
Photoperiodic Modulation in Immune and Reproductive Systems in Japanese Quails (Coturnix japonica): A Morphometric Perspective

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Hematological Analysis
2.3. Gross Morphometry and Histological Sample Preparation
2.4. Histomorphometric Analysis
2.5. Hormonal Analysis
2.6. Statistical Analysis
3. Results
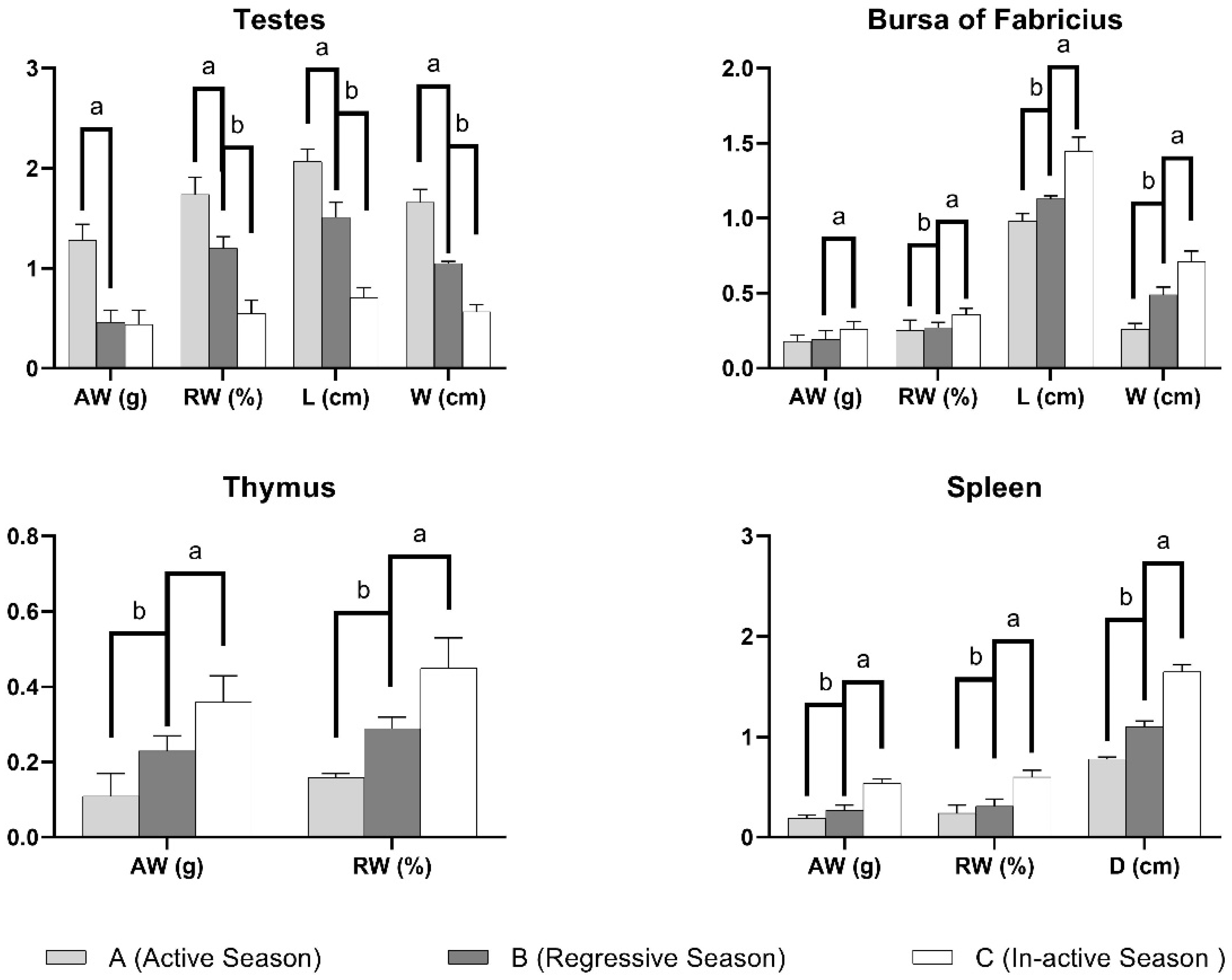
3.1. Anatomical Parameters
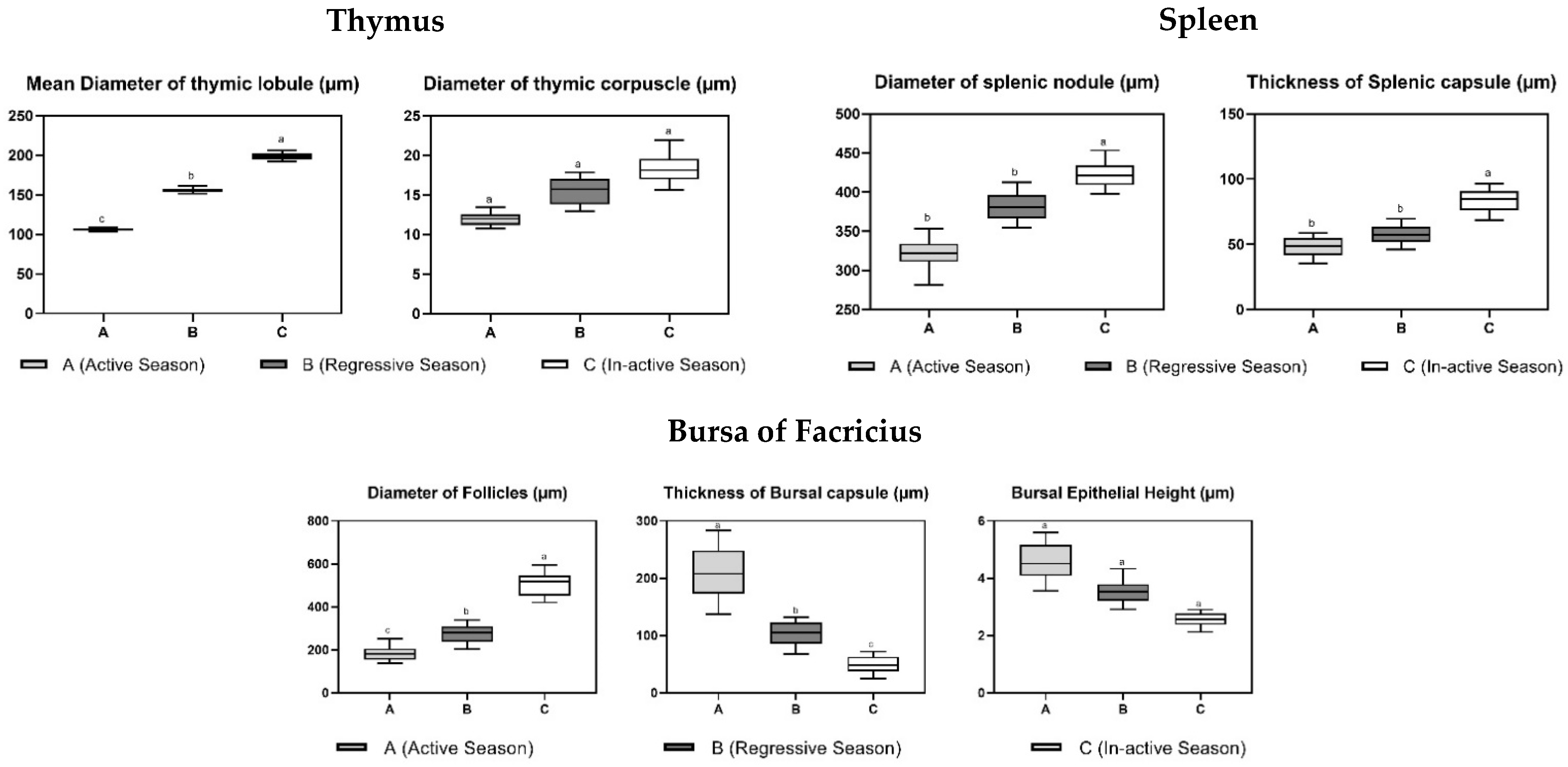
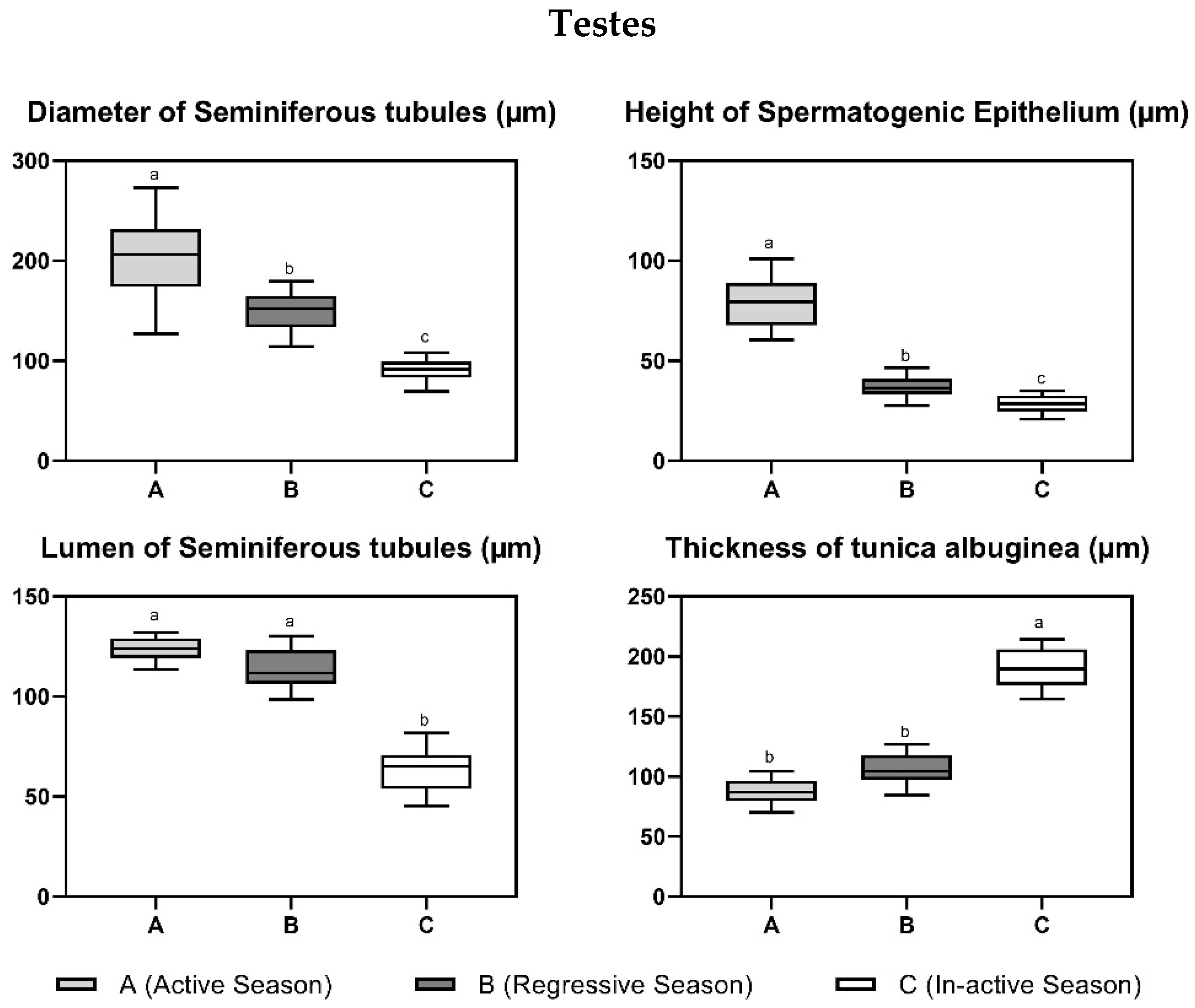
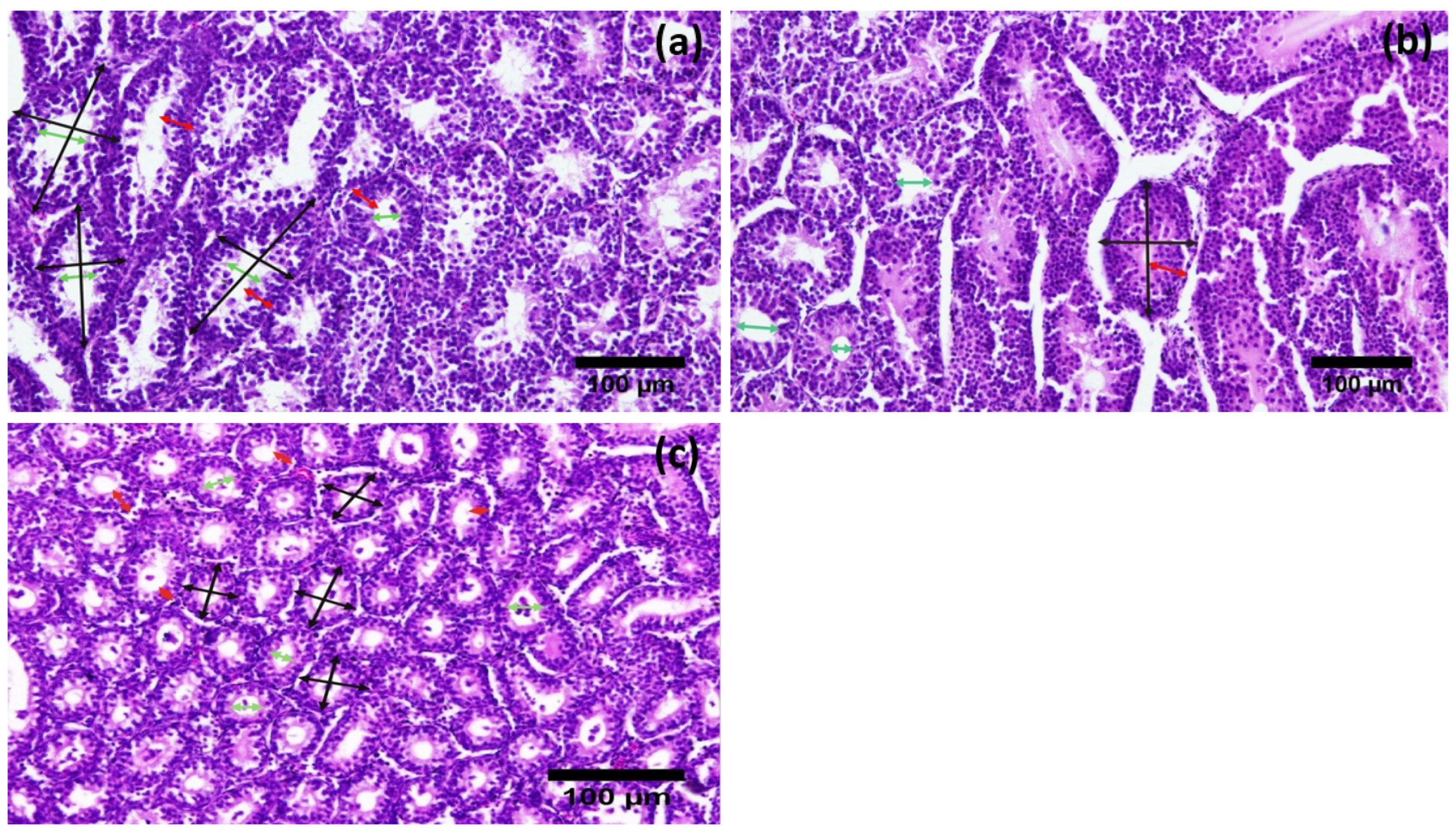
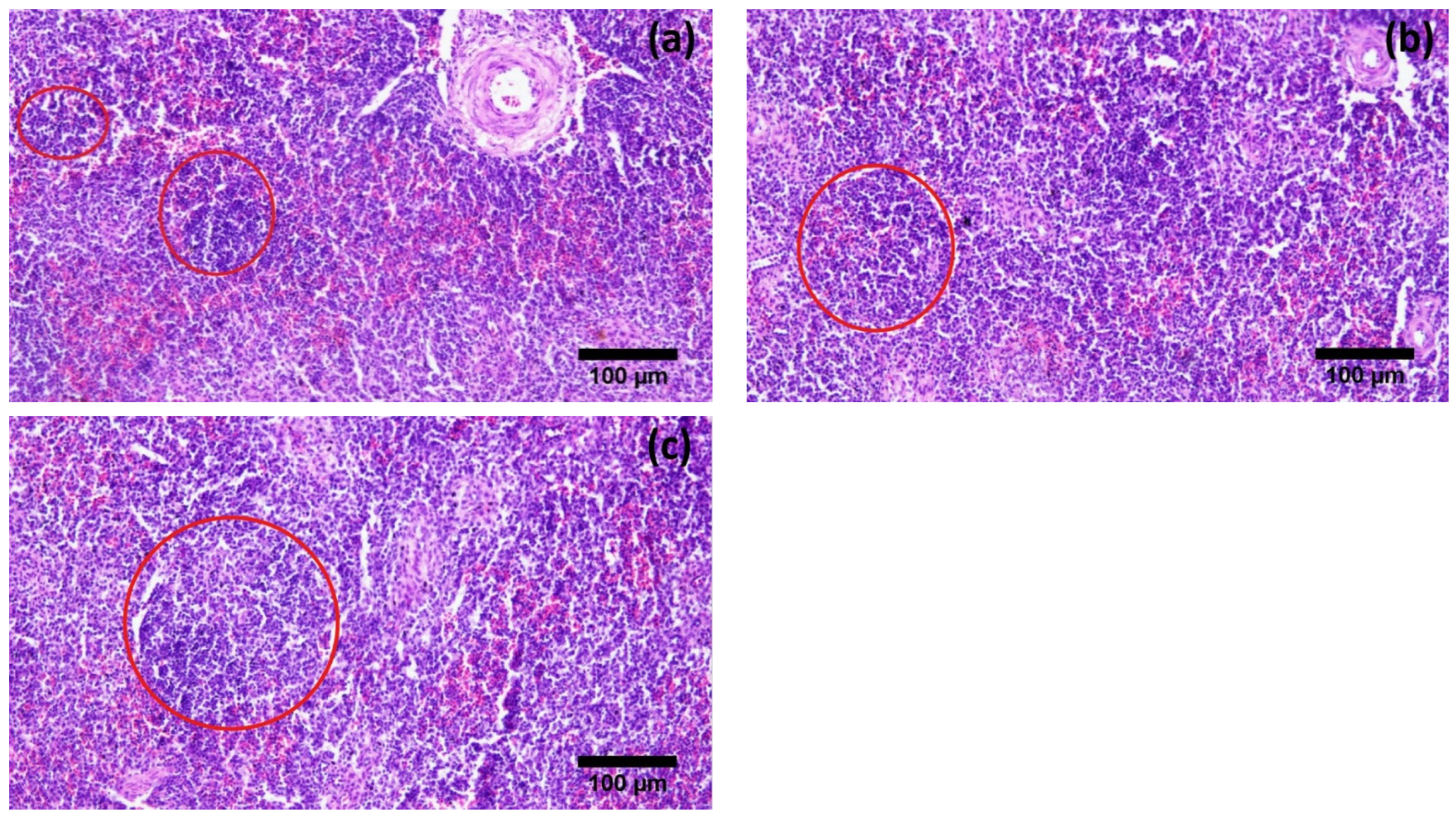
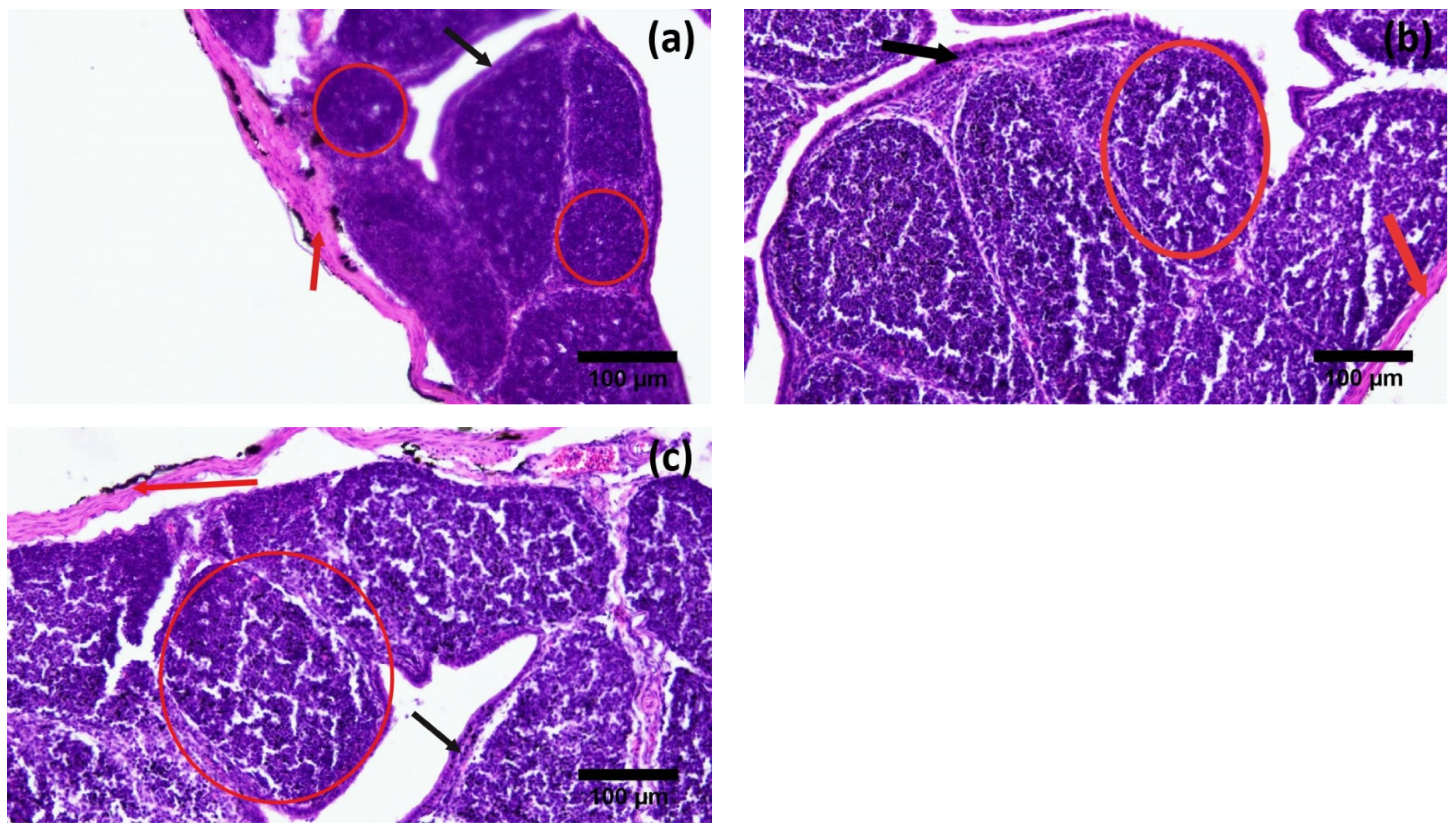
3.2. Histological Parameters
3.3. Hematological Parameters
3.4. Hormonal Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Williams, T.D. Physiological Adaptations for Breeding in Birds; Princeton University Press: Princeton, NJ, USA, 2012; ISBN 978-1-4008-4279-7. [Google Scholar]
- Ruiz, M.; French, S.S.; Demas, G.E.; Martins, E.P. Food supplementation and testosterone interact to influence reproductive behavior and immune function in Sceloporus graciosus. Horm. Behav. 2010, 57, 134–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasselquist, D. Comparative immunoecology in birds: Hypotheses and tests. J. Ornithol. 2007, 148, 571–582. [Google Scholar] [CrossRef]
- Koutsos, E.A.; Klasing, K.C. Chapter 17—Factors Modulating the Avian Immune System. In Avian Immunology, 2nd ed.; Schat, K.A., Kaspers, B., Kaiser, P., Eds.; Academic Press: Boston, MA, USA, 2014; pp. 299–313. ISBN 978-0-12-396965-1. [Google Scholar]
- Akbar, Z.; Qureshi, A.S. Effects of Seasonal Variation in Different Reproductive Phases on the Cellular Response of Bursa and Testes in Japanese Quail (Coturnix japonica). Pak. Vet. J. 2012, 32, 525–529. [Google Scholar]
- Ali, M.Z.; Qureshi, A.S.; Rehan, S.; Akbar, S.Z.; Manzoor, A. Seasonal variations in histomorphology of testes and bursa, immune parameters and serum testosterone concentration in male guinea fowl (Numida meleagris). Pak. Vet. J. 2015, 35, 88–92. [Google Scholar]
- John, J.L. The avian spleen: A neglected organ. Q. Rev. Biol. 1994, 69, 327–351. [Google Scholar] [CrossRef]
- Smith, K.G.; Hunt, J.L. On the use of spleen mass as a measure of avian immune system strength. Oecologia 2004, 138, 28–31. [Google Scholar] [CrossRef]
- Brendolan, A.; Rosado, M.M.; Carsetti, R.; Selleri, L.; Dear, T.N. Development and function of the mammalian spleen. Bioessays 2007, 29, 166–177. [Google Scholar] [CrossRef]
- Pabst, R. Plasticity and heterogeneity of lymphoid organs: What are the criteria to call a lymphoid organ primary, secondary or tertiary? Immunol. Lett. 2007, 112, 1–8. [Google Scholar] [CrossRef]
- Møller, A.P.; Erritzøe, J.; Saino, N. Seasonal Changes in Immune Response and Parasite Impact on Hosts. Am. Nat. 2003, 161, 657–671. [Google Scholar] [CrossRef] [Green Version]
- Dunn, J.C.; Goodman, S.J.; Benton, T.G.; Hamer, K.C. Avian blood parasite infection during the non-breeding season: An overlooked issue in declining populations? BMC Ecol. 2013, 13, 30. [Google Scholar] [CrossRef] [Green Version]
- Cornelius, J.; Zylberberg, M.; Breuner, C.; Gleiss, A.C.; Hahn, T. Assessing the role of reproduction and stress in the spring emergence of haematozoan parasites in birds. J. Exp. Biol. 2014, 217, 841–849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majewski, P.; Markowska, M.; Pawlak, J.; Piesiewicz, A.; Turkowska, E.; Skwarlo-Sonta, K. Pineal gland and melatonin: Impact on the seasonality of immune defence in mammals and birds. Adv. Neuroimmune Biol. 2012, 3, 95–108. [Google Scholar] [CrossRef]
- Lopes, L.E.; Teixeira, J.P.G.; Meireles, R.C.; Bastos, D.S.S.; de Oliveira, L.L.; Solar, R. High seasonal variation of plasma testosterone levels for a tropical grassland bird resembles patterns of temperate birds. Physiol. Biochem. Zool. 2021, 94, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.S.; Haldar, C. Melatonin prevents testosterone-induced suppression of immune parameters and splenocyte proliferation in Indian tropical jungle bush quail, Perdicula asiatica. Gen. Comp. Endocrinol. 2005, 141, 226–232. [Google Scholar] [CrossRef]
- Aire, T.A. Morphological changes in the efferent ducts during the main phases of the reproductive cycle of birds. J. Morphol. 2002, 253, 64–75. [Google Scholar] [CrossRef]
- Nasar, A.; Rahman, A.; Hoque, N.; Talukder, A.K.; Das, Z.C. A survey of Japanese quail (Coturnix coturnix japonica) farming in selected areas of Bangladesh. Vet. World 2016, 9, 940–947. [Google Scholar] [CrossRef] [Green Version]
- Lukanov, H. Domestic quail (Coturnix japonica domestica), is there such farm animal? Worlds Poult. Sci. J. 2019, 75, 547–558. [Google Scholar] [CrossRef]
- Singh, S.S.; Haldar, C. Peripheral melatonin modulates seasonal immunity and reproduction of Indian tropical male bird Perdicula asiatica. Comp. Biochem. Physiol. Part. A Mol. Integr. Physiol. 2007, 146, 446–450. [Google Scholar] [CrossRef]
- Hau, M. Timing of Breeding in Variable Environments: Tropical Birds as Model Systems. Horm. Behav. 2001, 40, 281–290. [Google Scholar] [CrossRef]
- Pap, P.L.; Czirják, G.Á.; Vágási, C.I.; Barta, Z.; Hasselquist, D. Sexual dimorphism in immune function changes during the annual cycle in house sparrows. Naturwissenschaften 2010, 97, 891–901. [Google Scholar] [CrossRef]
- Kuhlman, J.R.; Martin, L.B. Captivity affects immune redistribution to skin in a wild bird. Funct. Ecol. 2010, 24, 830–837. [Google Scholar] [CrossRef]
- Martin, L.B.; Brace, A.J.; Urban, A.; Coon, C.A.C.; Liebl, A.L. Does immune suppression during stress occur to promote physical performance? J. Exp. Biol. 2012, 215, 4097–4103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leopold, A.S. Age Determination in Quail. J. Wildl. Manag. 1939, 3, 261. [Google Scholar] [CrossRef]
- Pereira, D.F.; Batista, E.D.; Sanches, F.T.; Gabriel Filho, L.R.; Bueno, L.G. Diferenças comportamentais de poedeiras em diferentes ambientes térmicos. Energ. Na Agric. 2015, 30, 33–40. [Google Scholar] [CrossRef] [Green Version]
- Raharjo, S.; Rahayu, E.S.; Purnomo, S.H. Factors affecting quail egg production under the changing climate at Kulonprogo Regency, Indonesia. IOP Conf. Ser. Earth Environ. Sci. 2018, 200, 012012. [Google Scholar] [CrossRef] [Green Version]
- Molino, A.B.; Garcia, E.A.; Santos, G.C.; Vieira Filho, J.A.; Baldo, G.A.A.; Almeida Paz, I.C.L. Photostimulation of Japanese quail. Poult. Sci. 2015, 94, 156–161. [Google Scholar] [CrossRef] [PubMed]
- Bancroft, J.D.; Gamble, M. Theory and Practice of Histological Techniques; Elsevier Health Sciences: Amsterdam, The Netherlands, 2008; ISBN 0-443-10279-1. [Google Scholar]
- Carisch, L.; Stirn, M.; Hatt, J.M.; Federer, K.; Hofmann-Lehmann, R.; Riond, B. White blood cell count in birds: Evaluation of a commercially available method. BMC Vet. Res. 2019, 15, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Kriegsfeld, L.J.; Ubuka, T.; Bentley, G.E.; Tsutsui, K. Seasonal control of gonadotropin-inhibitory hormone (GnIH) in birds and mammals. Front. Neuroendocr. 2015, 37, 65–75. [Google Scholar] [CrossRef] [Green Version]
- Aire, T.A.; Ozegbe, P.C. The testicular capsule and peritubular tissue of birds: Morphometry, histology, ultrastructure and immunohistochemistry. J. Anat. 2007, 210, 731–740. [Google Scholar] [CrossRef]
- Leska, A.; Kiezun, J.; Kaminska, B.; Dusza, L. Seasonal changes in the expression of the androgen receptor in the testes of the domestic goose (Anser anser f. domestica). Gen. Comp. Endocrinol. 2012, 179, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Dixit, A.S.; Singh, N.S. Environmental control of seasonal reproduction in the wild and captive Eurasian Tree Sparrow (Passer montanus) with respect to variations in gonadal mass, histology, and sex steroids. Can. J. Zool. 2013, 91, 302–312. [Google Scholar] [CrossRef]
- Tae, H.; Jang, B.; Ahn, D.; Choi, E.; Kang, H.; Kim, N.; Lee, J.; Park, S.; Yang, H.; Kim, I. Morphometric studies on the testis of Korean ring-necked pheasant (Phasianus colchicus karpowi) during the breeding and non-breeding seasons. Vet. Res. Commun. 2005, 29, 629–643. [Google Scholar] [CrossRef] [PubMed]
- Silverin, B.; Fänge, R.; Viebke, P.-A.; Westin, J. Seasonal Changes in Mass and Histology of the Spleen in Willow Tits Parus montanus. J. Avian Biol. 1999, 30, 255–262. [Google Scholar] [CrossRef]
- Höhn, E.O. Seasonal recrudescence of the thymus in adult birds. Can. J. Biochem. Physiol. 1956, 34, 90–101. [Google Scholar] [CrossRef] [PubMed]
- Węgrzyn, E. Resource allocation between growth and endothermy allows rapid nestling development at low feeding rates in a species under high nest predation. J. Avian Biol. 2013, 44, 383–389. [Google Scholar] [CrossRef]
- Deerenberg, C.; Biebach, H.; Bauchinger, U. Spleen size variation during long-distance migration in the garden warbler Sylvia borin. Avian Sci. 2002, 2, 217–226. [Google Scholar]
- Thompson, H.L.; Smithey, M.J.; Surh, C.D.; Nikolich-Žugich, J. Functional and homeostatic impact of age-related changes in lymph node stroma. Front. Immunol. 2017, 8, 706. [Google Scholar] [CrossRef] [Green Version]
- Ciriaco, E.; Píñera, P.P.; Díaz-Esnal, B.; Laurà, R. Age-related changes in the avian primary lymphoid organs (thymus and bursa of Fabricius). Microsc. Res. Tech. 2003, 62, 482–487. [Google Scholar] [CrossRef]
- Mešťanová, V.; Varga, I. Morphological view on the evolution of the immunity and lymphoid organs of vertebrates, focused on thymus. Biologia 2016, 71, 1080–1097. [Google Scholar] [CrossRef]
- Kannan, T.A.; Ramesh, G.; Ushakumari, S.; Vairamuthu, S.; Sivakumar, M. Age related ultrastructural changes in lymphoid organs of Nandanam layer chicken (Gallus domesticus). J. Entomol. Zool. Stud. 2018, 6, 1417–1421. [Google Scholar]
- Bentley, G.E.; Demas, G.E.; Nelson, R.J.; Ball, G.F. Melatonin, immunity and cost of reproductive state in male European starlings. Proc. R. Soc. Lond. Ser. B Biol. Sci. 1998, 265, 1191–1195. [Google Scholar] [CrossRef] [PubMed]
- Abdul-Rahman, I.I.; Obese, F.Y.; Jeffcoate, I.A. Developmental changes in the histological structure of the testes, and testosterone profiles in male guinea fowls (Numida meleagris). Theriogenology 2017, 101, 114–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Annual Reproductive Season | ||
|---|---|---|---|
| Active Season (April–May) | Regressive Season (September–October) | Nonactive Season (January–February) | |
| Testes | |||
| Absolute Weight (g) | 1.28 ± 0.16 a | 0.46 ± 0.12 b | 0.44 ± 0.14 b |
| Relative Weight (%) | 1.74 ± 0.17 a | 1.2 ± 0.12 b | 0.55 ± 0.13 c |
| Length (cm) | 2.07 ± 0.12 a | 1.51 ± 0.15 b | 0.71 ± 0.1 c |
| Width (cm) | 1.66 ± 0.13 a | 1.05 ± 0.02 b | 0.57 ± 0.07 c |
| Bursa of Fabricius | |||
| Absolute Weight (g) | 0.18 ± 0.04 b | 0.19 ± 0.06 b | 0.26 ± 0.05 a |
| Relative Weight (%) | 0.25 ± 0.07 c | 0.27 ± 0.035 b | 0.36 ± 0.04 a |
| Length (cm) | 0.98 ± 0.05 b | 1.13 ± 0.02 b | 1.45 ± 0.09 a |
| Width (cm) | 0.26 ± 0.04 c | 0.49 ± 0.05 b | 0.71 ± 0.07 a |
| Thymus | |||
| Absolute Weight (g) | 0.11 ± 0.06 c | 0.23 ± 0.04 b | 0.36 ± 0.07 a |
| Relative Weight (%) | 0.16 ± 0.01 c | 0.29 ± 0.03 b | 0.45 ± 0.08 a |
| Spleen | |||
| Absolute Weight (g) | 0.19 ± 0.03 c | 0.27 ± 0.05 b | 0.54 ± 0.04 a |
| Relative Weight (%) | 0.24 ± 0.08 b | 0.31 ± 0.07 b | 0.60 ± 0.07 a |
| Diameter (cm) | 0.78 ± 0.02 c | 1.1 ± 0.06 b | 1.65 ± 0.07 a |
| Parameter | Annual Reproductive Season | ||
|---|---|---|---|
| Active Phase (April–May) | Regressive Phase (September–October) | Nonactive Phase (January–February) | |
| Testes | |||
| Diameter of seminiferous tubules (µm) | 203.37 ± 65.22 a | 151.71 ± 28.22 b | 91.04 ± 11.41 b |
| Height of spermatogenic epithelium (µm) | 80.66 ± 21.22 a | 37.06 ± 9.67 b | 27.80 ± 7.22 c |
| Lumen of seminiferous tubules (µm) | 122.71 ± 9.98 b | 114.4 ± 15.93 b | 63.24 ± 19.11 a |
| Thickness of tunica albuginea (µm) | 86.81 ± 17.75 b | 105.29 ± 22.15 b | 191.10 ± 26.85 a |
| Bursa of Fabricius | |||
| Diameter of follicles (µm) | 195.5 ± 58.54 c | 271.13 ± 69.39 b | 508.52 ± 91.06 a |
| Thickness of capsule (µm) | 213.46 ± 78.49 a | 101.42 ± 37.08 b | 48.51 ± 23.94 c |
| Epithelial height (µm) | 4.57 ± 1.09 a | 3.61 ± 0.73 a | 2.53 ± 0.4 a |
| Thymus | |||
| Mean diameter of thymic lobule (µm) | 105.63 ± 2.59 c | 156.73 ± 5.47 b | 199.01 ± 7.03 a |
| Diameter of thymic corpuscle (µm) | 12.11 ± 1.34 a | 15.16 ± 2.71 a | 18.79 ± 3.25 a |
| Spleen | |||
| Diameter of splenic nodule (µm) | 324.58 ± 33.34 b | 383.74 ± 28.86 b | 426.52 ± 29.47 a |
| Thickness of capsule (µm) | 46.68 ± 11.96 b | 57.74 ± 12.03 b | 82.19 ± 14.57 a |
| Parameter | Annual Reproductive Season | ||
|---|---|---|---|
| Active Phase (April–May) | Regressive Phase (September–October) | Nonactive Phase (January–February) | |
| Immunological Analysis | |||
| Total Leukocyte Count (×103/µL) | 109.5 ± 1.95 a | 157.25 ± 2.37 b | 189.3 ± 6.53 c |
| Lymphocyte Count (×103/µL) | 79.34 ± 2.79 a | 113.94 ± 3.16 b | 137.18 ± 4.92 c |
| Hormonal Analysis | |||
| Serum Testosterone (ng/mL) | 2.45 ± 0.035 a | 0.60 ± 0.024 b | 0.27 ± 0.011 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hayat, K.; Raza, A.; Anas, A.; Qureshi, A.S.; Rehan, S.; Rabbani, A.H.; Rehman, H.F.u.; Qamar, A.G.; Rehman, T.; Deeba, F.; et al. Photoperiodic Modulation in Immune and Reproductive Systems in Japanese Quails (Coturnix japonica): A Morphometric Perspective. Vet. Sci. 2022, 9, 248. https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci9050248
Hayat K, Raza A, Anas A, Qureshi AS, Rehan S, Rabbani AH, Rehman HFu, Qamar AG, Rehman T, Deeba F, et al. Photoperiodic Modulation in Immune and Reproductive Systems in Japanese Quails (Coturnix japonica): A Morphometric Perspective. Veterinary Sciences. 2022; 9(5):248. https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci9050248
Chicago/Turabian StyleHayat, Khizar, Ali Raza, Aitzaz Anas, Anas Sarwar Qureshi, Sarmad Rehan, Ameer Hamza Rabbani, Hafiz Faseeh ur Rehman, Abdul Ghaffar Qamar, Tayyab Rehman, Farah Deeba, and et al. 2022. "Photoperiodic Modulation in Immune and Reproductive Systems in Japanese Quails (Coturnix japonica): A Morphometric Perspective" Veterinary Sciences 9, no. 5: 248. https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci9050248