
Multilocus Genotyping of Giardia duodenalis Occurring in Korean Native Calves
by
 ,
,
Sang-Ik Oh
1 ,
,
Suk-Han Jung
1, 
Han-Kyoung Lee
2,
Changyong Choe
1,
Tai-Young Hur
1 and
Kyoung-Min So
1,* 1
Division of Animal Diseases & Health, National Institute of Animal Science, Rural Development Administration, Wanju 55365, Korea
2
Happy Veterinary Clinic, 289 Nambuk-ro, Gimje 54373, Korea
*
Author to whom correspondence should be addressed.
Vet. Sci. 2021, 8(7), 118; https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci8070118
Submission received: 24 May 2021
/
Revised: 14 June 2021
/
Accepted: 21 June 2021
/
Published: 23 June 2021
Abstract
:Giardia duodenalis is one of the most widely occurring zoonotic protozoan parasites causing diarrheal disease in calves. This study aimed to investigate the prevalence of G. duodenalis in Korean native calves and elucidate the causal factors associated with giardiasis in these animals. We investigated the sequences of three genes (ssu, bg, and gdh) of G. duodenalis in fecal samples collected from 792 Korean native calves during 2019–2020. Data were analyzed with regard to age, sex, sampling season, and the fecal sample type (based on its physical characteristics). The samples were screened for the three genes mentioned above, and 44 samples (5.6%) were G. duodenalis-positive. Polymerase chain reaction results showed a significantly higher prevalence of the infection in calves aged ≥1 month and in those with watery diarrhea in spring season. Screening for the gene sequences ssu (87.5%), bg (96.2%), and gdh (96.7%) revealed that most of the G. duodenalis-positive samples belonged to assemblage E. Four of the G. duodenalis-positive samples belonged to the zoonotic assemblage A. This study highlights the importance of continuous surveillance of genetic mutations in G. duodenalis for the detection of emerging variants of zoonotic G. duodenalis in calves.
1. Introduction
Giardia duodenalis (synonyms Giardia intestinalis, Giardia lamblia) is a zoonotic protozoan parasite that causes intestinal diseases in various host species. Infection occurs following ingestion of infective stages of this pathogen (cysts) via the fecal–oral route or drinking contaminated water [1,2,3]. Giardiasis in cattle is caused by G. duodenalis and is one of the most important parasitic diseases associated with diarrhea in calves [4]. Although the pathogenesis of this protozoan parasite is not fully understood yet, the pathogen is known to cause diffuse shortening of brush border microvilli and malabsorption of water, sodium, and glucose [5]. Consequently, cattle infected with G. duodenalis show clinical signs such as malabsorptive diarrhea and reduced weight gain [5,6]. G. duodenalis has a broad host range, including humans, cattle, sheep, goats, pigs, dogs, cats, and rats, and is thus a potential threat to public health [7,8]. Previous studies have reported that cattle are one of the important hosts responsible for zoonotic transmission of giardiasis to humans, and molecular epidemiological analysis of G. duodenalis from cattle is required to elucidate the role of cattle as a potential zoonotic reservoir [9].
Recent studies have reported that G. duodenalis consists of eight distinct assemblages (A to H), which appear to have different host ranges [7,10]. Among these assemblages, A and B are infective in humans and many other animals, including ruminants, dogs, rodents, and marine mammals [2,7], whereas other assemblages are mostly host-specific; assemblage E is infective in ruminants and pigs, C and D in dogs, and G in rodents [7]. Thus far, assemblages A, B, and E have been detected in cattle [7,8]. Interestingly, previous studies suggested that calves are more frequently infected with zoonotic assemblages A and B than adult cattle are [6,11].
According to previous studies, the prevalence of G. duodenalis was 13.1% (2013–2015) and 10.0% (2017) in Korean native calves with diarrhea and normal feces, respectively [12,13]. These studies also reported that all the analyzed fecal samples from diarrheic calves in Korea in 2013–2015 belonged to assemblage E [12], and 2.2% of those analyzed from normal fecal samples in 2017 belonged to assemblage A [13]. These two studies indicated that G. duodenalis is a major concern of infection in Korean native cattle farms, and zoonotic G. duodenalis (assemblage A) could have emerged in Korea in 2017. Therefore, it is essential to continuously monitor by testing fecal samples from calves and adult cattle to determine the prevalence and identify the genotypes of G. duodenalis responsible for causing zoonotic infections.
Most of the previous studies conducted in Korea have identified G. duodenalis in calves by a single gene locus [12,13]. Therefore, in this study, we broadened the scope by performing a large-scale analysis of G. duodenalis, with multilocus genotyping based on sequences of the small subunit (SSU) rRNA, β-giardin, and glutamate dehydrogenase (GDH). We also analyzed the factors associated with the occurrence of G. duodenalis, including age and sex of calves, sampling season, and the fecal sample type (based on its physical characteristics).
2. Materials and Methods
2.1. Sample Collection
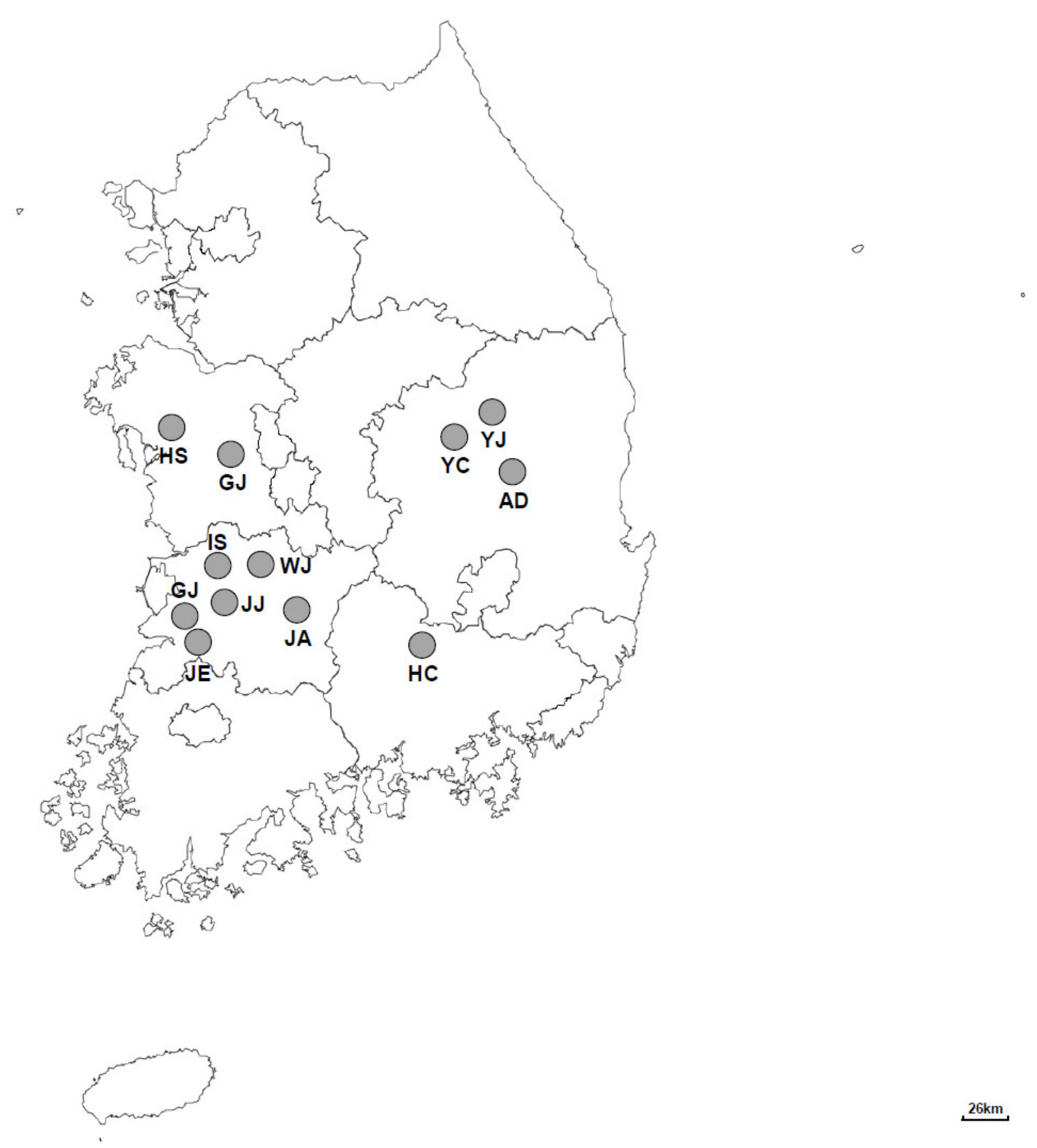
A total of 792 fecal samples were obtained from 291 Korean native cattle farms between May 2019 and September 2020. These cattle farms were located in 12 different regions (Gimje, Jeongeup, Youngju, Iksan, Gongju, Hongseong, Hapcheon, Jeonju, Andong, Wanju, Jinan, and Yecheon) in the central region of the Republic of Korea (Figure 1). The details of each calf were recorded, including geographical location, age, sex, and the sample collection date. The type of feces was also recorded and categorized as normal, pasty, watery, or hemorrhagic according to its physical characteristics. Fresh feces from calves were collected in 50-milliliter stool specimen collection tubes immediately after defecation, using polyethylene gloves. All the samples were packed in boxes with icepacks and sent to the laboratory. Feces were stored at 4 °C until the extraction of genomic DNA.
2.2. DNA Extraction and Polymerase Chain Reaction (PCR) Amplification
Genomic DNA was extracted from fecal samples using the QIAamp fast DNA Stool Mini Kit (QIAgen, Hilden, Germany) according to the manufacturer’s instructions. To amplify the SSU rRNA gene (ssu) and β-giardin gene (bg) fragments from G. duodenalis, nested PCR was conducted for each gene using primer sets described in a previous study [14]. For amplification of the GDH gene (gdh) fragments, semi-nested PCR was performed as reported previously [14].
2.3. Sequencing and Phylogenetic Analysis
All the fecal samples identified as G. duodenalis-positive by PCR were used for sequencing. Amplicons were sequenced directly in both directions using the Bigdye Terminator Cycle Sequencing Ready Reaction Kit V.3.1 (Applied Biosystems, Foster City, CA, USA) on the 3730XL Capillary DNA sequencer machine (Applied Biosystems) by SolGent Co., Ltd. (Daejeon, Korea). If the sequencing result was unclear, the experiment was repeated until a clear result was obtained. The obtained sequences of ssu, bg, and gdh were identified from the public data libraries of the National Center for Biotechnology Information (NCBI, USA) using the basic local alignment search tool (BLAST) and then aligned using Clustal Omega Alignment Tools (http://www.ebi.ac.uk/tools/msa/clustalo/, accessed on 9 April 2021). The nucleotide sequences of the G. duodenalis-positive samples were deposited in the NCBI GenBank database: MW876381–MW876412 for the SSU rRNA locus, MW876413–MW876438 for the β-giardin locus, and MW876439–MW876467 for the GDH locus. For phylogenetic analysis, we collected the nucleotide sequences of G. duodenalis variants from various host species in various countries from the NCBI GenBank database. Phylogenetic trees were constructed using the neighbor-joining method. The sequence data were sampled using 1000 replicates for bootstrap analysis using the Mega-X software (version 10.0.4) with the Kimura two-parameter option.
2.4. Statistical Analysis
A binominal logistic regression analysis was used to compare the prevalence of G. duodenalis by calf age and sex, sampling season, and fecal sample type. The variables were grouped as follows: six age groups (≤2, 3–4, 5–6, 7–8, 9–12, and >12 weeks), three groups for sex (male, female, and unknown), four groups for sampling season (spring, summer, autumn, and winter), and four fecal types (normal, pasty, watery, and hemorrhagic feces). The level of significance was set at p < 0.05. The odds ratio (OR) with 95% confidence intervals (CI) was estimated to show the strength of associations. Statistical analysis was performed using SPSS version 26.0 (IBM, Armonk, NY, USA). In addition, the Kappa (κ) values, which range from 1 to 0, were interpreted according to a previous study [15], where <0: no agreement; 0–0.20: slight agreement; 0.21–0.40: fair agreement; 0.41–0.60: moderate agreement; 0.61–0.80: substantial agreement; and 0.81–1.00: almost perfect agreement.
3. Results
3.1. Prevalence and Risk Factors of G. duodenalis
The prevalence of G. duodenalis was 5.6% (n = 44) among 792 calves from 37 farms (12.7% of 271 tested farms), as determined by PCR (Table 1). G. duodenalis was most prevalent in calves aged 7–8 weeks (15.4% of 26 calves) and ≥12 weeks (15.4% of 13 calves), followed by those aged 9–12 weeks (15.2% of 46 calves) and 5–6 weeks (14.0% of 107 calves). The calves aged less than 1 month showed a lower prevalence of G. duodenalis compared with those aged more than 1 month. There was no significant difference between the occurrence of G. duodenalis in male (5.7%) and female (5.6%) calves. With respect to seasons, the prevalence of G. duodenalis was 6.7% in spring (March–May), 6.0% in summer (June–August), 7.4% in autumn (September–November), and 1.5% in winter (December–February). Furthermore, G. duodenalis was frequently detected in hemorrhagic (12.5%) and watery (9.8%) diarrheal feces, but rarely detected in pasty (2.9%) and normal (2.3%) feces. Based on the statistical analysis, there was significantly lower prevalence in calves under 2 weeks of age (OR = 0.084, p = 0.006) than in those over 2 weeks old. In addition, G. duodenalis was significantly more prevalent in watery feces (OR = 4.319, p = 0.001) and in fecal samples from the spring (OR = 6.446, p = 0.018).
3.2. Detection of G. duodenalis by the Amplification of ssu, gdh, and bg
Among the 44 G. duodenalis-positive fecal samples, 32 fecal samples (72.7%) were ssu-positive, 26 samples (59.1%) were bg-positive, and 30 samples (68.2%) were gdh-positive (Figure 2). Of the 32 ssu rRNA gene-positive samples, 62.5% (n = 20) were bg-positive and 65.6% (n = 21) were gdh-positive. Only 31.8% (n = 14) of samples tested positive for all three genes. The κ-values of 0.68 and 0.66 suggest a good agreement between ssu- and bg-positive samples and ssu- and gdh-positive samples, respectively. A moderate agreement was observed between bg- and gdh-positive samples (κ = 0.59).
3.3. Nucleotide Sequencing
A total of 28 sequences were identical among the 32 ssu-positive sequences, three were different from the 28 sequences but identical to each other, and one sequence showed a marginal difference from the other sequences. Eight and six sequences were identified in bg and gdh sequences, respectively (Table 2 and Table 3). All the sequences were deposited in the NCBI GenBank database under the respective accession numbers.
3.4. Phylogenetic Analysis
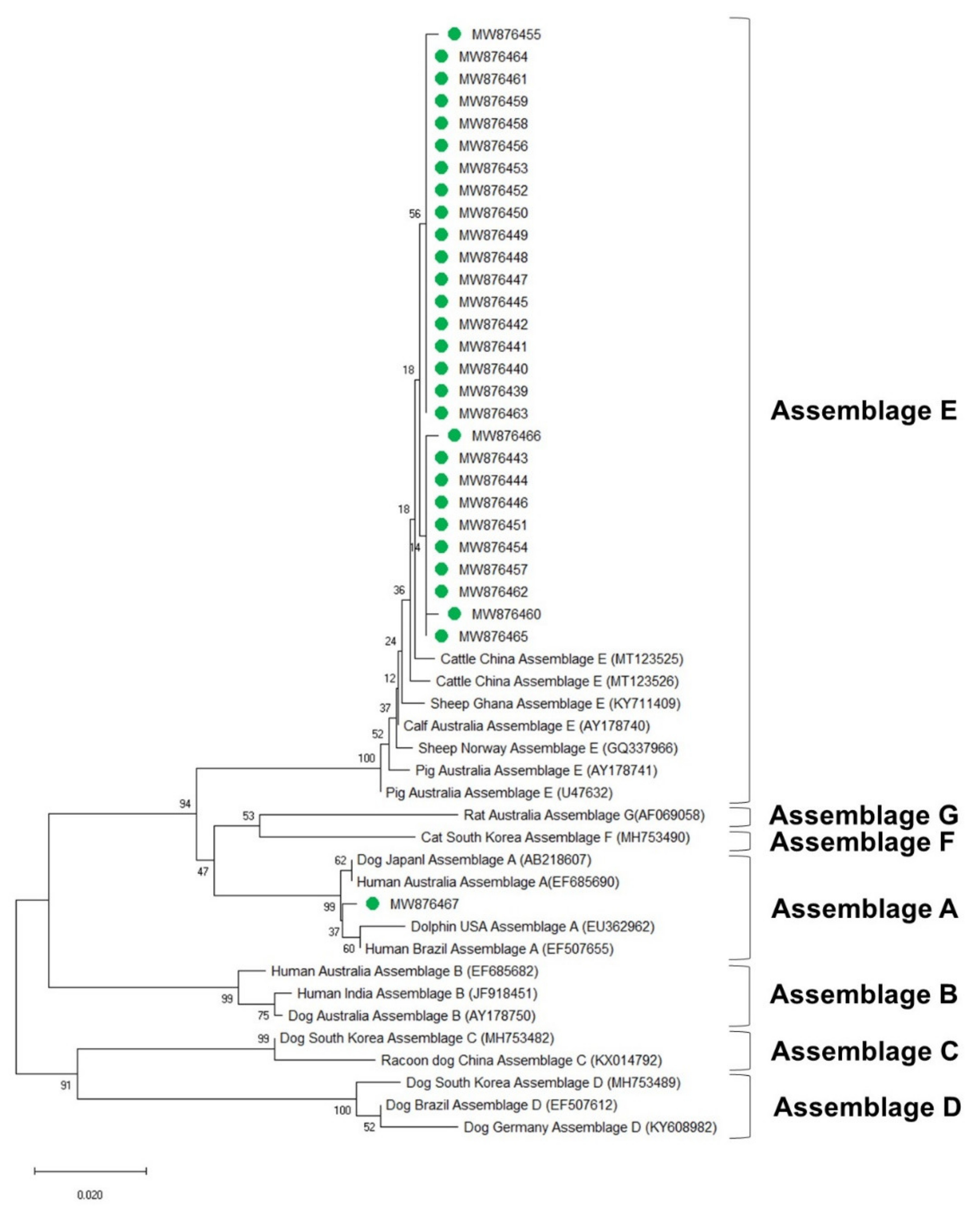
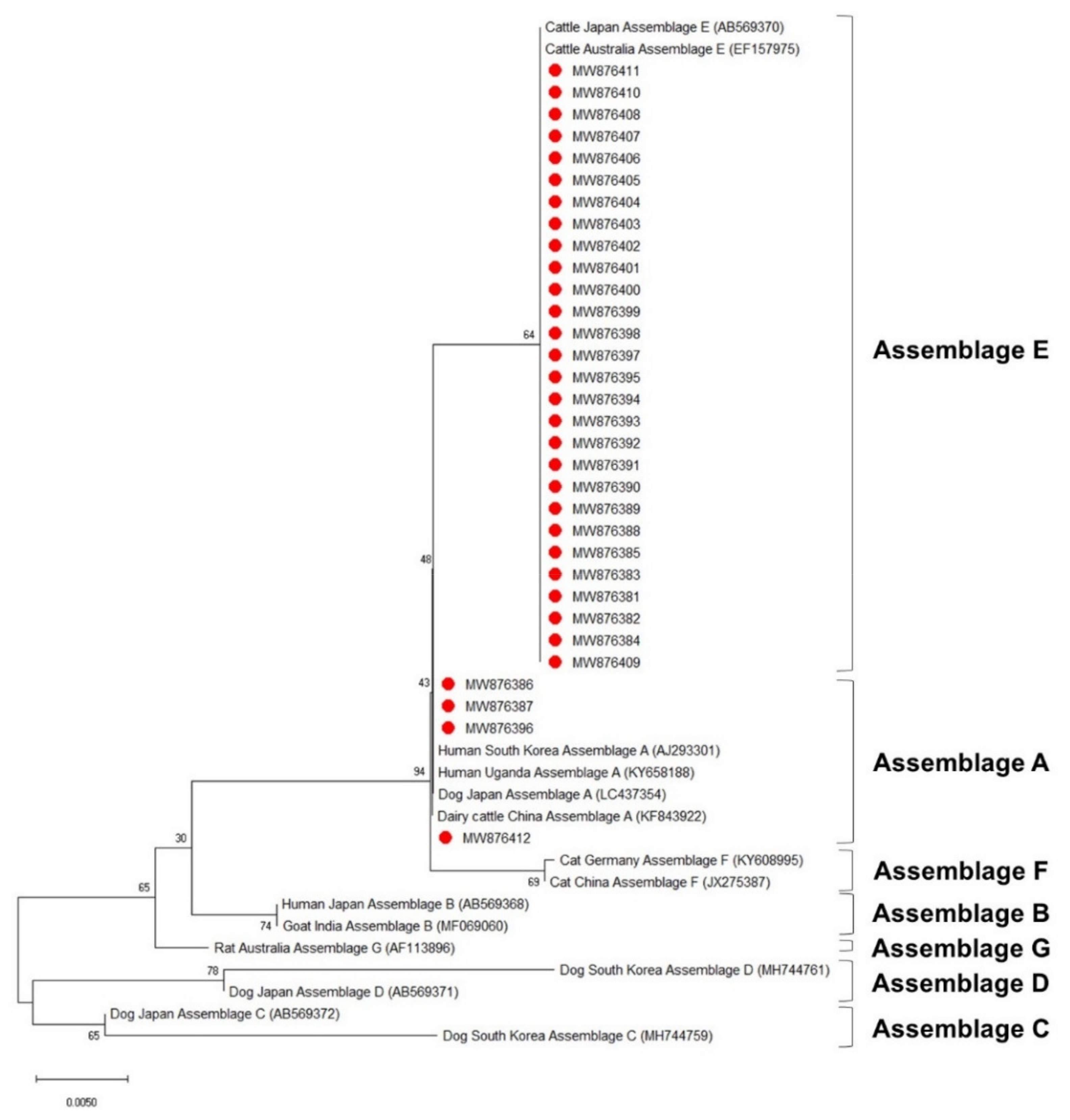
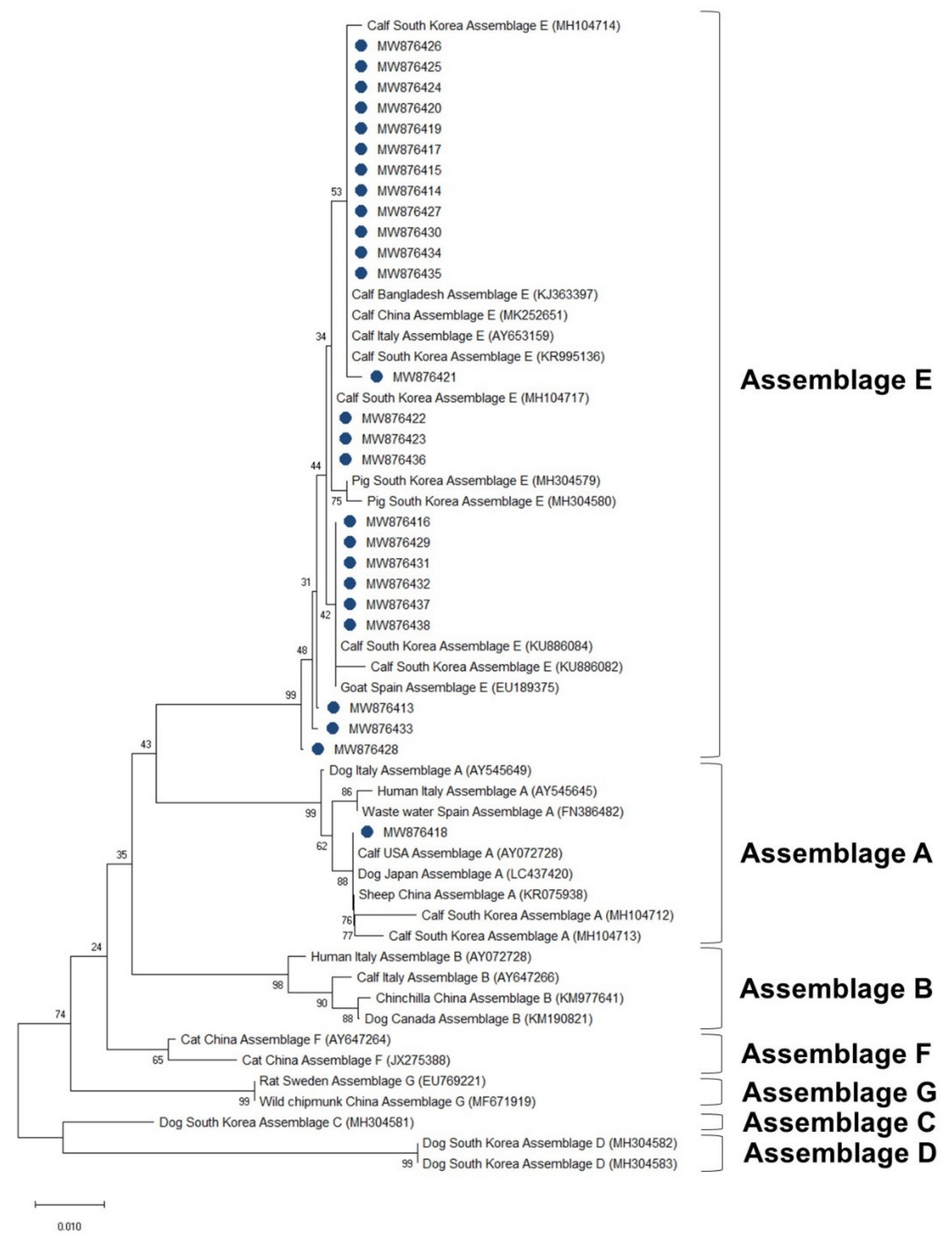
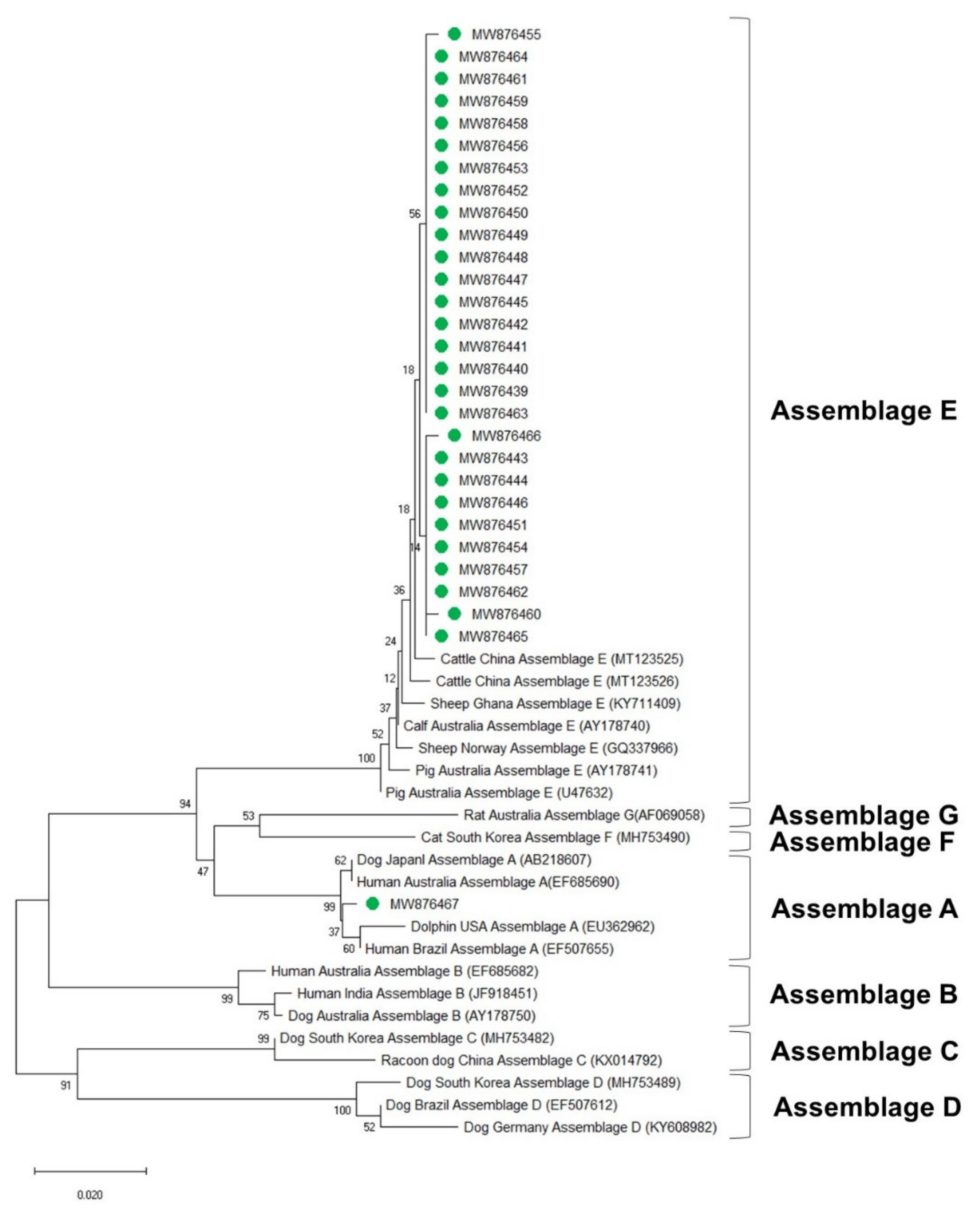
The phylogenetic analysis results of the ssu, bg, and gdh sequences are shown in Figure 3, Figure 4 and Figure 5, respectively. Most of the ssu-positive G. duodenalis samples from Korean native calves belonged to assemblage E (87.5%, n = 28). Four of the ssu-positive samples (12.5%) belonged to assemblage A (Figure 3). A total of 25 bg-positive samples (96.2%) belonged to assemblage E, and only one sample (3.8%) belonged to assemblage A (Figure 4). All gdh-positive fecal samples (96.7%) belonged to assemblage E except one (3.3%), which belonged to assemblage A (Figure 5).
4. Discussion
The present study (2019–2020) indicated that 5.6% of the tested Korean native calves were infected with G. duodenalis, which is lower than the number of calves reportedly infected in 2013–2015 (13.1%, n = 590) and 2017 (10.0%, n = 90) [12,13]. The rate of infection (5.6%) observed in this study was also lower than that reported from other countries, including the United States (44%), Belgium (31.3%), Egypt (13.3%), Vietnam (10.2%), China (9.2%), and Thailand (6.0%) [16,17,18,19,20,21]. These differences may be ascribed to the fact that the determinants of infection rates are varied and affected by many factors, including the geoecological conditions, diagnostic methods, sample size, sample collection period, and age of the sampled animals [6,12,22]. Therefore, the current study investigated the influential factors behind the occurrence of G. duodenalis infection in Korean native calves.
As shown in Table 1, the neonatal calves (≤2 weeks old) were associated (OR = 0.084, p = 0.006) with prevalence of G. duodenalis in feces. G. duodenalis was more prevalent in calves aged ≥1 month (14.6%) than in those aged <1 month (2.7%). This result was in line with the findings of previous studies in China in 2016 and 2020, which reported the highest prevalence of G. duodenalis infection in calves aged 1–3 months (19.6% and 10.7%, respectively) [19,23]. Thus, given that neonatal calf diarrhea commonly occurs in calves at around 2 weeks of age, G. duodenalis may not be the main pathogen responsible for this disease in these animals [24]. However, there is a need to continuously monitor this protozoan parasite to prevent outbreaks of giardiasis in calf-breeding farms. In the present study, no significant difference was observed in the prevalence of the infection between male (5.7%, p = 0.998) and female (5.6%, p = 0.161) calves, which was consistent with the findings of a previous study in Korea [25]. Furthermore, the prevalence of G. duodenalis infection was the highest in autumn (7.4%, n = 6 out of 81), followed by spring (6.7%, n = 17 out of 256), summer (6.0%, n = 19 out of 320), and winter (2.7%, n = 2 out of 135). Among the four seasons, the fecal samples of calves from the spring were significantly (OR = 6.446, p = 0.018) associated with the presence of G. duodenalis. The current findings are consistent with the findings of previous studies in Korea [12] and China [23], in which G. duodenalis infection was most frequently detected in the warm season. With regard to fecal sample type, the occurrence rate of G. duodenalis was the highest in calves with hemorrhagic feces (12.5%), followed by watery diarrheal (9.8%), pasty (2.9%), and normal (2.3%) feces. According to the binominal logistic regression model analysis, the watery fecal samples were significantly (OR = 4.319, p = 0.001) associated with the occurrence of G. duodenalis. This result is in accordance with the findings of a previous study in Korea, which reported a statistical association between G. duodenalis detection and the type of fecal sample [12]. However, other studies reported no significant association between the fecal sample type and G. duodenalis infection [13,26]. These overall results suggest that veterinarians and farm workers should suspect G. duodenalis infection in calves (≥1 month of age) with watery diarrhea in the spring. Moreover, hemorrhagic diarrhea could also be a result of co-infection with G. duodenalis, and thus, the calves must be screened for other pathogenic infections that could cause hemorrhagic diarrhea.
The results of this study highlight that there is no perfect PCR primer set to detect all G. duodenalis-positive samples at once. Among the three PCR primer sets used in this study, the SSU rRNA gene primer set was more effective in detecting G. duodenalis than the protein-coding gene (β-giardin and GDH) primer sets. This finding is in agreement with previous studies [27,28]. Although the three PCR primer sets (ssu, bg, and gdh) showed a markedly good agreement with each other according to their κ-values, laboratory PCR diagnosis of G. duodenalis should be conducted using different primers for detecting all infected calves. Further studies are needed to establish a PCR diagnostic standard that can determine G. duodenalis infection with high sensitivity and specificity.
G. duodenalis exhibits host-specific characteristics that vary depending on their assemblages [4,10]. The phylogenetic analysis in this study suggested that the SSU rRNA gene is strongly conserved among G. duodenalis assemblages. The ssu sequence analysis revealed that four sequences (MW876386, MW876387, MW876396, and MW876412) belong to zoonotic assemblage A, whereas bg and gdh had only one sequence each belonging to assemblage A (MW876418 and MW876467, respectively). The bg (MW876418) and gdh sequences (MW876467) belonging to assemblage A were derived from two samples that also contained two ssu sequences belonging to assemblage A (MW876386 and MW876412). These different phylogenetic results according to the sequences of each locus suggest that assigning one locus is insufficient for laboratory diagnosis and identification of the assemblage of G. duodenalis.
In assemblage E, we found only one sequence at the ssu locus but seven at the bg locus and five at the gdh locus. The gene bg showed the greatest variability, followed by gdh, then ssu. The results implied that bg is the most feasible target for determination of the genetic diversity of G. duodenalis. A total of seven different sequences were identified from the bg gene sequence of assemblage E, three of which were novel types compared with a previous study on calves in Korea [12]. This result suggests the occurrence of genetic mutations in assemblage E of G. duodenalis infecting calves in Korea. Therefore, continuous monitoring is needed to detect emerging novel types of G. duodenalis and obtain updated genetic information to prevent the outbreak of infections caused by a novel G. duodenalis strain. Moreover, many recent studies commonly performed multilocus genotyping with three genes, namely bg, gdh, and triose phosphate isomerase (tpi), which could identify multilocus genotypes (MLGs) of G. duodenalis [19,23]. Thus, further experiments with tpi sequencing are needed to reveal the MLGs of G. duodenalis from Korean native calves. The prevalence of G. duodenalis belonging to assemblage A (9.1%) was lower in this study than that observed in previous studies in China, the USA, Belgium, and Portugal [16,17,22,29]. According to the results of the present study, the four calves that harbored zoonotic G. duodenalis (belonging to assemblage A) were all aged 1–2 months and had watery diarrhea. These results are in contrast to the findings of a previous study, which showed that G. duodenalis assemblage A was identified in calves aged 1 month with normal fecal discharge [13]. Furthermore, despite the small number of fecal samples collected in autumn, most of the G. duodenalis-positive samples (75%) belonging to assemblage A were detected in this season. These results are similar to the findings of a previous study, which reported that all G. duodenalis-positive samples belonging to assemblage A were isolated in autumn [30]. As G. duodenalis belonging to assemblage A is not host-specific, some vectors with increased activity in autumn (e.g., humans, wild rodents, or dogs), may play an important role in pathogen transmission. Therefore, further studies are needed to identify the potential vectors for zoonotic transmission of G. duodenalis from cattle farms. The present findings provide valuable information for similar monitoring studies in the future.
5. Conclusions
This study provides updated data regarding the prevalence of and the factors associated with G. duodenalis infection in Korean native calves. The prevalence of G. duodenalis infection was 5.7% (12.7% at the farm level) in this study. Calves aged ≥1 month or producing watery feces in the spring should be carefully monitored by veterinarians to prevent G. duodenalis transmission to healthy calves. In addition, this is the first multilocus genotyping study of G. duodenalis from the feces of native calves in Korea. The results highlight that analysis of the ssu gene sequence is an effective tool to detect the presence of G. duodenalis in fecal samples and determine their assemblages. The bg gene is useful to investigate the genetic diversity in G. duodenalis. A total of four out of 792 samples (0.5%) belonged to assemblage A as identified by ssu sequencing and were potential zoonotic pathogens. The present findings could help to understand the genetic relationships between G. duodenalis variants distributed across Korea.
Author Contributions
Conceptualization, S.-I.O. and K.-M.S.; methodology, S.-I.O. and K.-M.S.; software, S.-I.O.; validation, S.-I.O. and K.-M.S.; formal analysis, S.-I.O.; investigation, S.-H.J., C.C., and K.-M.S.; resources, H.-K.L. and C.C.; data curation, S.-I.O. and K.-M.S.; writing—original draft preparation, S.-I.O.; writing—review and editing, S.-I.O. and K.-M.S.; visualization, S.-I.O.; supervision, K.-M.S. and T.-Y.H.; project administration, K.-M.S.; funding acquisition, K.-M.S., C.C., and T.-Y.H. All authors have read and agreed to the published version of the manuscript.
Funding
This work was carried out with the support of “Cooperative Research Program for Agriculture Science and Technology Development (Project title: Research of occurrence and control of giardia, cryptosporidium diseases in Hanwoo, Project No. PJ01433101)”, Rural Development Administration, Republic of Korea.
Institutional Review Board Statement
All experiments in this study were approved by the Institutional Animal Care and Use Committee at the National Institute of Animal Science, Republic of Korea (approval number: NIAS 2019-327).
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors would like to thank Mi-Suk Kim for her technical support. We are also grateful to the staff of Happy Veterinary Clinic for their assistance to collect fecal samples, and thankful to the farmers who participated in this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Cheun, H.-I.; Kim, C.-H.; Cho, S.-H.; Ma, D.-W.; Goo, B.-L.; Na, M.-S.; Youn, S.-K.; Lee, W.-J. The first outbreak of giardiasis with drinking water in Korea. Osong Public Health Res. Perspect. 2013, 4, 89–92. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.; Xiao, L. Zoonotic potential and molecular epidemiology of Giardia species and giardiasis. Clin. Microbiol. Rev. 2011, 24, 110–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koloren, Z.; Seferoğlu, O.; Karanis, P. Occurency of Giardia duodenalis assemblages in river water sources of Black Sea, Turkey. Acta Trop. 2016, 164, 337–344. [Google Scholar] [CrossRef]
- Thompson, R.A. The zoonotic significance and molecular epidemiology of Giardia and giardiasis. Vet. Parasitol. 2004, 126, 15–35. [Google Scholar] [CrossRef]
- Buret, A.G. Mechanisms of epithelial dysfunction in giardiasis. Gut 2007, 56, 316–317. [Google Scholar] [CrossRef] [PubMed]
- Geurden, T.; Vercruysse, J.; Claerebout, E. Is Giardia a significant pathogen in production animals? Exp. Parasitol. 2010, 124, 98–106. [Google Scholar] [CrossRef]
- Bartelt, L.A.; Sartor, R.B. Advances in understanding Giardia: Determinants and mechanisms of chronic sequelae. F1000Prime Rep. 2015, 7, 62. [Google Scholar] [CrossRef] [Green Version]
- Heyworth, M.F. Giardia duodenalis genetic assemblages and hosts. Parasite 2016, 23, 13. [Google Scholar] [CrossRef] [Green Version]
- Smith, H.V.; Cacciò, S.M.; Tait, A.; McLauchlin, J.; Thompson, R.A. Tools for investigating the environmental transmission of Cryptosporidium and Giardia infections in humans. Trends Parasitol. 2006, 22, 160–167. [Google Scholar] [CrossRef] [PubMed]
- Monis, P.T.; Caccio, S.M.; Thompson, R.A. Variation in Giardia: Towards a taxonomic revision of the genus. Trends Parasitol. 2009, 25, 93–100. [Google Scholar] [CrossRef]
- Khan, S.M.; Debnath, C.; Pramanik, A.K.; Xiao, L.; Nozaki, T.; Ganguly, S. Molecular evidence for zoonotic transmission of Giardia duodenalis among dairy farm workers in West Bengal, India. Vet. Parasitol. 2011, 178, 342–345. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; VanBik, D.; Kim, H.; Cho, A.; Kim, J.; Byun, J.; Oem, J.; Oh, S.; Kwak, D. Prevalence and molecular characterisation of Giardia duodenalis in calves with diarrhoea. Vet. Rec. 2016, 178, 633. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-J.; Han, D.-G.; Ryu, J.-H.; Chae, J.-B.; Chae, J.-S.; Yu, D.-H.; Park, J.; Park, B.-K.; Kim, H.-C.; Choi, K.-S. Identification of zoonotic Giardia duodenalis in Korean native calves with normal feces. J. Parasitol. Res. 2018, 117, 1969–1973. [Google Scholar] [CrossRef] [PubMed]
- Gillhuber, J.; Pallant, L.; Ash, A.; Thompson, R.A.; Pfister, K.; Scheuerle, M.C. Molecular identification of zoonotic and livestock-specific Giardia-species in faecal samples of calves in Southern Germany. Parasit. Vectors 2013, 6, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Landis, J.R.; Koch, G.G. The measurement of observer agreement for categorical data. Biometrics 1977, 33, 159–174. [Google Scholar] [CrossRef] [Green Version]
- Geurden, T.; Geldhof, P.; Levecke, B.; Martens, C.; Berkvens, D.; Casaert, S.; Vercruysse, J.; Claerebout, E. Mixed Giardia duodenalis assemblage A and E infections in calves. Int. J. Parasitol. 2008, 38, 259–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trout, J.M.; Santín, M.; Greiner, E.; Fayer, R. Prevalence of Giardia duodenalis genotypes in pre-weaned dairy calves. Vet. Parasitol. 2004, 124, 179–186. [Google Scholar] [CrossRef]
- Geurden, T.; Somers, R.; Thanh, N.; Vien, L.; Nga, V.; Giang, H.; Dorny, P.; Giao, H.; Vercruysse, J. Parasitic infections in dairy cattle around Hanoi, northern Vietnam. Vet. Parasitol. 2008, 153, 384–388. [Google Scholar] [CrossRef]
- Li, S.; Zou, Y.; Zhang, X.-L.; Wang, P.; Chen, X.-Q.; Zhu, X.-Q. Prevalence and Multilocus Genotyping of Giardia lamblia in Cattle in Jiangxi Province, China: Novel Assemblage E Subtypes Identified. Korean J. Parasitol. 2020, 58, 681. [Google Scholar] [CrossRef]
- Naguib, D.; El-Gohary, A.H.; Mohamed, A.A.; Roellig, D.M.; Arafat, N.; Xiao, L. Age patterns of Cryptosporidium species and Giardia duodenalis in dairy calves in Egypt. Parasitol. Int. 2018, 67, 736–741. [Google Scholar] [CrossRef]
- Inpankaew, T.; Jiyipong, T.; Thadtapong, N.; Kengradomkij, C.; Pinyopanuwat, N.; Chimnoi, W.; Jittapalapong, S. Prevalence and genotype of Giardia duodenalis in dairy cattle from northern and northeastern part of Thailand. Acta Parasitol. 2015, 60, 459–461. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhao, G.; Chen, G.; Jian, F.; Zhang, S.; Feng, C.; Wang, R.; Zhu, J.; Dong, H.; Hua, J. Multilocus genotyping of Giardia duodenalis in dairy cattle in Henan, China. PLoS ONE 2014, 9, e100453. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.X.; Tan, Q.D.; Zhao, G.H.; Ma, J.G.; Zheng, W.B.; Ni, X.T.; Zhao, Q.; Zhou, D.H.; Zhu, X.Q. Prevalence, risk factors and multilocus genotyping of Giardia intestinalis in dairy cattle, Northwest China. J. Eukaryot. Microbiol. 2016, 63, 498–504. [Google Scholar] [CrossRef] [PubMed]
- Windeyer, M.; Leslie, K.; Godden, S.M.; Hodgins, D.; Lissemore, K.; LeBlanc, S. Factors associated with morbidity, mortality, and growth of dairy heifer calves up to 3 months of age. Prev. Vet. Med. 2014, 113, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Kwak, D.; Seo, M.-G. Genetic Analysis of zoonotic gastrointestinal Protozoa and Microsporidia in shelter cats in South Korea. Pathogens 2020, 9, 894. [Google Scholar] [CrossRef] [PubMed]
- Barigye, R.; Dyer, N.W.; Newell, T.K.; Khaitsa, M.L.; Trout, J.M.; Santin, M.; Fayer, R. Molecular and immunohistochemical detection of assemblage E, Giardia duodenalis in scouring North Dakota calves. Vet. Parasitol. 2008, 157, 196–202. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Muñoz, M.T.; Cámara-Badenes, C.; del Carmen Martínez-Herrero, M.; Dea-Ayuela, M.A.; Pérez-Gracia, M.T.; Fernández-Barredo, S.; Santín, M.; Fayer, R. Multilocus genotyping of Giardia duodenalis in lambs from Spain reveals a high heterogeneity. Res. Vet. Sci. 2012, 93, 836–842. [Google Scholar] [CrossRef]
- Cacciò, S.M.; Ryan, U. Molecular epidemiology of giardiasis. Mol. Biochem. Parasitol. 2008, 160, 75–80. [Google Scholar] [CrossRef]
- Mendonça, C.; Almeida, A.; Castro, A.; de Lurdes Delgado, M.; Soares, S.; da Costa, J.M.C.; Canada, N. Molecular characterization of Cryptosporidium and Giardia isolates from cattle from Portugal. Vet. Parasitol. 2007, 147, 47–50. [Google Scholar] [CrossRef]
- Abeywardena, H.; Jex, A.R.; Firestone, S.M.; McPhee, S.; Driessen, N.; Koehler, A.V.; Haydon, S.R.; von Samson-Himmelstjerna, G.; Stevens, M.A.; Gasser, R.B. Assessing calves as carriers of Cryptosporidium and Giardia with zoonotic potential on dairy and beef farms within a water catchment area by mutation scanning. Electrophoresis 2013, 34, 2259–2267. [Google Scholar] [CrossRef]
Figure 1.
A map of the Republic of Korea showing the location of the sampling sites from where fecal samples were collected from Korean native calves. GJ: Gimje (n = 624); JE: Jeongeup (n = 97); YJ: Youngju (n = 19); IS: Iksan (n = 14); GJ: Gongju (n = 13); HS: Hongseong (n = 7); HC: Hapcheon (n = 6); JJ: Jeonju (n = 3); AD: Andong (n = 3); WJ: Wanju (n = 2); JA: Jinan (n = 2); YC: Yecheon (n = 2).
Figure 1.
A map of the Republic of Korea showing the location of the sampling sites from where fecal samples were collected from Korean native calves. GJ: Gimje (n = 624); JE: Jeongeup (n = 97); YJ: Youngju (n = 19); IS: Iksan (n = 14); GJ: Gongju (n = 13); HS: Hongseong (n = 7); HC: Hapcheon (n = 6); JJ: Jeonju (n = 3); AD: Andong (n = 3); WJ: Wanju (n = 2); JA: Jinan (n = 2); YC: Yecheon (n = 2).
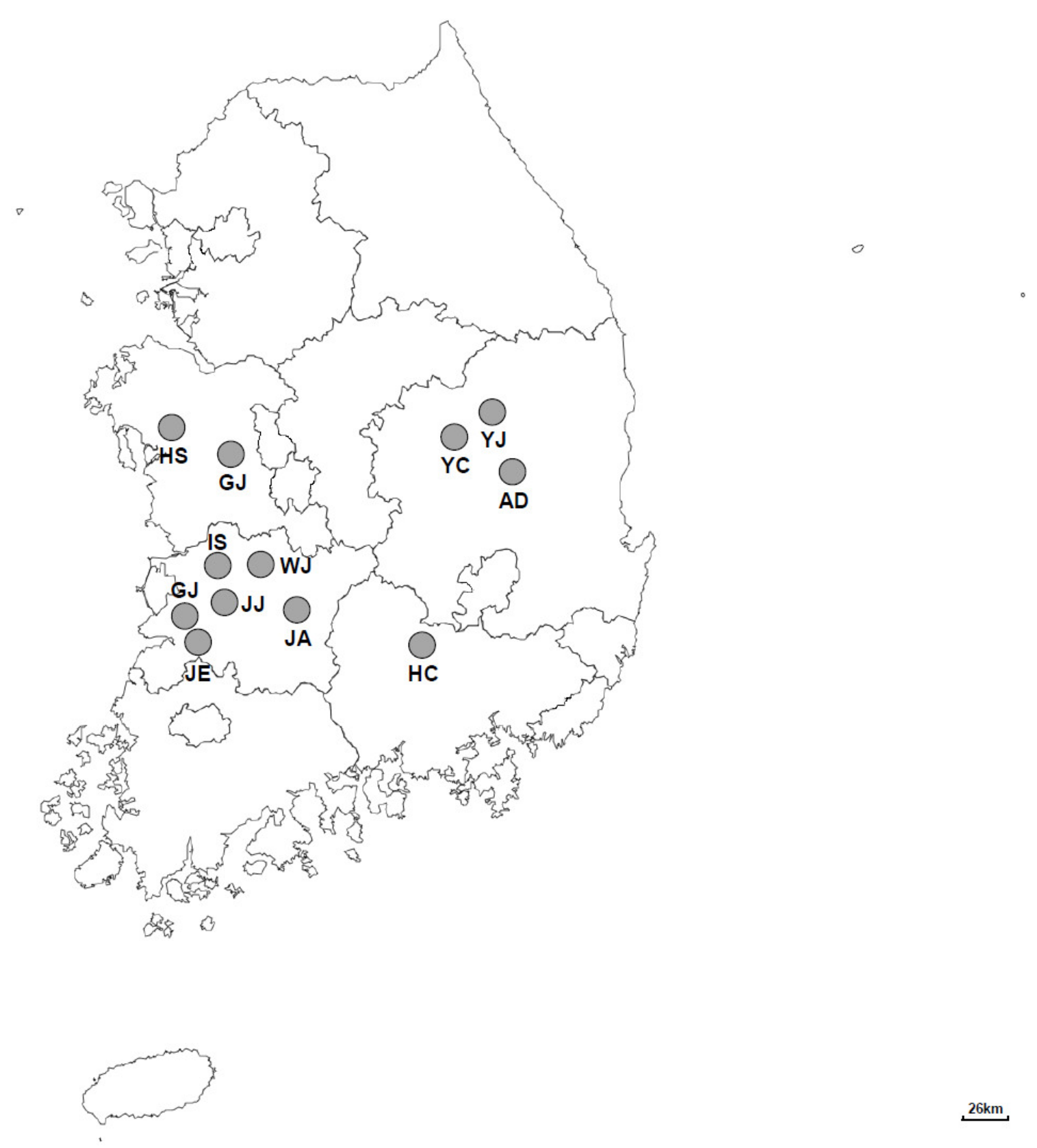
Figure 2.
Venn diagram of Giardia-duodenalis-positive samples (n = 44). Numbers of fecal samples that were PCR positive for ssu rRNA gene and/or bg gene and/or gdh gene are provided.
Figure 2.
Venn diagram of Giardia-duodenalis-positive samples (n = 44). Numbers of fecal samples that were PCR positive for ssu rRNA gene and/or bg gene and/or gdh gene are provided.
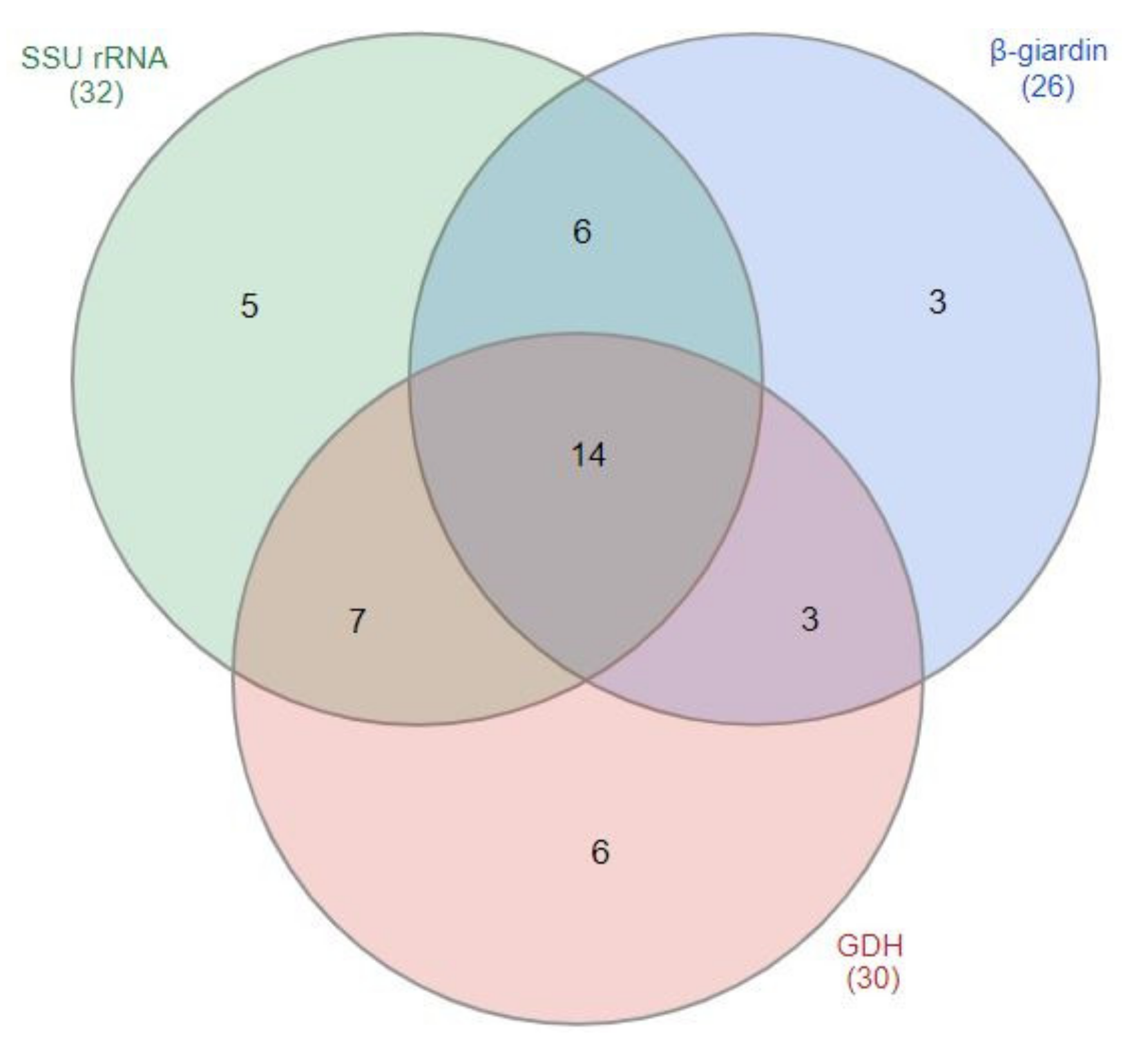
Figure 3.
Phylogenetic tree of G. duodenalis based on SSU rRNA gene sequences. The neighbor-joining method was used to construct the tree; the red circles indicate the sequences detected in this study. The GenBank accession numbers are shown in parentheses, and the G. duodenalis assemblages are indicated with one-sided square brackets. Tree reliability was tested by running 1000 bootstrap replicates.
Figure 3.
Phylogenetic tree of G. duodenalis based on SSU rRNA gene sequences. The neighbor-joining method was used to construct the tree; the red circles indicate the sequences detected in this study. The GenBank accession numbers are shown in parentheses, and the G. duodenalis assemblages are indicated with one-sided square brackets. Tree reliability was tested by running 1000 bootstrap replicates.
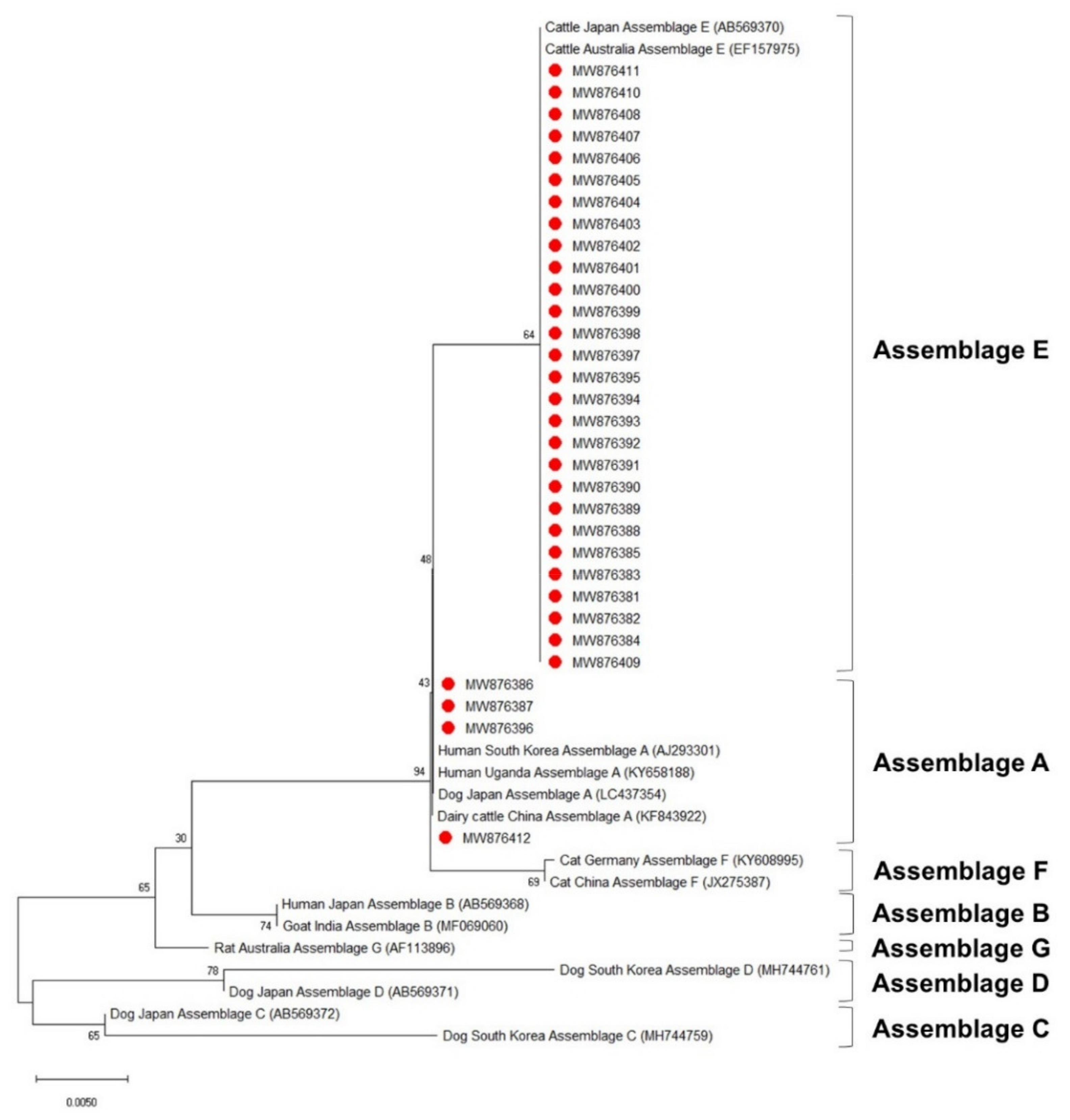
Figure 4.
Phylogenetic tree of G. duodenalis based on β-giardin gene sequences. The neighbor-joining method was used to construct the tree; the blue circles indicate the sequences detected in this study. The GenBank accession numbers are shown in parentheses, and the G. duodenalis assemblages are indicated with one-sided square brackets. Tree reliability was tested by running 1000 bootstrap replicates.
Figure 4.
Phylogenetic tree of G. duodenalis based on β-giardin gene sequences. The neighbor-joining method was used to construct the tree; the blue circles indicate the sequences detected in this study. The GenBank accession numbers are shown in parentheses, and the G. duodenalis assemblages are indicated with one-sided square brackets. Tree reliability was tested by running 1000 bootstrap replicates.
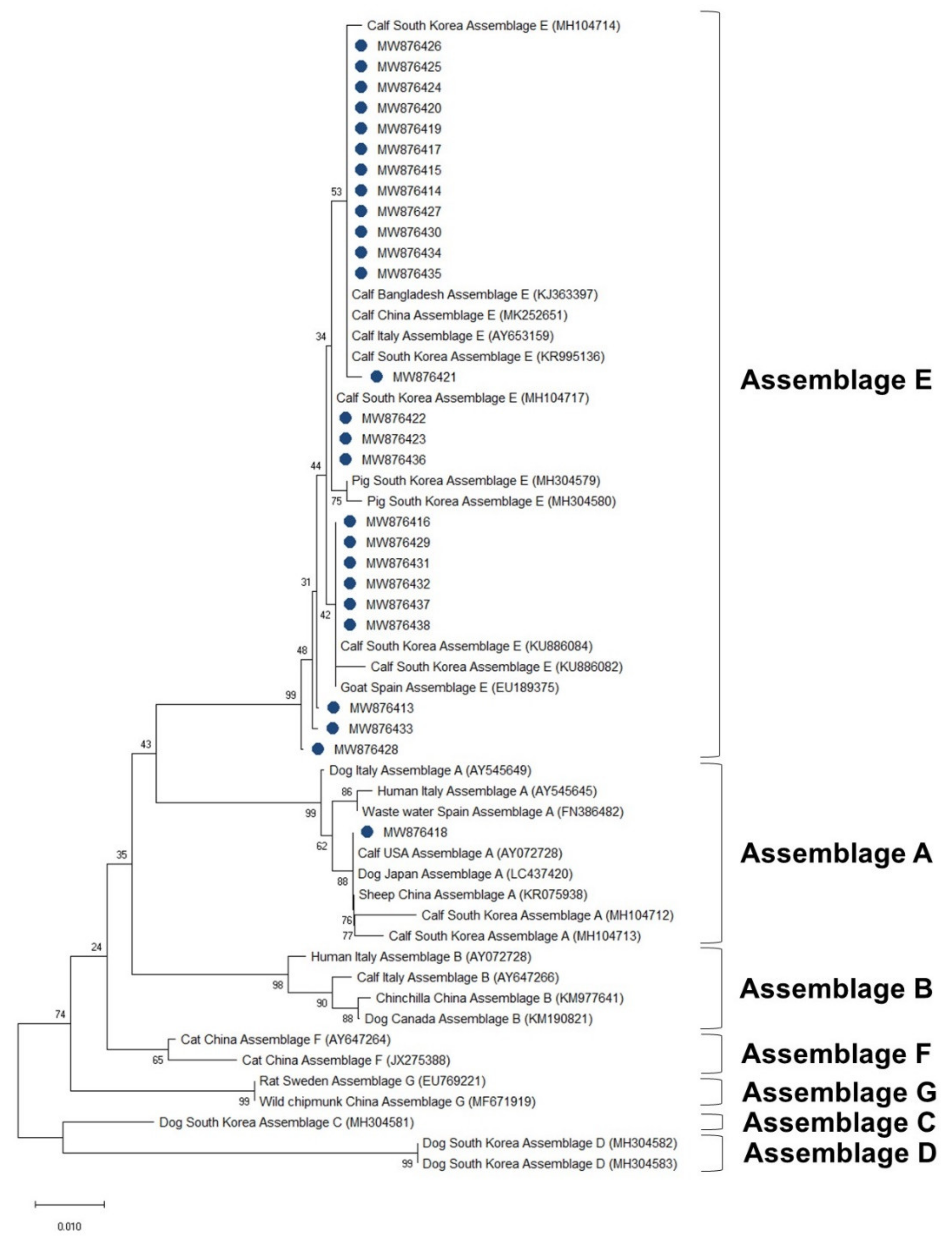
Figure 5.
Phylogenetic tree of G. duodenalis based on GDH gene sequences. The neighbor-joining method was used to construct the tree; the green circles indicate the sequences detected in this study. The GenBank accession numbers are shown in parentheses, and the G. duodenalis assemblages are indicated with one-sided square brackets. Tree reliability was tested by running 1000 bootstrap replicates.
Figure 5.
Phylogenetic tree of G. duodenalis based on GDH gene sequences. The neighbor-joining method was used to construct the tree; the green circles indicate the sequences detected in this study. The GenBank accession numbers are shown in parentheses, and the G. duodenalis assemblages are indicated with one-sided square brackets. Tree reliability was tested by running 1000 bootstrap replicates.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Detection of Giardia duodenalis by PCR in the feces of 792 Korean native calves.
| Classification | No. Tested | PCR | ||||
|---|---|---|---|---|---|---|
| No. of Positive Samples (%) | 95% CI * | OR * | p-Value | |||
| Constant | - | - | - | - | 0.06 | <0.001 |
| Age | ≤2 weeks ** | 462 | 7 (1.5) | 0.4–2.6 | 0.084 | 0.006 |
| 3–4 weeks | 138 | 9 (6.5) | 2.4–10.6 | 0.471 | 0.399 | |
| 5–6 weeks | 107 | 15 (14.0) | 7.4–20.6 | 1.081 | 0.929 | |
| 7–8 weeks | 26 | 4 (15.4) | 1.5–29.3 | 1.045 | 0.965 | |
| 9–12 weeks | 46 | 7 (15.2) | 4.8–25.6 | 1.072 | 0.940 | |
| ≥12 weeks | 13 | 2 (15.4) | 0–35.0 | - | - | |
| Sex | Male | 402 | 23 (5.7) | 3.5–8.0 | <0.001 | 0.998 |
| Female | 373 | 21 (5.6) | 3.3–8.0 | 0.618 | 0.161 | |
| Unknown | 17 | - | - | - | - | |
| Season | Spring ** | 256 | 17 (6.7) | 3.6–9.7 | 6.446 | 0.018 |
| Summer | 320 | 19 (6.0) | 3.4–8.5 | 3.393 | 0.113 | |
| Autumn | 81 | 6 (7.4) | 1.7–13.1 | 5.016 | 0.062 | |
| Winter | 135 | 2 (1.5) | 0–3.5 | - | - | |
| Fecal type | Normal | 349 | 8 (2.3) | 0.7–3.9 | - | - |
| Pasty | 137 | 4 (2.9) | 0.1–5.7 | 1.332 | 0.654 | |
| Watery ** | 234 | 23 (9.8) | 6.0–13.6 | 4.319 | 0.001 | |
| Hemorrhagic | 72 | 9 (12.5) | 4.9–20.1 | 2.123 | 0.169 | |
| Total | 792 | 44 (5.6) | 4.0–7.2 | - | - | |
* OR: Odds ratio, CI: Confidence interval. ** p < 0.05.
Table 2.
Nucleotide variations among the 26 β-giardin sequences detected from the feces of 792 calves.
Table 2.
Nucleotide variations among the 26 β-giardin sequences detected from the feces of 792 calves.
| Accession No. 1 | Nucleotides at Variable Positions | |||
|---|---|---|---|---|
| 45 | 150 | 252 | 393 | |
| MW876414, 15, 17, 19, 20, 24–27, 30, 34, 35 | T | A | T | T |
| MW876416, 29, 31, 32, 37, 38 | C | A | C | T |
| MW876422, 23, 36 | T | A | C | T |
| MW876413 | T | A | C | C |
| MW876421 | T | G | T | T |
| MW876428 | T | G | C | C |
| MW876433 | C | A | C | C |
1 MW876418 was marginally different from the other 25 β-giardin sequences.
Table 3.
Nucleotide variations among the 30 GDH sequences detected from the feces of 792 calves.
| Accession No. 1 | Nucleotides at Variable Positions | |||
|---|---|---|---|---|
| 258 | 306 | 342 | 387 | |
| MW876439–42, 45, 47–50, 52, 53, 56, 58, 59, 61, 63, 64 | C | T | A | C |
| MW876443, 44, 46, 51, 54, 57, 62, 65 | C | C | A | C |
| MW876455 | A | T | A | C |
| MW876460 | C | C | G | C |
| MW876466 | C | C | A | T |
1 MW876467 was marginally different from the other 29 GDH sequences.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Oh, S.-I.; Jung, S.-H.; Lee, H.-K.; Choe, C.; Hur, T.-Y.; So, K.-M. Multilocus Genotyping of Giardia duodenalis Occurring in Korean Native Calves. Vet. Sci. 2021, 8, 118. https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci8070118
AMA Style
Oh S-I, Jung S-H, Lee H-K, Choe C, Hur T-Y, So K-M. Multilocus Genotyping of Giardia duodenalis Occurring in Korean Native Calves. Veterinary Sciences. 2021; 8(7):118. https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci8070118
Chicago/Turabian StyleOh, Sang-Ik, Suk-Han Jung, Han-Kyoung Lee, Changyong Choe, Tai-Young Hur, and Kyoung-Min So. 2021. "Multilocus Genotyping of Giardia duodenalis Occurring in Korean Native Calves" Veterinary Sciences 8, no. 7: 118. https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci8070118
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.