
Computational Modeling of Blood Flow Hemodynamics for Biomechanical Investigation of Cardiac Development and Disease


Abstract
:1. Introduction
2. Numerical Modeling
2.1. Governing Equations in Solid and Fluid Domains
2.2. Meshing and Mesh Independence
2.3. Material Properties and Flow Characteristics
2.4. Boundary Conditions
2.5. Types of Numerical Models
2.6. Parameters for Hemodynamic Assessment
2.7. Uncertainty Quantification and Stochastic Sensitivity Analysis
3. Chicken Embryo Models
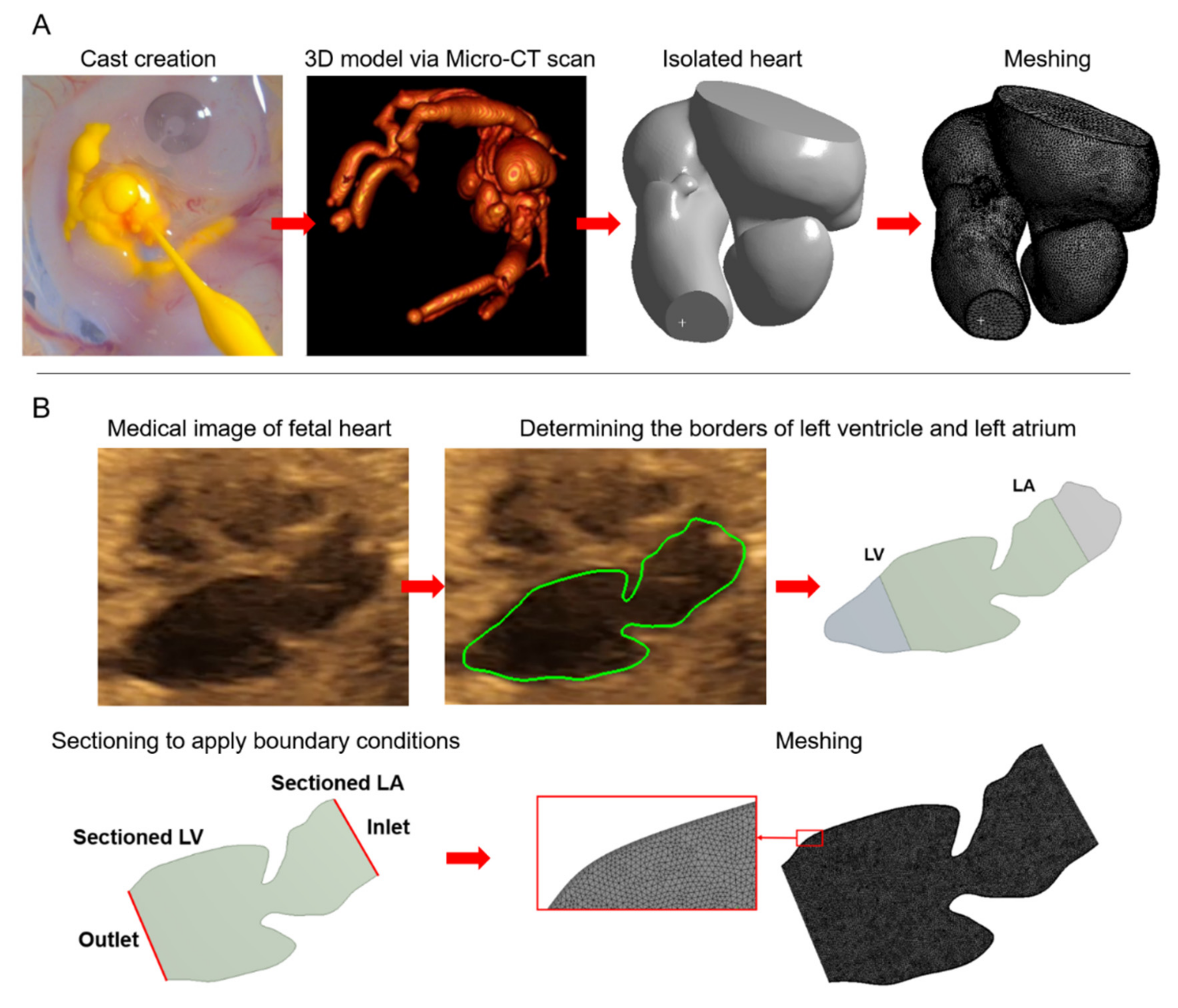
3.1. Imaging and 3D Model Generation
3.2. In Vivo Blood Velocity Measurements
3.3. CFD Results of Chicken Embryo Models
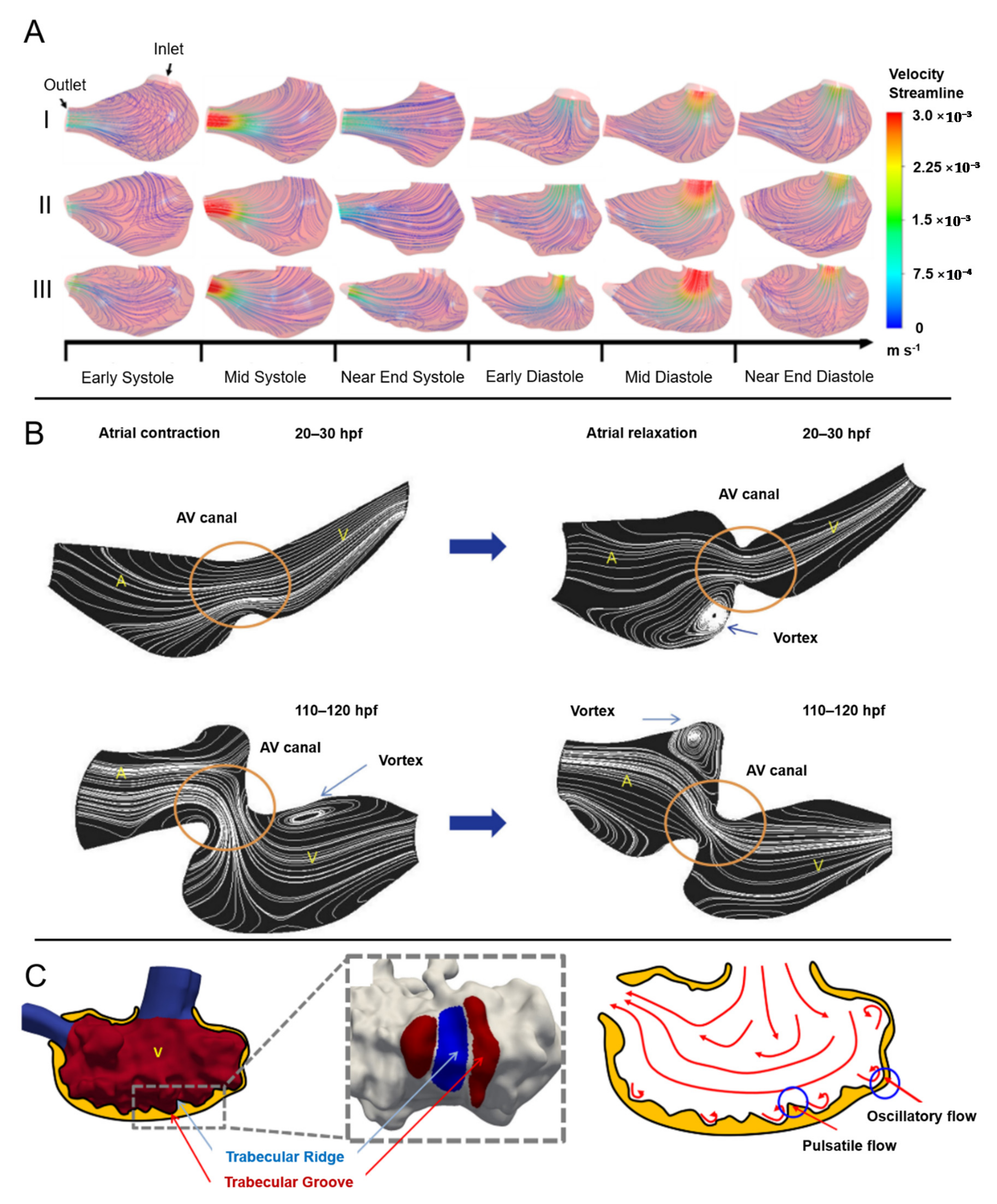
3.3.1. CFD Studies on Normally Developing Embryonic Chicken Hearts
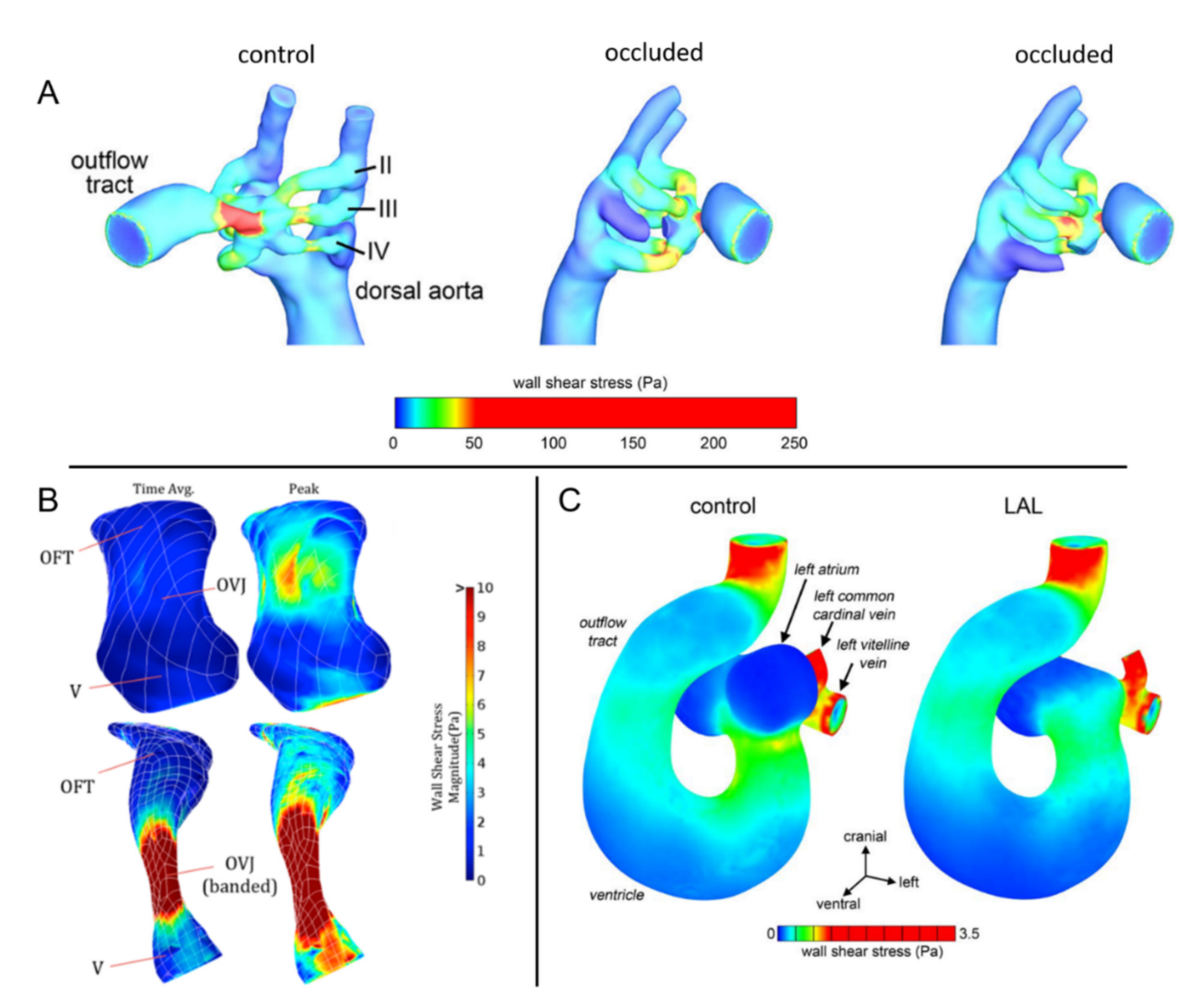
3.3.2. CFD Studies on Cardiac Defects in Chicken Embryos
4. Zebrafish Embryo Models
4.1. Microscopic Imaging of Zebrafish Embryo
4.2. CFD Studies for Normally Developing Zebrafish Embryo Hearts
4.3. CFD Studies on Defected Embryonic Zebrafish Hearts
5. Human Fetal Heart Models
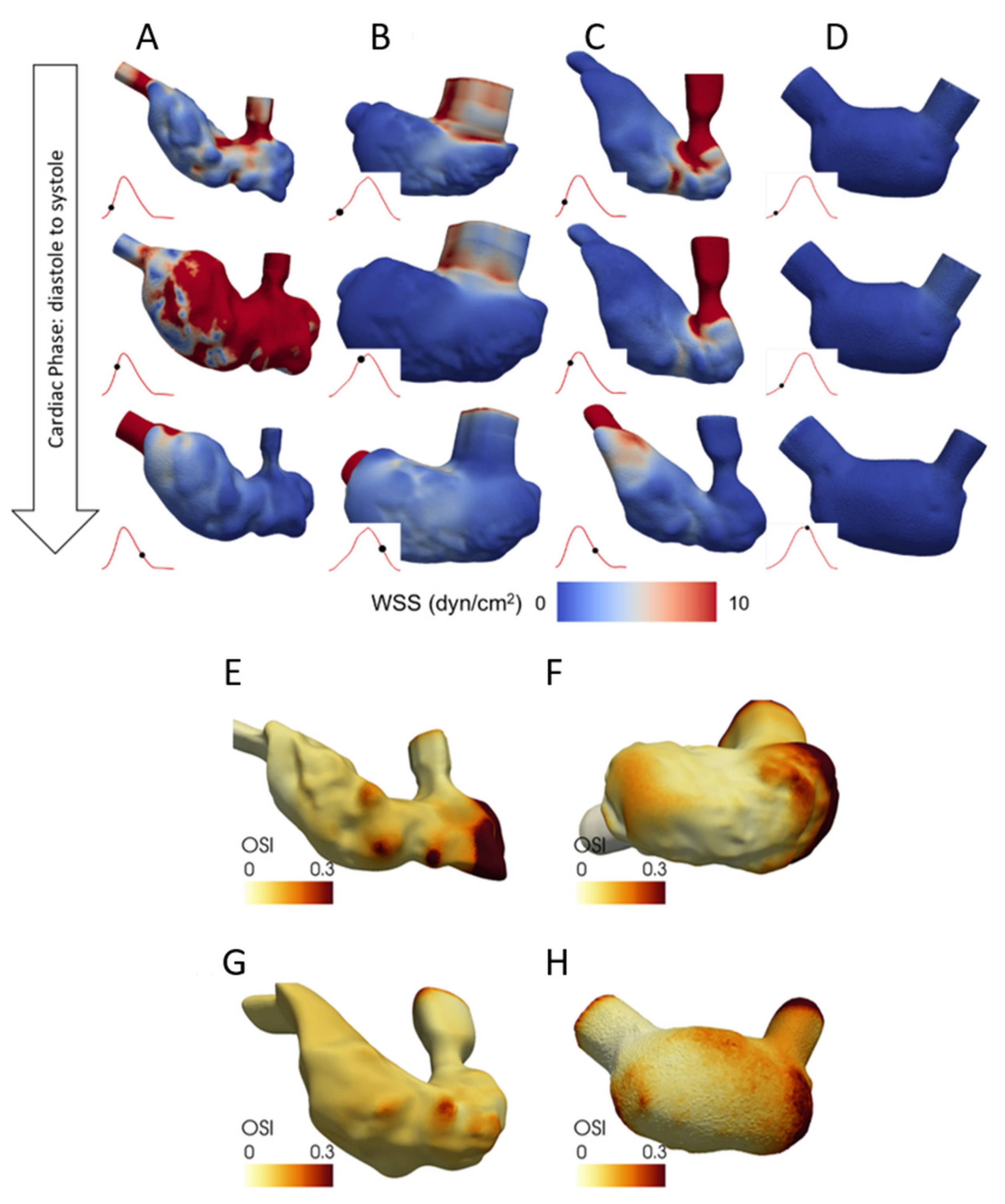
5.1. CFD Studies on Normal Human Fetal Hearts
5.2. CFD Studies on Defected Human Fetal Hearts
6. Clinical Utility of CFD Simulations
7. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Burggren, W.W. What Is the Purpose of the Embryonic Heart Beat? or How Facts Can Ultimately Prevail over Physiological Dogma. Physiol. Biochem. Zoöl. 2004, 77, 333–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salman, H.E.; Yazicioglu, Y. Computational analysis for non-invasive detection of stenosis in peripheral arteries. Med. Eng. Phys. 2019, 70, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, J.I.; Kaplan, S. The incidence of congenital heart disease. J. Am. Coll. Cardiol. 2002, 39, 1890–1900. [Google Scholar] [CrossRef] [Green Version]
- Øyen, N.; Poulsen, G.; Boyd, H.A.; Wohlfahrt, J.; Jensen, P.K.A.; Melbye, M. Recurrence of Congenital Heart Defects in Families. Circulation 2009, 120, 295–301. [Google Scholar] [CrossRef] [Green Version]
- Hierck, B.P.; van der Heiden, K.; Poelma, C.; Westerweel, J.; Poelmann, R.E. Fluid Shear Stress and Inner Curvature Remodeling of the Embryonic Heart. Choosing the Right Lane! Sci. World J. 2008, 8, 939501. [Google Scholar] [CrossRef]
- Poelmann, R.E.; Gittenberger-de Groot, A.C.; Hierck, B.P. The development of the heart and microcirculation: Role of shear stress. Med. Biol. Eng. Comput. 2008, 46, 479–484. [Google Scholar] [CrossRef] [Green Version]
- Groenendijk, B.C.; Hierck, B.P.; Vrolijk, J.; Baiker, M.; Pourquie, M.J.B.M.; Gittenberger-de Groot, A.C.; Poelmann, R.E. Changes in Shear Stress–Related Gene Expression After Experimentally Altered Venous Return in the Chicken Embryo. Circ. Res. 2005, 96, 1291–1298. [Google Scholar] [CrossRef] [Green Version]
- De Almeida, A.; McQuinn, T.; Sedmera, D. Increased Ventricular Preload Is Compensated by Myocyte Proliferation in Normal and Hypoplastic Fetal Chick Left Ventricle. Circ. Res. 2007, 100, 1363–1370. [Google Scholar] [CrossRef] [Green Version]
- Santhanakrishnan, A.; Miller, L.A. Fluid Dynamics of Heart Development. Cell Biochem. Biophys. 2011, 61, 1–22. [Google Scholar] [CrossRef]
- Courchaine, K.; Rugonyi, S. Quantifying blood flow dynamics during cardiac development: Demystifying computational methods. Philos. Trans. R. Soc. B Biol. Sci. 2018, 373, 20170330. [Google Scholar] [CrossRef] [Green Version]
- Courchaine, K.; Rykiel, G.; Rugonyi, S. Influence of blood flow on cardiac development. Prog. Biophys. Mol. Biol. 2018, 137, 95–110. [Google Scholar] [CrossRef] [PubMed]
- Khanafer, K.M.; Bull, J.L.; Upchurch, G.R.; Berguer, R. Turbulence Significantly Increases Pressure and Fluid Shear Stress in an Aortic Aneurysm Model under Resting and Exercise Flow Conditions. Ann. Vasc. Surg. 2007, 21, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Lindsey, S.E.; Butcher, J.T.; Yalcin, H.C. Mechanical regulation of cardiac development. Front. Physiol. 2014, 5, 318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campinho, P.; Vilfan, A.; Vermot, J. Blood Flow Forces in Shaping the Vascular System: A Focus on Endothelial Cell Behavior. Front. Physiol. 2020, 11, 552. [Google Scholar] [CrossRef]
- Lee, J.; Vedula, V.; Baek, K.I.; Chen, J.; Hsu, J.J.; Ding, Y.; Chang, C.-C.; Kang, H.; Small, A.; Fei, P.; et al. Spatial and temporal variations in hemodynamic forces initiate cardiac trabeculation. JCI Insight 2018, 3, e96672. [Google Scholar] [CrossRef] [Green Version]
- Samady, H.; Eshtehardi, P.; McDaniel, M.C.; Suo, J.; Dhawan, S.S.; Maynard, C.; Timmins, L.H.; Quyyumi, A.A.; Giddens, D.P. Coronary Artery Wall Shear Stress Is Associated With Progression and Transformation of Atherosclerotic Plaque and Arterial Remodeling in Patients With Coronary Artery Disease. Circulation 2011, 124, 779–788. [Google Scholar] [CrossRef] [Green Version]
- Matsui, H.; Germanakis, I.; Kulinskaya, E.; Gardiner, H.M. Temporal and spatial performance of vector velocity imaging in the human fetal heart. Ultrasound Obstet. Gynecol. 2011, 37, 150–157. [Google Scholar] [CrossRef]
- Paladini, D.; Lamberti, A.; Teodoro, A.; Arienzo, M.; Tartaglione, A.; Martinelli, P. Tissue Doppler imaging of the fetal heart. Ultrasound Obstet. Gynecol. 2000, 16, 530–535. [Google Scholar] [CrossRef]
- Norwood, W.I.; Lang, P.; Hansen, D.D. Physiologic Repair of Aortic Atresia-Hypoplastic Left Heart Syndrome. N. Engl. J. Med. 1983, 308, 23–26. [Google Scholar] [CrossRef]
- Tworetzky, W.; Wilkins-Haug, L.; Jennings, R.W.; van der Velde, M.E.; Marshall, A.C.; Marx, G.R.; Colan, S.D.; Benson, C.B.; Lock, J.E.; Perry, S.B. Balloon Dilation of Severe Aortic Stenosis in the Fetus. Circulation 2004, 110, 2125–2131. [Google Scholar] [CrossRef] [Green Version]
- Pickard, S.S.; Wong, J.B.; Bucholz, E.M.; Newburger, J.W.; Tworetzky, W.; la Franchi, T.; Benson, C.B.; Wilkins-Haug, L.E.; Porras, D.; Callahan, R.; et al. Fetal Aortic Valvuloplasty for Evolving Hypoplastic Left Heart Syndrome. Circ. Cardiovasc. Qual. Outcomes 2020, 13, e006127. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Klena, N.T.; Gabriel, G.C.; Liu, X.; Kim, A.J.; Lemke, K.; Chen, Y.; Chatterjee, B.; Devine, W.; Damerla, R.R.; et al. Global genetic analysis in mice unveils central role for cilia in congenital heart disease. Nature 2015, 521, 520–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henning, A.L.; Jiang, M.X.; Yalcin, H.C.; Butcher, J.T. Quantitative three-dimensional imaging of live avian embryonic morphogenesis via micro-computed tomography. Dev. Dyn. 2011, 240, 1949–1957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yalcin, H.C. Femtosecond laser photodisruption of vitelline vessels of avian embryos as a technique to study embryonic vascular remodeling. Exp. Biol. Med. 2014, 239, 1644–1652. [Google Scholar] [CrossRef]
- Yalcin, H.C.; Shekhar, A.; Nishimura, N.; Rane, A.A.; Schaffer, C.B.; Butcher, J.T. Two-photon microscopy-guided femtosecond-laser photoablation of avian cardiogenesis: Noninvasive creation of localized heart defects. Am. J. Physiol. Heart Circ. Physiol. 2010, 299, H1728–H1735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zakaria, Z.Z.; Benslimane, F.M.; Nasrallah, G.K.; Shurbaji, S.; Younes, N.N.; Mraiche, F.; Da’As, S.I.; Yalcin, H.C. Using Zebrafish for Investigating the Molecular Mechanisms of Drug-Induced Cardiotoxicity. BioMed Res. Int. 2018, 2018, 1642684. [Google Scholar] [CrossRef]
- Benslimane, F.M.; Alser, M.; Zakaria, Z.Z.; Sharma, A.; Abdelrahman, H.A.; Yalcin, H.C. Adaptation of a Mice Doppler Echocardiography Platform to Measure Cardiac Flow Velocities for Embryonic Chicken and Adult Zebrafish. Front. Bioeng. Biotechnol. 2019, 7, 96. [Google Scholar] [CrossRef]
- Shrestha, R.; Lieberth, J.; Tillman, S.; Natalizio, J.; Bloomekatz, J. Using Zebrafish to Analyze the Genetic and Environmental Etiologies of Congenital Heart Defects. Adv. Exp. Med. Biol. 2020, 1236, 189–223. [Google Scholar] [CrossRef]
- Sassen, W.A.; Köster, R.W. A molecular toolbox for genetic manipulation of zebrafish. Adv. Genom. Genet. 2015, 5, 151–163. [Google Scholar] [CrossRef] [Green Version]
- Yalcin, H.C.; Amindari, A.; Butcher, J.T.; Althani, A.; Yacoub, M. Heart function and hemodynamic analysis for zebrafish embryos. Dev. Dyn. 2017, 246, 868–880. [Google Scholar] [CrossRef] [Green Version]
- Vedula, V.; Lee, J.; Xu, H.; Kuo, C.C.J.; Hsiai, T.K.; Marsden, A.L. A method to quantify mechanobiologic forces during zebrafish cardiac development using 4-D light sheet imaging and computational modeling. PLoS Comput. Biol. 2017, 13, e1005828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bharadwaj, K.N.; Spitz, C.; Shekhar, A.; Yalcin, H.C.; Butcher, J.T. Computational Fluid Dynamics of Developing Avian Outflow Tract Heart Valves. Ann. Biomed. Eng. 2012, 40, 2212–2227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salman, H.E.; Ramazanli, B.; Yavuz, M.M.; Yalcin, H.C. Biomechanical Investigation of Disturbed Hemodynamics-Induced Tissue Degeneration in Abdominal Aortic Aneurysms Using Computational and Experimental Techniques. Front. Bioeng. Biotechnol. 2019, 7, 111. [Google Scholar] [CrossRef] [PubMed]
- Bathe, K.-J.; Hou, Z.; Ji, S. Finite element analysis of fluid flows fully coupled with structural interactions. Comput. Struct. 1999, 72, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Wiputra, H.; Chen, C.K.; Talbi, E.; Lim, G.L.; Soomar, S.M.; Biswas, A.; Mattar, C.N.Z.; Bark, D.; Leo, H.L.; Yap, C.H. Human fetal hearts with tetralogy of Fallot have altered fluid dynamics and forces. Am. J. Physiol. Heart Circ. Physiol. 2018, 315, H1649–H1659. [Google Scholar] [CrossRef]
- Elahi, S.; Blackburn, B.J.; Lapierre-Landry, M.; Gu, S.; Rollins, A.M.; Jenkins, M.W. Semi-automated shear stress measurements in developing embryonic hearts. Biomed. Opt. Express 2020, 11, 5297–5305. [Google Scholar] [CrossRef]
- Lashkarinia, S.S.; Çoban, G.; Ermek, E.; Çelik, M.; Pekkan, K. Spatiotemporal remodeling of embryonic aortic arch: Stress distribution, microstructure, and vascular growth in silico. Biomech. Model. Mechanobiol. 2020, 19, 1897–1915. [Google Scholar] [CrossRef]
- Courchaine, K.; Gray, M.J.; Beel, K.; Thornburg, K.; Rugonyi, S. 4-D Computational Modeling of Cardiac Outflow Tract Hemodynamics over Looping Developmental Stages in Chicken Embryos. J. Cardiovasc. Dev. Dis. 2019, 6, 11. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Garcia, M.J.; Doiron, A.L.; Steele, R.R.M.; Labouta, H.I.; Vafadar, B.; Shepherd, R.D.; Gates, I.D.; Cramb, D.T.; Childs, S.J.; Rinker, K.D. Nanoparticle localization in blood vessels: Dependence on fluid shear stress, flow disturbances, and flow-induced changes in endothelial physiology. Nanoscale 2018, 10, 15249–15261. [Google Scholar] [CrossRef]
- Wiputra, H.; Lai, C.Q.; Lim, G.L.; Heng, J.J.W.; Guo, L.; Soomar, S.M.; Leo, H.L.; Biwas, A.; Mattar, C.N.Z.; Yap, C.H.; et al. Fluid mechanics of human fetal right ventricles from image-based computational fluid dynamics using 4D clinical ultrasound scans. Am. J. Physiol. Heart Circ. Physiol. 2016, 311, H1498–H1508. [Google Scholar] [CrossRef] [Green Version]
- Lai, C.Q.; Lim, G.L.; Jamil, M.; Mattar, C.N.Z.; Biswas, A.; Yap, C.H. Fluid mechanics of blood flow in human fetal left ventricles based on patient-specific 4D ultrasound scans. Biomech. Model. Mechanobiol. 2016, 15, 1159–1172. [Google Scholar] [CrossRef] [PubMed]
- Scotti, C.M.; Finol, E.A. Compliant biomechanics of abdominal aortic aneurysms: A fluid–structure interaction study. Comput. Struct. 2007, 85, 1097–1113. [Google Scholar] [CrossRef]
- Scotti, C.M.; Jimenez, J.; Muluk, S.C.; Finol, E.A. Wall stress and flow dynamics in abdominal aortic aneurysms: Finite element analysis vs. fluid–structure interaction. Comput. Methods Biomech. Biomed. Eng. 2008, 11, 301–322. [Google Scholar] [CrossRef] [PubMed]
- Wolters, B.J.B.M.; Rutten, M.C.M.; Schurink, G.W.H.; Kose, U.; de Hart, J.; van de Vosse, F.N. A patient-specific computational model of fluid–structure interaction in abdominal aortic aneurysms. Med Eng. Phys. 2005, 27, 871–883. [Google Scholar] [CrossRef] [PubMed]
- Goenezen, S.; Chivukula, V.K.; Midgett, M.; Phan, L.; Rugonyi, S. 4D subject-specific inverse modeling of the chick embryonic heart outflow tract hemodynamics. Biomech. Model. Mechanobiol. 2016, 15, 723–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amindari, A.; Saltik, L.; Kirkkopru, K.; Yacoub, M.; Yalcin, H.C. Assessment of calcified aortic valve leaflet deformations and blood flow dynamics using fluid-structure interaction modeling. Inform. Med. Unlocked 2017, 9, 191–199. [Google Scholar] [CrossRef]
- Wiputra, H.; Lim, G.L.; Chia, D.A.K.; Mattar, C.N.Z.; Biswas, A.; Yap, C.H. Methods for fluid dynamics simulations of human fetal cardiac chambers based on patient-specific 4D ultrasound scans. J. Biomech. Sci. Eng. 2016, 11, 15-00608. [Google Scholar] [CrossRef]
- De Hart, J.; Peters, G.W.M.; Schreurs, P.J.G.; Baaijens, F.P.T. A three-dimensional computational analysis of fluid–structure interaction in the aortic valve. J. Biomech. 2003, 36, 103–112. [Google Scholar] [CrossRef]
- Ene, F.; DeLassus, P.; Morris, L. The influence of computational assumptions on analysing abdominal aortic aneurysm haemodynamics. Proc. Inst. Mech. Eng. Part H J. Eng. Med. 2014, 228, 768–780. [Google Scholar] [CrossRef]
- Kelsey, L.J.; Powell, J.T.; Norman, P.E.; Miller, K.; Doyle, B.J. A comparison of hemodynamic metrics and intraluminal thrombus burden in a common iliac artery aneurysm. Int. J. Numer. Methods Biomed. Eng. 2017, 33, e2821. [Google Scholar] [CrossRef] [Green Version]
- Di Achille, P.; Tellides, G.; Humphrey, J.D. Hemodynamics-driven deposition of intraluminal thrombus in abdominal aortic aneurysms. Int. J. Numer. Methods Biomed. Eng. 2017, 33, e2828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandra, S.; Raut, S.S.; Jana, A.; Biederman, R.W.; Doyle, M.; Muluk, S.C.; Finol, E.A. Fluid-Structure Interaction Modeling of Abdominal Aortic Aneurysms: The Impact of Patient-Specific Inflow Conditions and Fluid/Solid Coupling. J. Biomech. Eng. 2013, 135, 081001–08100114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khanafer, K.M.; Gadhoke, P.; Berguer, R.; Bull, J.L. Modeling pulsatile flow in aortic aneurysms: Effect of non-Newtonian properties of blood. Biorheology 2006, 43, 661–679. [Google Scholar] [PubMed]
- Al-Roubaie, S.; Jahnsen, E.D.; Mohammed, M.; Henderson-Toth, C.; Jones, E.A.V. Rheology of embryonic avian blood. Am. J. Physiol. Heart Circ. Physiol. 2011, 301, H2473–H2481. [Google Scholar] [CrossRef] [Green Version]
- Cho, Y.I.; Kensey, K.R. Effects of the non-Newtonian viscosity of blood on flows in a diseased arterial vessel. Part 1: Steady flows. Biorheology 1991, 28, 241–262. [Google Scholar] [CrossRef]
- Arzani, A. Accounting for residence-time in blood rheology models: Do we really need non-Newtonian blood flow modelling in large arteries? J. R. Soc. Interface 2018, 15, 20180486. [Google Scholar] [CrossRef] [Green Version]
- Thurston, G.B. Rheological parameters for the viscosity viscoelasticity and thixotropy of blood. Biorheology 1979, 16, 149–162. [Google Scholar] [CrossRef]
- Eckhardt, B.; Schneider, T.M.; Hof, B.; Westerweel, J. Turbulence Transition in Pipe Flow. Annu. Rev. Fluid Mech. 2006, 39, 447–468. [Google Scholar] [CrossRef] [Green Version]
- Salman, H.E.; Sert, C.; Yazicioglu, Y. Computational analysis of high frequency fluid–structure interactions in constricted flow. Comput. Struct. 2013, 122, 145–154. [Google Scholar] [CrossRef]
- Varghese, S.S.; Frankel, S.H. Numerical Modeling of Pulsatile Turbulent Flow in Stenotic Vessels. J. Biomech. Eng. 2003, 125, 445–460. [Google Scholar] [CrossRef]
- Menter, F.R. Two-equation eddy-viscosity turbulence models for engineering applications. AIAA J. 1994, 32, 1598–1605. [Google Scholar] [CrossRef] [Green Version]
- Les, A.S.; Shadden, S.C.; Figueroa, C.A.; Park, J.M.; Tedesco, M.M.; Herfkens, R.J.; Dalman, R.L.; Taylor, C.A. Quantification of Hemodynamics in Abdominal Aortic Aneurysms During Rest and Exercise Using Magnetic Resonance Imaging and Computational Fluid Dynamics. Ann. Biomed. Eng. 2010, 38, 1288–1313. [Google Scholar] [CrossRef] [PubMed]
- Arzani, A.; Suh, G.-Y.; Dalman, R.L.; Shadden, S.C. A longitudinal comparison of hemodynamics and intraluminal thrombus deposition in abdominal aortic aneurysms. Am. J. Physiol. Heart Circ. Physiol. 2014, 307, H1786–H1795. [Google Scholar] [CrossRef] [PubMed]
- Salman, H.E.; Yazicioglu, Y. Experimental and numerical investigation on soft tissue dynamic response due to turbulence-induced arterial vibration. Med. Biol. Eng. Comput. 2019, 57, 1737–1752. [Google Scholar] [CrossRef] [PubMed]
- Mooney, M. A Theory of Large Elastic Deformation. J. Appl. Phys. 1940, 11, 582–592. [Google Scholar] [CrossRef]
- Rivlin, R.S.; Saunders, D.W.; Andrade, E.N.D.C. Large elastic deformations of isotropic materials VII. Experiments on the deformation of rubber. Philos. Trans. R. Soc. Lond. Ser. A Math. Phys. Sci. 1951, 243, 251–288. [Google Scholar] [CrossRef]
- Ogden, R.W.; Hill, R. Large deformation isotropic elasticity—On the correlation of theory and experiment for incompressible rubberlike solids. Proc. R. Soc. Lond. A Math. Phys. Sci. 1972, 326, 565–584. [Google Scholar] [CrossRef]
- Salman, H.E.; Yalcin, H.C. Advanced blood flow assessment in Zebrafish via experimental digital particle image velocimetry and computational fluid dynamics modeling. Micron 2020, 130, 102801. [Google Scholar] [CrossRef]
- Deng, L.; Huang, X.; Yang, C.; Song, Y.; Tang, D. Patient-specific CT-based 3D passive FSI model for left ventricle in hypertrophic obstructive cardiomyopathy. Comput. Methods Biomech. Biomed. Eng. 2018, 21, 255–263. [Google Scholar] [CrossRef]
- Arzani, A.; Shadden, S.C. Characterizations and Correlations of Wall Shear Stress in Aneurysmal Flow. J. Biomech. Eng. 2015, 138. [Google Scholar] [CrossRef] [Green Version]
- Arzani, A.; Gambaruto, A.M.; Chen, G.; Shadden, S.C. Wall shear stress exposure time: A Lagrangian measure of near-wall stagnation and concentration in cardiovascular flows. Biomech. Model. Mechanobiol. 2017, 16, 787–803. [Google Scholar] [CrossRef] [PubMed]
- Roache, P.J. Discussion: “Uncertainties and CFD Code Validation” (Coleman, H.W. and Stern, F., 1997, ASME J. Fluids Eng., 119, pp. 795–803). J. Fluids Eng. 1998, 120, 635. [Google Scholar] [CrossRef] [Green Version]
- Lin, G.; Wan, X.; Su, C.; Karniadakis, G.E. Stochastic Computational Fluid Mechanics. Comput. Sci. Eng. 2007, 9, 21–29. [Google Scholar] [CrossRef] [Green Version]
- Richmond, M.; Kolios, A.; Pillai, V.S.; Nishino, T.; Wang, L. Development of a stochastic computational fluid dynamics approach for offshore wind farms. J. Phys. Conf. Ser. 2018, 1037, 072034. [Google Scholar] [CrossRef]
- Bozzi, S.; Morbiducci, U.; Gallo, D.; Ponzini, R.; Rizzo, G.; Bignardi, C.; Passoni, G. Uncertainty propagation of phase contrast-MRI derived inlet boundary conditions in computational hemodynamics models of thoracic aorta. Comput. Methods Biomech. Biomed. Eng. 2017, 20, 1104–1112. [Google Scholar] [CrossRef]
- Tran, J.S.; Schiavazzi, D.E.; Ramachandra, A.B.; Kahn, A.M.; Marsden, A.L. Automated tuning for parameter identification and uncertainty quantification in multi-scale coronary simulations. Comput. Fluids 2017, 142, 128–138. [Google Scholar] [CrossRef] [Green Version]
- Boccadifuoco, A.M.A.; Simona, C.; Nicola, M.; Salvetti, M.V. Uncertainty quantification in numerical simulations of the flow in thoracic aortic aneurysms. In Proceedings of the VII European Congress on Computational Methods in Applied Sciences and Engineering, Crete Island, Greece, 5–10 June 2016. [Google Scholar]
- Boccadifuoco, A.; Mariotti, A.; Celi, S.; Martini, N.; Salvetti, M.V. Impact of uncertainties in outflow boundary conditions on the predictions of hemodynamic simulations of ascending thoracic aortic aneurysms. Comput. Fluids 2018, 165, 96–115. [Google Scholar] [CrossRef]
- Sankaran, S.; Marsden, A.L. A Stochastic Collocation Method for Uncertainty Quantification and Propagation in Cardiovascular Simulations. J. Biomech. Eng. 2011, 133. [Google Scholar] [CrossRef]
- Boccadifuoco, A.; Mariotti, A.; Capellini, K.; Celi, S.; Salvetti, M.V. Validation of Numerical Simulations of Thoracic Aorta Hemodynamics: Comparison with In Vivo Measurements and Stochastic Sensitivity Analysis. Cardiovasc. Eng. Technol. 2018, 9, 688–706. [Google Scholar] [CrossRef]
- Midgett, M.; Rugonyi, S. Congenital heart malformations induced by hemodynamic altering surgical interventions. Front. Physiol. 2014, 5, 287. [Google Scholar] [CrossRef] [Green Version]
- Gregg, C.L.; Butcher, J.T. Quantitative in vivo imaging of embryonic development: Opportunities and challenges. Differentiation 2012, 84, 149–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kowalski, W.J.; Pekkan, K.; Tinney, J.P.; Keller, B.B. Investigating developmental cardiovascular biomechanics and the origins of congenital heart defects. Front. Physiol. 2014, 5, 408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Midgett, M.; Chivukula, V.K.; Dorn, C.; Wallace, S.; Rugonyi, S. Blood flow through the embryonic heart outflow tract during cardiac looping in HH13–HH18 chicken embryos. J. R. Soc. Interface 2015, 12, 20150652. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.H.; Ma, Y.S.; Zhao, Y.Q.; Liu, J.; Liu, J.H.; Lv, J.T.; Wang, Y. Measurement of the absolute velocity of blood flow in early-stage chick embryos using spectral domain optical coherence tomography. Appl. Opt. 2017, 56, 8832–8837. [Google Scholar] [CrossRef]
- Yalcin, H.C.; Shekhar, A.; Rane, A.A.; Butcher, J.T. An ex-ovo Chicken Embryo Culture System Suitable for Imaging and Microsurgery Applications. J. Vis. Exp. 2010, 10, e2154. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, M.W.; Adler, D.C.; Gargesha, M.; Huber, R.; Rothenberg, F.; Belding, J.; Watanabe, M.; Wilson, D.L.; Fujimoto, J.G.; Rollins, A.M. Ultrahigh-speed optical coherence tomography imaging and visualization of the embryonic avian heart using a buffered Fourier Domain Mode Locked laser. Opt. Express 2007, 15, 6251–6267. [Google Scholar] [CrossRef] [Green Version]
- Yelbuz, T.M.; Choma, M.A.; Thrane, L.; Kirby, M.L.; Izatt, J.A. Optical Coherence Tomography. Circulation 2002, 106, 2771–2774. [Google Scholar] [CrossRef] [Green Version]
- Rugonyi, S.; Shaut, C.; Liu, A.; Thornburg, K.; Wang, R.K. Changes in wall motion and blood flow in the outflow tract of chick embryonic hearts observed with optical coherence tomography after outflow tract banding and vitelline-vein ligation. Phys. Med. Biol. 2008, 53, 5077–5091. [Google Scholar] [CrossRef]
- Ma, Z.; Dou, S.; Zhao, Y.; Guo, C.; Liu, J.; Wang, Q.; Xu, T.; Wang, R.K.; Wang, Y. In vivo assessment of wall strain in embryonic chick heart by spectral domain optical coherence tomography. Appl. Opt. 2015, 54, 9253–9257. [Google Scholar] [CrossRef]
- Phan, L.; Grimm, C.; Rugonyi, S. Visualization Techniques for the Developing Chicken Heart. In Proceedings of Advances in Visual Computing; Springer: Cham, Switzerland, 2015; pp. 35–44. [Google Scholar]
- Tan, G.X.Y.; Jamil, M.; Tee, N.G.Z.; Zhong, L.; Yap, C.H. 3D Reconstruction of Chick Embryo Vascular Geometries Using Non-invasive High-Frequency Ultrasound for Computational Fluid Dynamics Studies. Ann. Biomed. Eng. 2015, 43, 2780–2793. [Google Scholar] [CrossRef]
- Oosterbaan, A.M.; Ursem, N.T.C.; Struijk, P.C.; Bosch, J.G.; van der Steen, A.F.W.; Steegers, E.A.P. Doppler flow velocity waveforms in the embryonic chicken heart at developmental stages corresponding to 5-8 weeks of human gestation. Ultrasound Obstet. Gynecol. 2009, 33, 638–644. [Google Scholar] [CrossRef] [PubMed]
- Peterson, L.M.; Jenkins, M.W.; Gu, S.; Barwick, L.; Watanabe, M.; Rollins, A.M. 4D shear stress maps of the developing heart using Doppler optical coherence tomography. Biomed. Opt. Express 2012, 3, 3022–3032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yalcin, H.C.; Shekhar, A.; McQuinn, T.C.; Butcher, J.T. Hemodynamic patterning of the avian atrioventricular valve. Dev. Dyn. 2011, 240, 23–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, A.; Rugonyi, S.; Pentecost, J.O.; Thornburg, K.L. Finite element modeling of blood flow-induced mechanical forces in the outflow tract of chick embryonic hearts. Comput. Struct. 2007, 85, 727–738. [Google Scholar] [CrossRef]
- Liu, A.; Nickerson, A.; Troyer, A.; Yin, X.; Cary, R.; Thornburg, K.; Wang, R.; Rugonyi, S. Quantifying blood flow and wall shear stresses in the outflow tract of chick embryonic hearts. Comput. Struct. 2011, 89, 855–867. [Google Scholar] [CrossRef] [Green Version]
- Ho, S.; Chan, W.X.; Rajesh, S.; Phan-Thien, N.; Yap, C.H. Fluid dynamics and forces in the HH25 avian embryonic outflow tract. Biomech. Model. Mechanobiol. 2019, 18, 1123–1137. [Google Scholar] [CrossRef]
- Celik, M.; Goktas, S.; Karakaya, C.; Cakiroglu, A.I.; Karahuseyinoglu, S.; Lashkarinia, S.S.; Ermek, E.; Pekkan, K. Microstructure of early embryonic aortic arch and its reversibility following mechanically altered hemodynamic load release. Am. J. Physiol. Heart Circ. Physiol. 2020, 318, H1208–H1218. [Google Scholar] [CrossRef]
- Gould, R.A.; Yalcin, H.C.; Mackay, J.L.; Sauls, K.; Norris, R.; Kumar, S.; Butcher, J.T. Cyclic Mechanical Loading Is Essential for Rac1-Mediated Elongation and Remodeling of the Embryonic Mitral Valve. Curr. Biol. 2016, 26, 27–37. [Google Scholar] [CrossRef] [Green Version]
- Martinsen, B.J. Reference guide to the stages of chick heart embryology. Dev. Dyn. 2005, 233, 1217–1237. [Google Scholar] [CrossRef]
- Wittig, J.G.; Münsterberg, A. The Early Stages of Heart Development: Insights from Chicken Embryos. J. Cardiovasc. Dev. Dis. 2016, 3, 12. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Dur, O.; Patrick, M.J.; Tinney, J.P.; Tobita, K.; Keller, B.B.; Pekkan, K. Aortic Arch Morphogenesis and Flow Modeling in the Chick Embryo. Ann. Biomed. Eng. 2009, 37, 1069–1081. [Google Scholar] [CrossRef] [PubMed]
- Liu, A.; Yin, X.; Shi, L.; Li, P.; Thornburg, K.L.; Wang, R.; Rugonyi, S. Biomechanics of the Chick Embryonic Heart Outflow Tract at HH18 Using 4D Optical Coherence Tomography Imaging and Computational Modeling. PLoS ONE 2012, 7, e40869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, S.; Tan, G.X.Y.; Foo, T.J.; Phan-Thien, N.; Yap, C.H. Organ Dynamics and Fluid Dynamics of the HH25 Chick Embryonic Cardiac Ventricle as Revealed by a Novel 4D High-Frequency Ultrasound Imaging Technique and Computational Flow Simulations. Ann. Biomed. Eng. 2017, 45, 2309–2323. [Google Scholar] [CrossRef] [PubMed]
- Kowalski, W.J.; Teslovich, N.C.; Menon, P.G.; Tinney, J.P.; Keller, B.B.; Pekkan, K. Left atrial ligation alters intracardiac flow patterns and the biomechanical landscape in the chick embryo. Dev. Dyn. 2014, 243, 652–662. [Google Scholar] [CrossRef]
- Lindsey, S.E.; Menon, P.G.; Kowalski, W.J.; Shekhar, A.; Yalcin, H.C.; Nishimura, N.; Schaffer, C.B.; Butcher, J.T.; Pekkan, K. Growth and hemodynamics after early embryonic aortic arch occlusion. Biomech. Model. Mechanobiol. 2015, 14, 735–751. [Google Scholar] [CrossRef] [Green Version]
- Lindsey, S.E.; Butcher, J.T.; Vignon-Clementel, I.E. Cohort-based multiscale analysis of hemodynamic-driven growth and remodeling of the embryonic pharyngeal arch arteries. Development 2018, 145, dev162578. [Google Scholar] [CrossRef] [Green Version]
- Kowalski, W.J.; Teslovich, N.C.; Dur, O.; Keller, B.B.; Pekkan, K. Computational hemodynamic optimization predicts dominant aortic arch selection is driven by embryonic outflow tract orientation in the chick embryo. Biomech. Model. Mechanobiol. 2012, 11, 1057–1073. [Google Scholar] [CrossRef]
- Midgett, M.; Goenezen, S.; Rugonyi, S. Blood flow dynamics reflect degree of outflow tract banding in Hamburger–Hamilton stage 18 chicken embryos. J. R. Soc. Interface 2014, 11, 20140643. [Google Scholar] [CrossRef]
- Menon, V.; Eberth, J.F.; Goodwin, R.L.; Potts, J.D. Altered Hemodynamics in the Embryonic Heart Affects Outflow Valve Development. J. Cardiovasc. Dev. Dis. 2016, 2, 108. [Google Scholar] [CrossRef]
- Chivukula, V.K.; Goenezen, S.; Liu, A.; Rugonyi, S. Effect of Outflow Tract Banding on Embryonic Cardiac Hemodynamics. J. Cardiovasc. Dev. Dis. 2016, 3, 1. [Google Scholar] [CrossRef]
- Eisa-Beygi, S.; Benslimane, F.M.; El-Rass, S.; Prabhudesai, S.; Abdelrasoul, M.K.A.; Simpson, P.M.; Yalcin, H.C.; Burrows, P.E.; Ramchandran, R. Characterization of Endothelial Cilia Distribution During Cerebral-Vascular Development in Zebrafish (Danio rerio). Arter. Thromb. Vasc. Biol. 2018, 38, 2806–2818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stainier, D.Y.; Kontarakis, Z.; Rossi, A. Making Sense of Anti-Sense Data. Dev. Cell 2015, 32, 7–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glickman, N.S.; Yelon, D. Cardiac development in zebrafish: Coordination of form and function. Semin. Cell Dev. Biol. 2002, 13, 507–513. [Google Scholar] [CrossRef] [PubMed]
- Poon, K.L.; Brand, T. The zebrafish model system in cardiovascular research: A tiny fish with mighty prospects. Glob. Cardiol. Sci. Pract. 2013, 2013, 9–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beis, D.; Bartman, T.; Jin, S.-W.; Scott, I.C.; Amico, L.A.; Ober, E.A.; Verkade, H.; Frantsve, J.; Field, H.A.; Wehman, A.; et al. Genetic and cellular analyses of zebrafish atrioventricular cushion and valve development. Development 2005, 132, 4193–4204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, R.T.; Bartman, T. Analysis of heart valve development in larval zebrafish. Dev. Dyn. 2009, 238, 1796–1802. [Google Scholar] [CrossRef] [PubMed]
- Benslimane, F.M.; Zakaria, Z.Z.; Shurbaji, S.; Abdelrasool, M.K.A.; Al-Badr, M.A.H.I.; Al Absi, E.S.K.; Yalcin, H.C. Cardiac function and blood flow hemodynamics assessment of zebrafish (Danio rerio) using high-speed video microscopy. Micron 2020, 136, 102876. [Google Scholar] [CrossRef]
- Yalcin, H.C. Hemodynamic Studies for Analyzing the Teratogenic Effects of Drugs in the Zebrafish Embryo. Methods Mol. Biol. 2018, 1797, 487–495. [Google Scholar] [CrossRef]
- Jamison, R.A.; Samarage, C.R.; Bryson-Richardson, R.J.; Fouras, A. In Vivo Wall Shear Measurements within the Developing Zebrafish Heart. PLoS ONE 2013, 8, e75722. [Google Scholar] [CrossRef]
- Bagatto, B.; Burggren, W. A Three-Dimensional Functional Assessment of Heart and Vessel Development in the Larva of the Zebrafish (Danio rerio). Physiol. Biochem. Zoöl. 2006, 79, 194–201. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Fei, P.; Packard, R.R.S.; Kang, H.; Xu, H.; Baek, K.I.; Jen, N.; Chen, J.; Yen, H.; Kuo, C.-C.J.; et al. 4-Dimensional light-sheet microscopy to elucidate shear stress modulation of cardiac trabeculation. J. Clin. Investig. 2016, 126, 1679–1690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trivedi, V.; Truong, T.V.; Trinh, L.A.; Holland, D.B.; Liebling, M.; Fraser, S.E. Dynamic structure and protein expression of the live embryonic heart captured by 2-photon light sheet microscopy and retrospective registration. Biomed. Opt. Express 2015, 6, 2056–2066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, M.; Huisken, J. Light sheet microscopy for real-time developmental biology. Curr. Opin. Genet. Dev. 2011, 21, 566–572. [Google Scholar] [CrossRef] [PubMed]
- Boselli, F.; Vermot, J. Live imaging and modeling for shear stress quantification in the embryonic zebrafish heart. Methods 2016, 94, 129–134. [Google Scholar] [CrossRef]
- Sidhwani, P.; Yelon, D. Chapter Eleven—Fluid forces shape the embryonic heart: Insights from zebrafish. In Current Topics in Developmental Biology; Wellik, D.M., Ed.; Academic Press: Cambridge, MA, USA, 2019; Volume 132, pp. 395–416. [Google Scholar]
- Hove, J.R. Quantifying Cardiovascular Flow Dynamics During Early Development. Pediatr. Res. 2006, 60, 6–13. [Google Scholar] [CrossRef] [Green Version]
- Hu, N.; Yost, H.J.; Clark, E.B. Cardiac morphology and blood pressure in the adult zebrafish. Anat. Rec. 2001, 264, 1–12. [Google Scholar] [CrossRef]
- Peshkovsky, C.; Totong, R.; Yelon, D. Dependence of cardiac trabeculation on neuregulin signaling and blood flow in zebrafish. Dev. Dyn. 2011, 240, 446–456. [Google Scholar] [CrossRef]
- Battista, N.A.; Douglas, D.R.; Lane, A.N.; Samsa, L.A.; Liu, J.; Miller, L.A. Vortex Dynamics in Trabeculated Embryonic Ventricles. J. Cardiovasc. Dev. Dis. 2019, 6, 6. [Google Scholar] [CrossRef] [Green Version]
- Foo, Y.Y.; Pant, S.; Tay, H.S.; Imangali, N.; Chen, N.; Winkler, C.; Yap, C.H. 4D modelling of fluid mechanics in the zebrafish embryonic heart. Biomech. Model. Mechanobiol. 2020, 19, 221–232. [Google Scholar] [CrossRef]
- Lee, J.; Moghadam, M.E.; Kung, E.; Cao, H.; Beebe, T.; Miller, Y.; Roman, B.L.; Lien, C.-L.; Chi, N.C.; Marsden, A.L.; et al. Moving Domain Computational Fluid Dynamics to Interface with an Embryonic Model of Cardiac Morphogenesis. PLoS ONE 2013, 8, e72924. [Google Scholar] [CrossRef] [Green Version]
- Miller, L.A. Fluid Dynamics of Ventricular Filling in the Embryonic Heart. Cell Biochem. Biophys. 2011, 61, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Hove, J.R.; Köster, R.W.; Forouhar, A.S.; Acevedo-Bolton, G.; Fraser, S.E.; Gharib, M. Intracardiac fluid forces are an essential epigenetic factor for embryonic cardiogenesis. Nature 2003, 421, 172–177. [Google Scholar] [CrossRef] [PubMed]
- Boselli, F.; Steed, E.; Freund, J.B.; Vermot, J. Anisotropic shear stress patterns predict the orientation of convergent tissue movements in the embryonic heart. Development 2017, 144, 4322–4327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bombardini, T.; Gemignani, V.; Bianchini, E.; Venneri, L.; Petersen, C.; Pasanisi, E.; Pratali, L.; Alonso-Rodriguez, D.; Pianelli, M.; Faita, F.; et al. Diastolic time—Frequency relation in the stress echo lab: Filling timing and flow at different heart rates. Cardiovasc. Ultrasound 2008, 6, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Zhou, Y.; Wang, J.; Liu, X.; Ge, S.; He, Y. Modeling of coarctation of aorta in human fetuses using 3D/4D fetal echocardiography and computational fluid dynamics. Echocardiography 2017, 34, 1858–1866. [Google Scholar] [CrossRef] [PubMed]
- Sundareswaran, K.S.; Zelicourt, D.D.; Pekkan, K.; Jayaprakash, G.; Kim, D.; Whited, B.; Rossignac, J.; Fogel, M.A.; Kanter, K.R.; Yoganathan, A.P. Anatomically Realistic Patient-Specific Surgical Planning of Complex Congenital Heart Defects Using MRI and CFD. In Proceedings of the 2007 29th Annual International Conference of the IEEE Engineering in Medicine and Biology Society, Lyon, France, 22–26 August 2007; pp. 202–205. [Google Scholar]
- Cibis, M.; Jarvis, K.; Markl, M.; Rose, M.; Rigsby, C.; Barker, A.J.; Wentzel, J.J. The effect of resolution on viscous dissipation measured with 4D flow MRI in patients with Fontan circulation: Evaluation using computational fluid dynamics. J. Biomech. 2015, 48, 2984–2989. [Google Scholar] [CrossRef] [Green Version]
- De Zélicourt, D.A.; Kurtcuoglu, V. Patient-Specific Surgical Planning, Where Do We Stand? The Example of the Fontan Procedure. Ann. Biomed. Eng. 2016, 44, 174–186. [Google Scholar] [CrossRef]
- Haggerty, C.M.; de Zélicourt, D.A.; Restrepo, M.; Rossignac, J.; Spray, T.L.; Kanter, K.R.; Fogel, M.A.; Yoganathan, A.P. Comparing Pre- and Post-operative Fontan Hemodynamic Simulations: Implications for the Reliability of Surgical Planning. Ann. Biomed. Eng. 2012, 40, 2639–2651. [Google Scholar] [CrossRef]
- Qian, Y.; Liu, J.L.; Itatani, K.; Miyaji, K.; Umezu, M. Computational Hemodynamic Analysis in Congenital Heart Disease: Simulation of the Norwood Procedure. Ann. Biomed. Eng. 2010, 38, 2302–2313. [Google Scholar] [CrossRef]
- Ceballos, A.; Divo, E.; Argueta-Morales, R.; Calderone, C.; Kassab, A.; DeCampli, W. A Multi-Scale CFD Analysis of the Hybrid Norwood Palliative Treatment for Hypoplastic Left Heart Syndrome: Effect of Reverse Blalock-Taussing Shunt Diameter. In Proceedings of the ASME 2013 International Mechanical Engineering Congress and Exposition, San Diego, CA, USA, 15–21 November 2013. [Google Scholar]
- Gundelwein, L.; Miró, J.; Barlatay, F.G.; Lapierre, C.; Rohr, K.; Duong, L. Personalized stent design for congenital heart defects using pulsatile blood flow simulations. J. Biomech. 2018, 81, 68–75. [Google Scholar] [CrossRef]
- Sotelo, J.A.; Urbina, J.; Valverde, I.; Tejos, C.; Irarrazaval, P.; Hurtado, D.E.; Uribe, S. 3D quantification of hemodynamics parameters of pulmonary artery and aorta using finite-element interpolations in 4D flow MR data. J. Cardiovasc. Magn. Reson. 2015, 17, Q27. [Google Scholar] [CrossRef] [Green Version]
- Biglino, G.; Capelli, C.; Bruse, J.; Bosi, G.M.; Taylor, A.M.; Schievano, S. Computational modelling for congenital heart disease: How far are we from clinical translation? Heart 2017, 103, 98–103. [Google Scholar] [CrossRef] [PubMed]
- Young, A.A.; Frangi, A.F. Computational cardiac atlases: From patient to population and back. Exp. Physiol. 2009, 94, 578–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Figueroa, C.A.; Baek, S.; Taylor, C.A.; Humphrey, J.D. A computational framework for fluid–solid-growth modeling in cardiovascular simulations. Comput. Methods Appl. Mech. Eng. 2009, 198, 3583–3602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marsden, A.L.; Feinstein, J.A. Computational modeling and engineering in pediatric and congenital heart disease. Curr. Opin. Pediatr. 2015, 27, 587–596. [Google Scholar] [CrossRef]
- Keller, B.B.; Kowalski, W.J.; Tinney, J.P.; Tobita, K.; Hu, N. Validating the Paradigm That Biomechanical Forces Regulate Embryonic Cardiovascular Morphogenesis and Are Fundamental in the Etiology of Congenital Heart Disease. J. Cardiovasc. Dev. Dis. 2020, 7, 23. [Google Scholar] [CrossRef]
- Olesen, S.-P.; Claphamt, D.; Davies, P. Haemodynamic shear stress activates a K+ current in vascular endothelial cells. Nature 1988, 331, 168–170. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
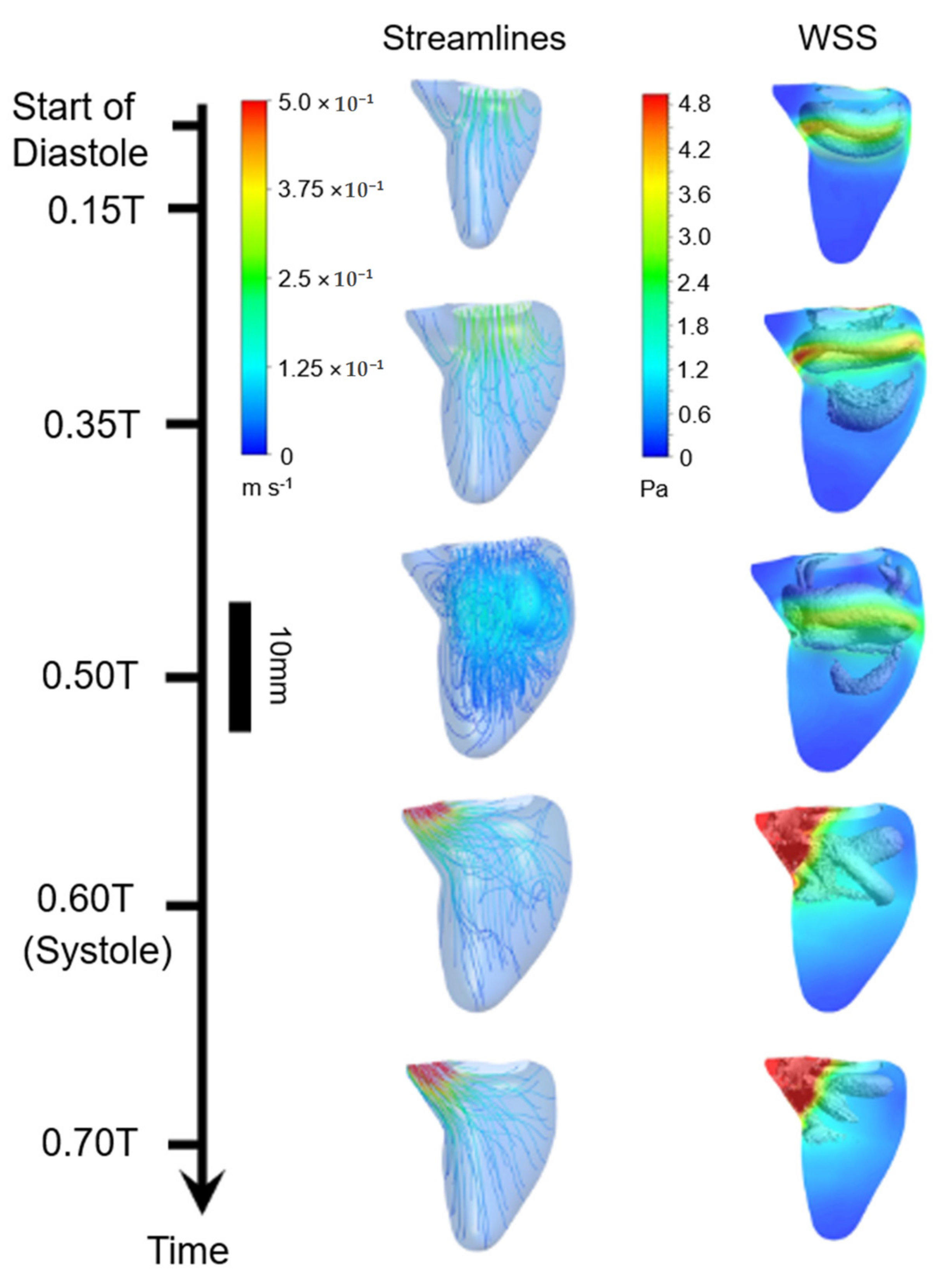{kind=link}
| Embryonic Stage–Investigated Region | Peak WSS (Dynes/cm2) | Spatially Averaged WSS at Peak Velocity (Dynes/cm2) | Spatially and Temporally Averaged WSS (Dynes/cm2) |
|---|---|---|---|
| HH16–OFT [32] | 18.16 (3.18) | 9.55 (0.40) | 3.03 (0.11) |
| HH17–AV canal [95] | 19.34 (4.45) | 9.17 (3.2) | 3.62 (0.32) |
| HH18–OFT [104] | 60 | ||
| HH18–Aortic Arch II left side [103] | - | 59.4 (14) | 17.9 (5) |
| HH18–Aortic Arch II right side [103] | - | 47.1 (12) | 14.3 (4) |
| HH18–Aortic Arch III left side [103] | - | 64.4 (18) | 19.4 (5) |
| HH18–Aortic Arch III right side [103] | - | 55.8 (17) | 17.0 (5) |
| HH18–Aortic Arch IV left side [103] | - | 41.7 (17) | 12.5 (5) |
| HH18–Aortic Arch IV right side [103] | - | 59.9 (18) | 18.2 (6) |
| HH21–OFT [96] | 31 | - | - |
| HH23–AV canal [95] | 78.33 (37.09) | 33.59 (16.84) | 6.79 (3.22) |
| HH23–Proximal OFT [32] | 59.36 (10.07) | 28.15 (7.47) | 4.23 (0.09) |
| HH23–Distal OFT [32] | 57.12 (9.04) | 31.69 (7.48) | 7.05 (0.82) |
| HH24–Aortic Arch III left side [103] | - | 104.9 (37) | 23.6 (8) |
| HH24–Aortic Arch III right side [103] | - | 82.2 (16) | 18.4 (4) |
| HH24–Aortic Arch IV left side [103] | - | 92.3 (56) | 20.8 (12) |
| HH24–Aortic Arch IV right side [103] | - | 122.9 (40) | 27.5 (9) |
| HH24–Aortic Arch VI left side [103] | - | 130.7 (28) | 29.3 (6) |
| HH24–Aortic Arch VI right side [103] | - | 40.4 (40) | 9.0 (9) |
| HH27–AV canal [95] | 250.09 (51.49) | 59.7 (4.6) | 6.1 (0.52) |
| HH27–OFT [32] | 236.07 (39.34) | 111.74 (21.7) | 39.49 (9.34) |
| HH27–Carotid arteries [92] | - | 4.60 | - |
| HH27–Pharyngeal aortic arches [92] | - | 18.90 | - |
| HH27–Left and right dorsal aortae [92] | - | 2.4 | - |
| HH27–Common dorsal aorta [92] | - | 7.3 | - |
| HH30–AV canal [95] | 287.18 (67.45) | 86.27 (8.6) | 9.11 (1.061) |
| HH30–RVOFT [32] | 671.24 (211.36) | 184.36 (34.26) | 100.67 (27.82) |
| HH30–LVOFT [32] | 400.93 (65.65) | 226.67 (20.41) | 136.5 (17.82) |
| Embryonic Stage–Investigated Region | Peak WSS (Dynes/cm2) |
|---|---|
| 20–30 hpf–AV canal [133] | 3.5 |
| 48 hpf–AV canal [126] | 70 |
| 40–50 hpf–AV canal [133] | 20 |
| 52 hpf–Ventral vein [39] | 3.4 |
| 60–70 hpf–AV canal [133] | 28 |
| 80–90 hpf–AV canal [133] | 58 |
| 108 hpf–AV canal [134] | 13.6 |
| 110–120 hpf–AV canal [133] | 82 |
| 120 hpf–Mid-ventricular segment [132] | 4–11 |
| 120 hpf–Ventricle inflow tract [132] | 130 |
| 120 hpf–Ventricle outflow tract [132] | 110 |
| Gestation Week–Heart Condition | Diastolic RV Peak WSS (Dynes/cm2) | Diastolic LV Peak WSS (Dynes/cm2) | Systolic RV Peak WSS (Dynes/cm2) | Systolic LV Peak WSS (Dynes/cm2) |
|---|---|---|---|---|
| 22 week–TOF-I [35] | 27.0 | 9.3 | 11.6 | 11.2 |
| 31 week–TOF-II [35] | 12.6 | 9.4 | 15.8 | 15.1 |
| 31 week–TOF-III [35] | 12.8 | 5.6 | 16.6 | 13.9 |
| 22 week–Normal [35] | 12.8 (6.1) | 7.4 (2.9) | 20.0 (2.8) | 15.1 (2.6) |
| 31 week–Normal [35] | 7.8 (1.4) | 8.1 (1.9) | 16.1 (1.8) | 15.8 (3.8) |
| 20 week–Normal [47] | 12.0 | - | 39.0 | - |
| Model Type | Embryonic Development Stage | Health Status of Investigated Embryonic Hearts | |
|---|---|---|---|
| Bharadwaj et al. (2012) [32] | Chicken embryo | HH16, HH23, HH27, HH30 | Normal |
| Yalcin et al. (2011) [95] | Chicken embryo | HH17, HH23, HH27, HH30 | Normal |
| Wang et al. (2009) [103] | Chicken embryo | HH18, HH24 | Normal |
| Liu et al. (2012) [104] | Chicken embryo | HH18 | Normal |
| Liu et al. (2007) [96] | Chicken embryo | HH21 | Normal |
| Tan et al. (2015) [92] | Chicken embryo | HH27 | Normal |
| Ho et al. (2017) [105] | Chicken embryo | HH25 | Normal |
| Kowalski et al. (2014) [106] | Chicken embryo | HH21 | Normal, Left atrial ligated |
| Lindsay et al. (2015) [107] | Chicken embryo | HH18, HH24 | Normal, Occluded pharyngeal arch artery |
| Menon et al. (2015) [111] | Chicken embryo | HH16/17 | Normal, Constricted ventricle junction/outflow tract |
| Lee et al. (2013) [133] | Zebrafish embryo | 20–30 hpf, 40–50 hpf, 60–70 hpf, 80–90 hpf, 110–120 hpf | Normal |
| Boselli and Vermot (2016) [126] | Zebrafish embryo | 48 hpf | Normal |
| Gomez-Garcia et al. (2018) [39] | Zebrafish embryo | 52 hpf | Normal |
| Miller (2011) [134] | Zebrafish embryo | 108 hpf | Normal |
| Foo et al. (2020) [132] | Zebrafish embryo | 120 hpf | Normal |
| Vedula et al. (2017) [31] | Zebrafish embryo | 4 dpf | Normal, Inhibited trabeculaction, Inhibited proliferation, Inhibited ventricle development |
| Lee et al. (2018) [15] | Zebrafish embryo | 4 dpf | Normal, Inhibited trabeculation, Inhibited contractility |
| Wiputra et al. (2018) [35] | Human fetus | 22 week, 31 week | Normal, Tetralogy of fallot |
| Wiputra et al. (2016) [47] | Human fetus | 20 week | Normal |
| Lai et al. (2016) [41] | Human fetus | 20 week | Normal |
| Wiputra et al. (2016) [40] | Human fetus | 20 week | Normal |
| Chen et al. (2017) [138] | Human fetus | 32 week | Normal, Aortic coarctation |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salman, H.E.; Yalcin, H.C. Computational Modeling of Blood Flow Hemodynamics for Biomechanical Investigation of Cardiac Development and Disease. J. Cardiovasc. Dev. Dis. 2021, 8, 14. https://0-doi-org.brum.beds.ac.uk/10.3390/jcdd8020014
Salman HE, Yalcin HC. Computational Modeling of Blood Flow Hemodynamics for Biomechanical Investigation of Cardiac Development and Disease. Journal of Cardiovascular Development and Disease. 2021; 8(2):14. https://0-doi-org.brum.beds.ac.uk/10.3390/jcdd8020014
Chicago/Turabian StyleSalman, Huseyin Enes, and Huseyin Cagatay Yalcin. 2021. "Computational Modeling of Blood Flow Hemodynamics for Biomechanical Investigation of Cardiac Development and Disease" Journal of Cardiovascular Development and Disease 8, no. 2: 14. https://0-doi-org.brum.beds.ac.uk/10.3390/jcdd8020014