Factor Influences for Diagnosis and Vaccination of Avian Infectious Bronchitis Virus (Gammacoronavirus) in Chickens

 ,
, 
Abstract
:1. Introduction
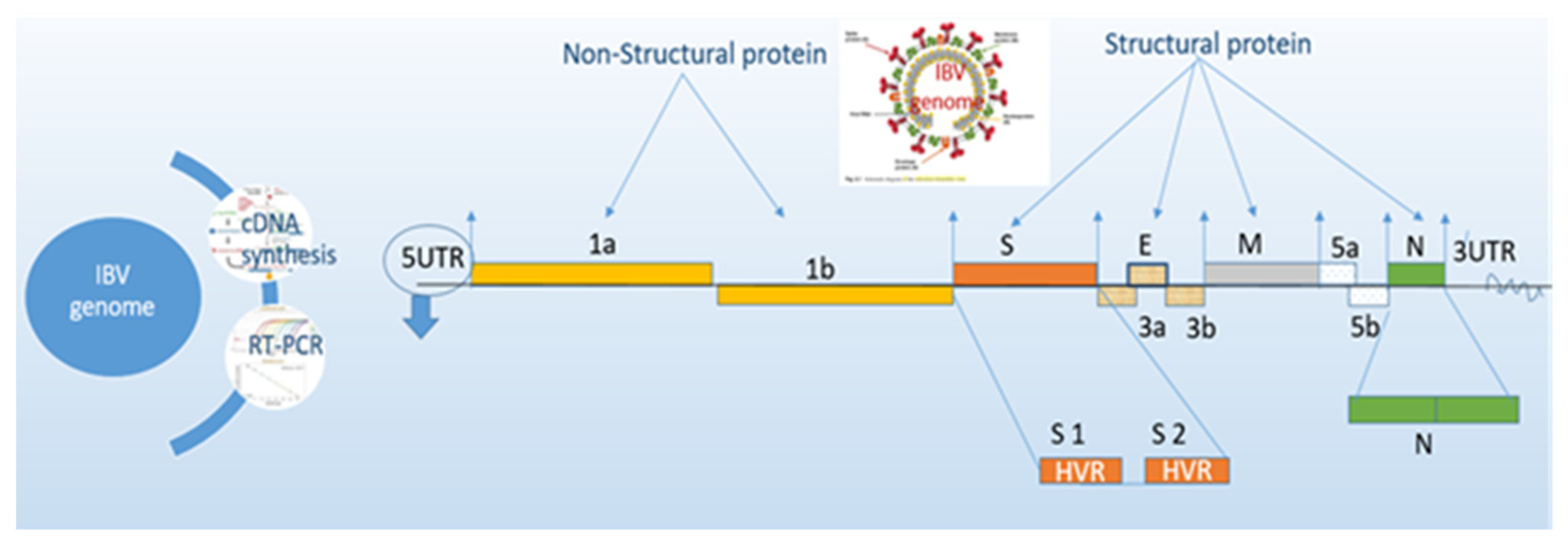
2. Factors Influencing IBV Diagnosis
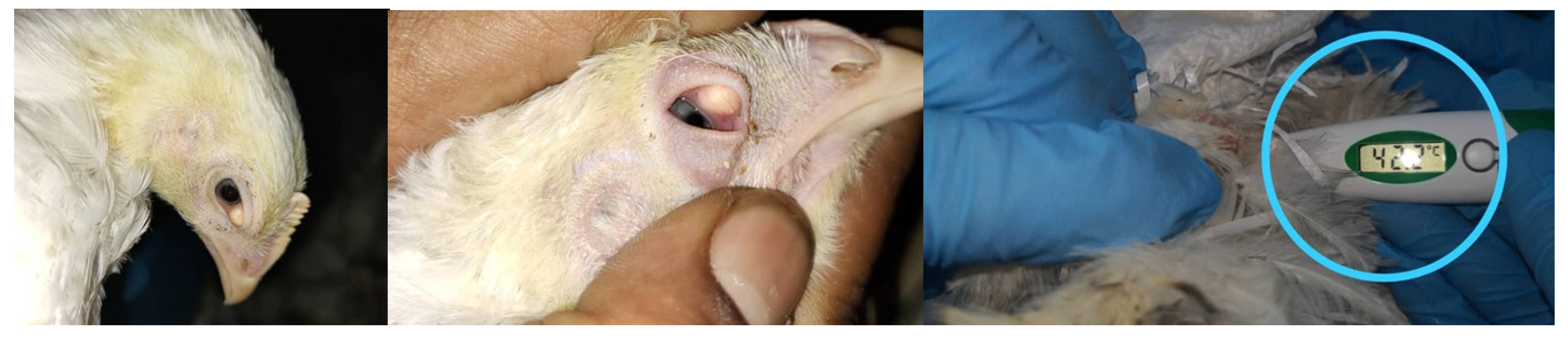
2.1. Clinical Signs
2.2. Vaccination
2.3. Assay Selection
2.4. Serology
2.5. Organ Selection and Sample Quality
2.6. Immunization and Challenge during Infection
2.7. Genetics and Immunosuppression
3. Vaccination Factor Influence
3.1. Stress and Vaccination
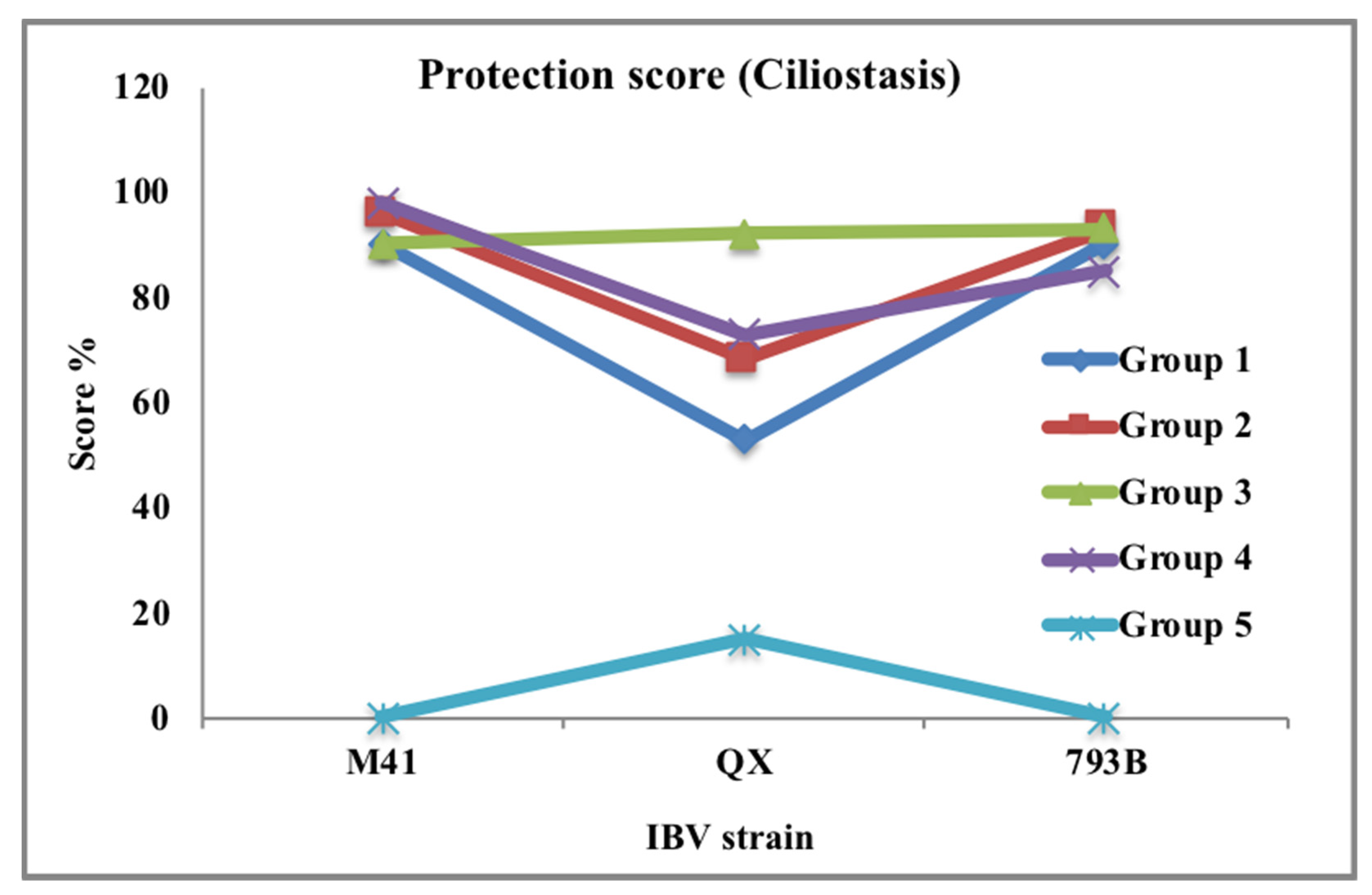
3.2. Methods for Protection Studies
3.3. Vaccine and Vaccination Program
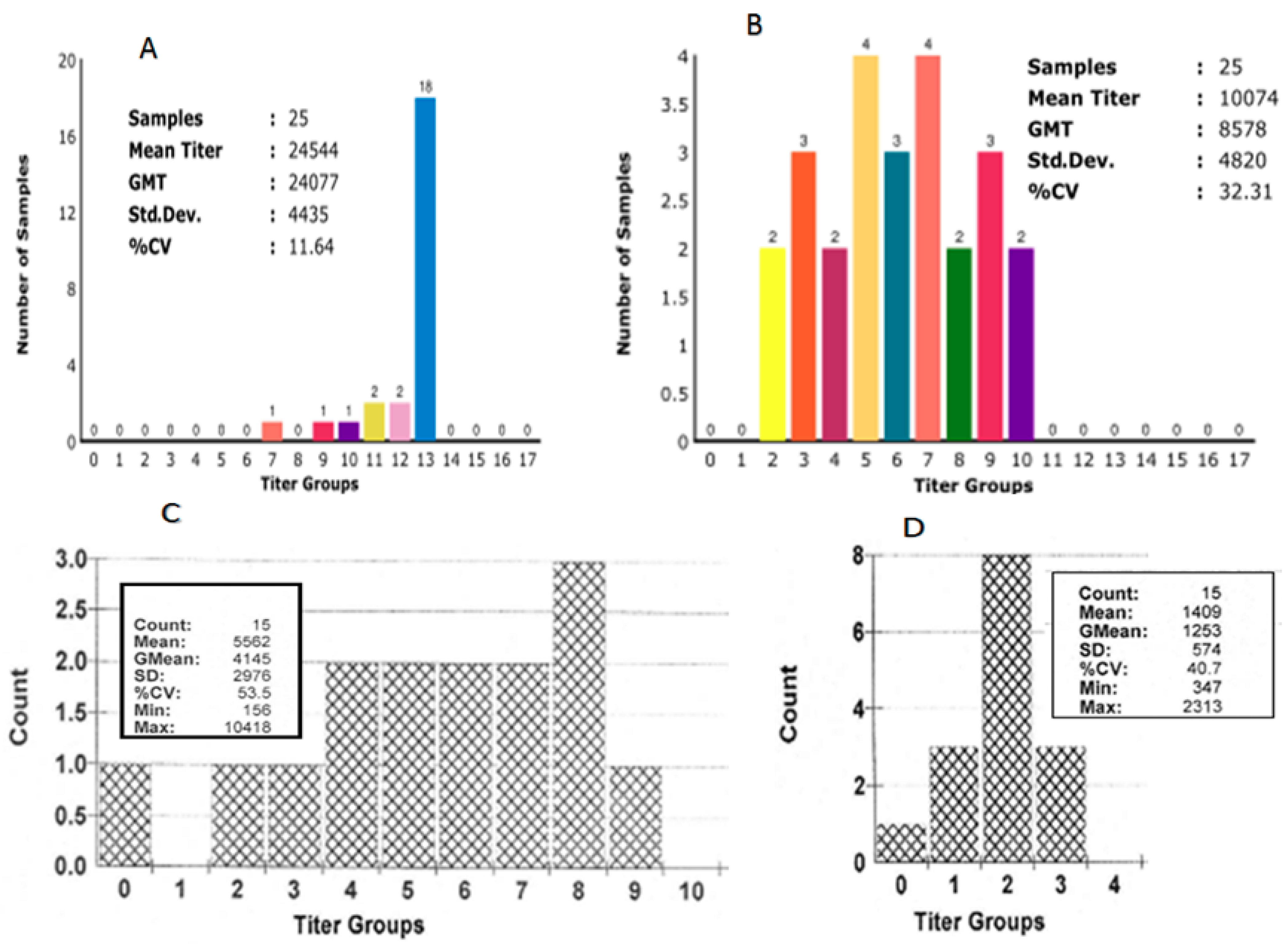
3.4. Group Size and Sample Number
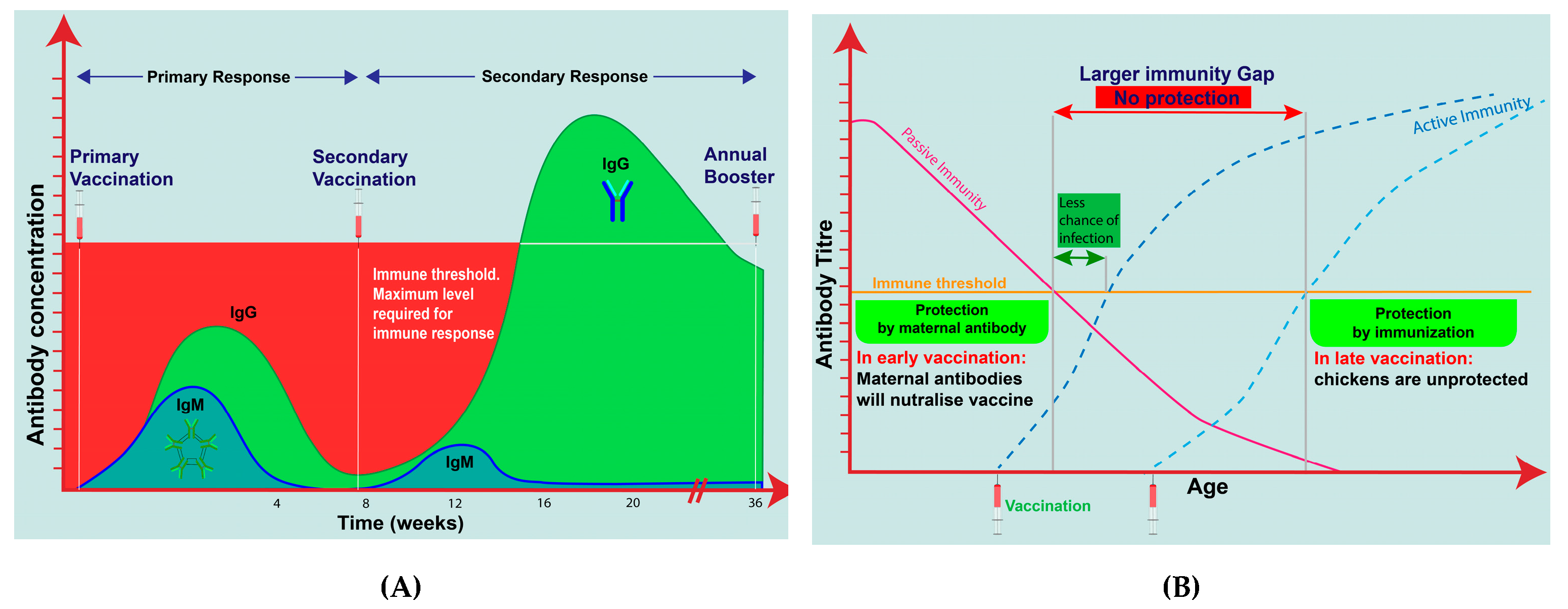
3.5. Immunity at Time of Vaccination/Infection
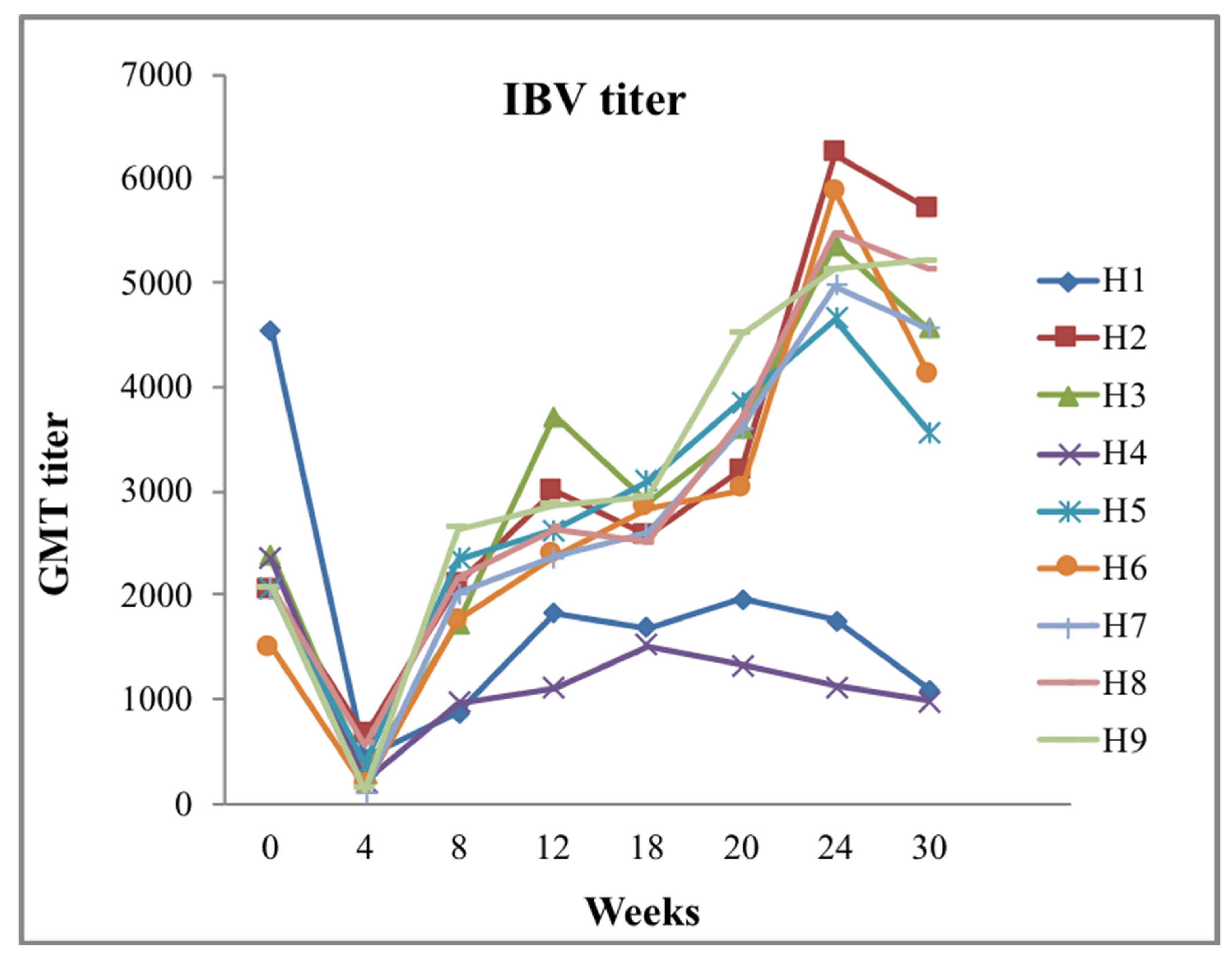
3.6. Age and Maternally Derived Antibodies
3.7. Genetics
3.8. Vaccine Dosage
3.9. Vaccination Schedule
3.10. Cross-Reactions between Serotypes
3.11. Vaccine Handling and Storage
3.12. Postvaccine Challenge and Reaction
3.13. Management Factors
Ventilation Quality
3.14. Possible Immunosuppression
3.15. Routes of IBV Vaccination
3.15.1. Spray Vaccination
3.15.2. Oculonasal
3.15.3. Drinking water
3.15.4. Intramuscular Injection
3.15.5. Gel Vaccination
3.16. Vaccine Index (VI)
4. Differential Diagnosis of Infectious Bronchitis Virus
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Papineau, A.; Berhane, Y.; Wylie, T.N.; Wylie, K.M.; Sharpe, S.; Lung, O. Genome organization of Canada Goose Coronavirus, a novel species identified in a mass die-off of Canada Geese. Sci. Rep. 2019, 9, e5954. [Google Scholar] [CrossRef] [PubMed]
- Legnardi, M.; Tucciarone, C.M.; Franzo, G.; Cecchinato, M. Infectious Bronchitis Virus Evolution, Diagnosis and Control. Vet. Sci. 2020, 7, 79. [Google Scholar] [CrossRef] [PubMed]
- Keep, S.; Oade, M.S.; Lidzbarski-Silvestre, F.; Bentley, K.; Stevenson-Leggett, P.; Freimanis, G.L.; Tennakoon, C.; Sanderson, N.; Hammond, J.A.; Jones, R.C.; et al. Multiple novel non-canonically transcribed sub-genomic mRNAs produced by avian coronavirus infectious bronchitis virus. J. Gen. Virol. 2020, 101, 1103–1118. [Google Scholar] [CrossRef]
- Decaro, N.; Lorusso, A. Novel human coronavirus (SARS-CoV-2): A lesson from animal coronaviruses. Vet. Microbiol. 2020, 104, 252–253. [Google Scholar] [CrossRef]
- Khan, S.; Ali, A.; Siddique, R.; Nabi, G. Novel coronavirus is putting the whole world on alert. J. Hosp. Infect. 2020, 104, 252–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mo, M.; Huang, B.; Wei, P.; Wei, T.; Chen, Q.; Wang, X.; Li, M.; Fan, W. Complete genome sequences of two Chinese virulent avian coronavirus infectious bronchitis virus variants. J. Virol. 2012, 86, 10903–10904. [Google Scholar] [CrossRef] [Green Version]
- Bande, F.; Arshad, S.S.; Omar, A.R.; Bejo, M.H.; Abubakar, M.S.; Abba, Y. Pathogenesis and diagnostic approaches of avian infectious bronchitis. Adv. Virol. 2016, 2016, 4621659. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Youn, H.N.; Kwon, J.S.; Lee, Y.J.; Kim, J.H.; Lee, J.B.; Park, S.Y.; Choi, S.; Song, C.S. Characterization of a novel live attenuated infectious bronchitis virus vaccine candidate derived from a Korean nephropathogenic strain. Vaccine 2010, 28, 2887–2894. [Google Scholar] [CrossRef]
- Valastro, V.; Holmes, E.C.; Britton, P.; Fusaro, A.; Jackwood, M.W.; Cattoli, G.; Monne, I. S1 gene-based phylogeny of infectious bronchitis virus: An attempt to harmonize virus classification. Infect. Genet. Evol. 2016, 39, 349–364. [Google Scholar] [CrossRef]
- De Wit, J.J.; Cook, J.K.A.; der Heijden, H.M. Infectious bronchitis virus variants: A review of the history, current situation and control measures. Avian Pathol. 2011, 40, 223–235. [Google Scholar] [CrossRef]
- Jackwood, M.W. Review of infectious bronchitis virus around the world. Avian Dis. 2012, 56, 634–641. [Google Scholar] [CrossRef] [Green Version]
- Cook, J.K.A.; Jackwood, M.; Jones, R.C. The long view: 40 years of infectious bronchitis research. Avian Pathol. 2012, 41, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Awad, F.; Chhabra, R.; Forrester, A.; Chantrey, J.; Baylis, M.; Lemiere, S.; Hussein, H.A.; Ganapathy, K. Experimental infection of IS/885/00-like infectious bronchitis virus in specific pathogen free and commercial broiler chicks. Res. Vet. Sci. 2016, 105, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Rohaim, M.A.; El Naggar, R.F.; Abdelsabour, M.A.; Mohamed, M.H.; El-Sabagh, I.M.; Munir, M. Evolutionary analysis of infectious bronchitis virus reveals marked genetic diversity and recombination events. Genes 2020, 11, 605. [Google Scholar] [CrossRef] [PubMed]
- Ennaji, Y.; Khataby, K.; Ennaji, M.M. Infectious Bronchitis Virus in Poultry: Molecular Epidemiology and Factors Leading to the Emergence and Reemergence of Novel Strains of Infectious Bronchitis Virus. Emerg. Reemerging Viral Pathog. 2020, 31–44. [Google Scholar]
- Cook, J.K.; Orbell, S.J.; Woods, M.A.; Huggins, M.B. Breadth of protection of the respiratory tract provided by different live-attenuated infectious bronchitis vaccines against challenge with infectious bronchitis viruses of heterologous serotypes. Avian Pathol. 1999, 28, 477–485. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Cheng, J.L.; Liu, X.Y.; Zhao, J.; Hu, Y.X.; Zhang, G.Z. Safety and efficacy of an attenuated Chinese QX-like infectious bronchitis virus strain as a candidate vaccine. Vet. Microbiol. 2015, 180, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Terregino, C.; Toffan, A.; Beato, M.S.; De Nardi, R.; Vascellari, M.; Meini, A.; Ortali, G.; Mancin, M.; Capua, I. Pathogenicity of a QX strain of infectious bronchitis virus in specific pathogen free and commercial broiler chickens, and evaluation of protection induced by a vaccination programme based on the Ma5 and 4/91 serotypes. Avian Pathol. 2008, 37, 487–493. [Google Scholar] [CrossRef]
- He, K.; Li, M.; Wei, P.; Mo, M.L.; Wei, T.C.; Li, K.R. Complete genome sequence of an infectious bronchitis virus chimera between co-circulating heterotypic strains. J. Virol. 2012, 86, 13887–13888. [Google Scholar] [CrossRef] [Green Version]
- Ignjatovic, J.; Gould, G.; Sapats, S. Isolation of a variant infectious bronchitis virus in Australia that further illustrates diversity among emerging strains. Arch. Virol. 2006, 151, 1567–1585. [Google Scholar] [CrossRef]
- Cavanagh, D. Coronavirus avian infectious bronchitis virus. Vet. Res. 2007, 38, 281–297. [Google Scholar] [CrossRef] [Green Version]
- McKinley, E.T.; Jackwood, M.W.; Hilt, D.A.; Kissinger, J.C.; Robertson, J.S.; Lemke, C.; Paterson, A.H. Attenuated live vaccine usage affects accurate measures of virus diversity and mutation rates in avian coronavirus infectious bronchitis virus. Virus Res. 2011, 158, 225–234. [Google Scholar] [CrossRef]
- Brun, A.; Albina, E.; Barret, T.; Chapman, D.A.; Czub, M.; Dixon, L.K.; Keil, G.M.; Klonjkowski, B.; Le Potier, M.F.; Libeau, G.; et al. Antigen delivery systems for veterinary vaccine development. Viral-Vector based delivery systems. Vaccine 2008, 26, 6508–6528. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.; Li, H.; Qian, J.; Yang, J.; Lu, Y.; Yi, Q. Optimizing dendritic cell vaccine for immunotherapy in multiple myeloma: Tumour lysates are more potent tumour antigens than idiotype protein to promote anti-tumour immunity. Clin. Exp. Immunol. 2012, 170, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Niewiesk, S. Maternal antibodies: Clinical significance, mechanism of interference with immune responses, and possible vaccination strategies. Front. Immunol. 2014, 5, 446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boelm, G.J. Influence of maternally derived antibodies on vaccination using a IBV H120 vaccine virus. J. Vet. Med. Res. 2018, 5, 1124. [Google Scholar]
- Bosha, J.A.; Nongo, N.N. Common breaches in poultry vaccine handling and administration in Makurdi metropolis: A recurrent phenomenon in the tropics. J. Vet. Med. Res. 2012, 9, 11–16. [Google Scholar]
- Ganguly, S.; Paul, I.; Mukhopadhayay, S.K. Different types of vaccines and vaccination—The most accepted trend to control and eradicate infections. Indian Pet. J. 2010, 5, 34–37. [Google Scholar]
- Abdullahi, U.S.; Adamu, S.B.; Ahmed, A.F. 2009 Investigations on some causes of poultry vaccination failures in Bauchi metropolis and environs. Niger. J. Exp. Appl Biol. 2009, 10, 47–50. [Google Scholar]
- De Herdt, P.; Ducatelle, A.R.; Uyttebroek, A.E.; Sneep, A.; Torbeyns, R. Infectious bronchitis serology in broilers and broiler breeders: Correlations between antibody titers and performance in vaccinated flocks. Avian Dis. 2001, 45, 612–619. [Google Scholar] [CrossRef] [PubMed]
- Van Leerdam, B.; Kuhne, P. Diagnosis of IBV Field Challenge—PoultryWorld. 2010. Available online: https://www.poultryworld.net/Home/General/2010/7/Diagnosis-of-IBV-field-challenge-WP007714W/ (accessed on 29 December 2020).
- Van Leerdam, B.; Kuhne, P. Understanding Elisa Results. 2017. Available online: https://www.biochek.com/wp-content/uploads/2017/08/IBV-Understanding-ELISA.pdf?x14571 (accessed on 30 December 2020).
- Messaï, C.R.; Salhi, O.; Khelef, D.; Lounas, A.; Mohamed-Cherif, A.; Kaidi, R.; Aït-Oudhia, K. Serological, clinical, and risk factors of the Newcastle disease on broilers flocks in Algeria. Vet World. 2019, 12, 938–944. [Google Scholar] [CrossRef]
- Cavanagha, D.; Gelb, J., Jr. Infectious Bronchitis. In Diseases of Poultry, 12th ed.; Saif, Y.M., Fadly, A.M., Glisson, J.R., McDougald, L.R., Nolan, L.K., Swayne, D.E., Eds.; Blackwell Publishing Professional: Ames, IA, USA, 2008; pp. 117–135. [Google Scholar]
- Subramanian, B.M.; Raj, G.D.; Kumanan, K.; Nachimuthu, K.; Nainar, A.M. Interaction between genomes of infectious bronchitis and Newcastle disease viruses studied by reverse transcription-polymerase chain reaction. Acta Virol. 2004, 48(2), 123–129. [Google Scholar] [PubMed]
- Capua, I.; Terregino, C.; Cattoli, G.; Mutinelli, F.; Rodriguez, J.F. Development of a DIVA-Differentiating infected from vaccinated animals strategy using a vaccine containing a heterologous neuraminidase for the control of avian influenza. Avian Pathol. 2003, 32, 47–55. [Google Scholar] [CrossRef]
- Goldenberg, D.; Lublin, A.; Rosenbluth, E.; Heller, E.D.; Pitcovski, J. Differentiating infected from vaccinated animals, and among virulent prototypes of reovirus. J. Virol Methods. 2011, 177, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Capua, I.; Cattoli, G.; Marangon, S. DIVA–a vaccination strategy enabling the detection of field exposure to avian influenza. Dev. Biol. 2004, 119, 229–233. [Google Scholar]
- Hamblin, C.; Kitching, R.P.; Donaldson, A.I.; Crowther, J.R.; Barnett, I.T.R. Enzyme-linked immunosorbent assay (ELISA) for the detection of antibodies against foot-and-mouth disease virus: III. Evaluation of antibodies after infection and vaccination. Epidemiol. Infect. 1987, 99, 733–744. [Google Scholar] [CrossRef] [Green Version]
- Clem, A.S. Fundamentals of vaccine immunology. J. Glob. Infect. Dis. 2011, 3, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Jordan, B. Vaccination against infectious bronchitis virus: A continuous challenge. Vet. Microbiol. 2017, 206, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishnan, S.; Kappala, D. Avian infectious bronchitis virus. In Recent Advances in Animal Virology; Springer: Singapore, 2019; pp. 301–319. [Google Scholar]
- Zavala, G. Fundamental Basics of Use and Interpretation of Serology. 2019. Available online: https://avicultura.info/en/fundamental-basics-of-the-use-and-interpretation-of-serology (accessed on 30 December 2020).
- Darbyshire, J.H.; Cook, J.K.; Peters, R.W. Growth comparisons of avian infectious bronchitis virus strains in organ cultures of chicken tissues. Arch. Virol. 1978, 56, 317–325. [Google Scholar] [CrossRef]
- Chen, H.T.; Zhang, J.; Ma, Y.P.; Ma, L.N.; Ding, Y.Z.; Liu, X.T.; Cai, X.P.; Ma, L.Q.; Zhang, Y.G.; Liu, Y.S. Reverse transcription loop-mediated isothermal amplification for the rapid detection of infectious bronchitis virus in infected chicken tissues. Mol. Cell Probes 2010, 24, 104–106. [Google Scholar] [CrossRef]
- Najimudeen, S.M.; Hassan, M.S.H.; Cork, S.C.; Abdul-Careem, M.F. Infectious Bronchitis Coronavirus Infection in Chickens: Multiple System Disease with Immune Suppression. Pathogens 2020, 9, 779. [Google Scholar] [CrossRef]
- Ren, M.; Han, Z.; Zhao, Y.; Sun, J.; Liu, S.; Ma, D. Multiple recombination events between field and vaccine strains resulted in the emergence of a novel infectious bronchitis virus with decreased pathogenicity and altered replication capacity. Poult Sci. 2020, 99, 1928–1938. [Google Scholar] [CrossRef]
- Villarreal, L.Y.B. Diagnosis of infectious bronchitis: An overview of concepts and tools. Rev. Bras. Cienc. Avic. 2010, 12, 111–114. [Google Scholar] [CrossRef] [Green Version]
- Santos, R.M.; Fernando, F.S.; Montassier, M.F.S.; Silva, K.R.; Lopes, P.D.; Pavani, C.; Borzi, M.M.; Okino, C.H.; Montassier, H.J. Memory immune responses and protection of chickens against a nephropathogenic infectious bronchitis virus strain by combining live heterologous and inactivated homologous vaccines. J. Vet. Med. Sci. 2019, 81, 612–619. [Google Scholar] [CrossRef] [PubMed]
- Jackwood, M.W.; de Wit, J.J. Infectious Bronchitis. In Diseases of Poultry, 3rd ed.; Swayne, D.E., Glisson, J.R., McDougald, L.R., Nolan, L.K., Suarez, D.L., Nair, V., Eds.; Blackwell Publishing Professional: Ames, IA, USA, 2013; pp. 117–135. [Google Scholar]
- Gharaibeh, S.; Mahmoud, K.; Al-Natour, M. Field evaluation of maternal antibody transfer to a group of pathogens in meat-type chickens. Poult Sci. 2008, 87, 1550–1555. [Google Scholar] [CrossRef] [PubMed]
- Coakley, C.M.; Staszewski, V.; Herborn, K.A.; Cunningham, E.J. Factors affecting the levels of protection transferred from mother to offspring following immune challenge. Front. Zool. 2014, 11(1), 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, K.; Cook, J.K.A.; Frazier, J.A.; Narita, M. Escherichia coli multiplication and lesions in the respiratory tract of chickens inoculated with infectious bronchitis virus and/or Escherichia coli. Avian Dis. 1992, 36, 881–890. [Google Scholar] [CrossRef]
- Hoerr, F.J. Clinical aspects of immunosuppression in poultry. Avian Dis. 2010, 54, 2–15. [Google Scholar] [CrossRef] [Green Version]
- Schat, K.A.; Skinner, M.A. Avian Immunosuppressive Diseases and Immunoevasion. Avian Immunol. 2014, 275–297. [Google Scholar] [CrossRef]
- Raj, G.D.; Jones, R.C. Immunopathogenesis of infection in SPF chicks and commercial broilers of a variant infectious bronchitis virus of economic importance. Avian Pathol. 1996, 25, 481–501. [Google Scholar] [CrossRef]
- Dantzer, R. Stress, stereotypies and welfare. Behav. Process. 1991, 25, 95–102. [Google Scholar] [CrossRef]
- Burns, V.E.; Carroll, D.; Drayson, M.; Whitham, M.; Ring, C. Life events, perceived stress and antibody response to influenza vaccination in young, healthy adults. J. Psychosom Res. 2003, 55, 569–572. [Google Scholar] [CrossRef]
- Takahashi, A.; Flanigan, M.E.; McEwen, B.S.; Russo, S.J. Aggression, Social Stress, and the Immune System in Humans and Animal Models. Front. Behav. Neurosci. 2018, 12, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hofmann, T.; Schmucker, S.S.; Bessei, W.; Grashorn, M.; Stefanski, V. Impact of Housing Environment on the Immune System in Chickens: A Review. Animals 2020, 10, 1138. [Google Scholar] [CrossRef]
- Kwok, A.H.Y.; Wang, Y.; Wang, C.Y.; Leung, F.C. Cloning of chicken glucocorticoid receptor (GR) and characterization of its expression in pituitary and extrapituitary tissues. Poult. Sci. 2007, 86, 423–430. [Google Scholar] [CrossRef]
- De Wit, J.J.; Cook, J.K. Factors influencing the outcome of infectious bronchitis vaccination and challenge experiments. Avian Pathol. 2014, 43, 485–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arvidson, Y.; Tannock, G.A.; Senthilselvan, A.; Zerbes, M. A model for determining immunogenic relationships between avian infectious bronchitis virus. Arch. Virol. 1990, 111, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Jackwood, M.W.; Jordan, B.J.; Roh, H.J.; Hilt, D.A.; Williams, S.M. Evaluating protection against infectious bronchitis virus by clinical signs, ciliostasis, challenge virus detection, and histopathology. Avian Dis. 2015, 59, 368–374. [Google Scholar] [CrossRef]
- Awad, F.; Hutton, S.; Forrester, A.; Baylis, M.; Ganapathy, K. Heterologous live infectious bronchitis virus vaccination in day-old commercial broiler chicks: Clinical signs, ciliary health, immune and protection against variant infectious bronchitis viruses. Avian Pathol. 2016, 45, 169–177. [Google Scholar] [CrossRef]
- Bande, F.; Arshad, S.S.; Bejo, M.H.; Moeini, H.; Omar, A.R. Progress and challenges toward the development of vaccines against avian infectious bronchitis. J. Immunol Res. 2015, 2015, 424860. [Google Scholar] [CrossRef] [Green Version]
- De Wit, J.J. Detection of infectious bronchitis virus. Avian Pathol. 2000, 29, 71–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karimi, V.; Mohammadi, P.; Ghalyanchilangeroudi, A.; Ghafouri, S.A.; Hashemzadeh, M.; Farahani, R.K.; Maghsouldoo, H.; Isakakroudi, N. Including 793/B type avian infectious bronchitis vaccine in 1-day-old chicken increased the protection against QX genotype. Trop. Anim. Health Prod. 2019, 51, 629–635. [Google Scholar] [CrossRef]
- De Wit, J.J.; Nieuwenhuisen-van Wilgen, J.; Hoogkamer, A.; van de Sande, H.; Zuidam, G.J.; Fabri, T.H. Induction of cystic oviducts and protection against early challenge with infectious bronchitis virus serotype D388 (genotype QX) by maternally derived antibodies and by early vaccination. Avian Pathol. 2011, 40, 463–471. [Google Scholar] [CrossRef] [Green Version]
- Awad, F.; Forrester, A.; Baylis, M.; Lemiere, S.; Ganapathy, K. Protection conferred by live infectious bronchitis vaccine viruses against variant Middle East IS/885/00-like and IS/1494/06-like isolates in commercial broiler chicks. Vet. Rec. Open 2015, 2, e000111. [Google Scholar] [CrossRef] [Green Version]
- Bru, T.; Vila, R.; Cabana, M.; Geerligs, H.J. Protection of chickens vaccinated with combinations of commercial live infectious bronchitis vaccines containing Massachusetts, Dutch and QX-like serotypes against challenge with virulent infectious bronchitis viruses 793B and IS/1494/06 Israel variant 2. Avian Pathol. 2017, 46, 52–58. [Google Scholar] [CrossRef] [Green Version]
- Vagnozzi, A.; García, M.; Riblet, S.M.; Zavala, G. Protection induced by infectious laryngotracheitis virus vaccines alone and combined with Newcastle disease virus and/or infectious bronchitis virus vaccines. Avian Dis. 2010, 54, 1210–1219. [Google Scholar] [CrossRef]
- De Wit, J.J. Underestimation of the difficulties of vaccination against viral respiratory diseases by mass application methods. In Proceedings of the XVIIIth International Congress of the World Veterinary Poultry Association, Nantes, France, 19–23 August 2013; pp. 63–67. [Google Scholar]
- Di Fabio, J.; Rossini, L.I.; Orbell, S.J.; Paul, G.; Huggins, M.B.; Malo, A.; Silva, B.G.; Cook, J.K.A. Characterization of infectious bronchitis viruses isolated from outbreaks of disease in commercial flocks in Brazil. Avian Dis. 2000, 44, 582–589. [Google Scholar] [CrossRef]
- De Wit, J.J.; Van Gerwe, T.W.J.M. Statistical approach of the required number of birds per group and number of TOCs per bird for ciliostasis testing in efficacy trials of IBV vaccines. In Proceedings of the IV International Symposium on Corona-And Pneumovirus Infections, Rauischholzhausen, Germany, 20–23 June 2004; pp. 213–219, VVB Laufersweiler Verlag. [Google Scholar]
- Benyeda, Z.; Mato, T.; Suveges, T.; Szabo, E.; Kardi, V.; Abonyi-Toth, Z.; Rusvai, M.; Palya, V. Comparison of the pathogenicity of QX-like, M41 and 793/B infectious bronchitis strains from different pathological conditions. Avian Pathol. 2009, 38, 449–456. [Google Scholar] [CrossRef] [Green Version]
- Geerligs, H.J.; Boelm, G.J.; Meinders, C.A.; Stuurman, B.G.; Symons, J.; Tarres-Call, J.; Bru, T.; Vila, R.; Mombarg, M.; Karaca, K.; et al. Efficacy and safety of an attenuated live QX-like infectious bronchitis virus strain as a vaccine for chickens. Avian Pathol. 2011, 40, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Greenwood, B. The contribution of vaccination to global health: Past, present and future. Phil. Trans. R. Soc. Lond. B. 2014, 369, 20130433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darbyshire, J.H.; Peters, R.W. Sequential development of humoral immunity and assessment of protection in chickens following vaccination and challenge with avian infectious bronchitis virus. Res Vet Sci. 1984, 37, 77–86. [Google Scholar] [CrossRef]
- Pensaert, M.; Lambrechts, C. Vaccination of chickens against a Belgian nephropathogenic strain of infectious bronchitis virus B1648 using attenuated homologous and heterologous strains. Avian Pathol. 1994, 23, 631–641. [Google Scholar] [CrossRef] [PubMed]
- De Wit, J.J.; Mekkes, D.R.; Kouwenhoven, B.; Verheijden, J.H. Sensitivity and specificity of serological tests for infectious bronchitis virus antibodies in broilers. Avian Pathol. 1997, 26, 105–118. [Google Scholar] [CrossRef] [PubMed]
- Chhabra, R.; Chantrey, J.; Ganapathy, K. Immune Responses to Virulent and Vaccine Strains of Infectious Bronchitis Viruses in Chickens. Viral Immunol. 2015, 28, 478–488. [Google Scholar] [CrossRef]
- Palgen, J.L.; Tchitchek, N.; Rodriguez-Pozo, A.; Jouhault, Q.; Abdelhouahab, H.; Dereuddre-Bosquet, N.; Contreras, V.; Martinon, F.; Cosma, A.; Lévy, Y.; et al. Innate and secondary humoral responses are improved by increasing the time between MVA vaccine immunizations. NPJ Vaccines 2020, 5, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Pulendran, B.; Ahmed, R. Immunological mechanisms of vaccination. Nat. Immunol. 2011, 12, 509–517. [Google Scholar] [CrossRef] [PubMed]
- Breytenbach, J.H. Guidelines for effective vaccination of broilers. Int. Poult. Prod. 1999, 13, 7–9. Available online: http://www.positiveaction.info/pdfs/articles/pp13.7p7.pdf (accessed on 30 December 2020).
- Bar-shira, E.; Sklan, D.; Friedman, A. Establishment of immune competence in the avian GALT during the immediate post-hatch period. Develop. Comp. Immunol. 2003, 27, 147–157. [Google Scholar] [CrossRef]
- Hamal, K.R.; Burgess, S.C.; Pevzner, I.Y.; Erf, G.F. Maternal antibody transfer from dams to their egg yolks, egg whites, and chicks in meat lines of chickens. Poult. Sci. 2006, 85, 1364–1372. [Google Scholar] [CrossRef]
- Kapczynski, D.R.; Hilt, D.A.; Shapiro, D.; Sellers, H.S.; Jackwood, M.W. Protection of chickens from infectious bronchitis by in ovo and intramuscular vaccination with a DNA vaccine expressing the S1 glycoprotein. Avian Dis. 2003, 47, 272–285. [Google Scholar] [CrossRef] [Green Version]
- Sahin, K.; Sahin, N.; Onderci, M.; Yaralioglu, S.; Kucuk, O. Protective role of supplemental vitamin E on lipid peroxidation, vitamins E, A, and some mineral concentrations of broilers reared under heat stress. Vet. Med. 2001, 46, 140–144. [Google Scholar] [CrossRef] [Green Version]
- Alizadeh, M.; Munyaka, P.; Yitbarek, A.; Echeverry, H.; Rodriguez-Lecompte, J.C. Maternal antibody decay and antibody-mediated immune responses in chicken pullets fed prebiotics and synbiotics. Poult Sci. 2017, 96, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Ignjatovic, J.; Galli, L. Immune responses to structural proteins of avian infectious bronchitis virus. Avian Pathol. 1995, 24, 313–332. [Google Scholar] [CrossRef]
- Lin, K.Y.; Wang, H.C.; Wang, C.H. Protective effect of vaccination in chicks with local infectious bronchitis viruses against field virus challenge. J. Microbiol. Immunol. Infect. 2005, 38, 25–30. [Google Scholar]
- Martin, M.P.; Wakenell, P.S.; Woolcock, P.; O’Connor, B. Evaluation of the effectiveness of two infectious bronchitis virus vaccine programs for preventing disease caused by a California IBV field isolate. Avian Dis. 2007, 51, 584–589. [Google Scholar] [CrossRef]
- Meir, R.; Krispel, S.; Simanov, L.; Pitcovski, J. Immune Responses to Mucosal Vaccination by the Recombinant S1 and N Proteins of Infectious Bronchitis Virus. Viral Immunol. 2012, 25, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Gelb, J.; Ladman, B.S.; Pope, C.R.; Ruano, J.M.; Brannick, E.M.; Bautista, D.A.; Coughlin, C.M.; Preskenis, L.A. Characterization of nephropathogenic infectious bronchitis virus DMV/1639/11 recovered from Delmarva broiler chickens in 2011. Avian Dis. 2013, 57, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Lim, T.H.; Kim, M.S.; Jang, J.H.; Lee, D.H.; Park, J.K.; Young, H.N.; Lee, J.B.; Park, S.Y.; Choi, I.S.; Song, C.S. Live attenuated nephropathogenic infectious bronchitis virus vaccine provides broad cross protection against new variant strains. Poult. Sci. 2012, 91, 89–94. [Google Scholar] [CrossRef]
- Rosenberger, J.K.; Alphin, R.L.; Krauss, W.S. Cross-protection studies with a Holland strain of IBV. Avian Dis. 1976, 20, 199–201. [Google Scholar] [CrossRef] [PubMed]
- Cook, J.K.A.; Otsuki, K.; Martin, N.R.D.S.; Ellis, M.M.; Huggins, M.B. The secretory antibody response of inbred lines of chickens to avian infectious bronchitis virus infection. Avian Pathol. 1992, 21, 681–692. [Google Scholar] [CrossRef]
- Bacon, L.D.; Hunter, D.B.; Zhang, H.M.; Brand, K.; Etches, R. Retrospective evidence that the MHC (B haplotype) of chickens influences genetic resistance to attenuated infectious bronchitis vaccine strains in chickens. Avian Pathol. 2004, 33, 605–609. [Google Scholar] [CrossRef]
- Joiner, K.S.; Hoerr, F.J.; Ewald, S.J.; van Santen, V.L.; Wright, J.C.; van Ginkel, F.W.; Toro, H. Pathogenesis of infectious bronchitis virus in vaccinated chickens of two different major histocompatibility B complex genotypes. Avian Dis. 2007, 51, 758–763. [Google Scholar] [CrossRef]
- Andrade, L.F.; Villegas, P.; Fletcher, O.J.; Laudencia, R. Evaluation of ciliary movement in tracheal rings to assess immunity against infectious bronchitis virus. Avian Dis. 1982, 26, 805–815. [Google Scholar] [CrossRef] [PubMed]
- Matthijs, M.G.; van Eck, J.H.; de Wit, J.J.; Bouma, A.; Stegeman, J.A. Effect of IBV-H120 vaccination in broilers on colibacillosis susceptibility after infection with a virulent Massachusetts-type IBV strain. Avian Dis. 2005, 49, 540–545. [Google Scholar] [CrossRef]
- Bailey, R.I.; Cheng, H.H.; Chase-Topping, M.; Mays, J.K.; Anacleto, O.; Dunn, J.R.; Doeschl-Wilson, A. Pathogen transmission from vaccinated hosts can cause dose-dependent reduction in virulence. PLoS Biol. 2020, 18, e3000619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leyson, C.M.; Hilt, D.A.; Jordan, B.J.; Jackwood, M.W. Minimum Infectious Dose Determination of the Arkansas Delmarva Poultry Industry Infectious Bronchitis Virus Vaccine Delivered by Hatchery Spray Cabinet. Avian Dis. 2017, 61, 123–127. [Google Scholar] [CrossRef] [PubMed]
- Landman, W.J. The downside of broiler vaccination. Vet. Q. 2012, 32, 121–122. [Google Scholar] [CrossRef] [Green Version]
- Ismail, M.I.; Tan, S.W.; Hair-Bejo, M.; Omar, A.R. Evaluation of the antigen relatedness and efficacy of a single vaccination with different infectious bronchitis virus strains against a challenge with Malaysian variant and QX-like IBV strains. J. Vet. Sci. 2020, 21, e76. [Google Scholar] [CrossRef] [PubMed]
- Aziz, T. Avian Infectious Bronchitis Virus: Infection, Evolution, and Immunity. 2020. Available online: https://www.thepoultrysite.com/articles/avian-infectious-bronchitis-virus-infection-evolution-and-immunity (accessed on 16 January 2021).
- Aston, E.J.; Jordan, B.J.; Williams, S.M.; García, M.; Jackwood, M.W. Effect of Pullet Vaccination on Development and Longevity of Immunity. Viruses 2019, 11, 135. [Google Scholar] [CrossRef] [Green Version]
- Sharif, A.; Ahmad, T. Preventing Vaccine Failure in Poultry Flocks. Retrieved from Google scholar data base on the World Wide Web. 2018. Available online: http://0-dx-doi-org.brum.beds.ac.uk/10.5772/intechopen.79330 (accessed on 16 January 2021).
- Lala, M.K.; Lala, K.R. Thermo stability of vaccines. Indian Pediatr. 2003, 40, 311–319. [Google Scholar]
- Vangroenweghe, F. Good vaccination practice: It all starts with a good vaccine storage temperature. Porc Health Manag. 2017, 3, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butcher, G.D.; Yegani, M. Investigating Vaccine Failure in Poultry Flocks; UF IFAS Extension, University of Florida: Gainesville, FL, USA, 2009. [Google Scholar]
- Dhama, K.; Singh, S.D.; Barathidasan, R.; Desingu, P.A.; Chakraborty, S.; Tiwari, R.; Kumar, M.A. Emergence of avian infectious bronchitis virus and its variants need better diagnosis, prevention and control strategies: A global perspective. Pakistan J. Biol. Sci. 2014, 17, 751–767. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.F.F.; Yan, J.Y.; Hu, Y.W.; Tucker, C.M.; Green, A.R.; Cheng, H.W. Immune Response of Laying Hens Exposed to 30 ppm Ammonia for 25 Weeks. Int. J. Poult. Sci. 2017, 16, 139–146. [Google Scholar] [CrossRef] [Green Version]
- Gudev, D.; Popova-Ralcheva, S. Yanchev, I.; Moneva, Petkov, E.; Ignatova, M. Effect of betaine on egg performance and some blood constituents in laying hens reared indoor under natural summer temperatures and varying levels of air ammonia. Bulg. J. Agric. Sci. 2011, 17, 859–866. [Google Scholar]
- Wei, F.X.; Xu, B.; Hu, X.F.; Li, S.Y.; Wang, L.Y. The Effect of Ammonia and Humidity in Poultry Houses on Intestinal Morphology and Function of Broilers. J. Anim Vet. Adv. 2012, 11, 3641–3646. [Google Scholar]
- Oyetunde, O.O.; Thomson, R.G.; Carlson, H.C. Aerosol exposure of ammonia, dust and Escherichia coli in broiler chickens. Can. Vet. J. 1978, 19, 187–193. [Google Scholar] [PubMed]
- Wang, Y.M.; Meng, Q.P.; Guo, Y.M.; Wang, Y.Z.; Wang, Z.; Yao, Z.L.; Shan, T.Z. Effect of atmospheric ammonia on growth performance and immunological response of broiler chickens. J. Anim. Vet. Adv. 2010, 9(22), 2802–2806. [Google Scholar] [CrossRef]
- Anderson, D.P.; Beard, C.W.; Hanson, R.P. The Adverse Effects of Ammonia on Chickens Including Resistance to Infection with Newcastle Disease Virus. Avian Dis. 1964, 8, 369–379. [Google Scholar] [CrossRef]
- Nimmermark, S.; Lund, V.; Gustafsson, G.; Eduard, W. Ammonia, dust and bacteria in welfare-oriented systems for laying hens. Ann. Agric. Environ. Med. 2009, 16, 103–113. [Google Scholar]
- Purswell, J.L.; Davis, J.D.; Luck, B.D.; Kim, E.J.; Olanrewaju, H.A.; Kiess, A.S.; Branton, S.L. Effects of elevated carbon dioxide concentrations on broiler chicken performance from 28 to 49 days. Int. J. Poult. Sci. 2011, 10, 597–602. [Google Scholar] [CrossRef] [Green Version]
- Broucek, J.; Cermák, B. Emission of harmful gases from poultry farms and possibilities of their reduction. Ekologia 2015, 34, 89–100. [Google Scholar] [CrossRef] [Green Version]
- David, B.; Mejdell, C.; Michel, V.; Lund, V.; Moe, R.O. Air Quality in Alternative Housing Systems may have an Impact on Laying Hen Welfare. Part II-Ammonia. Animals 2015, 5, 886–896. [Google Scholar] [CrossRef] [Green Version]
- Bell, D.D.; Weaver, W.D. Commercial Chicken Production Manual, 5th ed.; Kluwer Academic Publishers: Amsterdam, The Netherlands, 2002; p. 1416. [Google Scholar]
- De Wit, J.J.; Swart, W.A.J.M.; Fabri, T.H.F. Efficacy of infectious bronchitis virus vaccinations in the field: Association between the -IBV IgM response, protection and vaccine application parameters. Avian Pathol 2010, 39, 123–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, G.; Naqi, S. Cytotoxic activity of cells recovered from the respiratory tracts of chickens inoculated with infectious bronchitis virus. Avian Dis. 1997, 41, 690–694. [Google Scholar] [CrossRef]
- Saif, Y.M. Immunosuppression induced by infectious bursal disease virus. Vet. Immunol. Immunopathol. 1991, 30, 45–50. [Google Scholar] [CrossRef]
- Matthijs, M.G.; Ariaans, M.P.; Dwars, R.M.; van Eck, J.H.; Bouma, A.; Stegeman, A.; Vervelde, L. Course of infection and immune responses in the respiratory tract of IBV infected broilers after superinfection with E. coli. Vet. Immunol. Immunopathol. 2009, 127, 77–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinet, W.; Verheye, S.; De Meyer, G.R. Selective depletion of macrophages in atherosclerotic plaques via macrophage-specific initiation of cell death. Trends Cardiovasc. Med. 2007, 17, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Purswell, J.L.; Fritz, B.K.; Branton, S.L.; Leigh, S.A. Effects of system pressure and nozzle type on spray application of avian vaccines. Appl. Eng. Agric. 2008, 24, 785–789. [Google Scholar] [CrossRef]
- Roh, H.J.; Jordan, B.J.; Hilt, D.A.; Ard, M.B.; Jackwood, M.W. Hatchery spray cabinet administration does not damage avian coronavirus infectious bronchitis virus vaccine based on analysis by electron microscopy and virus titration. Avian Dis. 2015, 59, 149–152. [Google Scholar] [CrossRef]
- Jordan, B.J. Spray Application of Infectious Bronchitis Virus Vaccines in the Hatchery: How Efficient are We? In Proceedings of the XXIV Congreso de Avicultura Centroamericano y del Caribe, Antigua, Guatemala, 9–11 November 2016. [Google Scholar]
- Purswell, J.L.; Branton, S.L.; Evans, J.D. Performance of an Automated Whole-House Spray Vaccination System. J. Appl Poult Res. 2019, 28, 215–220. [Google Scholar] [CrossRef]
- Spray Vaccination of Day–Old Chicks at the Hatchery. Available online: https://www.thepoultrysite.com/articles/spray-vaccination-of-dayold-chicks-at-the-hatchery (accessed on 16 January 2021).
- Paniago, M. Spray Vaccination in the Hatcheries, Key Points to Achieve Proper Immunization of the Day-Old Chicks. 2006. Available online: https://5mpublishing.sirv.com/poultry/legacy/focus/contents/ceva/OnlineBulletins/ob_2006/Article-No4-Jan06.pdf (accessed on 16 January 2021).
- Following Best Practice in Administering Hatchery Spray Vaccination. Available online: https://www.wattagnet.com/articles/16484-following-best-practice-in-administering-hatchery-spray-vaccination (accessed on 16 January 2021).
- Perozo, F.; Villegas, P.; Estévez, C.; Alvarado, I.R.; Purvis, L.B.; Williams, S. Protection against infectious bursal disease virulent challenge conferred by a recombinant avian adenoassociated virus vaccine. Avian Dis. 2008, 52, 315–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russell, P.H.; Ezeifeka, G.O. The Hitchner B1 strain of Newcastle disease virus induces high levels of IgA, IgG and IgM in newly hatched chicks. Vaccine 1995, 13, 61–66. [Google Scholar] [CrossRef]
- Toro, H.; Espinoza, C.; Ponce, V.; Rojas, V.; Morales, M.A.; Kaleta, E.F. Infectious bronchitis: Effect of viral doses and routes on specific lacrimal and serum antibody responses in chickens. Avian Dis. 1997, 41, 379–387. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, J.W.; Randall, C.J.; McMartin, D.A.; Dagless, M.D. Immunity following inoculation of H120 and H52 vaccine strains of infectious bronchitis virus into the crop of the domestic fowl. Avian Pathol. 1983, 12, 379–383. [Google Scholar] [CrossRef]
- Ratanasethakul, C.; Cumming, R.B. Immune response of chickens to various routes of administration of Australian infectious bronchitis vaccine. Aust. Vet. J. 1983, 60, 214–216. [Google Scholar] [CrossRef]
- Guide to Successful Spray and Water Vaccination for Poultry Farmers. Available online: https://www.fwi.co.uk/livestock/poultry/guide-successful-spray-water-vaccination-poultry-farmers (accessed on 16 January 2021).
- Box, P.G.; Ellis, K.R. Infectious bronchitis in laying hens: Interference with response to emulsion vaccine by attenuated live vaccine. Avian Pathol. 1985, 14, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Danforth, H.D.; Lee, E.H.; Martin, A.; Dekich, M. Evaluation of a gel-immunization technique used with two different Immucox vaccine formulations in battery and floor-pen trials with broiler chickens. Parasitol. Res. 1997, 83, 445–451. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, T.; Lee, E.H. A gel delivery system for coccidiosis vaccine: Uniformity of distribution of oocysts. Can. Vet. J. 2000, 41, 613–616. [Google Scholar] [PubMed]
- Chapman, H.; Cherry, T.; Danforth, H.; Richards, G.; Shirley, M.; Williams, R. Sustainable coccidiosis control in poultry production: The role of live vaccines. Int. J. Parasitol. 2002, 32, 617–629. [Google Scholar] [CrossRef]
- Godoy, A.; Jordan, B.; El-Attrache, J. Replication dynamics of a live Infectious bronchitis virus vaccine in broiler chicks vaccinated by different routes. In Proceedings of the 20th World Veterinary Poultry Association Congress, Edinburgh, Scotland, 4–8 September 2017; p. 362. [Google Scholar]
- Tucciarone, C.M.; Franzo, G.; Bianco, A.; Berto, G.; Ramon, G.; Paulet, P.; Koutoulis, K.C.; Cecchinato, M. Infectious bronchitis virus gel vaccination: Evaluation of Mass-like (B-48) and 793/B-like (1/96) vaccine kinetics after combined administration at 1 day of age. Poult Sci. 2018, 97, 3501–3509. [Google Scholar] [CrossRef]
- Albanese, G.A.; Tensa, L.R.; Aston, E.J.; Hilt, D.A.; Jordan, B.J. Evaluation of a coccidia vaccine using spray and gel applications. Poult. Sci. 2018, 97, 1544–1553. [Google Scholar] [CrossRef]
- Tensa, L.R.; Jordan, B.J. Comparison of the application parameters of coccidia vaccines by gel and spray. Poult. Sci. 2019, 98, 634–641. [Google Scholar] [CrossRef] [PubMed]
- Leerdam, B.V.; Bosman, H. Vaccination Index: A new approach to evaluate Elisa. World Poul. Doetinchem. Elsevier 2010, 26, 12–15. [Google Scholar]
- Roberts, J.R.; Souillard, R.; Bertin, J. Avian diseases which affect egg production and quality. In Improving the Safety and Quality of Eggs and Egg Products; Woodhead Publishing: Cambridge, UK, 2011; pp. 376–393. [Google Scholar]
- Hassan, M.S.H.; Abdul-Careem, M.F. Avian Viruses that Impact Table Egg Production. Animals. 2020, 10, 1747. [Google Scholar] [CrossRef]
- Crispo, M.; Blackall, P.; Khan, A.; Shivaprasad, H.L.; Clothier, K.; Sentíes-Cué, C.G.; Cooper, G.; Blakey, J.; Pitesky, M.; Mountainspring, G.; et al. Characterization of an outbreak of infectious coryza (Avibacterium paragallinarum) in commercial chickens in central California. Avian Dis. 2019, 63, 486–494. [Google Scholar] [CrossRef]
- Wang, H.; Cong, F.; Guan, J.; Xiao, L.; Zhu, Y.; Lian, Y.; Huang, R.; Chen, M.; Guo, P. Development of a sensitive and specific xMAP assay for detection of antibodies against infectious laryngotracheitis and bronchitis viruses. Virol J. 2018, 15, 1–18. [Google Scholar] [CrossRef]
- Pattison, M.; Gough, R.E.; Dawson, P.S. The effect of live infectious bronchitis vaccine on the course of infectious laryngotracheitis. Vet. Rec. 1971, 75, 643–664. [Google Scholar] [CrossRef]
- Nakamura, K.; Cook, J.K.A.; Otsuki, K.; Huggins, M.B.; Frazier, J.A. Comparative study of respiratory lesions in two chicken lines of different susceptibility infected with infectious bronchitis virus: Histology, ultrastructure and immunohistochemistry. Avian Pathol. 1991, 20, 241–257. [Google Scholar] [CrossRef] [PubMed]
- Tawakol, M.M.; Nabil, N.M.; Samy, A. Evaluation of bacteriophage efficacy in reducing the impact of single and mixed infections with Escherichia coli and infectious bronchitis in chickens. Infect. Ecol. Epidemiol. 2019, 9, 1686822. [Google Scholar] [CrossRef] [Green Version]
- Goodwin, M.A.; Brown, J.; Smeltzer, M.A.; Girshick, T.; Miller, S.L.; Dickson, T.G. Relationship of common avian pathogen antibody-titres in so-called chicken anaemia agent (CAA)-antibodypositive chicks to titers in CAA-antibody-negative chicks. Avian Dis. 1992, 36, 356–358. [Google Scholar] [CrossRef]
- Fujisawa, S.; Murata, S.; Takehara, M.; Katakura, K.; Hmoon, M.M.; Win, S.Y.; Ohashi, K. Molecular detection and genetic characterization of Mycoplasma gallisepticum, Mycoplama synoviae, and infectious bronchitis virus in poultry in Myanmar. BMC Vet. Res. 2019, 15, 261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hopkins, S.R.; Yoder, H.W. Influence of infectious bronchitis strains and vaccines on the incidence of Mycoplasma synoviae airsacculitis. Avian Dis. 1982, 26, 741–775. [Google Scholar] [CrossRef] [PubMed]
- Uenaka, T.; Kishimoto, I.; Sato, S.; Animas, S.B.; Ito, T.; Otsuki, K.; Cook, J.K.A. Intracloacal infection with avian infectious bronchitis virus. Avian Pathol. 1998, 27, 309–312. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Types of Spray Vaccine | Droplet Size | Comments |
|---|---|---|
| ine spray | 50–80 µL | More suitable for booster vaccinations on the farm; however, vaccine can penetrate into respiratory epithelial cells causing a post-vaccination reaction if applied too early in chicks. |
| Coarse spray | 120–150 µL | Usually applied in day old chicks in hatcheries to administer respiratory vaccines and more convenient with high immune response. |
| Larger droplet sizes | <250 µL | Commonly applied to protect against Coccidia oocysts where chicks can easily gulp the droplet vaccine from their fluff. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhuiyan, M.S.A.; Amin, Z.; Bakar, A.M.S.A.; Saallah, S.; Yusuf, N.H.M.; Shaarani, S.M.; Siddiquee, S. Factor Influences for Diagnosis and Vaccination of Avian Infectious Bronchitis Virus (Gammacoronavirus) in Chickens. Vet. Sci. 2021, 8, 47. https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci8030047
Bhuiyan MSA, Amin Z, Bakar AMSA, Saallah S, Yusuf NHM, Shaarani SM, Siddiquee S. Factor Influences for Diagnosis and Vaccination of Avian Infectious Bronchitis Virus (Gammacoronavirus) in Chickens. Veterinary Sciences. 2021; 8(3):47. https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci8030047
Chicago/Turabian StyleBhuiyan, Md. Safiul Alam, Zarina Amin, Ag Muhammad Sagaf Abu Bakar, Suryani Saallah, Noor Hydayaty Md. Yusuf, Sharifudin Md. Shaarani, and Shafiquzzaman Siddiquee. 2021. "Factor Influences for Diagnosis and Vaccination of Avian Infectious Bronchitis Virus (Gammacoronavirus) in Chickens" Veterinary Sciences 8, no. 3: 47. https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci8030047