
Naturally Acquired Antibodies to Influenza A Virus in Fall-Migrating North American Mallards

 and
and 
Abstract
:1. Introduction
2. Materials and Methods
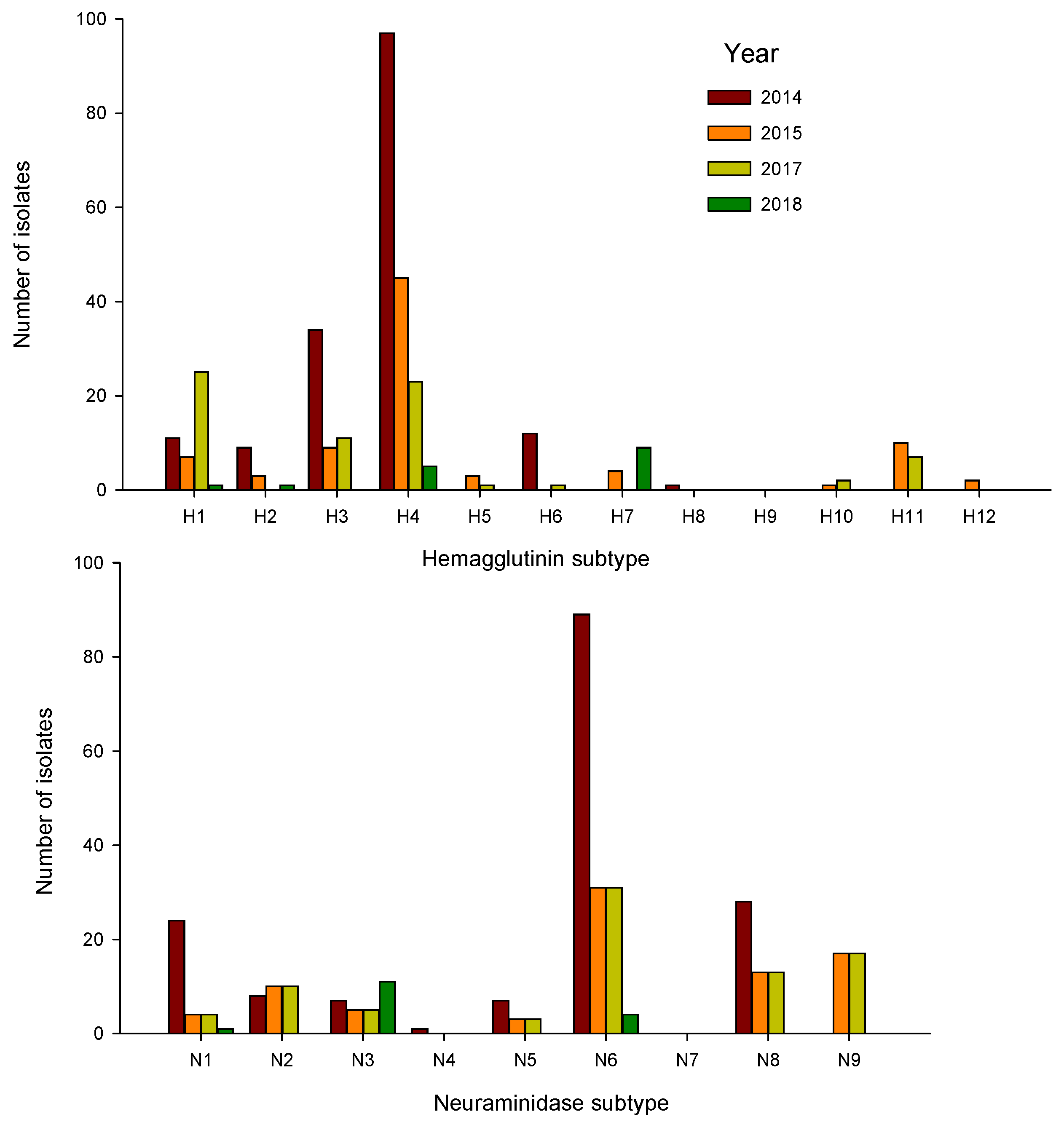
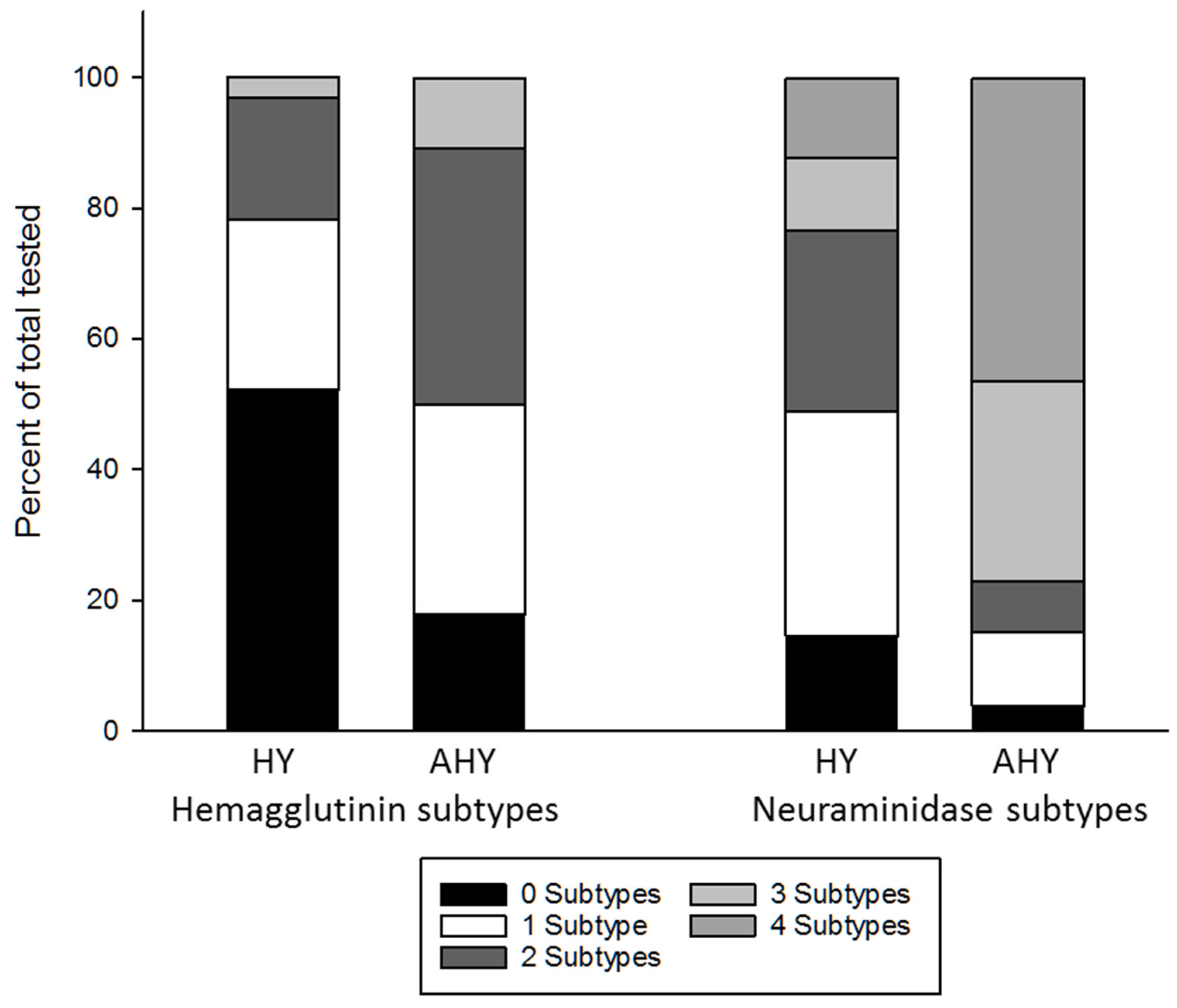
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| Year | Dates Sampled | Number Tested | Number IAV Positive | Percent Positive (95% Confidence Limits) | Subtypes Isolated (Subtype (Number)) |
|---|---|---|---|---|---|
| 2014 | 28 August 2014–13 September 2014 | 532 | 168 | 31.6% (0.28–0.36) | H1N1(7), H1N3(1), H1N5(1), H1N6(1), H1N8(1), H2N3(5), H2N5(3), H2N6(1), H3N1(5), H3N2(2), H3N5(1), H3N6(6), H3N8(19), H3Nx(1), H4Nx(1), H4N1(4), H4N2(6), H4N3(1), H4N5(1), H4N6(76), H4N8(9), H6N1(8), H6N5(1), H6N6(2), H6N8(1), H8N4(1), HxN5(1), HxN6(1), mixed a (1) |
| 2015 | 11 September 2015–23 September 2015 | 586 | 87 | 14.9% (0.12–0.18) | H1N1(2), H1N2(1), H1N5(1), H1N9(3), H2N3(1), H2N5(1), H2N9(1), H3N6(1), H3N8(7), H3N9(1), H4N1(1), H4N2(7), H4N6(29), H4N8(6), H4N9(1), H5N2(1), H5N9(2), H7Nx(2), H7N3(3), H10N9(1), H11N1(1), H11N2(1), H11N9(8), H12N5(1), HxN6(1), HxN9(2), mixed(1) |
| 2017 | 6 September 2017–21 September 2017 | 794 | 70 | 8.8% (0.07–0.11) | H1N1(24), H1N8(1), H3N1(1), H3N6(7), H3N8(3), H4N1(1), H4N2(2), H4N6(15), H4N8(5), H5N7(1), H6N1(1), H10N7(1), H10N9(1), H11N1(1), H11N8(1), H11N9(4), mixed(1) |
| 2018 | 30 August 2018–20 September 2018 | 273 | 25 | 9.2% (0.06–0.13) | H1N1(1), H2N3(1), H4N3(1), H4N6(4), H4Nx(2), H7N3(9), HxN6(1), HxNx b (6) |
References
- Diskin, E.R.; Friedman, K.; Krauss, S.; Nolting, J.M.; Poulson, R.L.; Slemons, R.D.; Stallknecht, D.E.; Webster, R.G.; Bowman, A.S. Subtype Diversity of Influenza A Virus in North American Waterfowl: A Multidecade Study. J. Virol. 2020, 94, e02022-19. [Google Scholar] [CrossRef]
- Hinshaw, V.S.; Webster, R.G.; Turner, B. The perpetuation of orthomyxoviruses and paramyxoviruses in Canadian waterfowl. Can. J. Microbiol. 1980, 26, 622–629. [Google Scholar] [CrossRef] [PubMed]
- van Dijk, J.G.; Hoye, B.J.; Verhagen, J.H.; Nolet, B.A.; Fouchier, R.A.; Klaassen, M. Juveniles and migrants as drivers for seasonal ep-izootics of avian influenza virus. J. Anim. Ecol. 2014, 83, 266–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinshaw, V.S.; Wood, J.M.; Webster, R.G.; Deibel, R.; Turner, B. Circulation of influenza viruses and paramyxoviruses in water-fowl originating from two different areas of North America. Bull. WHO 1985, 63, 711–719. [Google Scholar]
- Ramey, A.M.; Poulson, R.L.; González-Reiche, A.S.; Wilcox, B.R.; Walther, P.; Link, P.; Carter, D.L.; Newsome, G.M.; Müller, M.L.; Berghaus, R.D.; et al. Evidence for Seasonal Patterns in the Relative Abundance of Avian Influenza Virus Subtypes in Blue-Winged Teal (Anas discors). J. Wildl. Dis. 2014, 50, 916–922. [Google Scholar] [CrossRef]
- Carter, D.; Link, P.; Walther, P.; Ramey, A.; Stallknecht, D.; Poulson, R. Influenza A Prevalence and Subtype Diversity in Migrating Teal Sampled Along the United States Gulf Coast. Avian Dis. 2018, 63, 165–171. [Google Scholar] [CrossRef]
- Brown, J.D.; Luttrell, M.P.; Berghaus, R.D.; Kistler, W.; Keeler, S.P.; Howey, A.; Wilcox, B.; Hall, J.; Niles, L.; Dey, A.; et al. Prevalence of antibodies to type a influenza virus in wild avian species using two serologic assays. J. Wildl. Dis. 2010, 46, 896–911. [Google Scholar] [CrossRef] [PubMed]
- Hill, S.C.; Manvell, R.J.; Schulenburg, B.; Shell, W.; Wikramaratna, P.S.; Perrins, C.; Sheldon, B.; Brown, I.H.; Pybus, O.G. Antibody responses to avian influenza viruses in wild birds broaden with age. Proc. R. Soc. B Boil. Sci. 2016, 283, 20162159. [Google Scholar] [CrossRef]
- Ramey, A.M.; Uher-Koch, B.D.; Reeves, A.B.; Schmutz, J.A.; Poulson, R.L.; Stallknecht, D.E. Emperor geese (Anser canagicus) are exposed to a diversity of influenza A viruses, are infected during the non-breeding period and contribute to intercontinental viral dispersal. Transbound. Emerg. Dis. 2019, 66, 1958–1970. [Google Scholar] [CrossRef]
- Wohlbold, T.J.; Krammer, F. In the Shadow of Hemagglutinin: A Growing Interest in Influenza Viral Neuraminidase and Its Role as a Vaccine Antigen. Viruses 2014, 6, 2465–2494. [Google Scholar] [CrossRef]
- Chen, Y.-Q.; Wohlbold, T.J.; Zheng, N.-Y.; Huang, M.; Huang, Y.; Neu, K.E.; Lee, J.; Wan, H.; Rojas, K.T.; Kirkpatrick, E.; et al. Influenza Infection in Humans Induces Broadly Cross-Reactive and Protective Neuraminidase-Reactive Antibodies. Cell 2018, 173, 417–429.e10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hollander, L.P.; Fojtik, A.; Kienzle-Dean, C.; Davis-Fields, N.; Poulson, R.L.; Davis, B.; Mowry, C.; Stallknecht, D.E. Prevalence of In-fluenza A Viruses in Ducks Sampled in Northwestern Minnesota and Evidence for Predominance of H3N8 and H4N6 Sub-types in Mallards, 2007–2016. Avian Dis. 2019, 63, 126–130. [Google Scholar] [CrossRef] [PubMed]
- Wagner, R.; Matrosovich, M.; Klenk, H.-D. Functional balance between haemagglutinin and neuraminidase in influenza virus infections. Rev. Med Virol. 2002, 12, 159–166. [Google Scholar] [CrossRef]
- Verhagen, J.H.; Fouchier, R.A.; Lewis, N. Highly Pathogenic Avian Influenza Viruses at the Wild–Domestic Bird Interface in Europe: Future Directions for Research and Surveillance. Viruses 2021, 13, 212. [Google Scholar] [CrossRef]
- Pasick, J.; Berhane, Y.; Joseph, T.; Bowes, V.; Hisanaga, T.; Handel, K.; Alexandersen, S. Reassortant Highly Pathogenic Influenza A H5N2 Virus Containing Gene Segments Related to Eurasian H5N8 in British Columbia, Canada, 2014. Sci. Rep. 2015, 5, 9484. [Google Scholar] [CrossRef] [Green Version]
- Saito, T.; Tanikawa, T.; Uchida, Y.; Takemae, N.; Kanehira, K.; Tsunekuni, R. Intracontinental and intercontinental dissemination of Asian H5 highly pathogenic avian influenza virus (clade 2.3.4.4) in the winter of 2014-2015. Rev. Med Virol. 2015, 25, 388–405. [Google Scholar] [CrossRef]
- Lee, D.-H.; Bahl, J.; Torchetti, M.K.; Killian, M.L.; Ip, H.; Deliberto, T.J.; Swayne, D.E. Highly Pathogenic Avian Influenza Viruses and Generation of Novel Reassortants, United States, 2014–2015. Emerg. Infect. Dis. 2016, 22, 1283–1285. [Google Scholar] [CrossRef]
- Lee, D.-H.; Torchetti, M.K.; Killian, M.L.; Deliberto, T.J.; Swayne, D. Reoccurrence of Avian Influenza A(H5N2) Virus Clade 2.3.4.4 in Wild Birds, Alaska, USA, 2016. Emerg. Infect. Dis. 2017, 23, 365–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krauss, S.; Stallknecht, D.E.; Slemons, R.D.; Bowman, A.S.; Poulson, R.L.; Nolting, J.M.; Knowles, J.P.; Webster, R.G. The enigma of the apparent disappearance of Eurasian highly pathogenic H5 clade 2.3.4.4 influenza A viruses in North American waterfowl. Proc. Natl. Acad. Sci. USA 2016, 113, 9033–9038. [Google Scholar] [CrossRef] [Green Version]
- Costa, T.P.; Brown, J.D.; Howerth, E.W.; Stallknecht, D.E.; Swayne, D.E. Homo- and Heterosubtypic Low Pathogenic Avian Influenza Exposure on H5N1 Highly Pathogenic Avian Influenza Virus Infection in Wood Ducks (Aix sponsa). PLoS ONE 2011, 6, e15987. [Google Scholar] [CrossRef] [Green Version]
- Berhane, Y.; Embury-Hyatt, C.; Leith, M.; Kehler, H.; Suderman, M.; Pasick, J. Pre-exposing Canada Geese (Branta canadensis) to a low-pathogenic H1N1 avian influenza virus protects them against H5N1 HPAI virus challenge. J. Wildl. Dis. 2014, 50, 84–97. [Google Scholar] [CrossRef] [PubMed]
- Latorre-Margalef, N.; Brown, J.D.; Fojtik, A.; Poulson, R.L.; Carter, D.; França, M.; Stallknecht, D.E. Competition between influenza A virus subtypes through heterosubtypic immunity modulates re-infection and antibody dynamics in the mallard duck. PLoS Pathog. 2017, 13, e1006419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segovia, K.M.; Stallknecht, D.E.; Kapczynski, D.R.; Stabler, L.; Berghaus, R.; Fotjik, A.; Latorre-Margalef, N.; França, M.S. Adaptive Heterosubtypic Immunity to Low Pathogenic Avian Influenza Viruses in Experimentally Infected Mallards. PLoS ONE 2017, 12, e0170335. [Google Scholar] [CrossRef] [PubMed]
- Segovia, K.M.; França, M.S.; Leyson, C.L.; Kapczynski, D.R.; Chrzastek, K.; Bahnson, C.S.; Stallknecht, D.E. Heterosubtypic immunity increases infectious dose required to infect Mallard ducks with Influenza A virus. PLoS ONE 2018, 13, e0196394. [Google Scholar] [CrossRef] [Green Version]
- Hanson, B.A.; Stallknecht, D.E.; Swayne, D.; Lewis, L.A.; Senne, D.A. Avian Influenza Viruses in Minnesota Ducks During 1998–2000. Avian Dis. 2003, 47, 867–871. [Google Scholar] [CrossRef]
- Wilcox, B.R.; Knutsen, G.A.; Berdeen, J.; Goekjian, V.; Poulson, R.; Goyal, S.; Sreevatsan, S.; Cardona, C.; Berghaus, R.; Swayne, D.; et al. Influenza-A Viruses in Ducks in Northwestern Minnesota: Fine Scale Spatial and Temporal Variation in Prevalence and Subtype Diversity. PLoS ONE 2011, 6, e24010. [Google Scholar] [CrossRef]
- Stallknecht, D.E.; Shane, S.M.; Zwank, P.J.; Senne, D.A.; Kearney, M.T. Avian Influenza Viruses from Migratory and Resident Ducks of Coastal Louisiana. Avian Dis. 1990, 34, 398. [Google Scholar] [CrossRef]
- Spackman, E. Avian Influenza Virus Detection and Quantitation by Real-Time RT-PCR. Methods Mol. Biol. 2020, 2123, 137–148. [Google Scholar] [CrossRef]
- Lebarbenchon, C.; Poulson, R.; Shannon, K.; Slagter, J.; Slusher, M.J.; Wilcox, B.R.; Berdeen, J.; Knutsen, G.A.; Cardona, C.J.; Stallknecht, D.E. Isolation of Influenza A Viruses from Wild Ducks and Feathers in Minnesota (2010–2011). Avian Dis. 2013, 57, 677–680. [Google Scholar] [CrossRef]
- Shriner, S.A.; VanDalen, K.K.; Root, J.J.; Sullivan, H.J. Evaluation and optimization of a commercial blocking ELISA for detecting antibodies to influenza A virus for research and surveillance of mallards. J. Virol. Methods 2016, 228, 130–134. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.D.; Stallknecht, D.E.; Berghaus, R.D.; Luttrell, M.P.; Velek, K.; Kistler, W.; Costa, T.; Yabsley, M.J.; Swayne, D. Evaluation of a commercial blocking enzyme-linked immunosorbent assay to detect avian influenza virus antibodies in multiple experi-mentally infected avian species. Clin. Vaccine Immunol. 2009, 16, 824–829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spivey, T.J.; Lindberg, M.S.; Meixell, B.W.; Smith, K.R.; Puryear, W.B.; Davis, K.R.; Runstadler, J.A.; Stallknecht, D.E.; Ramey, A.M. Maintenance of influenza A viruses and antibody response in mallards (Anas platyrhynchos) sampled during the non-breeding season in Alaska. PLoS ONE 2017, 12, e0183505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, J.; Couzens, L.; Eichelberger, M.C. Measuring Influenza Neuraminidase Inhibition Antibody Titers by Enzyme-linked Lectin Assay. J. Vis. Exp. 2016, 6, e54573. [Google Scholar] [CrossRef] [PubMed]
- Couzens, L.; Gao, J.; Westgeest, K.; Sandbulte, M.; Lugovtsev, V.; Fouchier, R.; Eichelberger, M. An optimized enzyme-linked lectin assay to measure influenza A virus neuraminidase inhibition antibody titers in human sera. J. Virol. Methods 2014, 210, 7–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, N.J.; Takekawa, J.Y.; Ackerman, J.T.; Hobson, K.A.; Herring, G.; Cardona, C.J.; Runstadler, J.A.; Boyce, W.M. Migration strategy affects avian influenza dynamics in mallards (Anas platyrhynchos). Mol. Ecol. 2012, 21, 5986–5999. [Google Scholar] [CrossRef] [PubMed]
- Hill, S.C.; Hansen, R.; Watson, S.; Coward, V.; Russell, C.; Cooper, J.; Essen, S.; Everest, H.; Parag, K.V.; Fiddaman, S.; et al. Comparative micro-epidemiology of pathogenic avian influenza virus outbreaks in a wild bird population. Philos. Trans. R. Soc. B Biol. Sci. 2019, 374, 20180259. [Google Scholar] [CrossRef] [Green Version]


| Year | Age Class 1 | Number Tested | Number Positive | Percent Positive (95% Confidence Limits) |
|---|---|---|---|---|
| 2014 | HY | 89 | 67 | 75% (0.65–0.84) |
| AHY | 22 | 20 | 91% (0.71–0.99) | |
| 2015 | HY | 353 | 179 | 51% (0.45–0.56) |
| AHY | 82 | 76 | 93% (0.85–0.97) | |
| 2017 | HY | 92 | 70 | 76% (0.66–0.84) |
| AHY | 57 | 49 | 86% (0.74–0.94) | |
| 2018 | HY | 51 | 31 | 61% (0.46–0.74) |
| AHY | 41 | 37 | 90% (0.77–0.97) | |
| Total | HY | 585 | 347 | 59% (0.55–0.63) |
| AHY | 202 | 182 | 90% (0.85–0.93) |
| Neuraminidase Subtype | Hemagglutinin Subtype | Total | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| H1 | H2 | H3 | H4 | H5 | H6 | H7 | H8 | H9 | H10 | H11 | H12 | ||
| N1 | 34 | 6 | 6 | 9 | 2 | 57 | |||||||
| N2 | 1 | 2 | 15 | 1 | 1 | 20 | |||||||
| N3 | 1 | 7 | 2 | 12 | 22 | ||||||||
| N4 | 1 | 1 | |||||||||||
| N5 | 2 | 4 | 1 | 1 | 1 | 1 | 10 | ||||||
| N6 | 1 | 1 | 14 | 124 | 2 | 142 | |||||||
| N7 | 1 | 1 | 2 | ||||||||||
| N8 | 2 | 29 | 20 | 1 | 1 | 53 | |||||||
| N9 | 3 | 1 | 1 | 1 | 2 | 2 | 12 | 22 | |||||
| Total | 44 | 13 | 53 | 169 | 4 | 13 | 12 | 1 | 0 | 3 | 16 | 1 | 329 |
| Year | Age Class 1 | n | Microneutralization (Number Positive (% Positive)) | ELLA (Number Positive (% Positive)) | |||||
|---|---|---|---|---|---|---|---|---|---|
| H3 | H4 | H5 | N1 | N2 | N6 | N8 | |||
| 2014 2 | HY | 21 | 2 (10%) | 4 (20%) | 1 (5%) | 3 (16%) | 3 (14%) | 14 (67%) | 4 (20%) |
| AHY | 5 | 0 | 2 (40%) | 0 | 0 | 1 (20%) | 4 (80%) | 1 (20%) | |
| 2015 | HY | 26 | 9 (35%) | 9 (35%) | 10 (38%) | 4 (15%) | 8 (31%) | 19 (73%) | 12 (46%) |
| AHY | 5 | 2 (40%) | 1 (20%) | 5 (100%) | 2 (40%) | 5 (100%) | 3 (60%) | 5 (100%) | |
| 2017 | HY | 24 | 5 (21%) | 9 (38%) | 6 (25%) | 6 (25%) | 7 (29%) | 19 (79%) | 12 (50%) |
| AHY | 4 | 1 (25%) | 1 (25%) | 3 (75%) | 3 (75%) | 4 (100%) | 3 (75%) | 3 (75%) | |
| 2018 | HY | 21 | 4 (19%) | 6 (29%) | 5 (24%) | 5 (24%) | 10 (48%) | 20 (95%) | 10 (48%) |
| AHY | 14 | 10 (71%) | 6 (43%) | 9 (64%) | 9 (64%) | 12 (86%) | 13 (93%) | 13 (93%) | |
| Total | HY | 92 | 20 (22%) | 28 (30%) | 22 (24%) | 18 (20%) | 28 (30%) | 72 (78%) | 38 (41%) |
| AHY | 28 | 13 (46%) | 10 (36%) | 17 (61%) | 14 (54%) | 22 (79%) | 23 (83%) | 22 (79%) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stallknecht, D.E.; Fojtik, A.; Carter, D.L.; Crum-Bradley, J.A.; Perez, D.R.; Poulson, R.L. Naturally Acquired Antibodies to Influenza A Virus in Fall-Migrating North American Mallards. Vet. Sci. 2022, 9, 214. https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci9050214
Stallknecht DE, Fojtik A, Carter DL, Crum-Bradley JA, Perez DR, Poulson RL. Naturally Acquired Antibodies to Influenza A Virus in Fall-Migrating North American Mallards. Veterinary Sciences. 2022; 9(5):214. https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci9050214
Chicago/Turabian StyleStallknecht, David E., Alinde Fojtik, Deborah L. Carter, Jo Anne Crum-Bradley, Daniel R. Perez, and Rebecca L. Poulson. 2022. "Naturally Acquired Antibodies to Influenza A Virus in Fall-Migrating North American Mallards" Veterinary Sciences 9, no. 5: 214. https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci9050214