
Deletion of a Hand1 lncRNA-Containing Septum Transversum Enhancer Alters lncRNA Expression but Is Not Required for Hand1 Expression


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mouse Strains and Genotyping
2.2. Histology
2.3. Cloning
2.4. In-Situ Hybridization
2.5. Quantitative Real Time PCR
2.6. ATAC-Seq
3. Results
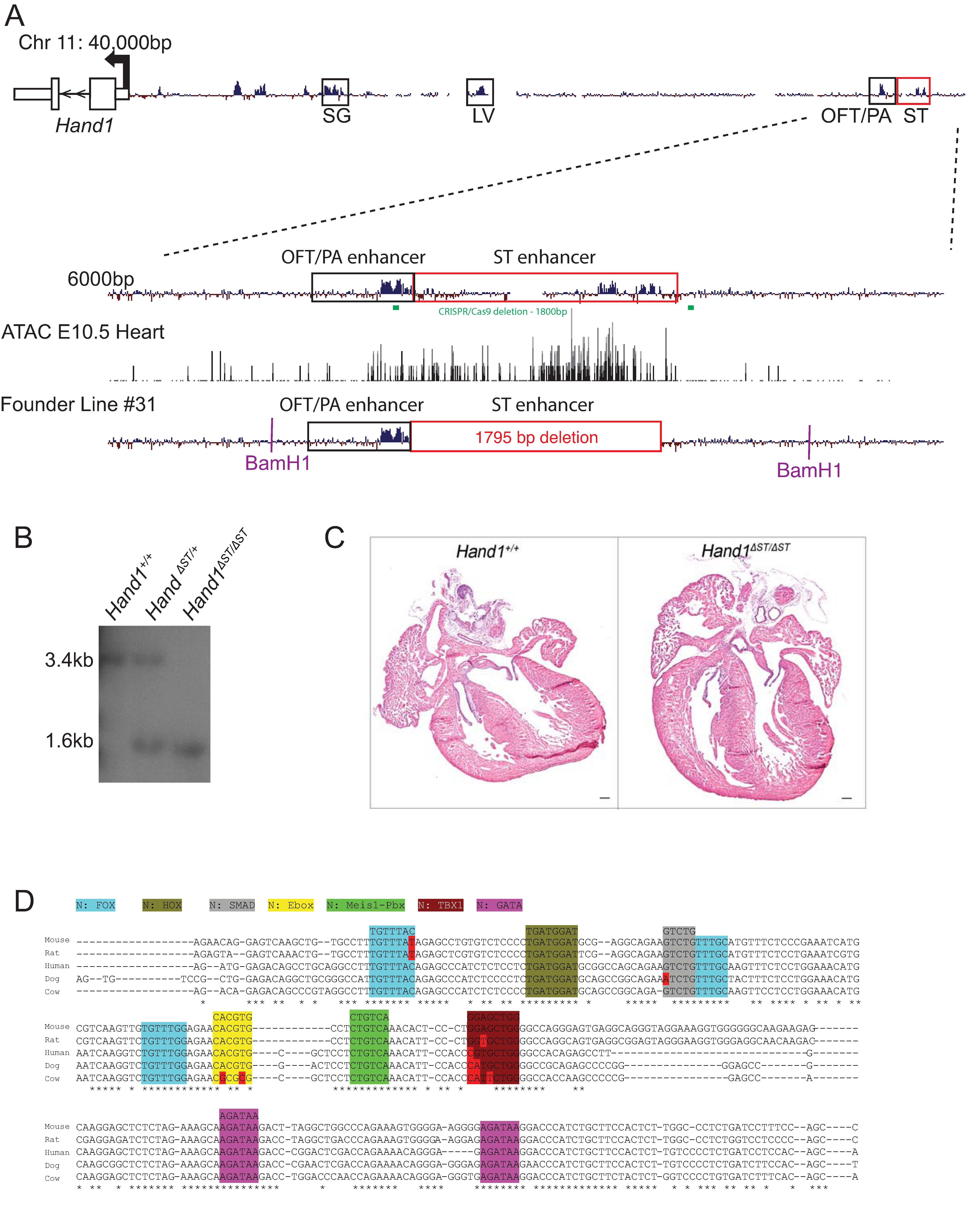
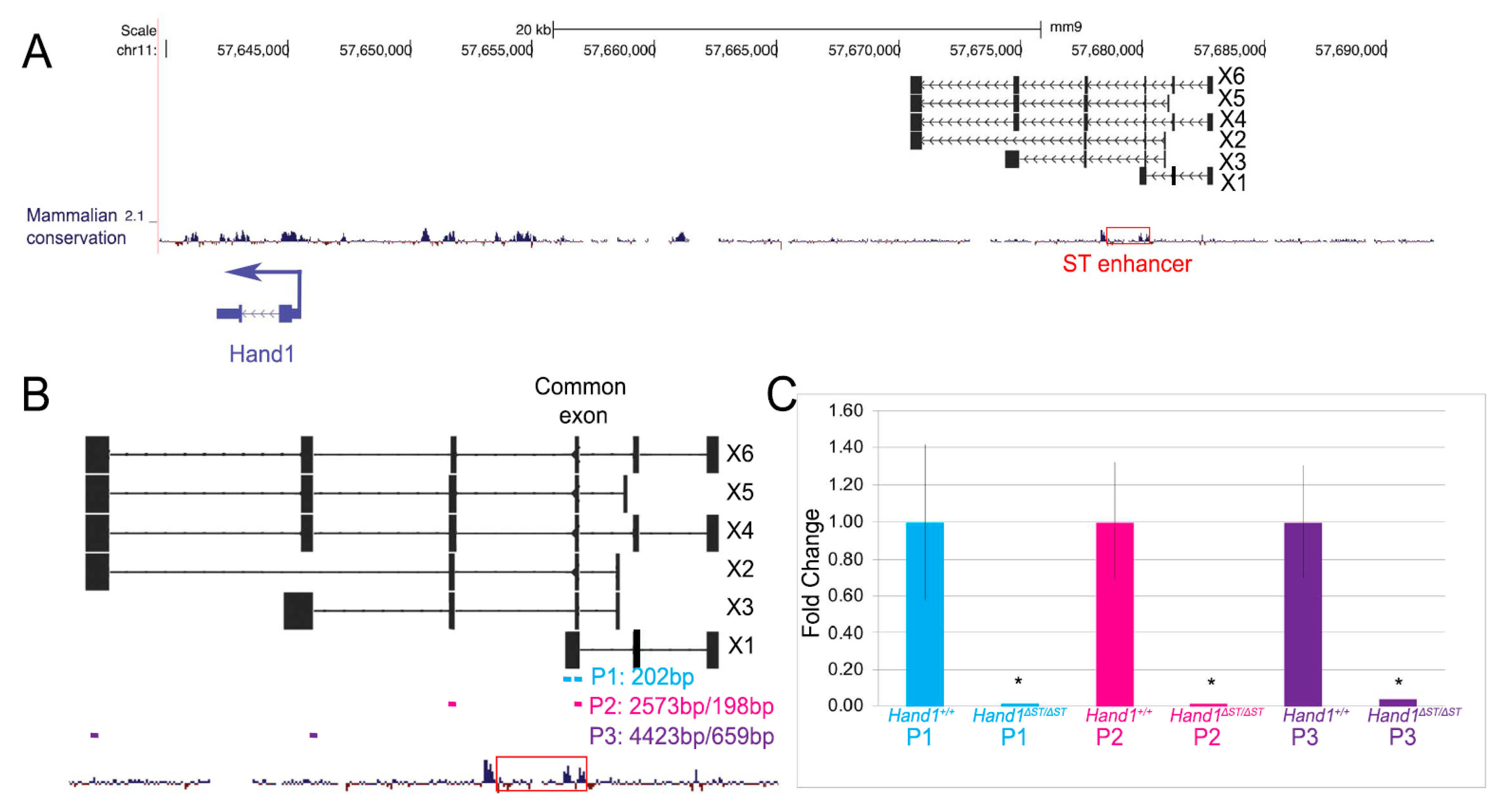
3.1. CRISPR/Cas9 Mediated Deletion of the ST Enhancer
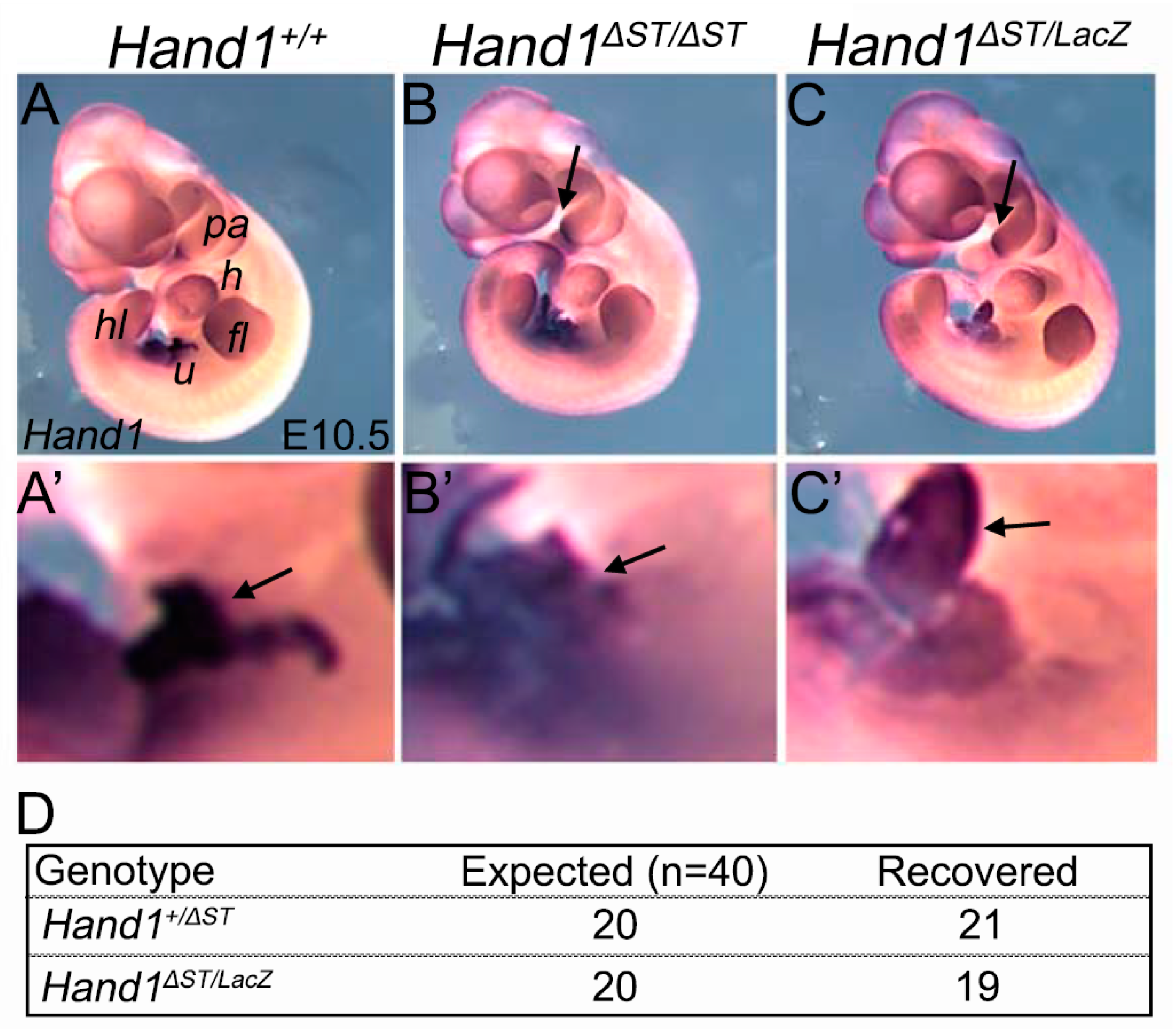
3.2. Loss of a Hand1 Allele over the ST Enhancer Knockout Doesnot Lead to Embryonic Lethality
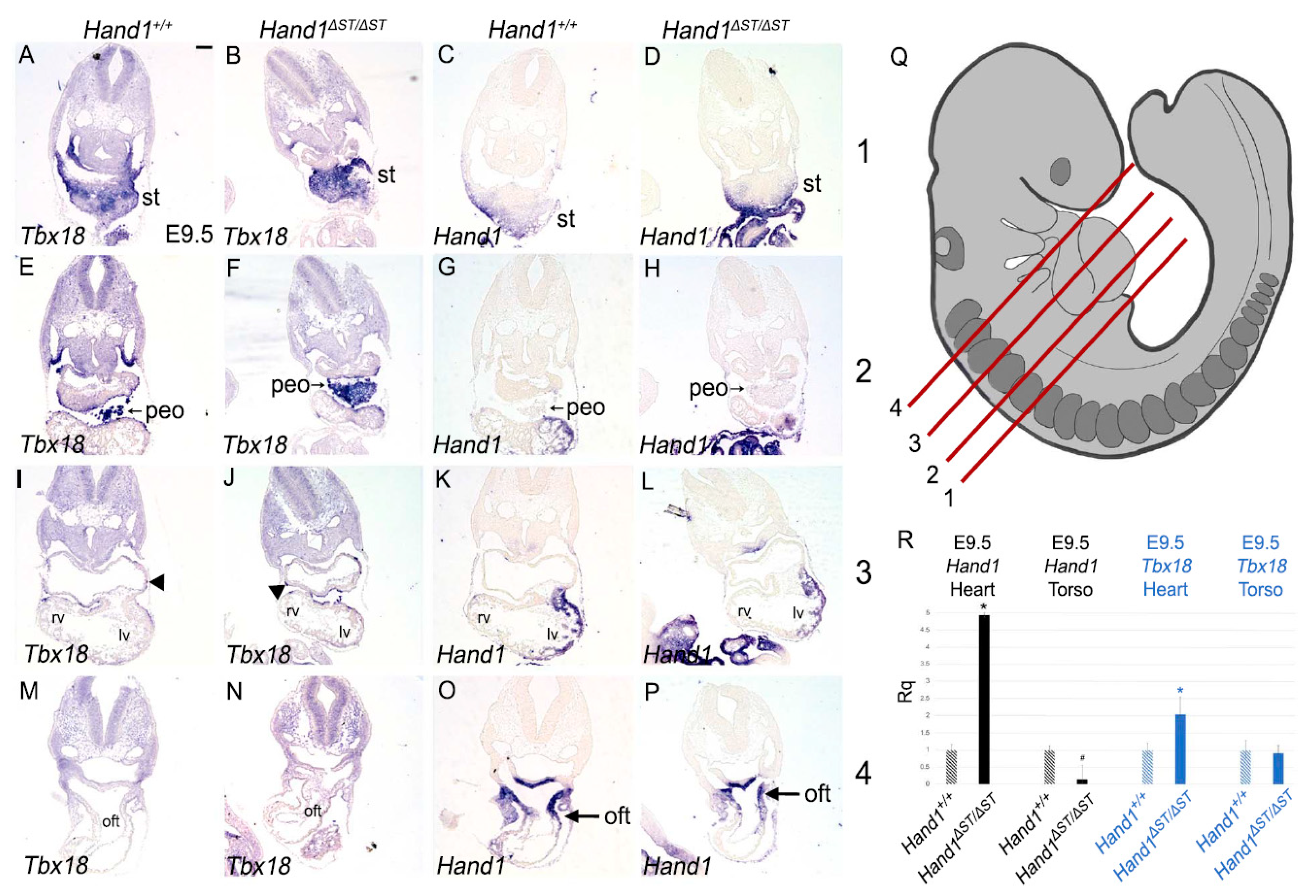
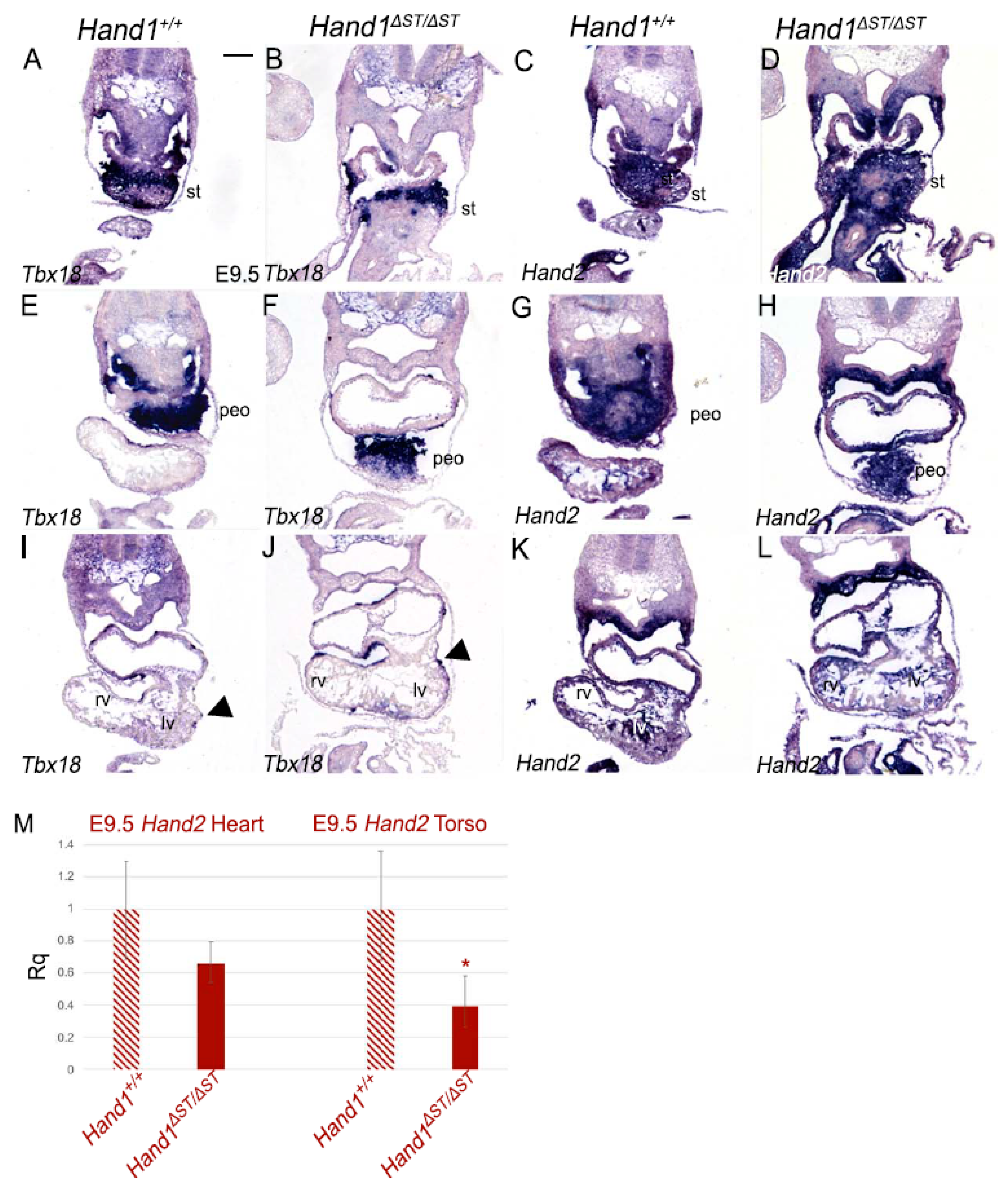
3.3. ST and PEO In Hand1ΔST/ΔST Embryo
3.4. Expression of Hand2 in Hand1ΔST/ΔST Embryos
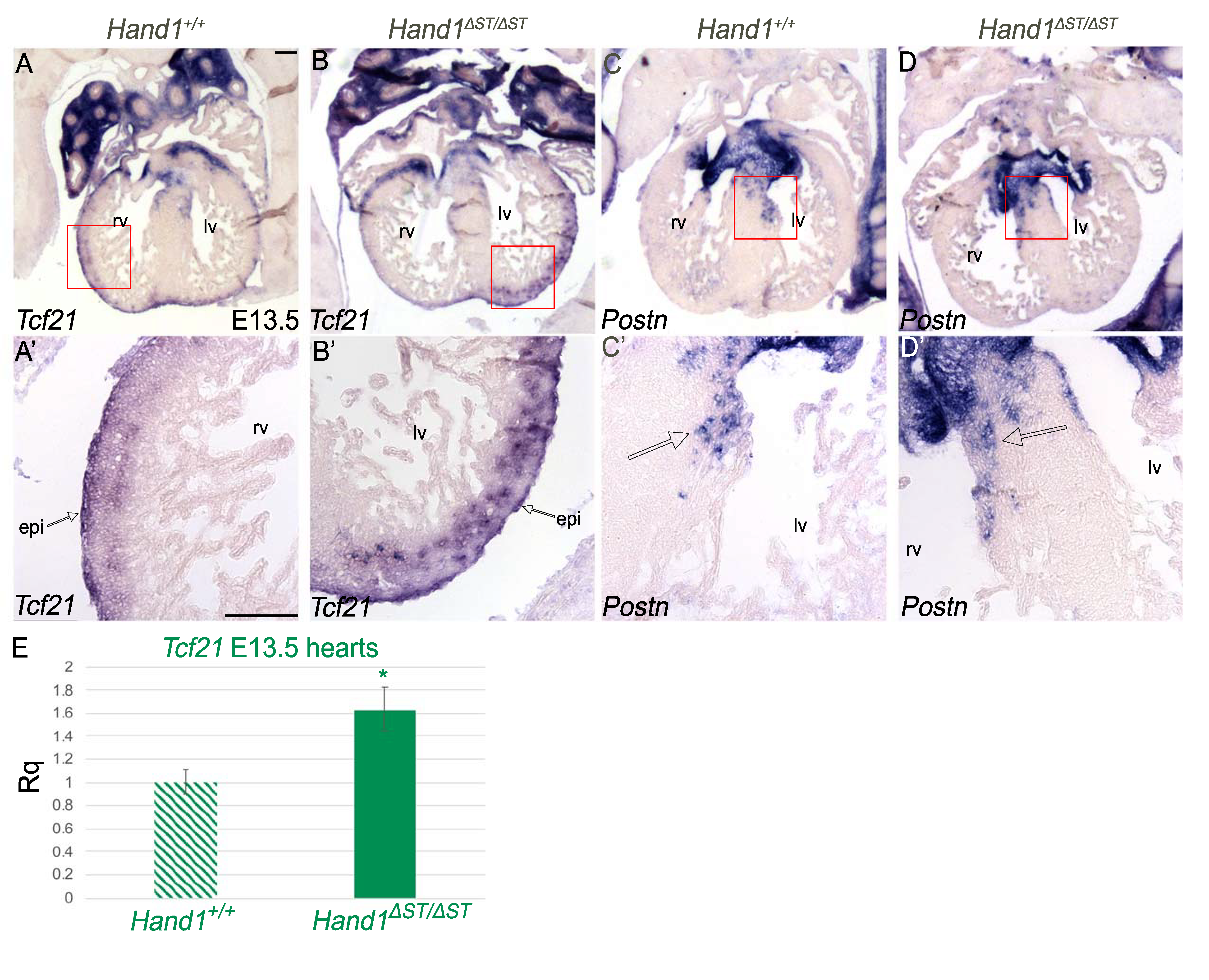
3.5. Development of the Epicardium Is Unaffected In Hand ΔST/ΔST Embryo
3.6. Loss of Putative LncRNA Family Does Not Affect Heart Development
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Simoes, C.F.; Riley, P.R. The ontogeny, activation and function of the epicardium during heart development and regeneration. Development 2018, 145. [Google Scholar] [CrossRef] [Green Version]
- Ho, E.; Shimada, Y. Formation of the epicardium studied with the scanning electron microscope. Dev. Biol. 1978, 66, 579–585. [Google Scholar] [CrossRef]
- Viragh, S.; Challice, C.E. The origin of the epicardium and the embryonic myocardial circulation in the mouse. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 1981, 201, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Hiruma, T.; Hirakow, R. Epicardial formation in embryonic chick heart: Computer-aided reconstruction, scanning, and transmission electron microscopic studies. Am. J. Anat. 1989, 184, 129–138. [Google Scholar] [CrossRef]
- Manner, J.; Pérez-Pomares, J.; Macías, D.; Muñoz-Chápuli, R. The Origin, Formation and Developmental Significance of the Epicardium: A Review. Cells Tissues Organs 2001, 169, 89–103. [Google Scholar] [CrossRef] [PubMed]
- Poelmann, R.E.; Groot, A.C.G.-D.; Mentink, M.M.; Bökenkamp, R.; Hogers, B. Development of the cardiac coronary vascular endothelium, studied with antiendothelial antibodies, in chicken-quail chimeras. Circ. Res. 1993, 73, 559–568. [Google Scholar] [CrossRef] [Green Version]
- Wessels, A.; Hoff, M.J.V.D.; Adamo, R.F.; Phelps, A.L.; Lockhart, M.M.; Sauls, K.; Briggs, L.E.; Norris, R.A.; van Wijk, B.; Perez-Pomares, J.M.; et al. Epicardially derived fibroblasts preferentially contribute to the parietal leaflets of the atrioventricular valves in the murine heart. Dev. Biol. 2012, 366, 111–124. [Google Scholar]
- Yamaguchi, Y.; Cavallero, S.; Patterson, M.; Shen, H.; Xu, J.; Kumar, S.R.; Sucov, H.M. Adipogenesis and epicardial adipose tissue: a novel fate of the epicardium induced by mesenchymal transformation and PPARgamma activation. Proc. Natl. Acad. Sci. USA 2015, 112, 2070–2075. [Google Scholar] [PubMed] [Green Version]
- Zangi, L.; Oliveira, M.S.; Ye, L.Y.; Ma, Q.; Sultana, N.; Hadas, Y.; Chepurko, E.; Später, D.; Zhou, B.; Chew, W.L.; et al. Insulin-Like Growth Factor 1 Receptor-Dependent Pathway Drives Epicardial Adipose Tissue Formation After Myocardial Injury. Circulation 2017, 135, 59–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srivastava, D.; Thomas, T.; Lin, Q.; Kirby, M.L.; Brown, D.D.; Olson, E.N. Regulation of cardiac mesodermal and neural crest development by the bHLH transcription factor, dHAND. Nat. Genet. 1997, 16, 154–160. [Google Scholar] [PubMed]
- Firulli, A.B.; McFadden, D.G.; Lin, Q.; Srivastava, D.; Olson, E.N. Heart and extra-embryonic mesodermal defects in mouse embryos lacking the bHLH transcription factor Hand1. Nat. Genet. 1998, 18, 266–270. [Google Scholar] [CrossRef] [PubMed]
- George, R.M.; Firulli, A.B. Hand Factors in Cardiac Development. Anat. Rec. 2019, 302, 101–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnes, R.M.; Firulli, B.A.; VanDusen, N.J.; Morikawa, Y.; Conway, S.J.; Cserjesi, P.; Vincentz, J.W.; Firulli, A.B. Hand2 Loss-of-Function inHand1-Expressing Cells Reveals Distinct Roles in Epicardial and Coronary Vessel Development. Circ. Res. 2011, 108, 940–949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, B.; Ma, Q.; Rajagopal, S.K.; Wu, S.M.; Domian, I.J.; Rivera-Feliciano, J.; Jiang, D.; Von Gise, A.; Ikeda, S.; Chien, K.R.; et al. Epicardial progenitors contribute to the cardiomyocyte lineage in the developing heart. Nat. Cell Biol. 2008, 454, 109–113. [Google Scholar] [CrossRef] [Green Version]
- Vincentz, J.W.; Casasnovas, J.J.; Barnes, R.M.; Que, J.; Clouthier, D.E.; Wang, J.; Firulli, A.B. Exclusion of Dlx5/6 expression from the distal-most mandibular arches enables BMP-mediated specification of the distal cap. Proc. Natl. Acad. Sci. USA 2016, 113, 7563–7568. [Google Scholar] [CrossRef] [Green Version]
- Firulli, B.A.; George, R.M.; Harkin, J.; Toolan, K.P.; Gao, H.; Liu, Y.; Zhang, W.; Field, L.J.; Liu, Y.; Shou, W.; et al. HAND1 loss-of-function within the embryonic myocardium reveals survivable congenital cardiac defects and adult heart failure. Cardiovasc. Res. 2019, 116, 605–618. [Google Scholar]
- Vincentz, W.J.; Clouthier, D.E.; Firulli, A.B. Mis-Expression of a Cranial Neural Crest Cell-Specific Gene Program in Cardiac Neural Crest Cells Modulates HAND Factor Expression, Causing Cardiac Outflow Tract Phenotypes. J. Cardiovasc. Dev. Dis. 2020, 7, 13. [Google Scholar] [CrossRef]
- Buenrostro, J.D.; Wu, B.; Chang, H.Y.; Greenleaf, W.J. ATAC-seq: A Method for Assaying Chromatin Accessibility Genome-Wide. Curr. Protoc. Mol. Biol. 2015, 109, 21–29. [Google Scholar] [CrossRef]
- Kraus, F.; Haenig, B.; Kispert, A. Cloning and expression analysis of the mouse T-box gene Tbx18. Mech. Dev. 2001, 100, 83–86. [Google Scholar] [CrossRef]
- Wu, S.-P.; Dong, X.-R.; Regan, J.N.; Su, C.; Majesky, M.W. Tbx18 regulates development of the epicardium and coronary vessels. Dev. Biol. 2013, 383, 307–320. [Google Scholar]
- Ratajska, A.; Czarnowska, E.; Ciszek, B. Embryonic development of the proepicardium and coronary vessels. Int. J. Dev. Biol. 2008, 52, 229–236. [Google Scholar]
- Vincentz, J.W.; Toolan, K.P.; Zhang, W.; Firulli, A.B. Hand factor ablation causes defective left ventricular chamber development and compromised adult cardiac function. PLoS Genet. 2017, 13, e1006922. [Google Scholar] [CrossRef] [PubMed]
- Acharya, A.; Baek, S.T.; Huang, G.; Eskiocak, B.; Goetsch, S.; Sung, C.Y.; Banfi, S.; Sauer, M.F.; Olsen, G.S.; Duffield, J.S.; et al. The bHLH transcription factor Tcf21 is required for lineage-specific EMT of cardiac fibroblast progenitors. Development 2012, 139, 2139–2149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lie-Venema, H.; Eralp, I.; Markwald, R.R.; Akker, N.M.S.V.D.; Wijffels, M.C.E.F.; Kolditz, D.P.; Van Der Laarse, A.; Schalij, M.J.; Poelmann, R.E.; Bogers, A.J.J.C.; et al. Periostin expression by epicardium-derived cells is involved in the development of the atrioventricular valves and fibrous heart skeleton. Differentiation 2008, 76, 809–819. [Google Scholar] [CrossRef] [PubMed]
- Vincentz, J.W.; VanDusen, N.J.; Fleming, A.B.; Rubart, M.; Firulli, B.A.; Howard, M.J.; Firulli, A.B. A Phox2- and Hand2-Dependent Hand1 cis-Regulatory Element Reveals a Unique Gene Dosage Requirement for Hand2 during Sympathetic Neurogenesis. J. Neurosci. 2012, 32, 2110–2120. [Google Scholar] [CrossRef]
- Barnes, R.M.; Firulli, B.A.; Conway, S.J.; Vincentz, J.W.; Firulli, A.B. Analysis of the Hand1 cell lineage reveals novel contributions to cardiovascular, neural crest, extra-embryonic, and lateral mesoderm derivatives. Dev. Dyn. 2010, 239, 3086–3097. [Google Scholar]
- Balmer, G.M.; Bollini, S.; Dubé, K.N.; Martinez-Barbera, J.P.; Williams, O.; Riley, P.R. Dynamic haematopoietic cell contribution to the developing and adult epicardium. Nat. Commun. 2014, 5, 4054. [Google Scholar] [CrossRef] [Green Version]
- Firulli, B.A.; Howard, M.J.; McDaid, J.R.; McIlreavey, L.; Dionne, K.M.; E Centonze, V.; Cserjesi, P.; Virshup, D.M.; Firulli, A.B. PKA, PKC, and the Protein Phosphatase 2A Influence HAND Factor Function. Mol. Cell 2003, 12, 1225–1237. [Google Scholar] [CrossRef]
- Firulli, B.A.; Krawchuk, D.; Centonze, V.E.; Virshup, D.M.; Conway, S.J.; Cserjesi, P.; Laufer, E.; Firulli, A.B. Altered Twist1 and Hand2 dimerization is associated with Saethre-Chotzen syndrome and limb abnormalities. Nat. Genet. 2005, 37, 373–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Firulli, B.A.; Redick, B.A.; Conway, S.J.; Firulli, A.B. Mutations within Helix I of Twist1 Result in Distinct Limb Defects and Variation of DNA Binding Affinities. J. Biol. Chem. 2007, 282, 27536–27546. [Google Scholar] [CrossRef] [Green Version]
- George, M.R.; Duan, Q.; Nagle, A.; Kathiriya, I.S.; Huang, Y.; Rao, K.; Haldar, S.M.; Bruneau, B.G. Minimal in vivo requirements for developmentally regulated cardiac long intergenic non-coding RNAs. Development 2019, 146, dev185314. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
George, R.M.; Firulli, A.B. Deletion of a Hand1 lncRNA-Containing Septum Transversum Enhancer Alters lncRNA Expression but Is Not Required for Hand1 Expression. J. Cardiovasc. Dev. Dis. 2021, 8, 50. https://0-doi-org.brum.beds.ac.uk/10.3390/jcdd8050050
George RM, Firulli AB. Deletion of a Hand1 lncRNA-Containing Septum Transversum Enhancer Alters lncRNA Expression but Is Not Required for Hand1 Expression. Journal of Cardiovascular Development and Disease. 2021; 8(5):50. https://0-doi-org.brum.beds.ac.uk/10.3390/jcdd8050050
Chicago/Turabian StyleGeorge, Rajani M., and Anthony B. Firulli. 2021. "Deletion of a Hand1 lncRNA-Containing Septum Transversum Enhancer Alters lncRNA Expression but Is Not Required for Hand1 Expression" Journal of Cardiovascular Development and Disease 8, no. 5: 50. https://0-doi-org.brum.beds.ac.uk/10.3390/jcdd8050050