
Changes in Neuropeptide Prohormone Genes among Cetartiodactyla Livestock and Wild Species Associated with Evolution and Domestication

Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Prohormone Identification across Species
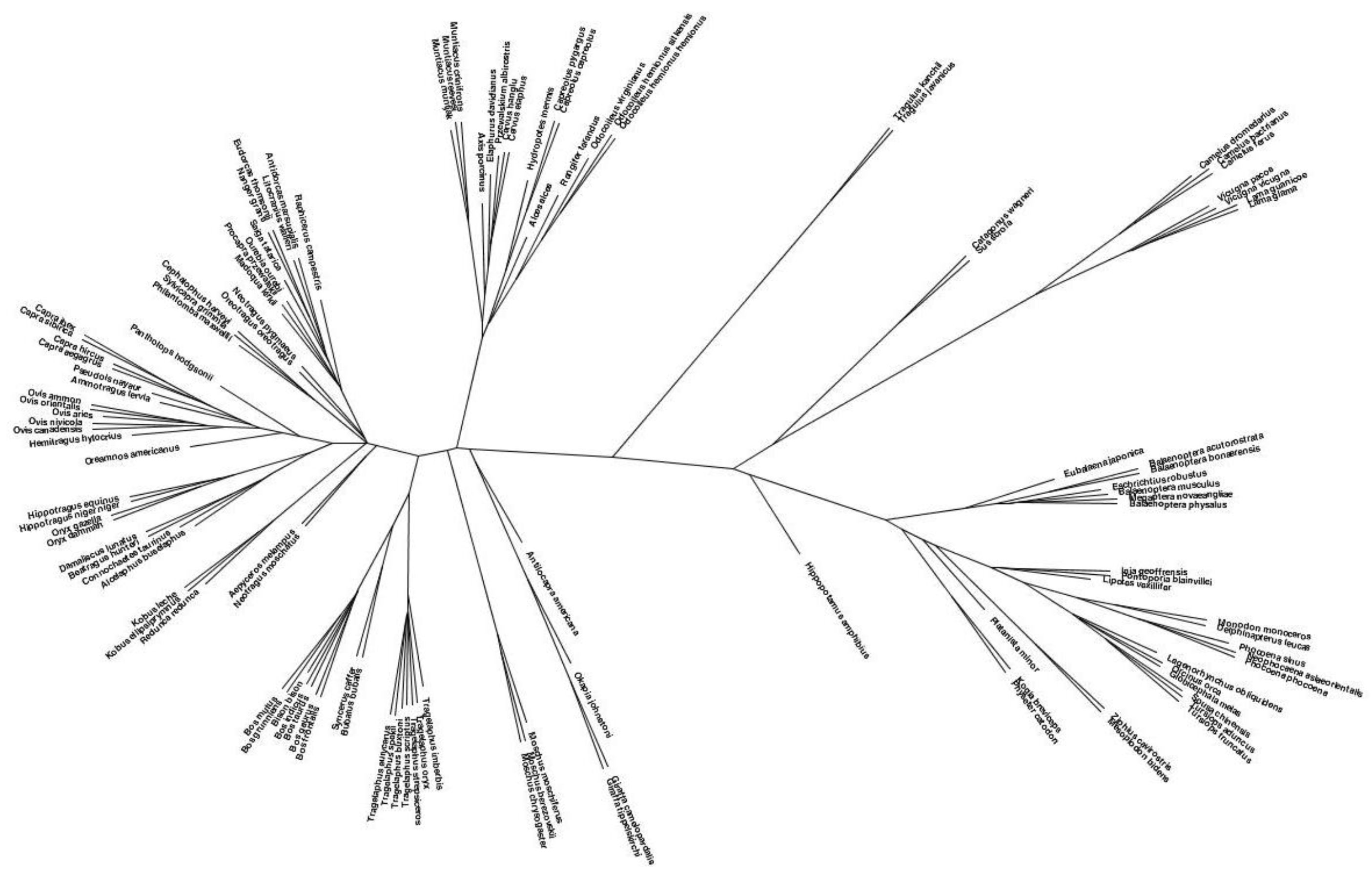
3.2. Species Tree Derived from Prohormone Sequences
3.2.1. Species Tree
3.2.2. Correlation of InterSpecies Distances Based on Individual and All Neuropeptide Prohormone Genes
3.3. Evolutionary Model
4. Discussion
4.1. Prohormone Complement
4.2. Evolutionary Model
4.3. Domestication
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zeller, U.; Göttert, T. The relations between evolution and domestication reconsidered—Implications for systematics, ecology, and nature conservation. Glob. Ecol. Conserv. 2019, 20, e00756. [Google Scholar] [CrossRef]
- Zeder, M.A. Out of the Fertile Crescent: The dispersal of domestic livestock through Europe and Africa. In Human Dispersal and Species Movement: From Prehistory to the Present; Petraglia, M., Boivin, N., Crassard, R., Eds.; Cambridge University Press: Cambridge, UK, 2017; pp. 261–303. [Google Scholar]
- McHugo, G.P.; Dover, M.J.; MacHugh, D.E. Unlocking the origins and biology of domestic animals using ancient DNA and paleogenomics. BMC Biol. 2019, 17, 98. [Google Scholar] [CrossRef] [Green Version]
- Frantz, L.A.F.; Bradley, D.G.; Larson, G.; Orlando, L. Animal domestication in the era of ancient genomics. Nat. Rev. Genet. 2020, 21, 449–460. [Google Scholar] [CrossRef]
- Lott, R.; Williamson, C. Cetaceans in Captivity. In Marine Mammal Welfare: Human Induced Change in the Marine Environment and its Impacts on Marine Mammal Welfare; Butterworth, A., Ed.; Springer International Publishing: Cham, Switzerland, 2017; pp. 161–181. [Google Scholar]
- Fan, R.; Gu, Z.; Guang, X.; Marin, J.C.; Varas, V.; Gonzalez, B.A.; Wheeler, J.C.; Hu, Y.; Li, E.; Sun, X.; et al. Genomic analysis of the domestication and post-Spanish conquest evolution of the llama and alpaca. Genome Biol. 2020, 21, 159. [Google Scholar] [CrossRef] [PubMed]
- Roed, K.H.; Flagstad, O.; Nieminen, M.; Holand, O.; Dwyer, M.J.; Rov, N.; Vila, C. Genetic analyses reveal independent domestication origins of Eurasian reindeer. Proc. Biol. Sci. 2008, 275, 1849–1855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herbeck, Y.E.; Gulevich, R.G. Neuropeptides as facilitators of domestication. Cell Tissue Res. 2019, 375, 295–307. [Google Scholar] [CrossRef] [PubMed]
- Irwin, D.M. Variation in the Evolution and Sequences of Proglucagon and the Receptors for Proglucagon-Derived Peptides in Mammals. Front. Endocrinol. 2021, 12, 700066. [Google Scholar] [CrossRef] [PubMed]
- Wolf Horrell, E.M.; Boulanger, M.C.; D’Orazio, J.A. Melanocortin 1 Receptor: Structure, Function, and Regulation. Front. Genet. 2016, 7, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Simoni Gouveia, J.J.; da Silva, M.V.; Paiva, S.R.; de Oliveira, S.M. Identification of selection signatures in livestock species. Genet. Mol. Biol. 2014, 37, 330–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dib, L.; San-Jose, L.M.; Ducrest, A.L.; Salamin, N.; Roulin, A. Selection on the Major Color Gene Melanocortin-1-Receptor Shaped the Evolution of the Melanocortin System Genes. Int. J. Mol. Sci. 2017, 18, 2618. [Google Scholar] [CrossRef] [Green Version]
- Von Heijne, G. The signal peptide. J. Membr. Biol. 1990, 115, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Southey, B.R.; Rodriguez-Zas, S.L.; Sweedler, J.V. Prediction of neuropeptide prohormone cleavages with application to RFamides. Peptides 2006, 27, 1087–1098. [Google Scholar] [CrossRef] [PubMed]
- Southey, B.R.; Sweedler, J.V.; Rodriguez-Zas, S.L. A python analytical pipeline to identify prohormone precursors and predict prohormone cleavage sites. Front. Neuroinform. 2008, 2, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Southey, B.R.; Romanova, E.V.; Rodriguez-Zas, S.L.; Sweedler, J.V. Bioinformatics for Prohormone and Neuropeptide Discovery. Methods Mol. Biol 2018, 1719, 71–96. [Google Scholar] [CrossRef] [PubMed]
- Southey, B.R.; Rodriguez-Zas, S.L.; Sweedler, J.V. Characterization of the prohormone complement in cattle using genomic libraries and cleavage prediction approaches. BMC Genom. 2009, 10, 228. [Google Scholar] [CrossRef] [Green Version]
- Porter, K.I.; Southey, B.R.; Sweedler, J.V.; Rodriguez-Zas, S.L. First survey and functional annotation of prohormone and convertase genes in the pig. BMC Genom. 2012, 13, 582. [Google Scholar] [CrossRef] [Green Version]
- Hu, C.K.; Southey, B.R.; Romanova, E.V.; Maruska, K.P.; Sweedler, J.V.; Fernald, R.D. Identification of prohormones and pituitary neuropeptides in the African cichlid, Astatotilapia burtoni. BMC Genom. 2016, 17, 660. [Google Scholar] [CrossRef] [Green Version]
- Giuffra, E.; Kijas, J.M.; Amarger, V.; Carlborg, O.; Jeon, J.T.; Andersson, L. The origin of the domestic pig: Independent domestication and subsequent introgression. Genetics 2000, 154, 1785–1791. [Google Scholar] [CrossRef]
- Verdugo, M.P.; Mullin, V.E.; Scheu, A.; Mattiangeli, V.; Daly, K.G.; Maisano Delser, P.; Hare, A.J.; Burger, J.; Collins, M.J.; Kehati, R.; et al. Ancient cattle genomics, origins, and rapid turnover in the Fertile Crescent. Science 2019, 365, 173–176. [Google Scholar] [CrossRef]
- Ciani, E.; Mastrangelo, S.; Da Silva, A.; Marroni, F.; Ferencakovic, M.; Ajmone-Marsan, P.; Baird, H.; Barbato, M.; Colli, L.; Delvento, C.; et al. On the origin of European sheep as revealed by the diversity of the Balkan breeds and by optimizing population-genetic analysis tools. Genet. Sel. Evol. 2020, 52, 25. [Google Scholar] [CrossRef]
- Sayers, E.W.; Beck, J.; Brister, J.R.; Bolton, E.E.; Canese, K.; Comeau, D.C.; Funk, K.; Ketter, A.; Kim, S.; Kimchi, A.; et al. Database resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2020, 48, D9–D16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The Bactrian Camels Genome Sequencing and Analysis Consortium; Jirimutu; Wang, Z.; Ding, G.; Chen, G.; Sun, Y.; Sun, Z.; Zhang, H.; Wang, L.; Hasi, S.; et al. Genome sequences of wild and domestic bactrian camels. Nat. Commun. 2012, 3, 1202. [Google Scholar] [CrossRef]
- Amare, A.; Hummon, A.B.; Southey, B.R.; Zimmerman, T.A.; Rodriguez-Zas, S.L.; Sweedler, J.V. Bridging neuropeptidomics and genomics with bioinformatics: Prediction of mammalian neuropeptide prohormone processing. J. Proteome Res. 2006, 5, 1162–1167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tegge, A.N.; Southey, B.R.; Sweedler, J.V.; Rodriguez-Zas, S.L. Comparative analysis of neuropeptide cleavage sites in human, mouse, rat, and cattle. Mamm. Genome 2008, 19, 106–120. [Google Scholar] [CrossRef] [PubMed]
- Alim, F.Z.D.; Romanova, E.V.; Tay, Y.L.; Rahman, A.; Chan, K.G.; Hong, K.W.; Rogers, M.; Southey, B.R.; Greenwood, M.P.; Mecawi, A.S.; et al. Seasonal adaptations of the hypothalamo-neurohypophyseal system of the dromedary camel. PLoS ONE 2019, 14, e0216679. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schaffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [Green Version]
- Birney, E.; Clamp, M.; Durbin, R. GeneWise and Genomewise. Genome Res. 2004, 14, 988–995. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.K.; Wootton, J.C.; Altschul, S.F. The compositional adjustment of amino acid substitution matrices. Proc. Natl. Acad. Sci. USA 2003, 100, 15688–15693. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Gil, M.; Zanetti, M.S.; Zoller, S.; Anisimova, M. CodonPhyML: Fast maximum likelihood phylogeny estimation under codon substitution models. Mol. Biol. Evol. 2013, 30, 1270–1280. [Google Scholar] [CrossRef] [Green Version]
- Mirarab, S.; Reaz, R.; Bayzid, M.S.; Zimmermann, T.; Swenson, M.S.; Warnow, T. ASTRAL: Genome-scale coalescent-based species tree estimation. Bioinformatics 2014, 30, i541–i548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirarab, S.; Warnow, T. ASTRAL-II: Coalescent-based species tree estimation with many hundreds of taxa and thousands of genes. Bioinformatics 2015, 31, i44–i52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, J.; Zhang, C.; Mirarab, S. ASTRAL-MP: Scaling ASTRAL to very large datasets using randomization and parallelization. Bioinformatics 2019, 35, 3961–3969. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Rabiee, M.; Sayyari, E.; Mirarab, S. ASTRAL-III: Polynomial time species tree reconstruction from partially resolved gene trees. BMC Bioinform. 2018, 19, 153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wise, M.J. Mean protein evolutionary distance: A method for comparative protein evolution and its application. PLoS ONE 2013, 8, e61276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braun, E.L. An evolutionary model motivated by physicochemical properties of amino acids reveals variation among proteins. Bioinformatics 2018, 34, i350–i356. [Google Scholar] [CrossRef]
- Sneath, P.H. Relations between chemical structure and biological activity in peptides. J. Biol. 1966, 12, 157–195. [Google Scholar] [CrossRef]
- Grantham, R. Amino acid difference formula to help explain protein evolution. Science 1974, 185, 862–864. [Google Scholar] [CrossRef]
- Xia, X.; Li, W.H. What amino acid properties affect protein evolution? J. Mol. Evol. 1998, 47, 557–564. [Google Scholar] [CrossRef]
- Jones, D.T.; Taylor, W.R.; Thornton, J.M. The rapid generation of mutation data matrices from protein sequences. Comput. Appl. Biosci. 1992, 8, 275–282. [Google Scholar] [CrossRef]
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Autom. Control 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Chen, L.; Qiu, Q.; Jiang, Y.; Wang, K.; Lin, Z.; Li, Z.; Bibi, F.; Yang, Y.; Wang, J.; Nie, W.; et al. Large-scale ruminant genome sequencing provides insights into their evolution and distinct traits. Science 2019, 364, eaav6202. [Google Scholar] [CrossRef] [PubMed]
- Heckeberg, N.S. The systematics of the Cervidae: A total evidence approach. PeerJ 2020, 8, e8114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Southey, B.R.; Rodriguez-Zas, S.L.; Rhodes, J.S.; Sweedler, J.V. Characterization of the prohormone complement in Amphiprion and related fish species integrating genome and transcriptome assemblies. PLoS ONE 2020, 15, e0228562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, H.; Yoshida, T.; Miyamoto, N.; Motoike, T.; Kurosu, H.; Shibata, K.; Yamanaka, A.; Williams, S.C.; Richardson, J.A.; Tsujino, N.; et al. Characterization of a family of endogenous neuropeptide ligands for the G protein-coupled receptors GPR7 and GPR8. Proc. Natl. Acad. Sci. USA 2003, 100, 6251–6256. [Google Scholar] [CrossRef] [Green Version]
- Couzens, M.; Liu, M.; Tuchler, C.; Kofler, B.; Nessler-Menardi, C.; Parker, R.M.; Klocker, H.; Herzog, H. Peptide YY-2 (PYY2) and pancreatic polypeptide-2 (PPY2): Species-specific evolution of novel members of the neuropeptide Y gene family. Genomics 2000, 64, 318–323. [Google Scholar] [CrossRef]
- Southey, B.R.; Zhang, P.; Keever, M.R.; Rymut, H.E.; Johnson, R.W.; Sweedler, J.V.; Rodriguez-Zas, S.L. Effects of maternal immune activation in porcine transcript isoforms of neuropeptide and receptor genes. J. Integr. Neurosci. 2021, 20, 21–31. [Google Scholar] [CrossRef]
- Zhang, Y.; Lu, L.; Furlonger, C.; Wu, G.E.; Paige, C.J. Hemokinin is a hematopoietic-specific tachykinin that regulates B lymphopoiesis. Nat. Immunol. 2000, 1, 392–397. [Google Scholar] [CrossRef]
- Lok, S.; Johnston, D.S.; Conklin, D.; Lofton-Day, C.E.; Adams, R.L.; Jelmberg, A.C.; Whitmore, T.E.; Schrader, S.; Griswold, M.D.; Jaspers, S.R. Identification of INSL6, a new member of the insulin family that is expressed in the testis of the human and rat. Biol. Reprod. 2000, 62, 1593–1599. [Google Scholar] [CrossRef] [Green Version]
- Fenger, M.; Johnsen, A.H. Alpha-amidated peptides derived from pro-opiomelanocortin in normal human pituitary. Biochem. J. 1988, 250, 781–788. [Google Scholar] [CrossRef] [Green Version]
- Bohlen, P.; Esch, F.; Shibasaki, T.; Baird, A.; Ling, N.; Guillemin, R. Isolation and characterization of a gamma 1-melanotropin-like peptide from bovine neurointermediate pituitary. FEBS Lett. 1981, 128, 67–70. [Google Scholar] [CrossRef] [Green Version]
- Jégou, S.; Cone, R.D.; Eberlé, A.N.; Vaudry, H. Chapter 111—Melanocortins. In Handbook of Biologically Active Peptides, 2nd ed.; Kastin, A.J., Ed.; Academic Press: Boston, MA, USA, 2013; pp. 838–844. [Google Scholar]
- Wikberg, J.E.; Muceniece, R.; Mandrika, I.; Prusis, P.; Lindblom, J.; Post, C.; Skottner, A. New aspects on the melanocortins and their receptors. Pharm. Res 2000, 42, 393–420. [Google Scholar] [CrossRef] [PubMed]
- Humphreys, M.H. Gamma-MSH, sodium metabolism, and salt-sensitive hypertension. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2004, 286, R417–R430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harmer, S.C.; Bicknell, A.B. Role of gamma-MSH peptides in the regulation of adrenal steroidogenesis. Peptides 2005, 26, 1944–1951. [Google Scholar] [CrossRef] [PubMed]
- Klusa, V.; Svirskis, S.; Opmane, B.; Muceniece, R.; Wikberg, J.E. Behavioural responses of gamma-MSH peptides administered into the rat ventral tegmental area. Acta Physiol. Scand. 1999, 167, 99–104. [Google Scholar] [CrossRef]
- Nyberg, F.; Hallberg, M. Neuropeptides in hyperthermia. Prog. Brain Res. 2007, 162, 277–293. [Google Scholar] [CrossRef]
- Russell, F.A.; King, R.; Smillie, S.-J.; Kodji, X.; Brain, S.D. Calcitonin gene-related peptide: Physiology and pathophysiology. Physiol. Rev. 2014, 94, 1099–1142. [Google Scholar] [CrossRef] [Green Version]
- Granstein, R.D.; Wagner, J.A.; Stohl, L.L.; Ding, W. Calcitonin gene-related peptide: Key regulator of cutaneous immunity. Acta Physiol. 2015, 213, 586–594. [Google Scholar] [CrossRef] [Green Version]
- Brain, S.D.; Grant, A.D. Vascular actions of calcitonin gene-related peptide and adrenomedullin. Physiol. Rev. 2004, 84, 903–934. [Google Scholar] [CrossRef] [Green Version]
- Pilot, M.; Malewski, T.; Moura, A.E.; Grzybowski, T.; Olenski, K.; Kaminski, S.; Fadel, F.R.; Alagaili, A.N.; Mohammed, O.B.; Bogdanowicz, W. Diversifying Selection Between Pure-Breed and Free-Breeding Dogs Inferred from Genome-Wide SNP Analysis. G3 2016, 6, 2285–2298. [Google Scholar] [CrossRef] [Green Version]
- Hekman, J.P.; Johnson, J.L.; Edwards, W.; Vladimirova, A.V.; Gulevich, R.G.; Ford, A.L.; Kharlamova, A.V.; Herbeck, Y.; Acland, G.M.; Raetzman, L.T.; et al. Anterior Pituitary Transcriptome Suggests Differences in ACTH Release in Tame and Aggressive Foxes. G3 2018, 8, 859–873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katafuchi, T.; Yasue, H.; Osaki, T.; Minamino, N. Calcitonin receptor-stimulating peptide: Its evolutionary and functional relationship with calcitonin/calcitonin gene-related peptide based on gene structure. Peptides 2009, 30, 1753–1762. [Google Scholar] [CrossRef] [PubMed]
- Dougherty, D.A. Cation-pi interactions involving aromatic amino acids. J. Nutr. 2007, 137, 1504S–1508S; discussion 1516S–1517S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahrestani, F.S. Bos frontalis and Bos gaurus (Artiodactyla: Bovidae). Mamm. Species 2018, 50, 34–50. [Google Scholar] [CrossRef]
- Frantz, L.A.; Schraiber, J.G.; Madsen, O.; Megens, H.J.; Cagan, A.; Bosse, M.; Paudel, Y.; Crooijmans, R.P.; Larson, G.; Groenen, M.A. Evidence of long-term gene flow and selection during domestication from analyses of Eurasian wild and domestic pig genomes. Nat. Genet. 2015, 47, 1141–1148. [Google Scholar] [CrossRef]
- Lewis, J.E.; Brameld, J.M.; Jethwa, P.H. Neuroendocrine Role for VGF. Front. Endocrinol. 2015, 6, 3. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Reichert, S.; Singh, C.; Oikonomou, G.; Rihel, J.; Prober, D.A. Light-Dependent Regulation of Sleep and Wake States by Prokineticin 2 in Zebrafish. Neuron 2017, 95, 153–168.e156. [Google Scholar] [CrossRef] [Green Version]
- Sapolsky, R.M.; Romero, L.M.; Munck, A.U. How do glucocorticoids influence stress responses? Integrating permissive, suppressive, stimulatory, and preparative actions. Endocr. Rev. 2000, 21, 55–89. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Suborder and IF or Fam 1 | N 2 | Parvorder/Subfamily/Tribe/Genus 3 | ||
|---|---|---|---|---|
| D | W | |||
| Ruminantia | ||||
| Bovidae | 7 | 50 | Bovinae: 4,12; Caprinae: 2,12; Alcelaphinae: 0,4; Reduncinae: 0,3; Cephalophinae:0,3; Hippotraginae: 0,4; Antilopinae: 0,12; Aepycerotinae: 0,1 | |
| Cervidae | 1 | 15 | Muntiacinae: 0,3; Cervinae: 0,5; Hydropotinae: 0,1; Odocoileinae: 1,6 | |
| Other | 0 | 9 | Moschidae: 0,3; Tragulidae: 0,2; Giraffidae: 0,3; Antilocapridae: 0,1 | |
| Suina | ||||
| 1 | o | Sus: 1,0; Catagonus: 0,1 | ||
| Tylopoda | ||||
| Camelidae | 4 | 3 | Camelini: 2,1; Lamini: 2,2 | |
| Whippomorpha | ||||
| 0 | 27 | Hippopotamidae: 0,1; Odontoceti: 0,19; Mysticeti: 0,7 | ||
| Total | 13 | 105 | ||
| Taxonomic Group 1 | mS 2 | Parameter 3 | |||||||
|---|---|---|---|---|---|---|---|---|---|
| A | C | P | V | ||||||
| − | + | − | + | − | + | − | + | ||
| Overall | |||||||||
| All | 81.5 | 52 | 5 | 4 | 64 | 17 | 19 | 51 | 3 |
| Domestic | 10 | 31 | 5 | 2 | 32 | 11 | 13 | 26 | 5 |
| Wild | 75.5 | 50 | 6 | 3 | 62 | 17 | 18 | 49 | 6 |
| Wild terrestrial | 56 | 51 | 3 | 1 | 56 | 11 | 23 | 48 | 3 |
| Ruminantia | |||||||||
| All | 55 | 51 | 5 | 3 | 50 | 13 | 24 | 51 | 5 |
| Domestic | 6 | 23 | 5 | 3 | 27 | 5 | 12 | 18 | 4 |
| Wild | 52 | 43 | 5 | 0 | 54 | 11 | 22 | 47 | 4 |
| Bovidae | 38 | 35 | 5 | 6 | 44 | 10 | 17 | 38 | 5 |
| Bovidae Antilopinae | 11 | 18 | 8 | 2 | 24 | 7 | 4 | 24 | 3 |
| Bovidae Bovinae | 10 | 22 | 1 | 6 | 20 | 10 | 6 | 15 | 6 |
| Bovidae Caprinae | 7 | 13 | 7 | 4 | 22 | 8 | 14 | 25 | 2 |
| Cervidae | 11 | 11 | 2 | 6 | 2 | 7 | 1 | 4 | 0 |
| Tylopoda | |||||||||
| All | 4 | 20 | 1 | 13 | 10 | 9 | 7 | 10 | 1 |
| Whippomorpha | |||||||||
| All | 20 | 32 | 3 | 6 | 23 | 13 | 11 | 39 | 0 |
| Cetacea Mysticeti | 5 | 16 | 6 | 4 | 16 | 18 | 8 | 13 | 2 |
| Cetacea Odontoceti | 14 | 30 | 3 | 7 | 21 | 11 | 10 | 31 | 3 |
| Parameter 1 | ||||||||
|---|---|---|---|---|---|---|---|---|
| A | C | P | V | |||||
| Symbol 2 | All | Rum | All | Rum | All | Rum | All | Rum |
| Domestic | ||||||||
| APLN | 3.62 | 9.80 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Wild terrestrial | ||||||||
| ADCYAP1 | −1.04 | −1.27 | 0.00 | 0.00 | 0.00 | −0.95 | 0.00 | 0.00 |
| AVP | 0.00 | 0.00 | 0.74 | 1.22 | 0.00 | 0.00 | 0.00 | 0.00 |
| CCK | 0.00 | 1.70 | 2.03 | 2.18 | 0.00 | 0.00 | 0.00 | −0.90 |
| GHRH | −1.35 | −1.26 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| TOR2X | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | −1.32 | −1.57 |
| VGF | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | −1.23 | −1.39 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Southey, B.R.; Rodriguez-Zas, S.L. Changes in Neuropeptide Prohormone Genes among Cetartiodactyla Livestock and Wild Species Associated with Evolution and Domestication. Vet. Sci. 2022, 9, 247. https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci9050247
Southey BR, Rodriguez-Zas SL. Changes in Neuropeptide Prohormone Genes among Cetartiodactyla Livestock and Wild Species Associated with Evolution and Domestication. Veterinary Sciences. 2022; 9(5):247. https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci9050247
Chicago/Turabian StyleSouthey, Bruce R., and Sandra L. Rodriguez-Zas. 2022. "Changes in Neuropeptide Prohormone Genes among Cetartiodactyla Livestock and Wild Species Associated with Evolution and Domestication" Veterinary Sciences 9, no. 5: 247. https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci9050247