
Effects of Transportation Stress on Complete Blood Count, Blood Chemistry, and Cytokine Gene Expression in Heifers

 , , ,
, , ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Water Analysis and Quality
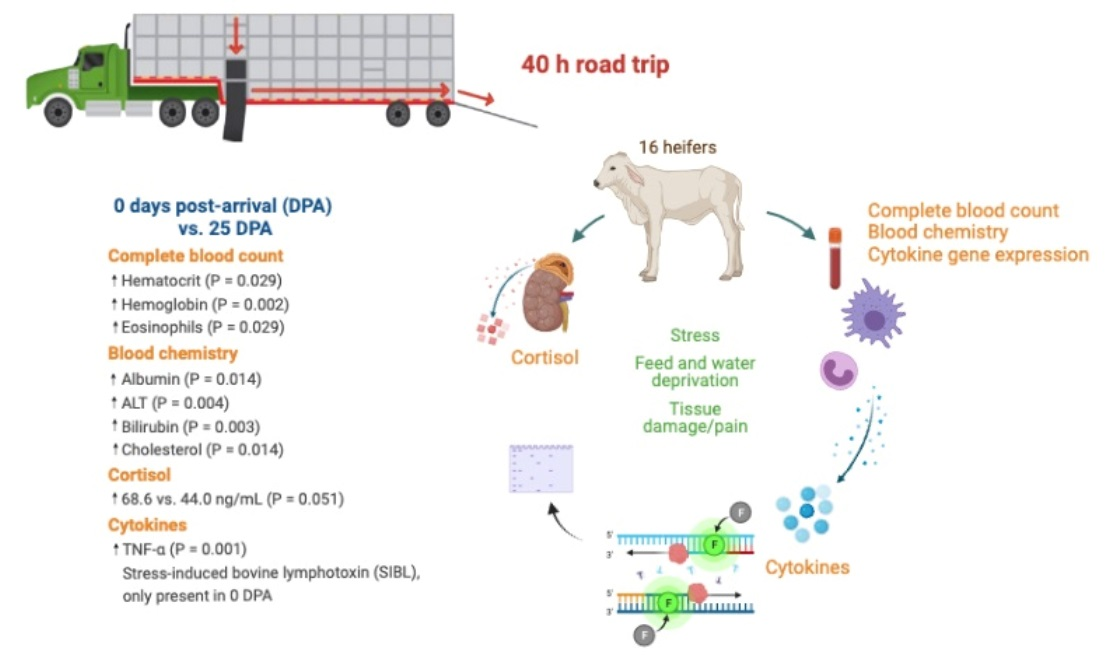
2.2. Heifers, Transportation, and Reception Diet
2.3. Sampling Procedures
2.4. Complete Blood Count, Blood Chemistry, and Cortisol Assays
2.5. RNA Extraction and Reverse Transcription—Polymerase Chain Reaction (RT-PCR)
2.6. Sequencing Non-Specific Bands
2.7. Statistical Analysis
3. Results
3.1. Complete Blood Count
3.2. Blood Chemistry
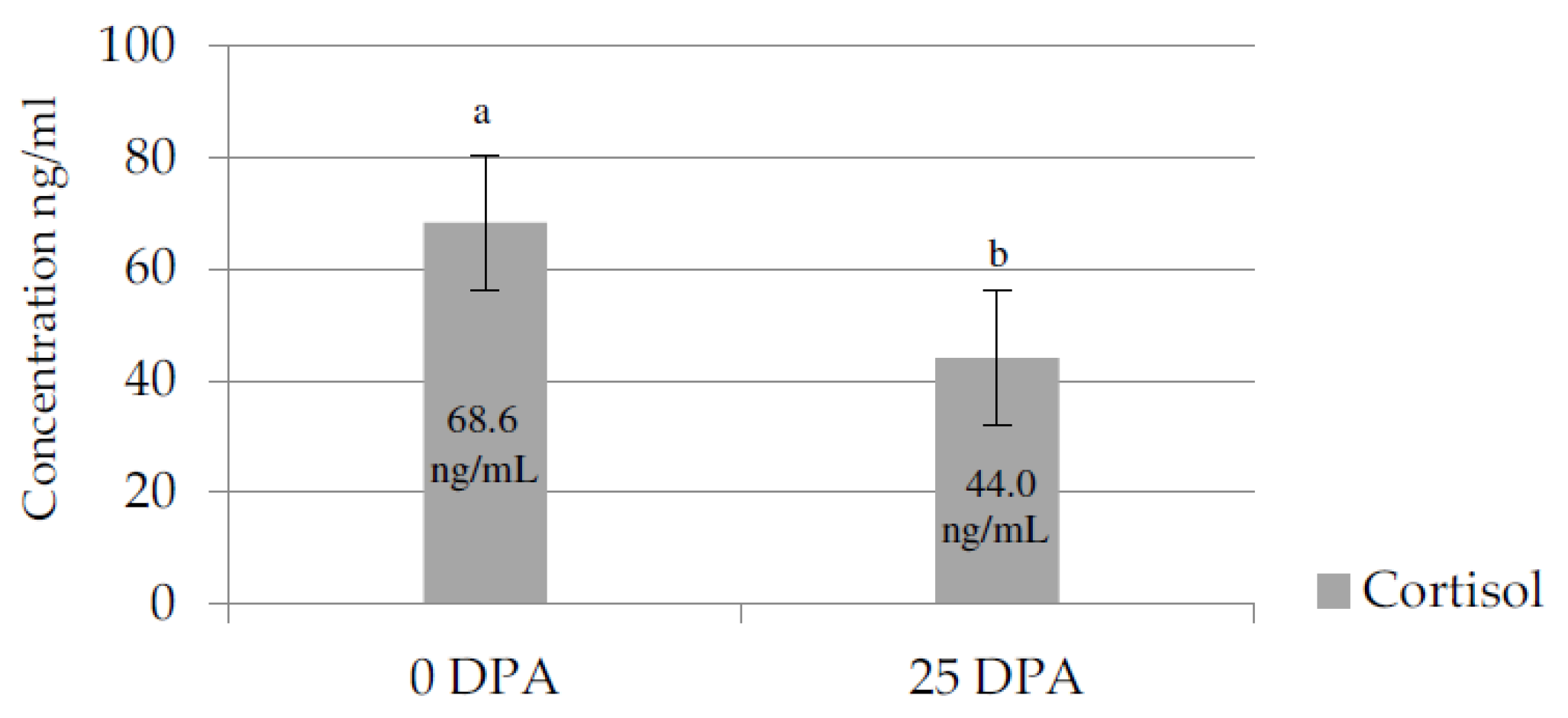
3.3. Cortisol
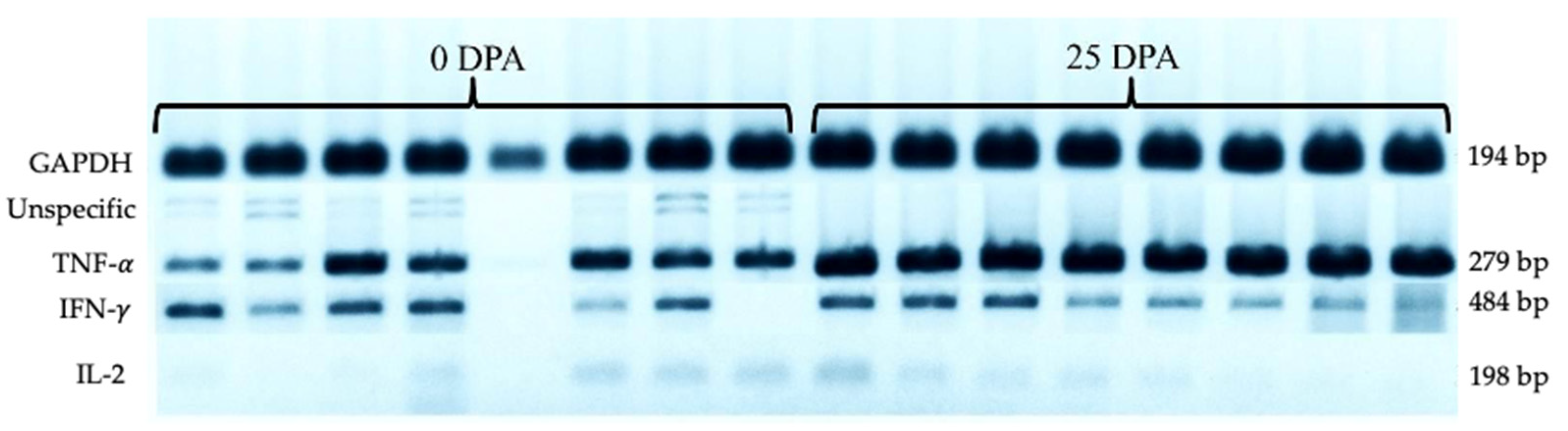
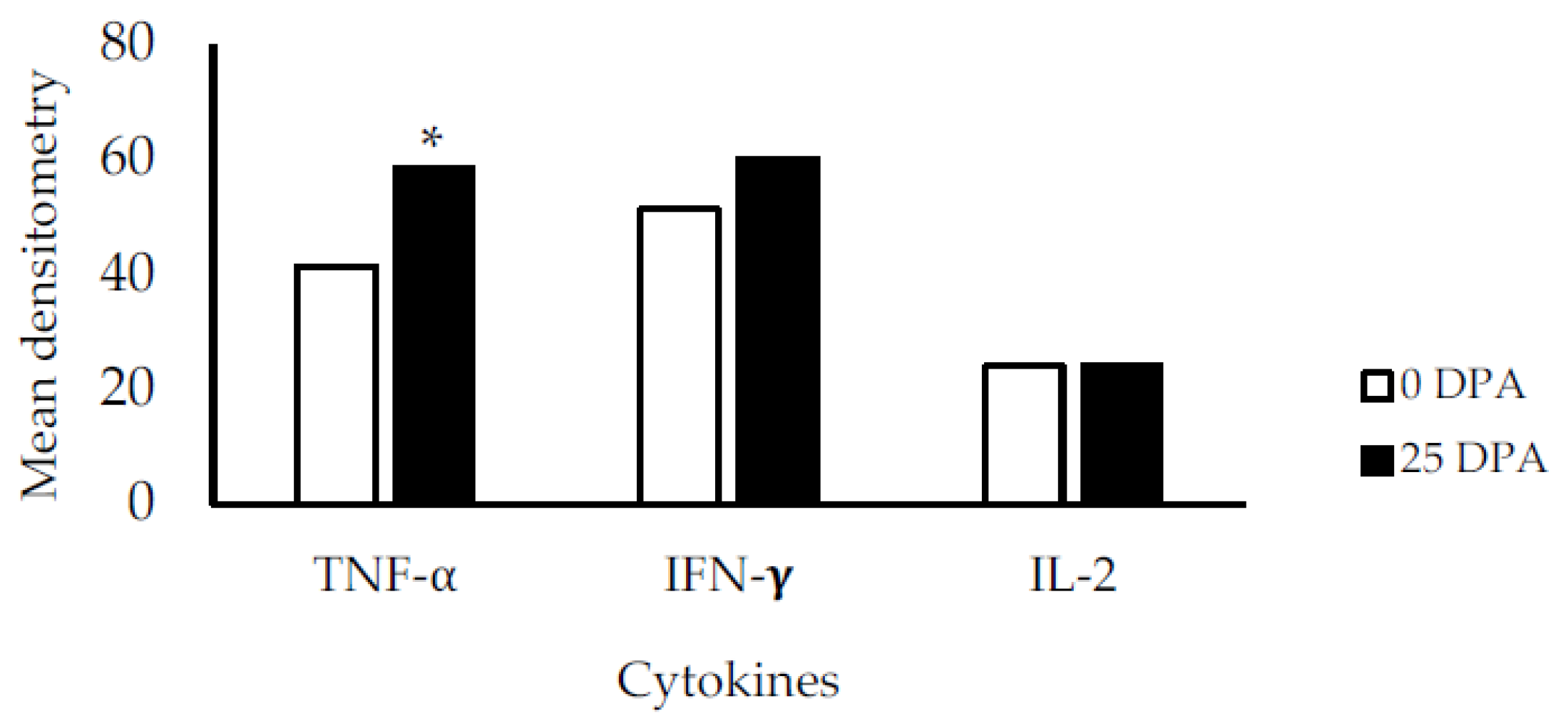
3.4. Cytokine Gene Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schwartzkopf, K.S.; Faucitano, L.; Dadgar, P.; Shand, P.; González, L.A.; Crowe, T.G. Road transport of cattle, swine and poultry in North America and its impact on animal welfare, carcass, and meat quality: A review. Meat Sci. 2012, 92, 227–243. [Google Scholar] [CrossRef]
- Appleby, M.C.; Cussen, V.; Garces, L. Long Distance Transport and Welfare of Farm Animals, 1st ed.; CABI: Cambridge, MA, USA, 2008; pp. 157–181. [Google Scholar]
- Cernicchiaro, N.; White, B.J.; Renter, D.J.; Babcock, A.H.; Kelly, L.; Slattery, R. Associations between the distance traveled from sale barns to commercial feedlots in the United States and overall performance, risk of respiratory disease, and cumulative mortality in feeder cattle during 1997–2009. J. Anim. Sci. 2012, 90, 1929–1939. [Google Scholar] [CrossRef]
- Buckham, S.; Xiao, L.; Tempelman, R.J.; Burton, J.L.; Earley, B.; Crowe, M.A. Transportation stress alters physiological circulating parameters that may be effective biomarkers of stress. J. Anim. Sci. 2008, 86, 1325–1334. [Google Scholar] [CrossRef]
- Swanson, J.C.; Morrow, J. Cattle transport: Historical research and future perspectives. J. Anim. Sci. 2001, 79, E102–E109. [Google Scholar] [CrossRef] [Green Version]
- Keeling, L.J. Healthy and happy: Animal welfare as an integral part of sustainable agriculture. Ambio. 2005, 34, 316–319. [Google Scholar] [CrossRef]
- Hughes, H.D.; Carroll, J.A.; Burdick, N.C.; Richeson, J.T. Natural variations in the stress and acute phase responses of cattle. Innate Immun. 2014, 20, 888–896. [Google Scholar] [CrossRef] [Green Version]
- Carroll, J.A.; Forsberg, N.E. Influence of stress and nutrition on cattle immunity. Vet. Clin. N. Am. Food Anim. Pract. 2007, 23, 105–149. [Google Scholar] [CrossRef]
- Hartung, J.; Nowak, B.; Springorum, A.C. Animal Welfare and Meat Quality. Improving the Sensory and Nutritional Quality of fresh Meat, 1st ed.; Woodhead Publishing: Sawston, UK, 2009; pp. 628–646. [Google Scholar]
- Kahn, C.M.; Line, S. The Merck Veterinary Manual, 10th ed.; Merck & Co, Inc.: Whitehouse Station, NJ, USA, 2010. [Google Scholar]
- Official Mexican Norm (NOM-051-ZOO-1995). Humane Treatment in the Mobilization of Animals; ISO: Geneva, Switzerland, 1998; pp. 1–20. [Google Scholar]
- Zaros, L.G.; Bricarello, P.A.; Amarante, A.F.T.; Coutinho, L.L. Quantification of bovine cytokine gene expression using real-time RT-PCR methodology. Genet. Mol. Biol. 2007, 30, 575–579. [Google Scholar] [CrossRef]
- Romero, M.H.; Uribe-Velásquez, L.F.; Sánchez, J.A. Biomarcadores de estrés como indicadores de bienestar animal en ganado de carne. Biosalud 2011, 10, 71–87. [Google Scholar]
- Caulfield, M.P.; Cambridge, H.; Foster, S.F.; McGreevy, P.D. Heat stress: A major contributor to poor animal welfare associated with long-haul live export voyages. Vet. J. 2014, 199, 223–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parker, A.J.; Hamlin, G.P.; Coleman, C.J.; Fitzpatrick, L.A. Excess cortisol interferes with a principal mechanism of resistance to dehydration in Bos indicus steers. J. Anim. Sci. 2004, 82, 1037–1045. [Google Scholar] [CrossRef]
- Cole, N.A.; Hutcheson, D.P. Influence of prefast feed intake on recovery from feed and water deprivation by beef steers. J. Anim. Sci. 1985, 60, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishizaki, H.; Kariya, Y. Road transportation stress promptly increases bovine peripheral blood absolute NK cell counts and cortisol levels. J. Vet. Med. Sci. 2010, 72, 747–753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez, O.; Marco, I.; Montane, J.; Lavin, S. Transport stress in southern chamois (Rupicapra pyrenaica) and its modulation by acepromazine. Vet. J. 2006, 720, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Marcato, F.; Van Den Brand, H.; Jansen, C.; Rutten, V.; Kemp, B.; Engel, B.; Wolthuis, M.; Van Reenen, K. Effects of pre-transport diet, transport duration and transport condition on immune cell subsets, haptoglobin, cortisol and bilirubin in young veal calves. PLoS ONE 2021, 16, e0246959. [Google Scholar] [CrossRef] [PubMed]
- Ishiwata, T.; Uetake, K.; Eguchi, Y.; Tanaka, T. Steer stress levels during long distance transport throughout the year in Japan. Anim. Sci. J. 2008, 79, 510–517. [Google Scholar] [CrossRef]
- Grandin, T. Assessment of stress during handling and transport. J. Anim. Sci. 1997, 75, 249–257. [Google Scholar] [CrossRef] [Green Version]
- Hopster, H.; Van der Werf, J.T.; Erkens, J.H.; Blokhuis, H.J. Effects of repeated jugular puncture on plasma cortisol concentrations in loose-housed dairy cows. J. Anim. Sci. 1999, 77, 708–714. [Google Scholar] [CrossRef]
- Dunn, C.S. Stress reactions of cattle undergoing ritual slaughter using two methods of restraint. Vet. Rec. 1990, 126, 522–525. [Google Scholar]
- Kushibiki, S. Tumor necrosis factor-α-induced inflammatory responses in cattle. Anim. Sci. J. 2011, 82, 504–511. [Google Scholar] [CrossRef]
- Cludts, I.; Cleuter, Y.; Kettmann, R.; Burny, A.; Droogmans, L. Cloning and characterization of the tandemly arranged bovine lymphotoxin and tumour necrosis factor-alpha genes. Cytokine 1993, 5, 336–341. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | Content (g/kg) |
|---|---|
| Buffer grass hay | 250 |
| Steam-flaked sorghum | 440 |
| Distillers’ dried grain | 100 |
| Soybean meal | 84.5 |
| Whole cottonseed | 50 |
| Molasses | 60 |
| Mineral–vitamin premix 1 | 2.5 |
| Calcium carbonate | 8 |
| Urea | 5 |
| Gene | Sequence (5′ → 3′) | Amplicon (bp) 1 | Annealing (°C) | Melting (°C) | Extension (°C) |
|---|---|---|---|---|---|
| TNF-α | F-TAACAAGCCGGTAGCCCACG | 279 | 57 | 88.5 | 72 |
| R-TCTTGATGGCAGACAGGATG | |||||
| IFN-γ | F-GCCAAATTGTCTCCTTCTACTTC | 484 | 61 | 85 | 72 |
| R-GGGTCAAGTGAAATAGTCACAGG | |||||
| IL-2 | F-TCCAAGCAAAAACCTGAACC | 198 | 57 | 82.5 | 72 |
| GADPH | R-CAGCGTTTACTGTTGCATCATCF-GGCGTGAACCACGAGAAGTATAA | 194 | 59 | 86 | 72 |
| R-CCCTCCACGATGCCAAAGT |
| Analysis | Reference Values a | Days after Arrival | SE | p | |
|---|---|---|---|---|---|
| 0 | 25 | ||||
| Hematocrit (%) | 24–46 | 48.4 | 40.6 | 2.14 | 0.029 |
| Hemoglobin (g/dL) | 8.0–14.0 | 15.7 | 13.0 | 0.48 | 0.002 |
| Neutrophils (%) b | 15–61 | 32.9 | 28.8 | 3.46 | 0.430 |
| Lymphocytes (%) | 26–68 | 54.0 | 61.0 | 3.91 | 0.227 |
| Monocytes (%) b | 0–12 | 7.6 | 8.5 | 1.41 | 0.750 |
| Eosinophils (%) b | 0–28 | 5.25 | 1.75 | 1.29 | 0.029 |
| N:L b | 0.5 | 0.69 | 0.49 | 0.129 | 0.372 |
| Platelets (K/µL) | 175–500 | 572 | 688 | 125.1 | 0.521 |
| Analysis | Reference Values a | Days Post-Arrival | SE | p | |
|---|---|---|---|---|---|
| 0 | 25 | ||||
| Glucose (mg/dL) | 40–100 | 68.4 | 80.0 | 5.79 | 0.177 |
| BUN (mg/dL) | 10–25 | 12.0 | 9.25 | 1.059 | 0.088 |
| Creatinine (mg/dL) | 0.5–2.2 | 0.95 | 0.94 | 0.052 | 0.867 |
| Phosphorus (mg/dL) | 5.6–8.0 | 7.93 | 9.23 | 0.516 | 0.098 |
| Calcium (mg/dL) | 8.0–11.4 | 11.0 | 10.8 | 0.285 | 0.674 |
| Total protein (g/dL) | 6.7–7.5 | 7.29 | 7.26 | 0.207 | 0.934 |
| Albumin (g/dL) | 2.5–3.8 | 2.91 | 2.56 | 0.088 | 0.014 |
| Globulin (g/dL) | 3.0–3.5 | 4.63 | 4.71 | 0.108 | 0.579 |
| ALT (U/L) | 6.9–35 | 74.4 | 40.1 | 6.96 | 0.004 |
| ALP (U/L) b | 18–153 | 153 | 136 | 26.15 | 0.958 |
| Bilirubin (mg/dL) b | 0.0–1.6 | 1.1 | 0.49 | 0.104 | 0.001 |
| Cholesterol (mg/dL) | 62–193 | 100.3 | 66.6 | 8.46 | 0.014 |
| Amylase (U/L) b | 41–98 | 56.8 | 12.5 | 8.41 | 0.139 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Avila-Jaime, B.; Ramos-Zayas, Y.; Franco-Molina, M.A.; Alvarado-Avila, R.; Zamora-Avila, D.E.; Fimbres-Durazo, H.; Zárate-Ramos, J.J.; Kawas, J.R. Effects of Transportation Stress on Complete Blood Count, Blood Chemistry, and Cytokine Gene Expression in Heifers. Vet. Sci. 2021, 8, 231. https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci8100231
Avila-Jaime B, Ramos-Zayas Y, Franco-Molina MA, Alvarado-Avila R, Zamora-Avila DE, Fimbres-Durazo H, Zárate-Ramos JJ, Kawas JR. Effects of Transportation Stress on Complete Blood Count, Blood Chemistry, and Cytokine Gene Expression in Heifers. Veterinary Sciences. 2021; 8(10):231. https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci8100231
Chicago/Turabian StyleAvila-Jaime, Benito, Yareellys Ramos-Zayas, Moisés A. Franco-Molina, René Alvarado-Avila, Diana E. Zamora-Avila, Héctor Fimbres-Durazo, Juan J. Zárate-Ramos, and Jorge R. Kawas. 2021. "Effects of Transportation Stress on Complete Blood Count, Blood Chemistry, and Cytokine Gene Expression in Heifers" Veterinary Sciences 8, no. 10: 231. https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci8100231