic acid activates Nrf2 and protects from acetaminophen hepatotoxicity via Nrf2-dependent and Nrf2-independent processes. Biochem. Pharmacol. 2009, 77, 1273–1282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, M.; Tao, S.; De La Vega, M.R.; Jiang, T.; Wen, Q.; Park, S.L.; Zhang, D.D.; Wondrak, G.T. Nrf2-dependent suppression of azoxymethane/dextran sulfate sodium-induced colon carcinogenesis by the cinnamon-derived dietary factor cinnamaldehyde. Cancer Prev. Res. 2015, 8, 444–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, M.; Vemu, B.; Veenstra, J.; Petiwala, S.M.; Johnson, J.J. Carnosol, a dietary diterpene from rosemary (Rosmarinus officinalis) activates Nrf2 leading to sestrin 2 induction in colon cells. Integr. Mol. Med. 2018, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunnumakkara, A.B.; Bordoloi, D.; Padmavathi, G.; Monisha, J.; Roy, N.K.; Prasad, S.; Aggarwal, B.B. Curcumin, the golden nutraceutical: Multitargeting for multiple chronic diseases. Br. J. Pharmacol. 2017, 174, 1325–1348. [Google Scholar] [CrossRef] [Green Version]
- Yao, L.; Hou, G.; Wang, L.; Zuo, X.; Liu, Z. Protective effects of thymol on LPS-induced acute lung injury in mice. Microb. Pathog. 2018, 116, 8–12. [Google Scholar] [CrossRef]
- De la Vega, M.R.; Krajisnik, A.; Zhang, D.D.; Wondrak, G.T. Targeting NRF2 for improved skin barrier function and photoprotection: Focus on the achiote-derived apocarotenoid bixin. Nutrients 2017, 9, 1371. [Google Scholar] [CrossRef] [Green Version]
- Schadich, E.; Hlaváč, J.; Volná, T.; Varanasi, L.; Hajdúch, M.; Džubák, P. Effects of ginger phenylpropanoids and quercetin on Nrf2-ARE pathway in human BJ fibroblasts and HaCaT keratinocytes. Biomed. Res. Int. 2016, 2016. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.W.; Kim, M.J.; Shin, Y.; Jung, S.K.; Kim, Y.-J. Green pepper (Piper nigrum L.) extract suppresses oxidative stress and LPS-induced inflammation via regulation of JNK signaling pathways. Appl. Sci. 2020, 10, 2519. [Google Scholar] [CrossRef] [Green Version]
- Balstad, T.R.; Carlsen, H.; Myhrstad, M.C.W.; Kolberg, M.; Reiersen, H.; Gilen, L.; Ebihara, K.; Paur, I.; Blomhoff, R. Coffee, broccoli and spices are strong inducers of electrophile response element-dependent transcription in vitro and in vivo—Studies in electrophile response element transgenic mice. Mol. Nutr. Food Res. 2011, 55, 185–197. [Google Scholar] [CrossRef]
- Pall, M.L.; Levine, S. Nrf2, a master regulator of detoxification and also antioxidant, anti-inflammatory and other cytoprotective mechanisms, is raised by health promoting factors. Sheng Li Xue Bao 2015, 67, 1–18. [Google Scholar]
- Noda, Y.; Kneyuki, T.; Igarashi, K.; Mori, A.; Packer, L. Antioxidant activity of nasunin, an anthocyanin in eggplant peels. Toxicology 2000, 148, 119–123. [Google Scholar] [CrossRef]
- Reuland, D.J.; Khademi, S.; Castle, C.J.; Irwin, D.C.; McCord, J.M.; Miller, B.F.; Hamilton, K.L. Upregulation of phase II enzymes through phytochemical activation of Nrf2 protects cardiomyocytes against oxidant stress. Free Radic. Biol. Med. 2013, 56, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Huélamo, M.; Rodríguez-Morató, J.; Boronat, A.; de la Torre, R. Modulation of Nrf2 by olive oil and wine polyphenols and neuroprotection. Antioxidants 2017, 6, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senger, D.R.; Cao, S. Diabetic wound healing and activation of Nrf2 by herbal medicine. J. Nat. Sci. 2016, 2, e247. [Google Scholar] [PubMed]
- Kim, J.; Oh, J.; Averilla, J.N.; Kim, H.J.; Kim, J.S.; Kim, J.S. Grape peel extract and resveratrol inhibit wrinkle formation in mice model through activation of Nrf2/HO-1 signaling pathway. J. Food Sci. 2019, 84, 1600–1608. [Google Scholar] [CrossRef]
- Bowtell, J.; Kelly, V. Fruit-derived polyphenol supplementation for athlete recovery and performance. Sport. Med. 2019, 49, 3–23. [Google Scholar] [CrossRef] [Green Version]
- Surh, Y.J.; Kundu, J.K.; Na, H.K. Nrf2 as a master redox switch in turning on the cellular signaling involved in the induction of cytoprotective genes by some chemopreventive phytochemicals. Planta Med. 2008, 74, 1526–1539. [Google Scholar] [CrossRef] [Green Version]
- Robledinos-Antón, N.; Fernández-Ginés, R.; Manda, G.; Cuadrado, A. Activators and inhibitors of NRF2: A review of their potential for clinical development. Oxid. Med. Cell. Longev. 2019, 2019, 9372182. [Google Scholar] [CrossRef]
- Copetti, C.; Franco, F.W.; Machado, E.D.; Soquetta, M.B.; Quatrin, A.; Ramos, V.D.; Moreira, J.C.; Emanuelli, T.; Sautter, C.K.; Penna, N.G. Acute consumption of bordo grape juice and wine improves serum antioxidant status in healthy individuals and inhibits reactive oxygen species production in human neuron-like cells. J. Nutr. Metab. 2018, 2018, 4384012. [Google Scholar] [CrossRef] [Green Version]
- Heron, M. National Vital Statistics Reports Volume 68, Number 6, June 24, 2019, Deaths: Leading Causes for 2017; U.S. Department of Health & Human Services: Hyattsville, MD, USA, 2019.
- Yuan, L.; Liu, J.; Zhen, J.; Xu, Y.; Chen, S.; Halm-Lutterodt, N.V.; Xiao, R. Vegetable and fruit juice enhances antioxidant capacity and regulates antioxidant gene expression in rat liver, brain and colon. Genet. Mol. Biol. 2017, 40, 134–141. [Google Scholar] [CrossRef] [Green Version]
- Rubió, L.; Motilva, M.J.; Romero, M.P. Recent advances in biologically active compounds in herbs and spices: A review of the most effective antioxidant and anti-inflammatory active principles. Crit. Rev. Food Sci. Nutr. 2013, 53, 943–953. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, C.R.B.; Cadavid, C.O.M.; Carmona, L.; Peña, L.; De Paula, O.R. Pasteurized orange juice rich in carotenoids protects caenorhabditis elegans against oxidative stress and β-amyloid toxicity through direct and indirect mechanisms. Oxid. Med. Cell. Longev. 2019, 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bar-Ya’akov, I.; Tian, L.; Amir, R.; Holland, D. Primary metabolites, anthocyanins, and hydrolyzable tannins in the pomegranate fruit. Front. Plant Sci. 2019, 10, 620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Husain, H.; Latief, U.; Ahmad, R. Pomegranate action in curbing the incidence of liver injury triggered by Diethylnitrosamine by declining oxidative stress via Nrf2 and NFκB regulation. Sci. Rep. 2018, 8, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; He, S.S.; Yin, P.; Li, D.Y.; Mei, C.; Yu, X.H.; Shi, Y.R.; Jiang, L.S.; Liu, F.H. Punicalagin induces Nrf2 translocation and HO-1 expression via PI3K/Akt, protecting rat intestinal epithelial cells from oxidative stress. Int. J. Hyperth. 2016, 32, 465–473. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Goel, N. Phenolic acids: Natural versatile molecules with promising therapeutic applications. Biotechnol. Rep. 2019, 24, e00370. [Google Scholar] [CrossRef]
- Tang, J.S.; Vissers, M.C.M.; Anderson, R.F.; Sreebhavan, S.; Bozonet, S.M.; Scheepens, A.; Melton, L.D. Bioavailable blueberry-derived phenolic acids at physiological concentrations enhance Nrf2-regulated antioxidant responses in human vascular endothelial cells. Mol. Nutr. Food Res. 2018, 62, 1–34. [Google Scholar] [CrossRef]
- Soyalan, B.; Minn, J.; Schmitz, H.J.; Schrenk, D.; Will, F.; Dietrich, H.; Baum, M.; Eisenbrand, G.; Janzowski, C. Apple juice intervention modulates expression of ARE-dependent genes in rat colon and liver. Eur. J. Nutr. 2011, 50, 135–143. [Google Scholar] [CrossRef]
- McClatchey, W. From Polynesian healers to health food stores: Changing perspectives of Morinda citrifolia (Rubiaceae). Integr. Cancer Ther. 2003, 1, 110–120. [Google Scholar] [CrossRef]
- Chen, J.; Shi, X.; Chen, Y.; Liang, H.; Cheng, C.; He, Q. Neuroprotective effects of chloroform and aqueous fractions of noni juice against t-butyl hydroperoxide-induced oxidative damage in SH-SY5Y cells. Food Nutr. Res. 2018, 62, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Manavalan, A.; Qader, M.; Wu, X.; Cao, S. Nrf2 activation by Morinda citrifolia L. (Noni) fruit juices. World J. Tradit. Chinese Med. 2020. In press. [Google Scholar]
- Masci, A.; Mattioli, R.; Costantino, P.; Baima, S.; Morelli, G.; Punzi, P.; Giordano, C.; Pinto, A.; Donini, L.M.; D’Erme, M.; et al. Neuroprotective effect of brassica oleracea sprouts crude juice in a cellular model of alzheimer’s disease. Oxid. Med. Cell. Longev. 2015, 2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawal, A.O.; Lawal, A.F.; Ologundudu, A.; Adeniran, O.Y.; Omonkhua, A.; Obi, F. Antioxidant effects of heated garlic juice on cadmium-induced liver damage in rats as compared to ascorbic acid. J. Toxicol. Sci. 2011, 36, 549–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krajka-Kuźniak, V.; Szaefer, H.; Bartoszek, A.; Baer-Dubowska, W. Modulation of rat hepatic and kidney phase II enzymes by cabbage juices: Comparison with the effects of indole-3-carbinol and phenethyl isothiocyanate. Br. J. Nutr. 2011, 105, 816–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krajka-Kuźniak, V.; Paluszczak, J.; Szaefer, H.; Baer-Dubowska, W. Betanin, a beetroot component, induces nuclear factor erythroid-2-related factor 2-mediated expression of detoxifying/antioxidant enzymes in human liver cell lines. Br. J. Nutr. 2013, 110, 2138–2149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.; Li, X.; Li, G.; Dai, B.; Tan, W. Actinidia chinensis Planch. Improves the indices of antioxidant and anti-inflammation status of type 2 diabetes mellitus by activating keap1 and Nrf2 via the upregulation of MicroRNA-424. Oxid. Med. Cell. Longev. 2017, 2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lukacs, P. Inventing Wine: A New History of One of the World’s Most Ancient Pleasures, 1st ed.; W.W. Nortan & Company, Inc.: New York, NY, USA, 2012; ISBN 9780393239645. [Google Scholar]
- Klatsky, A.L. Alcohol and cardiovascular diseases. Expert Rev. Cardiovasc. Ther. 2009, 7, 499–506. [Google Scholar] [CrossRef]
- Del Pino-García, R.; González-SanJosé, M.L.; Rivero-Pérez, M.D.; García-Lomillo, J.; Muñiz, P. The effects of heat treatment on the phenolic composition and antioxidant capacity of red wine pomace seasonings. Food Chem. 2017, 221, 1723–1732. [Google Scholar] [CrossRef] [Green Version]
- Guaita, M.; Bosso, A. Polyphenolic characterization of grape skins and seeds of four Italian red cultivars at harvest and after fermentative maceration. Foods 2019, 8, 395. [Google Scholar] [CrossRef] [Green Version]
- Nile, S.H.; Kim, S.H.; Ko, E.Y.; Park, S.W. Polyphenolic contents and antioxidant properties of different grape (V. vinifera, V. labrusca, and V. hybrid) cultivars. Biomed. Res. Int. 2013, 2013. [Google Scholar] [CrossRef] [Green Version]
- Brighenti, E.; Casagrande, K.; Cardoso, P.Z.; da Silveira Pasa, M.; Ciotta, M.N.; Brighenti, A.F. Total polyphenols contents in different grapevine varieties in highlands of southern brazil. BIO Web Conf. 2017, 9, 01024. [Google Scholar] [CrossRef] [Green Version]
- Narayanan, S.V.; Dave, K.R.; Saul, I.; Perez-Pinzon, M.A. Resveratrol preconditioning protects against cerebral ischemic injury via nuclear erythroid 2-related factor 2. Stroke 2015, 46, 1626–1632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, J.; Fan, C.; Chen, N.; Huang, J.; Yang, Q. Resveratrol pretreatment attenuates cerebral ischemic injury by upregulating expression of transcription factor Nrf2 and HO-1 in Rats. Neurochem. Res. 2011, 36, 2352–2362. [Google Scholar] [CrossRef] [PubMed]
- Ghanim, H.; Sia, C.L.; Korzeniewski, K.; Lohano, T.; Abuaysheh, S.; Marumganti, A.; Chaudhuri, A.; Dandona, P. A resveratrol and polyphenol preparation suppresses oxidative and inflammatory stress response to a high-fat, high-carbohydrate meal. J. Clin. Endocrinol. Metab. 2011, 96, 1409–1414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, C.; Cheng, W.; Yu, P.; Wang, L.; Zhou, L.; Zeng, L.; Yang, Q. Resveratrol pretreatment attenuates injury and promotes proliferation of neural stem cells following oxygen-glucose deprivation/reoxygenation by upregulating the expression of Nrf2, HO-1 and NQO1 in vitro. Mol. Med. Rep. 2016, 14, 3646–3654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bishayee, A.; Barnes, K.F.; Bhatia, D.; Darvesh, A.S.; Carroll, R.T. Resveratrol suppresses oxidative stress and inflammatory response in diethylnitrosamine-initiated rat hepatocarcinogenesis. Cancer Prev. Res. 2010, 3, 753–763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurusamy, N.; Ray, D.; Lekli, I.; Das, D.K. Red wine antioxidant resveratrol-modified cardiac stem cells regenerate infarcted myocardium. J. Cell. Mol. Med. 2010, 14, 2235–2239. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Xu, X.; Tao, Z.; Wang, X.J.; Pan, Y. Resveratrol dimers, nutritional components in grape wine, are selective ROS scavengers and weak Nrf2 activators. Food Chem. 2015, 173, 218–223. [Google Scholar] [CrossRef]
- Zghonda, N.; Yoshida, S.; Araki, M.; Kusunoki, M.; Mliki, A.; Ghorbel, A.; Miyazaki, H. Greater effectiveness of ε-viniferin in red wine than its monomer resveratrol for inhibiting vascular smooth muscle cell proliferation and migration. Biosci. Biotechnol. Biochem. 2011, 75, 1259–1267. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Yue, Y.; Li, J.; Li, Z.; Li, X.; Niu, Y.; Xiang, J.; Ding, H. Procyanidin B2 attenuates neurological deficits and blood-brain barrier disruption in a rat model of cerebral ischemia. Mol. Nutr. Food Res. 2015, 59, 1930–1941. [Google Scholar] [CrossRef]
- Bahia, P.K.; Rattray, M.; Williams, R.J. Dietary flavonoid (-)epicatechin stimulates phosphatidylinositol 3-kinase-dependent anti-oxidant response element activity and up-regulates glutathione in cortical astrocytes. J. Neurochem. 2008, 106, 2194–2204. [Google Scholar] [CrossRef] [PubMed]
- Lan, X.; Han, X.; Li, Q.; Wang, J. (−)-Epicatechin, a natural flavonoid compound, protects astrocytes against hemoglobin toxicity via Nrf2 and AP-1 signaling pathways. Mol. Neurobiol. 2017, 54, 7898–7907. [Google Scholar] [CrossRef] [Green Version]
- Dong, F.; Wang, S.; Wang, Y.; Yang, X.; Jiang, J.; Wu, D.; Qu, X.; Fan, H.; Yao, R. Quercetin ameliorates learning and memory via the Nrf2-ARE signaling pathway in d-galactose-induced neurotoxicity in mice. Biochem. Biophys. Res. Commun. 2017, 491, 636–641. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Ramiro, I.; Ramos, S.; Bravo, L.; Goya, L.; Martín, M.Á. Procyanidin B2 induces Nrf2 translocation and glutathione S-transferase P1 expression via ERKs and p38-MAPK pathways and protect human colonic cells against oxidative stress. Eur. J. Nutr. 2012, 51, 881–892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weng, C.J.; Chen, M.J.; Yeh, C.T.; Yen, G.C. Hepatoprotection of quercetin against oxidative stress by induction of metallothionein expression through activating MAPK and PI3K pathways and enhancing Nrf2 DNA-binding activity. New Biotechnol. 2011, 28, 767–777. [Google Scholar] [CrossRef] [PubMed]
- Chu, C.; Gao, X.; Li, X.; Zhang, X.; Ma, R.; Jia, Y.; Li, D.; Wang, D.; Xu, F. Involvement of estrogen receptor-α in the activation of Nrf2-antioxidative signaling pathways by silibinin in pancreatic β-cells. Biomol. Ther. 2020, 28, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Son, Y.; Byun, S.J.; Pae, H.O. Involvement of heme oxygenase-1 expression in neuroprotection by piceatannol, a natural analog and a metabolite of resveratrol, against glutamate-mediated oxidative injury in HT22 neuronal cells. Amino Acids 2013, 45, 393–401. [Google Scholar] [CrossRef]
- Erukainure, O.L.; Ijomone, O.M.; Sanni, O.; Aschner, M.; Islam, M.S. Type 2 diabetes induced oxidative brain injury involves altered cerebellar neuronal integrity and elemental distribution, and exacerbated Nrf2 expression: Therapeutic potential of raffia palm (Raphia hookeri) wine. Metab. Brain Dis. 2019, 34, 1385–1399. [Google Scholar] [CrossRef]
- Jeon, M.; Rahman, N.; Kim, Y.S. Cytoprotective effect of makgeolli lees on paraquat induced oxidative stress in A549 cells via activation of NRF2 and antioxidant genes. J. Microbiol. Biotechnol. 2015, 26, 277–286. [Google Scholar] [CrossRef]
- Zhai, X.; Chi, J.; Tang, W.; Ji, Z.; Zhao, F.; Jiang, C.; Lv, H.; Guo, H. Yellow wine polyphenolic compounds inhibit matrix metalloproteinase-2, -9 expression and improve atherosclerotic plaque in LDL-receptor–knockout mice. J. Pharmacol. Sci. 2014, 125, 132–141. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.; Zhang, J.; Ni, T.; Lin, N.; Meng, L.; Gao, F.; Luo, H.; Liu, X.; Chi, J.; Guo, H. Yellow wine polyphenolic compounds prevents doxorubicin-induced cardiotoxicity through activation of the Nrf2 signalling pathway. J. Cell. Mol. Med. 2019, 23, 6034–6047. [Google Scholar] [CrossRef] [Green Version]
- Cai, Y.-S.; Xu, J.; Chen, M.; Wang, D.; Yang, Y.; Manavalan, A.; Wu, X.; Liu, Y.; Cao, S. Compound analysis of jing liqueur and Nrf2 activation by jing liqueur—One of the most popular beverages in China. Beverages 2019, 6, 1. [Google Scholar] [CrossRef] [Green Version]
- Lakenbrink, C.; Lapczynski, S.; Maiwald, B.; Engelhardt, U.H. Flavonoids and other polyphenols in consumer brews of tea and other caffeinated beverages. J. Agric. Food Chem. 2000, 48, 2848–2852. [Google Scholar] [CrossRef]
- Ackar, D.; Valek Lendić, K.; Valek, M.; Šubarić, D.; Miličević, B.; Babić, J.; Nedić, I. cocoa polyphenols: Can we consider cocoa and chocolate as potential functional food? J. Chem. 2013, 2013, 289392. [Google Scholar] [CrossRef]
- Richelle, M.; Tavazzi, I.; Offord, E. Comparison of the antioxidant activity of commonly consumed polyphenolic beverages (coffee, cocoa, and tea) prepared per cup serving. J. Agric. Food Chem. 2001, 49, 3438–3442. [Google Scholar] [CrossRef]
- Moreira, A.S.P.; Nunes, F.M.; Domingues, M.R.; Coimbra, M.A. Coffee melanoidins: Structures, mechanisms of formation and potential health impacts. Food Funct. 2012, 3, 903–915. [Google Scholar] [CrossRef]
- Shi, A.; Shi, H.; Wang, Y.; Liu, X.; Cheng, Y.; Li, H.; Zhao, H.; Wang, S.; Dong, L. Activation of Nrf2 pathway and inhibition of NLRP3 inflammasome activation contribute to the protective effect of chlorogenic acid on acute liver injury. Int. Immunopharmacol. 2018, 54, 125–130. [Google Scholar] [CrossRef]
- Yao, J.; Peng, S.; Xu, J.; Fang, J. Reversing ROS-mediated neurotoxicity by chlorogenic acid involves its direct antioxidant activity and activation of Nrf2-ARE signaling pathway. BioFactors 2019, 45, 616–626. [Google Scholar] [CrossRef]
- Liang, N.; Kitts, D.D. Amelioration of oxidative stress in Caco-2 cells treated with pro-inflammatory proteins by chlorogenic acid isomers via activation of the Nrf2-Keap1-ARE-signaling pathway. J. Agric. Food Chem. 2018, 66, 11008–11017. [Google Scholar] [CrossRef]
- Trinh, K.; Andrews, L.; Krause, J.; Hanak, T.; Lee, D.; Gelb, M.; Pallanck, L. Decaffeinated coffee and nicotine-free tobacco provide neuroprotection in Drosophila models of Parkinson’s disease through an NRF2-dependent mechanism. J. Neurosci. 2010, 30, 5525–5532. [Google Scholar] [CrossRef]
- Ren, Y.; Wang, C.; Xu, J.; Wang, S. Cafestol and kahweol: A review on their bioactivities and pharmacological properties. Int. J. Mol. Sci. 2019, 20, 4328. [Google Scholar] [CrossRef] [Green Version]
- Higgins, L.G.; Cavin, C.; Itoh, K.; Yamamoto, M.; Hayes, J.D. Induction of cancer chemopreventive enzymes by coffee is mediated by transcription factor Nrf2. Evidence that the coffee-specific diterpenes cafestol and kahweol confer protection against acrolein. Toxicol. Appl. Pharmacol. 2008, 226, 328–337. [Google Scholar] [CrossRef]
- Cavin, C.; Holzhaeuser, D.; Scharf, G.; Constable, A.; Huber, W.W.; Schilter, B. Cafestol and kahweol, two coffee specific diterpenes with anticarcinogenic activity. Food Chem. Toxicol. 2002, 40, 1155–1163. [Google Scholar] [CrossRef]
- Hao, W.R.; Sung, L.C.; Chen, C.C.; Hong, H.J.; Liu, J.C.; Chen, J.J. Cafestol activates nuclear factor erythroid-2 related factor 2 and inhibits urotensin ii-induced cardiomyocyte hypertrophy. Am. J. Chin. Med. 2019, 47, 337–350. [Google Scholar] [CrossRef]
- Tsai, Y.T.; Sung, L.C.; Haw, W.R.; Chen, C.C.; Huang, S.F.; Liu, J.C.; Cheng, T.H.; Chen, P.Y.; Loh, S.H.; Tsai, C.S. Cafestol, a coffee diterpene, inhibits urotensin II-induced interleukin-8 expression in human umbilical vein endothelial cells. Eur. J. Pharmacol. 2018, 820, 106–112. [Google Scholar] [CrossRef]
- Fukuma, Y.; Sakai, E.; Nishishita, K.; Okamoto, K.; Tsukuba, T. Cafestol has a weaker inhibitory effect on osteoclastogenesis than kahweol and promotes osteoblast differentiation. BioFactors 2015, 41, 222–231. [Google Scholar] [CrossRef]
- Volz, N.; Boettler, U.; Winkler, S.; Teller, N.; Schwarz, C.; Bakuradze, T.; Eisenbrand, G.; Haupt, L.; Griffiths, L.R.; Stiebitz, H.; et al. Effect of coffee combining green coffee bean constituents with typical roasting products on the Nrf2/ARE pathway in vitro and in vivo. J. Agric. Food Chem. 2012, 60, 9631–9641. [Google Scholar] [CrossRef]
- Shen, J.; Wang, G.; Zuo, J. Caffeic acid inhibits HCV replication via induction of IFNα antiviral response through p62-mediated Keap1/Nrf2 signaling pathway. Antivir. Res. 2018, 154, 166–173. [Google Scholar] [CrossRef]
- Fratantonio, D.; Speciale, A.; Canali, R.; Natarelli, L.; Ferrari, D.; Saija, A.; Virgili, F.; Cimino, F. Low nanomolar caffeic acid attenuates high glucose-induced endothelial dysfunction in primary human umbilical-vein endothelial cells by affecting NF-κB and Nrf2 pathways. BioFactors 2017, 43, 54–62. [Google Scholar] [CrossRef]
- Nilnumkhum, A.; Kanlaya, R.; Yoodee, S.; Thongboonkerd, V. Caffeine inhibits hypoxia-induced renal fibroblast activation by antioxidant mechanism. Cell Adhes. Migr. 2019, 13, 259–271. [Google Scholar] [CrossRef] [Green Version]
- Rowley, T.J.; Bitner, B.F.; Ray, J.D.; Lathen, D.R.; Smithson, A.T.; Dallon, B.W.; Plowman, C.J.; Bikman, B.T.; Hansen, J.M.; Dorenkott, M.R.; et al. Monomeric cocoa catechins enhance β-cell function by increasing mitochondrial respiration. J. Nutr. Biochem. 2017, 49, 30–41. [Google Scholar] [CrossRef]
- Cordero-Herrera, I.; Martín, M.A.; Goya, L.; Ramos, S. Cocoa flavonoids protect hepatic cells against high-glucose-induced oxidative stress: Relevance of MAPKs. Mol. Nutr. Food Res. 2015, 59, 597–609. [Google Scholar] [CrossRef] [Green Version]
- Cheng, T.; Wang, W.; Li, Q.; Han, X.; Xing, J.; Qi, C.; Lan, X.; Wan, J.; Potts, A.; Guan, F.; et al. Cerebroprotection of flavanol (-)-epicatechin after traumatic brain injury via Nrf2-dependent and-independent pathways. Free Radic. Biol. Med. 2016, 92, 15–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, Z.A.; Li, R.C.; Ahmad, A.S.; Kensler, T.W.; Yamamoto, M.; Biswal, S.; Doré, S. The flavanol (-)-epicatechin prevents stroke damage through the Nrf2/HO1 pathway. J. Cereb. Blood Flow Metab. 2010, 30, 1951–1961. [Google Scholar] [CrossRef] [Green Version]
- Funakoshi-Tago, M.; Nonaka, Y.; Tago, K.; Takeda, M.; Ishihara, Y.; Sakai, A.; Matsutaka, M.; Kobata, K.; Tamura, H. Pyrocatechol, a component of coffee, suppresses LPS-induced inflammatory responses by inhibiting NF-κB and activating Nrf2. Sci. Rep. 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Boettler, U.; Sommerfeld, K.; Volz, N.; Pahlke, G.; Teller, N.; Somoza, V.; Lang, R.; Hofmann, T.; Marko, D. Coffee constituents as modulators of Nrf2 nuclear translocation and ARE (EpRE)-dependent gene expression. J. Nutr. Biochem. 2011, 22, 426–440. [Google Scholar] [CrossRef]
- Liang, N.; Dupuis, J.H.; Yada, R.Y.; Kitts, D.D. Chlorogenic acid isomers directly interact with Keap 1-Nrf2 signaling in Caco-2 cells. Mol. Cell. Biochem. 2019, 457. [Google Scholar] [CrossRef] [Green Version]
- Fürstenau, C.R.; de Souza, I.C.C.; de Oliveira, M.R. The effects of kahweol, a diterpene present in coffee, on the mitochondria of the human neuroblastoma SH-SY5Y cells exposed to hydrogen peroxide. Toxicol. Vitro 2019, 61. [Google Scholar] [CrossRef]
- Hwang, Y.P.; Jeong, H.G. The coffee diterpene kahweol induces heme oxygenase-1 via the PI3K and p38/Nrf2 pathway to protect human dopaminergic neurons from 6-hydroxydopamine-derived oxidative stress. FEBS Lett. 2008, 582, 2655–2662. [Google Scholar] [CrossRef] [Green Version]
- Chang, W.-C.; Wu, S.-C.; Xu, K.-D.; Liao, B.-C.; Wu, J.-F.; Cheng, A.-S. Scopoletin protects against methylglyoxal-induced hyperglycemia and insulin resistance mediated by suppression of advanced glycation endproducts (AGEs) generation and anti-glycation. Molecules 2015, 20, 2786–2801. [Google Scholar] [CrossRef] [Green Version]
- Kumar, H.; Kim, I.S.; More, S.V.; Kim, B.W.; Choi, D.K. Natural product-derived pharmacological modulators of Nrf2/ARE pathway for chronic diseases. Nat. Prod. Rep. 2014, 31, 109–139. [Google Scholar] [CrossRef]
- Kim, W.D.; Kim, Y.W.; Cho, I.J.; Lee, C.H.; Kim, S.G. E-cadherin inhibits nuclear accumulation of Nrf2: Implications for chemoresistance of cancer cells. J. Cell Sci. 2012, 125, 1284–1295. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.D.; Chapman, E. The role of natural products in revealing NRF2 function. Nat. Prod. Rep. 2020, 37, 797–826. [Google Scholar] [CrossRef]
- Lin, H.; Qiao, Y.; Yang, H.; Nan, Q.; Qu, W.; Feng, F.; Liu, W.; Chen, Y.; Sun, H. Small molecular Nrf2 inhibitors as chemosensitizers for cancer therapy. Future Med. Chem. 2020, 12, 243–267. [Google Scholar] [CrossRef]
- Tarumoto, T.; Nagai, T.; Ohmine, K.; Miyoshi, T.; Nakamura, M.; Kondo, T.; Mitsugi, K.; Nakano, S.; Muroi, K.; Komatsu, N.; et al. Ascorbic acid restores sensitivity to imatinib via suppression of Nrf2-dependent gene expression in the imatinib-resistant cell line. Exp. Hematol. 2004, 32, 375–381. [Google Scholar] [CrossRef] [PubMed]
- Gao, A.M.; Ke, Z.P.; Wang, J.N.; Yang, J.Y.; Chen, S.Y.; Chen, H. Apigenin sensitizes doxorubicin-resistant hepatocellular carcinoma BEL-7402/ADM cells to doxorubicin via inhibiting PI3K/Akt/Nrf2 pathway. Carcinogenesis 2013, 34, 1806–1814. [Google Scholar] [CrossRef]
- Kweon, M.H.; Adhami, V.M.; Lee, J.S.; Mukhtar, H. Constitutive overexpression of Nrf2-dependent heme oxygenase-1 in A549 cells contributes to resistance to apoptosis induced by epigallocatechin 3-gallate. J. Biol. Chem. 2006, 281, 33761–33772. [Google Scholar] [CrossRef] [Green Version]
- Xiu, J.W.; Hayes, J.D.; Henderson, C.J.; Wolf, C.R. Identification of retinoic acid as an inhibitor of transcription factor Nrf2 through activation of retinoic acid receptor alpha. Proc. Natl. Acad. Sci. USA 2007, 104, 19589–19594. [Google Scholar] [CrossRef] [Green Version]
- Debelo, H.; Novotny, J.A.; Ferruzzi, M.G. Vitamin A. Adv. Nutr. 2017, 8, 992–994. [Google Scholar] [CrossRef] [Green Version]
- Tang, X.; Wang, H.; Fan, L.; Wu, X.; Xin, A.; Ren, H.; Wang, X.J. Luteolin inhibits Nrf2 leading to negative regulation of the Nrf2/ARE pathway and sensitization of human lung carcinoma A549 cells to therapeutic drugs. Free Radic. Biol. Med. 2011, 50, 1599–1609. [Google Scholar] [CrossRef]
- Lin, Y.; Shi, R.; Wang, X.; Shen, H.-M. Luteolin, a Flavonoid with Potential for Cancer Prevention and Therapy. Curr. Cancer Drug Targets 2008, 8, 634–646. [Google Scholar] [CrossRef] [PubMed]
- Arlt, A.; Sebens, S.; Krebs, S.; Geismann, C.; Grossmann, M.; Kruse, M.L.; Schreiber, S.; Schäfer, H. Inhibition of the Nrf2 transcription factor by the alkaloid trigonelline renders pancreatic cancer cells more susceptible to apoptosis through decreased proteasomal gene expression and proteasome activity. Oncogene 2013, 32, 4825–4835. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Production of excess or insufficient amount of RONS in the biological systems will leads to oxidative stress or reductive stress, respectively, and related diseases. Though RONS cause detrimental effects to the biological systems, they are important as cellular signaling inducers. As a result, RONS could activate transcription factors like Nrf2 and induce antioxidant genes. This process will restore the equilibrium of reactive species in biological systems.
Figure 1.
Production of excess or insufficient amount of RONS in the biological systems will leads to oxidative stress or reductive stress, respectively, and related diseases. Though RONS cause detrimental effects to the biological systems, they are important as cellular signaling inducers. As a result, RONS could activate transcription factors like Nrf2 and induce antioxidant genes. This process will restore the equilibrium of reactive species in biological systems.

Figure 2.
Natural compounds (phytochemicals) responsible for the activation of Nrf2. (Glu- glucose).
Figure 2.
Natural compounds (phytochemicals) responsible for the activation of Nrf2. (Glu- glucose).
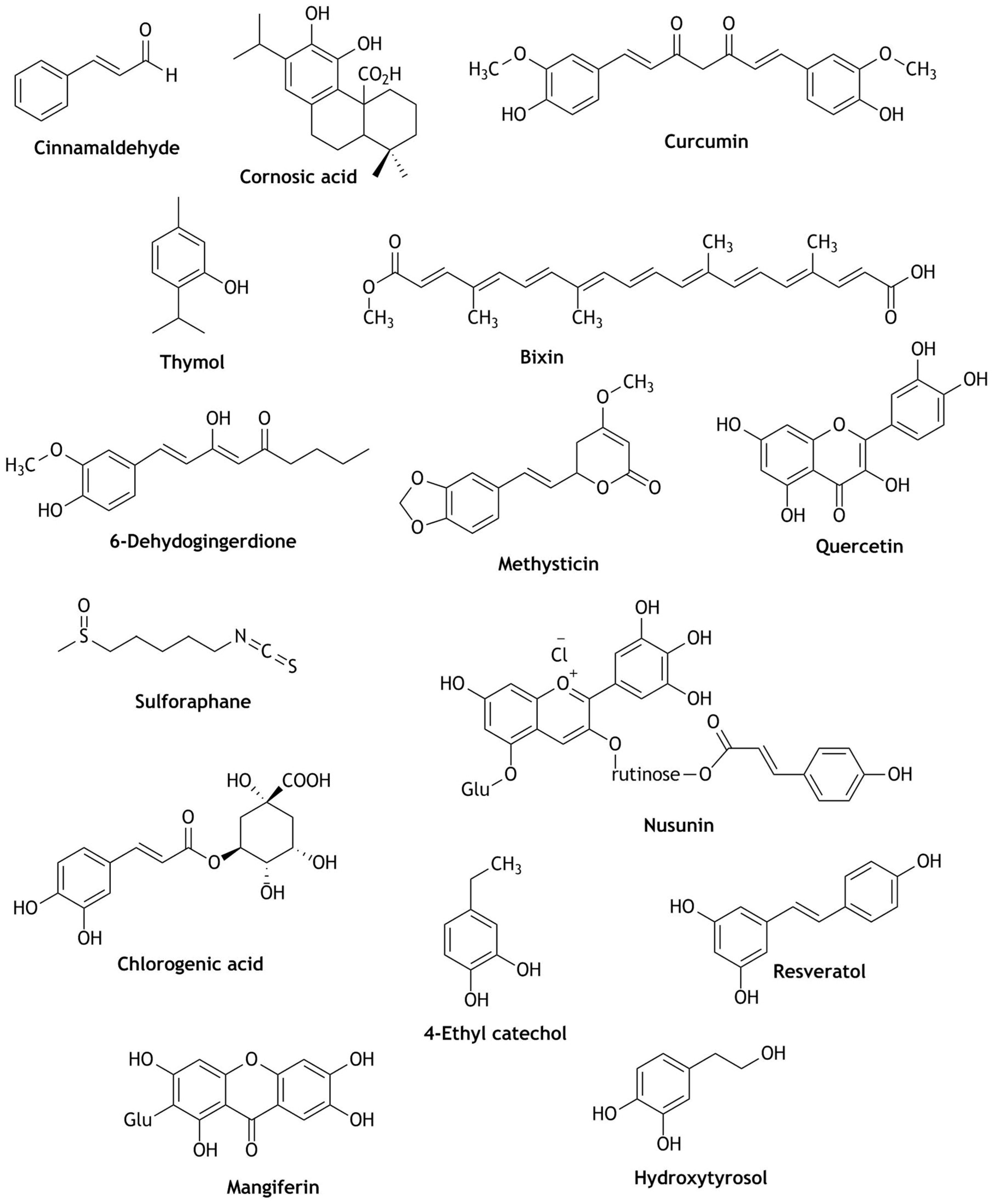
Figure 3.
Nrf2 activators present in juices, wines, coffee, and cocoa. In cytosol, Nrf2 separates from the Keap1, and then translocates into the nucleus. In the nucleus, Nrf2 induces the production of smaller proteins like sMaf, binds to phase 2 of the antioxidant response elements (ARE) in the DNA promoter region, and initiates the expression of antioxidant genes.
Figure 3.
Nrf2 activators present in juices, wines, coffee, and cocoa. In cytosol, Nrf2 separates from the Keap1, and then translocates into the nucleus. In the nucleus, Nrf2 induces the production of smaller proteins like sMaf, binds to phase 2 of the antioxidant response elements (ARE) in the DNA promoter region, and initiates the expression of antioxidant genes.

Figure 4.
Exponential interest in the scientific community towards searching for Nrf2 activators from natural phytochemicals during the last 20 years (2001 to 2019). PubMed database shows. that in the first six months of 2020, there were 35 articles published.
Figure 4.
Exponential interest in the scientific community towards searching for Nrf2 activators from natural phytochemicals during the last 20 years (2001 to 2019). PubMed database shows. that in the first six months of 2020, there were 35 articles published.

Figure 5.
Structures of phytochemicals identified in fruit juices as Nrf2 activators.
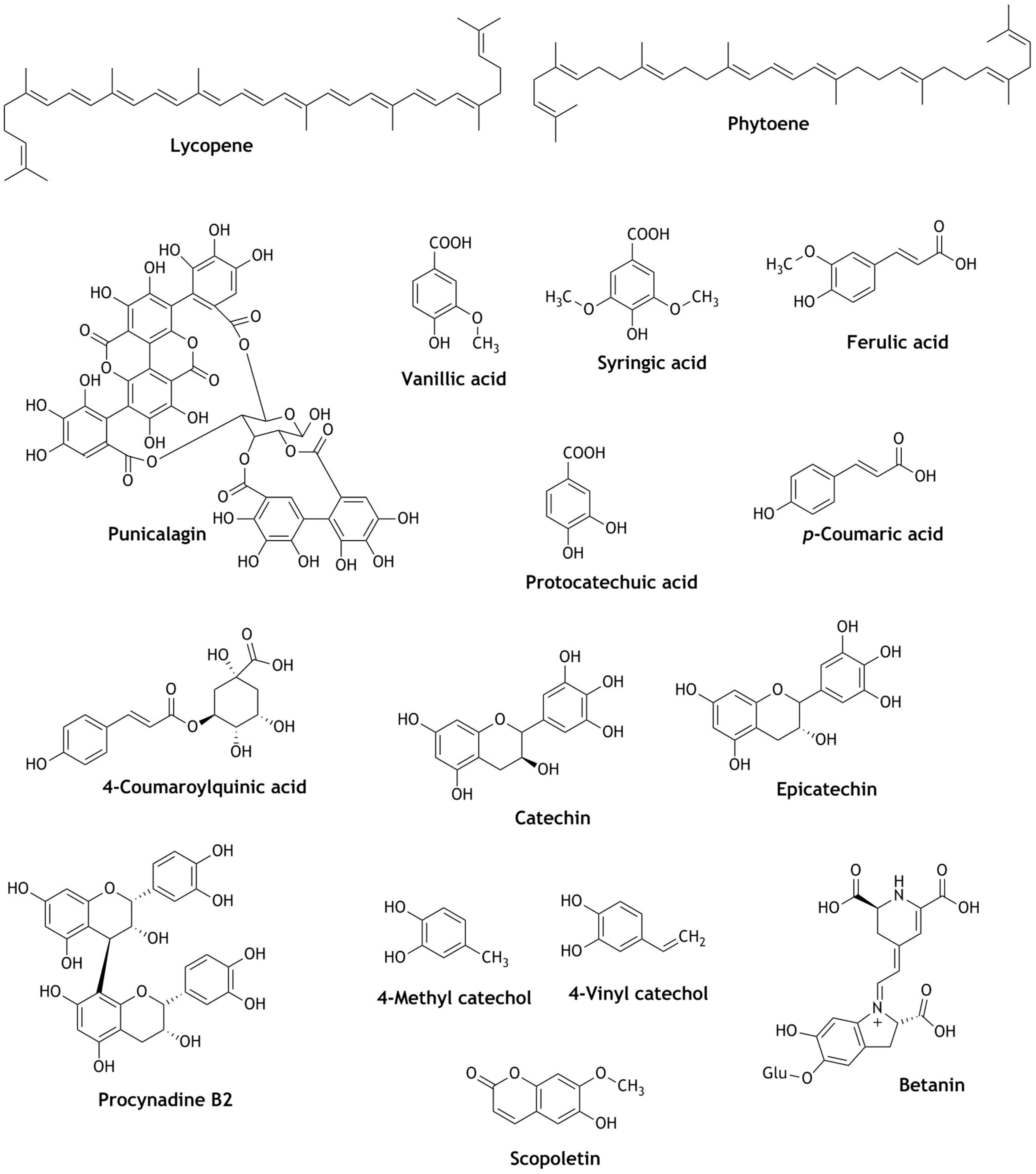
Figure 6.
Structures of phytochemicals identified in wine as Nrf2 activators.

Figure 7.
Structures of phytochemicals identified in coffee and cocoa as Nrf2 activators.
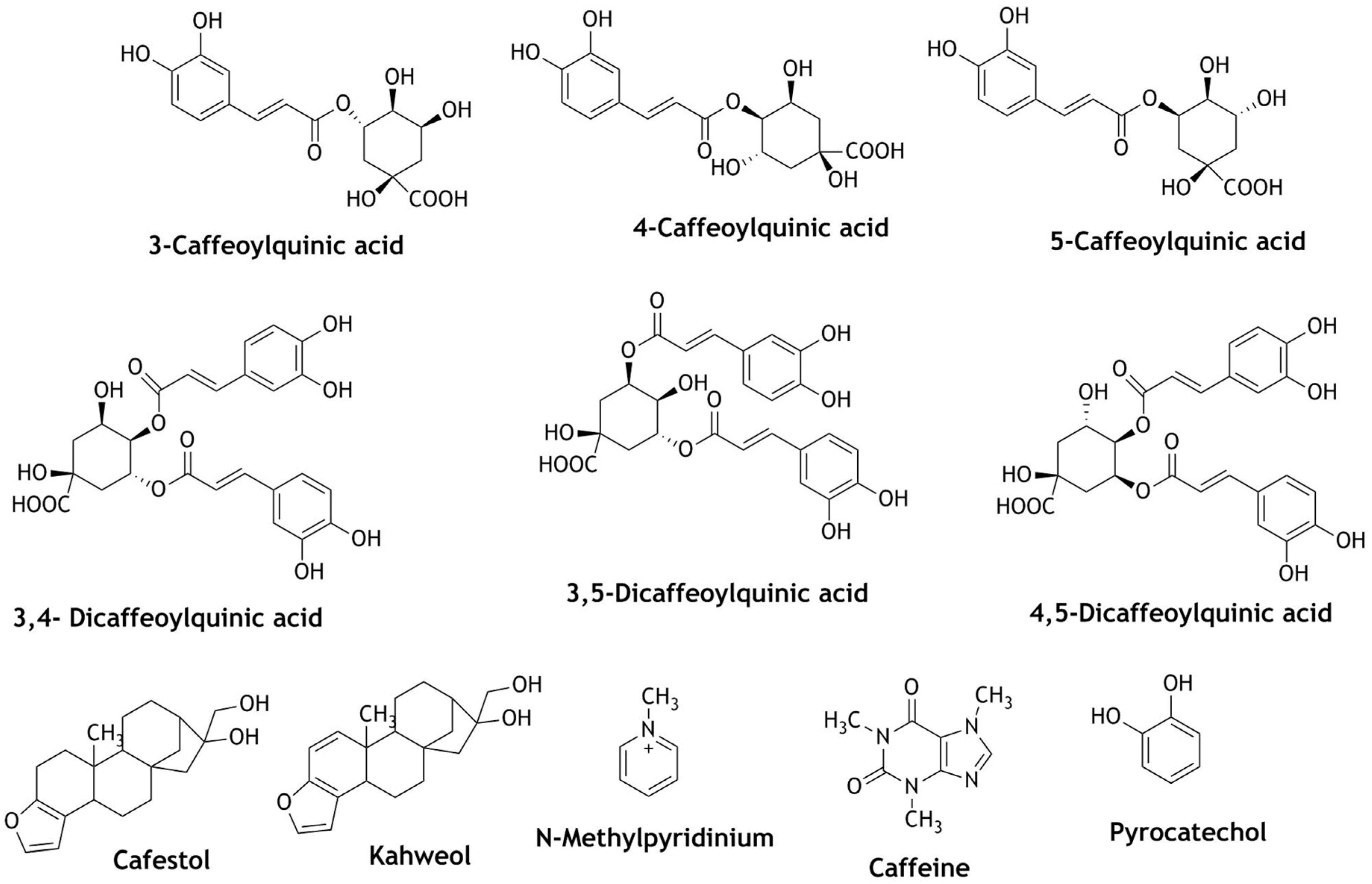
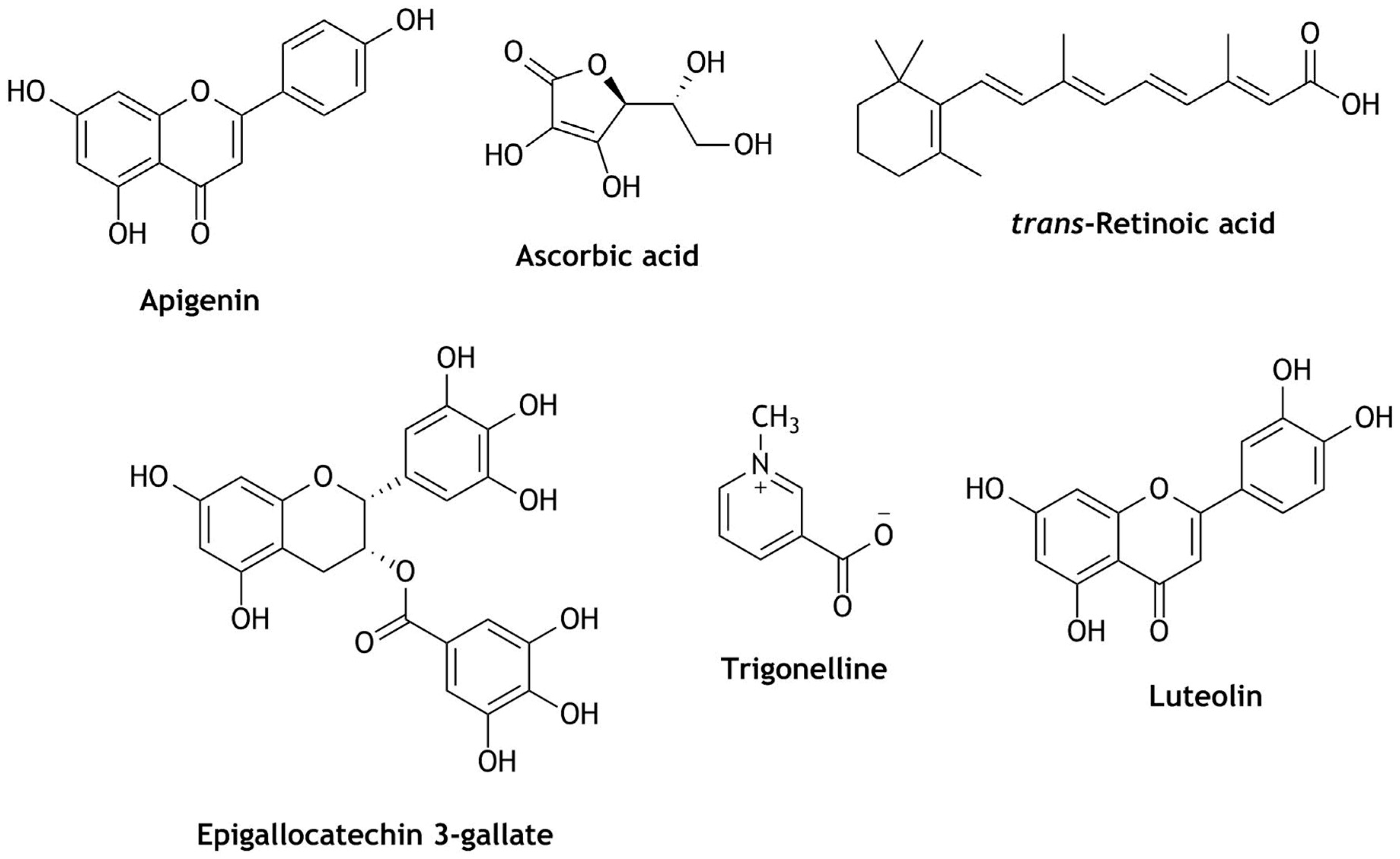
Figure 8.
Structures of phytochemicals identified as Nrf2 inhibitors from coffee, juices, and tea.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Active reactive oxygen and nitrogen species (RONS) in biological systems.
| Reactive Oxygen Species (ROS) | Reactive Nitrogen Species (RNS) |
|---|---|
| Superoxide anions (O2•−) | Nitric oxide (NO•) |
| Hydroxyl radical (•OH) | Nitrogen dioxide (NO2•) |
| Peroxyl radical (RO2•) | Peroxy-nitrite (ONOO−) |
| Alkoxyl radical (RO•) | Nitroxyl anion (NO−) |
| Hydroperoxyl radical (HO2•) | Nitrous oxide (N2O) |
| Hydrogen peroxide (H2O2) | Nitrogen oxides (NO2, N2O4) |
| Hypochlorous acid (HOCl) | Nitrous acid (HNO2) |
| Hypobromus acid (HOBr) | Peroxynitrous acid (HNO3) |
| S-nitrosothiols (RSNO) | |
| Fe-dinitrosyl complexes |
Table 2.
List of phytochemicals present in fruit juices as Nrf2 activators.
| Sources | Bioactive Natural Compound/s | Study Observations | Therapeutic Indications | Reference |
|---|---|---|---|---|
| Apple juice | Chlorogenic acid, 4-coumaroylquinic acid, epicatechin, procyanidin B2 | In distal colon of mice GPX2, GSR, CAT and in liver GPX1 and NOQ-1 mRNA were significantly up-regulated. | Protection against ROS associated toxicity | [62] |
| Beetroot juices | Betanin | Activation and translocation of Nrf2 and significantly increased the expression of GSTP, GSTM, GSTT and NQO-1. | Cytoprotective, Anticarcinogenic Hepatoprotective | [69] |
| Blueberry juice | Chlorogenic acid, vanillic acid, syringic acid, trans-ferulic acid, = protocatechuic acid, p-coumaric acid | Activated Nrf2 and up-regulation of HO-1 and glutamate-cysteine ligase modifier subunit (GCLM) | Beneficial to endothelial cell activity and vascular function | [61] |
| Broccoli sprout crude juice | N/A | Protected against β-amyloid peptide –induced cytotoxicity and apoptosis; up-regulated the intracellular glutathione content and mRNA levels or activity of HO-1, thioredoxin, thioredoxin reductase (TrxR), and NAD(P)H:quinone (NQO-1) | Neuroprotective | [66] |
| Cabbage and sauerkraut juices | N/A | Sauerkraut juice significantly increased the activity of GST and NQO-1; translocation of Nrf2 and up-regulation of GST and NQO-1 by both juices. | Chemo-preventive | [68] |
| Fruit juice of Actinidia chinensis | N/A | Increased Keap1 and Nrf2 activity; up-regulation of SOD and GSH and down-regulation of ALT and AST diabetic patients | Antidiabetic Anti-inflammatory | [70] |
| Garlic juice | N/A | Reduces the ROS in presence of toxic heavy metal Cd; significantly induce the SOD and CAT activity; Nrf2 and NQO-1 expression was significantly increased; HO-1 expression not significant. | Prevents heavy metal (Cd) induced liver damages | [67] |
| Noni fruit juice | N/A | Aqueous and chloroform fractions protect cells from tert-Butyl hydroperoxide (TBHP)-induced cell damage; significantly decreases the TBHP cytotoxicity, ROS generation, mitochondrial membrane depolarization and apoptotic; nuclear accumulation of Nrf2 and HO-1, CAT, SOD-1. | Neuroprotective | [64] |
| 4-Methyl catechol, 4-Ethyl catechol, 4-Vinyl catechol, Scopoletin | Increased Nrf2 nuclear translocation, expression of HO-1, NQO-1 and glucose 6-phosphate dehydrogenase activity | Diabetic wound healing Cytotoxicity | [47,65] | |
| Orange juice | Lycopene, phytoene, all-E-β-carotene, and other carotenoids | Up-regulated the expression of GCS-1. GST-4, SOD-4, HSP-16.2 genes; significantly increased the ROS reductions, gene expression activation, oxidative stress resistance; induces GST-4::GFP expressions and increased SKN-1/Nrf2 transcription factor. | Neuroprotective and Suppress oxidative stress | [56] |
| Pomegranate juice | Anthocyanin and hydrolysable tannins | Decreased in SOD, GST, CAT and membrane-ATPases; significant increase in Nrf2 and NF-κB expression in nitrosodiethylamine (NDEA)-induced fibrotic rats. | Liver fibrosis Hepatoprotective | [58] |
| Pomegranate juice | Punicalagin | Increased Nrf2 translocation and up-regulation of HO-1; decreased the generation of RONS, NO; increased the production of SOD activity | Intestinal injuries | [59] |
N/A: Not available; Cd: Cadmium.
Table 3.
List of phytochemicals present in wine as Nrf2 activators.
| Source | Bioactive Natural Compound/s | Study Observations | Therapeutic Indications | Reference |
|---|---|---|---|---|
| Chinese Jing wine | Calycosin, ethyl ferulate and two cinnamic acid derivatives | Crude extract increased the Nrf2-ARE reporter activity by seven–eight fold; strongly activates the Nrf2-ARE reporter activity. | Cytotoxic | [97] |
| Makgeolli lees (ML) fermented raw rice wine | N/A | Decreased the production of intercellular ROS generated in 1,1′-dimethyl-4,4′-bipyridinium dichloride toxicity; up-regulation of expression and translocation of Nrf2 and ARE-GFP reporter activity; up-regulation the expression of NQO-1, HO-1, GPX, SOD, CAT, peroxiredoxin 3 and 4. | Suppress oxidative stress chemoprotective | [94] |
| Raffia Palm (Raphia hookeri) wine | N/A | Up-regulation of Nrf2 expression in diabetic brain cerebellar cortexes; significantly increased GSH, SOD and CAT activity. | Antidiabetic | [93] |
| Wine | Resveratrol | Significant decrease in hepatic lipid peroxidation, protein oxidation and expression of nitric oxide synthase and 3-nitrotyrosine. Dose of 100 or 300 mg/kg increased in Nrf2 mRNA expression in diethyl nitrosamine induced liver tumorigenesis | Chemo-preventive | [81] |
| Enhanced expression of Nrf2 and redox effector factor-1 (Ref-1) enhanced by resveratol; enhanced stem cell survival, proliferation and regeneration by expression of EGFP. | Improved cardiac function and regeneration | [82] | ||
| Dose of 20 mM protects against oxygen-glucose deprivation/reoxy-generation; activation of Nrf2 via upregulation of NQO-1 and HO-1; increase in SOD and GSH activity | Neuroprotective | [80] | ||
| Activation of Nrf2 via up-regulation of NQO-1, HO-1, GST-P1, GSH | Protect brain against cerebral ischemia, suppress oxidative and inflammatory stress | [77,78,79] | ||
| Parthenocissin A, quadrangularin A, pallidol (resveratrol dimer) | Selective singlet oxygen scavengers; deactivate hydroxyl or superoxide anions; pallidol activates ARE-dependent firefly luciferase reporter gene at 30 μM dose | Selective ROS scavengers | [83] | |
| ε-Viniferin (dehydro-dimer of resveratrol) | Inhibited platelet-derived growth factor-induced cell proliferation, migration, and ROS production, inducing nitric oxide (NO) generation; activation of Nrf2 and up-regulation of HO-1 | Prevent atherosclerosis and anti-proliferative | [84] | |
| Silibinin | Dose of 100 mg/kg induces the up-regulation of estrogen receptor-α (ERα), Nrf2 and HO-1 in pancreatic β-cells; increase insulin biosynthesis and secretion and decrease in ROS. | Antidiabetic | [91] | |
| Piceatannol | Dose of 10 μM activates the expression of HO-1 via Nrf2 activation; protected neuronal cells against glutamate-induced cell death by inducing HO-1 | Neuroprotective | [92] | |
| Catechin, epicatechin | Activation of Nrf2 and up-regulation of HO-1, GSH, NADPH, GST, NQO-1. | Cytoprotective, protect brain cells against cerebral ischemia. | [85,86,87] | |
| Quercetin | Activation of Nrf2 and translocation to the nucleus in mice model for traumatic brain injury; prevention of changes in cell morphology, apoptosis, activation of Nrf2 and increased expression of HO-1 and SOD in galactose induce mice neurotoxicity | Protection from traumatic brain injury, Neuroprotective | [83,88] | |
| Procyanidin B2 | Nuclear translocation of Nrf2 and up-regulation of expression of the glutathione S-transferase P1 (GSTP1), extracellular signal-regulated protein kinases (ERKs) and the p38 mitogen-activated protein kinase (MAPK) | Prevention of oxidative stress related intestinal injury and gut pathologies. | [89] | |
| Quercetin, epicatechin, morin, rutin, luteolin, naringenin, naringin, hesperidin | Quercetin is the strongest antioxidant and induce the expression of Metallothioneins 1 and 2 (MT-1/2); induces the transcriptional MT via the activation of JNK, p38 and PI3K/Akt signaling pathways; increased nuclear translocation of Nrf2 and DNA-binding activity | Hepatoprotective | [90] | |
| Yellow rice wine | Polyphenols | Dose of 30 mg/kg of polyphenols improved doxorubicin (DOX)-induced cardiac dysfunctions; prevent mitochondria-mediated cardiac apoptosis; down-regulation of transforming growth factor beta 1 pathway by promoting Nrf2 translocation. | Cardioprotective | [96] |
Table 4.
List of phytochemicals present in coffee and cocoa as Nrf2 activators.
| Sources | Bioactive Natural Compound/s | Study Observations | Therapeutic Indications | Reference |
|---|---|---|---|---|
| Cocoa: Cocoa Cocoa/tea | Catechin | Nuclear translocation of Nrf2, expression of Nrf2 target genes: nuclear respiratory factor 1 (Nrf1), GA binding protein transcription factor alpha subunit (GABPA), essential genes for increasing mitochondrial function, improve cellular redox state, mitochondrial respiration, stimulates glucose secretion and β-cell function | Antidiabetic | [116] |
| Epicatechin | Decreased ROS production; activated nuclear Nrf2 without up-regulation of GPx, SOD1, HO-1 and GR activity, promotes cell protection and survival; did not alter HO-1 expression, inhibition of activator protein 1 (AP-1) | Antidiabetic, prevention of oxidative stress, cerebro-protection (brain neuro protection), ischemic heart diseases, ischemic brain diseases (stroke) | [117,118,119] | |
| Procyanidin B2 | Nuclear translocation of Nrf2 and up-regulation of expression of the glutathione S-transferase P1 (GSTP1), extracellular signal-regulated protein kinases (ERKs) and the p38 mitogen-activated protein kinase (MAPK) activity. | Prevention of oxidative stress related intestinal injury and gut pathologies | [89] | |
| Coffee: Coffee/tea Coffee (raw and roasted) Coffee/regular coffee/decaffeinated coffee Coffee | Chlorogenic acid | Activation of Nrf2 and up-regulation of HO-1, NQO-1, and GCLC. Reduces liver abnormalities, and activation of NLR3 inflammasomes; activation of Nrf2 and up-regulation of HO-1, NQO-1, glutathione, thioredoxin reductase 1 (TrxR), and thioredoxin 1. | Hepatoprotection, Neuroprotective | [102,103] |
| Pyrocatechol | Activation of Nrf2, inhibition of NF-κB, up-regulation of HO-1 and NQO-1 genes | Anti-inflammatory | [120] | |
| 5-O-Caffeoylquinic acid N-methylpyridinium | Increased Nrf2 translocation and up-regulation of NQO-1 and glutathione-S-transferase-α1 (GSTA1) and increased GST activity | Chemo-preventive | [112,121] | |
| Cafesterol | Decreased production of ROS, translocation of Nrf2 and up-regulation of HO-1, inhibition of redox signaling and cell proliferation; Nrf2 activation, gsrD1-GFP reporter activation | Cardioprotective Cancer therapy, Anti-angiogenic Parkinson’s, Neuroprotection | [105,109,110] | |
| 3-CQA (caffeoylquinic acid), 4-CQA, 5-CQA, 3,4/3,5/4,5-diCQA | diCQA show greater free radical scavenging activity and Nrf2 activation; up-regulation of genes related to Nrf2 expression: GSH, HO-1, NQO-1 | Prevention of oxidative stress | [104,122] | |
| Cafesterol Kahweol | Up-regulation of NQO-1, GSTA1, UDP-glucuronosyl transferase 1A6 (UGT1A6) and the glutamate cysteine ligase catalytic (GCLC) activity and Nrf2 nuclear translocation; induction of enzymes: glutathione S-transferases, glucuronosyl S-transferases, increased expression of gamma-glutamyl cysteine synthetase and HO-1 | Chemo-preventive, Anti-carcinogenic, Bone diseases | [107,108,111] | |
| Kahweol | Reduced production of ROS, suppress the effects of H2O2 on oxidative phosphorylation, activation of caspase-3 protein, up-regulation of HO-1 and Nrf2 | Prevention of oxidative stress, Neuroprotection | [123,124] | |
| Caffeic acid | Induce HO-1 activity and Nrf2, modulate Keap1/Nrf2 interaction via increasing p62 expression. Up-regulation of Nrf2/electrophile responsive element (EpRE) | Antiviral, Anti-inflammatory, Antidiabetic | [113,114] | |
| Caffeine | Decreased intercellular ROS, catalase activity, up-regulation of nuclear Nrf2, | Prevention of oxidative stress | [115] | |
| Scopoletin | Activation of Nrf2, catalyzing methylglyoxal to lactic acid, inhibits glycation end products | Antidiabetic | [125] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Qader, M.; Xu, J.; Yang, Y.; Liu, Y.; Cao, S. Natural Nrf2 Activators from Juices, Wines, Coffee, and Cocoa. Beverages 2020, 6, 68. https://0-doi-org.brum.beds.ac.uk/10.3390/beverages6040068
AMA Style
Qader M, Xu J, Yang Y, Liu Y, Cao S. Natural Nrf2 Activators from Juices, Wines, Coffee, and Cocoa. Beverages. 2020; 6(4):68. https://0-doi-org.brum.beds.ac.uk/10.3390/beverages6040068
Chicago/Turabian StyleQader, Mallique, Jian Xu, Yuejun Yang, Yuancai Liu, and Shugeng Cao. 2020. "Natural Nrf2 Activators from Juices, Wines, Coffee, and Cocoa" Beverages 6, no. 4: 68. https://0-doi-org.brum.beds.ac.uk/10.3390/beverages6040068
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.