An Ecological Assessment of Isaria fumosorosea Applications Compared to a Neonicotinoid Treatment for Regulating Invasive Ficus Whitefly

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Treatment Application
2.3. Fungal Treatment Efficacy on Ficus Whitefly Population
2.4. Fungal Identification on Leaf Phylloplane
2.5. Identification of Enzootics Isolated from Ficus Whitefly
2.6. Data Analysis
3. Results
3.1. Field Environmental Conditions
3.1.1. Pre-Treatment
3.1.2. Post-Treatment
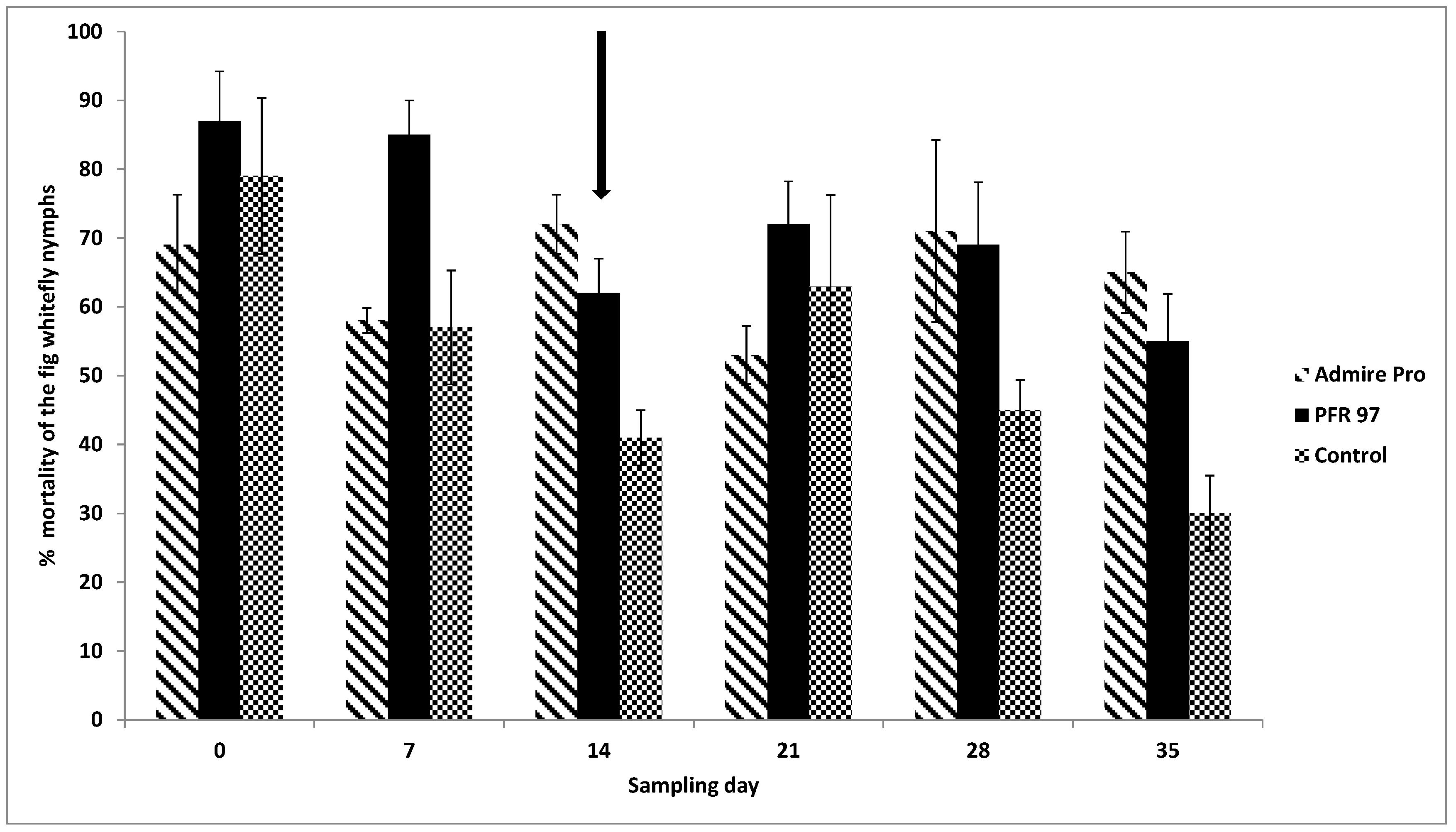
3.2. Insect Pests and Natural Enemies
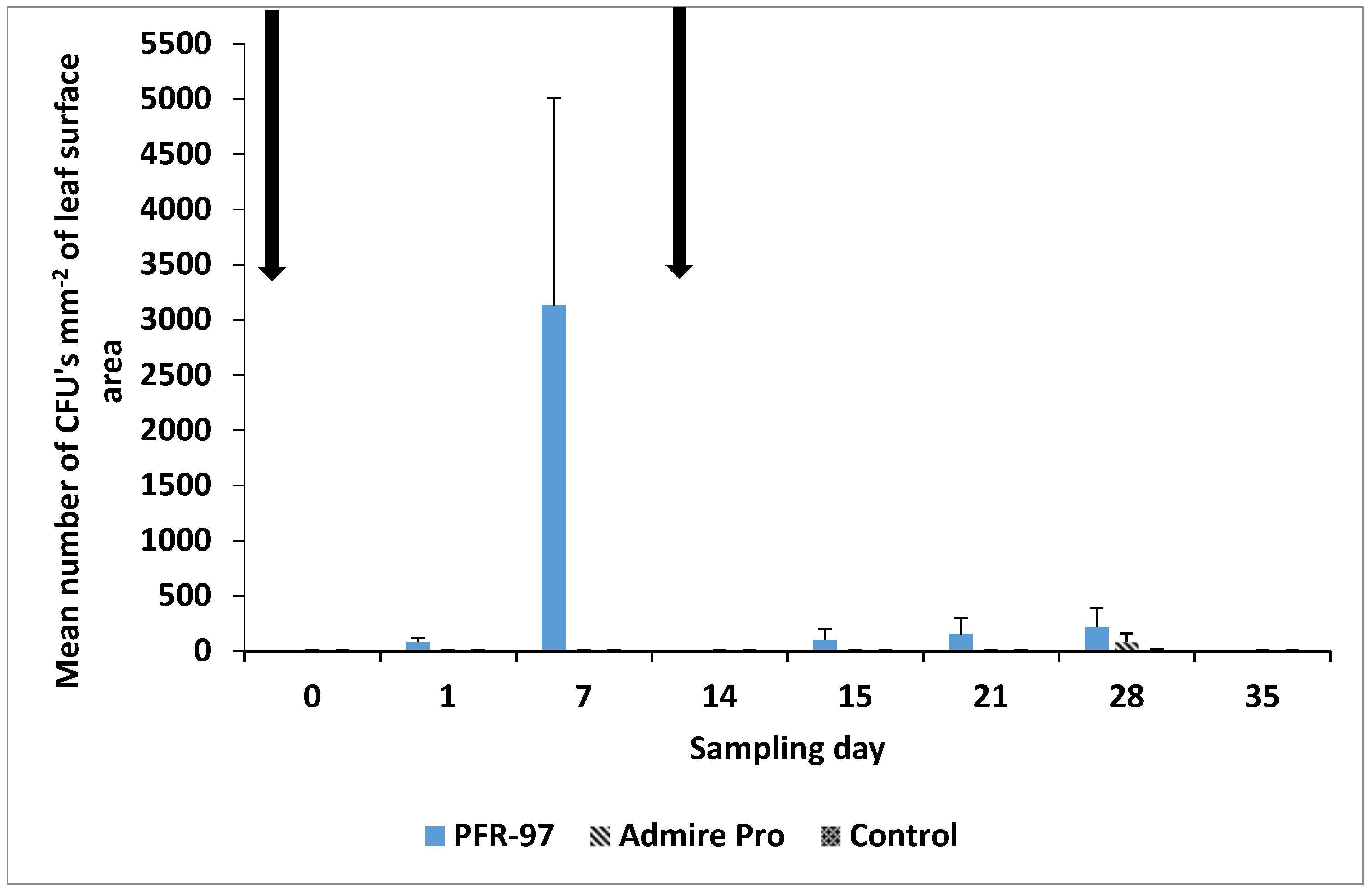
3.3. Blastospore Deposition
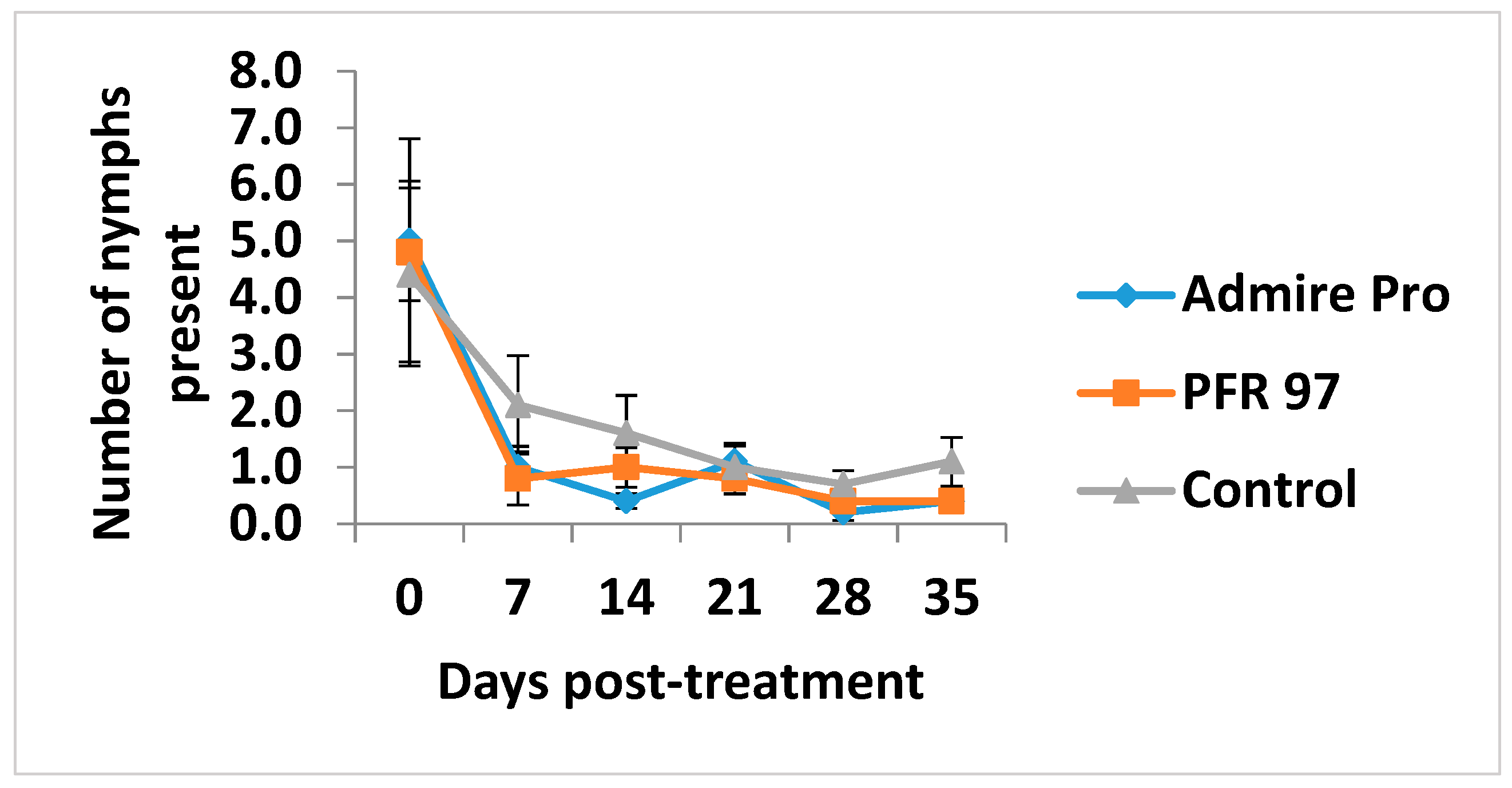
3.4. Population Density of and Treatment Effects on Whitefly Nymphs
3.5. Effects of Treatment on the Occurrence of Enzootic Fungal Species
3.6. Isaria Fumosorosea: Ecological Assessment
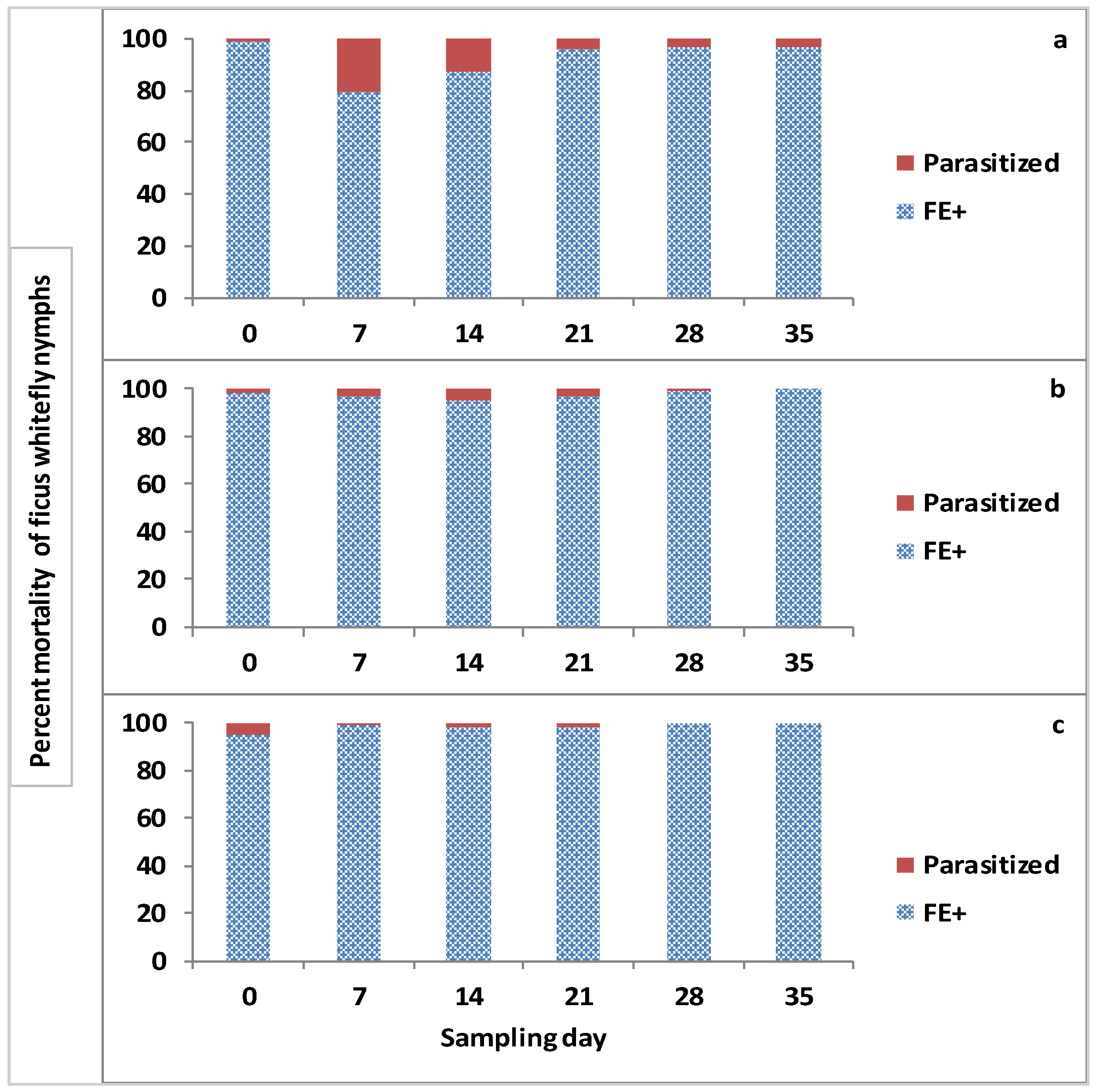
3.7. Effect of Treatments on Parasitism Rate of Parasitoids
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hodges, G. The fig whitefly Singhiella simplex (Singh) (Hemiptera: Aleyrodidae): A new exotic whitefly found on Ficus species in South Florida. Pest Alert DPI-FDACS Bul. 2007. Available online: http://www.freshfromflorida.com/pi/pest-alerts/singhiella-simplex.html (accessed on 19 December 2018).
- Avery, P.B.; Mannion, C.M.; Powell, C.A.; McKenzie, C.L.; Osborne, L.S. Natural enemies managing the invasion of the fig whitefly, Singhiella simplex (Hemiptera: Aleyrodidae), infesting a Ficus benjamina hedge. Fla. Entomol. 2011, 94, 696–698. [Google Scholar] [CrossRef]
- Singh, K. A contribution towards our knowledge of the Aleyrodidae (Whiteflies) of India. Mems. Dept. Agric. India 1931, 12, 1–98. [Google Scholar]
- Singh, K. On some new Rhynchota of the family Aleyrodidae from Burma. Rec. Indian Mus. 1932, 34, 81–88. [Google Scholar]
- David, B.V.; Subramaniam, T.R. Studies on some Indian Aleyrodidae. Rec. Zool. Surv. India 1976, 70, 133–233. [Google Scholar]
- Mannion, C.; Osborne, L.; Hunsberger, A.; Mayer, H.; Hodges, G. Ficus Whitefly: A New Pest in South Florida. University of Florida, IFAS Extension. August 2008. Available online: https://mrec.ifas.ufl.edu/lso/IAWG/FIG/The%20Fig%20Whitefly.htm (accessed on 19 December 2018).
- Mannion, C. Ficus Whitefly Management in the Landscape. University of Florida, IFAS Extension. February 2010. Available online: https://cisr.ucr.edu/pdf/ficus_whitefly_feb2010_fact_sheet.pdf (accessed on 19 December 2018).
- Van Meter, R.J.; Glinski, D.A.; Henderson, W.M.; Garrison, A.W.; Cyterski, M.; Purucker, S.T. Pesticide uptake across the amphibian dermis through soil and overspray exposures. Arch. Environ. Contam. Toxicol. 2015, 69, 545–556. [Google Scholar] [CrossRef]
- Gorgoglione, A.; Bombardelli, F.A.; Pitton, B.J.L.; Oki, L.R.; Haver, D.L.; Young, T.M. Role of sediments in insecticide runoff from urban surfaces: Analysis and modelling. Int. J. Environ. Res. Public Health 2018, 15, 1464. [Google Scholar] [CrossRef]
- Bass, C.; Denholm, I.; Williamson, M.S.; Nauen, R. The global status of insecticide resistance to neonicotinoid insecticides. Pestic. Biochem. Physiol. 2015, 121, 78–87. [Google Scholar] [CrossRef]
- Dângelo, R.A.C.; Michereff-Filho, M.; Campos, M.R.; da Silva, P.S.; Guedes, R.N.C. Insecticide resistance and control failure likelihood of the whitefly Bemisia tabaci (MEAM 1; B biotype): A neotropical scenario. Ann. Appl. Biol. 2018, 172, 88–99. [Google Scholar] [CrossRef]
- Addy-Orduna, L.M.; Brodeur, J.C.; Mateo, R. Oral acute toxicity of imidacloprid, thiamethoxam and clothianidin in eared doves: A contribution for the risk assessment of neonicotinoids in birds. Sci. Total Environ. 2019, 650, 1216–1223. [Google Scholar] [CrossRef]
- Mach, B.M.; Bondarenko, S.; Potter, D.A. Uptake and dissipation of neonicotinoid residues in nectar and foliage of systemically treated woody landscape plants. Environ. Toxicol. Chem. 2018, 37, 860–870. [Google Scholar] [CrossRef]
- Elliot, S.L.; Sabelis, M.W.; Janssen, A.; van der Geest, L.P.S.; Beerling, E.A.M.; Fransen, J. Can plants use entomopathogens as bodyguards? Ecol. Lett. 2000, 3, 228–235. [Google Scholar] [CrossRef] [Green Version]
- Torres-Barragán, A.; Anaya, A.L.; Alatorre, R.; Toriello, C. Entomopathogenic fungi from ‘El Eden’ Ecological Reserve, Quintana, Roo, Mexico. Mycopathologia 2004, 158, 61–71. [Google Scholar] [CrossRef]
- Nielsen, C.; Hajek, A.E. Control of invasive soybean aphid, Aphis glycines (Hemiptera: Aphididae), populations by existing natural enemies in New York State, with emphasis on entomopathogenic fungi. Environ. Entomol. 2005, 34, 1036–1047. [Google Scholar] [CrossRef]
- Lacey, L.A.; Frutos, R.; Kaya, H.K.; Vail, P. Insect pathogens as biological control agents: Do they have a future? Biol. Control 2001, 21, 230–248. [Google Scholar] [CrossRef]
- Meyling, N.V.; Eilenberg, J. Ecology of the entomopathogenic fungi Beauveria bassiana and Metarhizium anisopliae in temperate agroecosystems: Potential for conservation biological control. Biol. Control 2007, 2, 145–155. [Google Scholar] [CrossRef]
- Jackson, M.A.; Dunlap, C.A.; Jaronski, S.T. Ecological considerations in producing and formulating fungal entomopathogens for use in insect control. BioControl 2010, 55, 129–145. [Google Scholar] [CrossRef]
- Vega, F.E.; Goettel, M.S.; Blackwell, M.; Chandler, D.; Jackson, M.A.; Keller, S.; Koike, M.; Maniania, N.K.; Monzón, A.; Ownley, B.H.; et al. Fungal entomopathogens: New insights on their ecology. Fungal Ecol. 2009, 2, 149–159. [Google Scholar] [CrossRef]
- Avery, P.B.; Faull, J.; Simmonds, M.S.J. Effect of different photoperiods on the growth, infectivity and colonization of Trinidadian strains of Paecilomyces fumosoroseus on the greenhouse whitefly, Trialeurodes vaporariorum, using a glass slide bioassay. J. Insect Sci. 2004, 4, 38. [Google Scholar] [CrossRef]
- Avery, P.B.; Faull, J.; Simmonds, M.S.J. Effects of Paecilomyces fumosoroseus and Encarsia formosa on the control of the greenhouse whitefly: Preliminary assessment of a compatibility study. BioControl 2008, 53, 303–316. [Google Scholar] [CrossRef]
- Feng, M.G.; Chen, B.; Ying, S.H. Trials of Beauveria bassiana, Paecilomyces fumosoroseus and imidacloprid for management of Trialeurodes vaporariorum (Homoptera: Aleyrodidae) on greenhouse grown lettuce. Biocontrol Sci. Technol. 2004, 14, 531–544. [Google Scholar] [CrossRef]
- Cuthbertson, A.G.S.; Walters, K.F.A.; Deppe, C. Compatibility of entomopathogenic fungus Lecanicillium muscarium and insecticides for eradication of sweetpotato whitefly, Bemisia tabaci. Mycopathologia 2005, 160, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Fransen, J.J.; Winkelman, K.; van Lenteren, J.C. The differential mortality at various life stages of the greenhouse whitefly, Trialeurodes vaporariorum (Homoptera: Aleyrodidae), by infection with the fungus Aschersonia aleyrodis (Deuteromycotina: Coelomycetes). J. Inverter. Pathol. 1987, 50, 158–165. [Google Scholar] [CrossRef]
- Fransen, J.J.; van Lenteren, J.C. Host selection and survival of the parasitoid Encarsia formosa on greenhouse whitefly, Trialeurodes vaporariorum, in the presence of host infected with the fungus Aschersonia aleyrodis. Entomol. Exp. Appl. 1993, 69, 239–249. [Google Scholar] [CrossRef]
- Vidal, C.; Fargues, J.; Lacey, L. Intraspecific variability of Paecilomyces fumosoroseus: Effect of the temperature on the vegetative growth. J. Invertebr. Pathol. 1997, 70, 18–26. [Google Scholar] [CrossRef]
- Osborne, L.; Landa, Z. Biological control of whiteflies with entomopathogenic fungi. Fla. Entomol. 1992, 75, 456–471. [Google Scholar] [CrossRef]
- Zimmermann, G. The entomopathogenic fungi Isaria farinosa (formerly Paecilomyces farinosus) and the Isaria fumosorosea species complex (formerly Paecilomyces fumosoroseus): Biology, ecology and use in biological control. Biocontrol Sci. Technol. 2008, 18, 865–901. [Google Scholar] [CrossRef]
- Kim, J.S.; Yeon, H.J.; Skinner, M.; Parker, B.L. An oil-based formulation of Isaria fumosorosea blastospores for management of greenhouse whitefly Trialeurodes vaporariorum (Homoptera: Aleyrodidae). Pest Manag. Sci. 2013, 69, 576–581. [Google Scholar] [CrossRef] [PubMed]
- Sterk, G.; Bolckmans, K.; de Jonghe, R.; de Wael, L.; Vermeulen, J. Side-effects of the microbial insecticide PreFeRal WG (Paecilomyces fumosoroseus, strain Apopka 97) on Bombus terrestris. Meded. Fac. Landbouww. Rijksuniv. 1995, 60, 713–717. [Google Scholar]
- Sterk, G.; Bolckmans, K.; van de Veire, M.; Sels, B.; Stepman, W. Side-effects of the microbial insecticide PreFeRal WG (Paecilomyces fumosoroseus, strain Apopka 97) on different species of beneficial arthropods. Meded. Fac. Landbouww. Rijksuniv. 1995, 60, 719–724. [Google Scholar]
- Alma, C.R.; Goettel, M.S.; Roitberg, B.D.; Gillespie, D.R. Combined effects of the entomopathogenic fungus, Paecilomyces fumosoroseus Apopka-97, and the generalist predator, Dicyphus hesperus, on whitefly populations. BioControl 2007, 52, 669–681. [Google Scholar] [CrossRef]
- Zhou, F.; Ali, S.; Huang, Z. Influence of the entomopathogenic fungus Isaria fumosorosea on Axinoscymnus cardilobus (Coleoptera: Coccinellidae) under laboratory conditions. Biocontrol Sci. Technol. 2010, 20, 709–722. [Google Scholar] [CrossRef]
- Sánchez Barahona, C.F.; Threlkeld, B.S.; Avery, P.B.; Francis, A.W.; Cave, R.D. Compatibility and efficacy of the ladybird beetle Thalassa montezumae and the entomopathogenic fungus Isaria fumosorosea for biological control of the green croton scale: Laboratory and greenhouse investigations. Arthropod Plant Interact. 2018, 12, 715–723. [Google Scholar] [CrossRef]
- Chase, A.R.; Osborne, L.S.; Ferguson, V.M. Selective isolation of the entomopathogenic fungi Beauveria bassiana and Metarhizium anisopliae from an artificial potting medium. Fla. Entomol. 1986, 69, 285–292. [Google Scholar] [CrossRef]
- Meyling, N.V. Methods for Isolation of Entomopathogenic Fungi from the Soil Environment-Laboratory Manual. Available online: http://orgprints.org/11200/ (accessed on 20 December 2018).
- Hall, D.G.; Nguyen, R. Toxicity of pesticides to Tamarixia radiata, a parasitoid of the Asian citrus psyllid. BioControl 2010, 55, 601–611. [Google Scholar] [CrossRef]
- Püntener, W. Manual for Field Trials in Plant Protection, 2nd ed.; Agricultural Division, Ciba-Geigy, Ltd.: Basle, Switzerland, 1981. [Google Scholar]
- Cabanillas, H.E.; Jones, W.A. Pathogenicity of Isaria sp. (Hypocreales: Clavicipitaceae) against the sweet potato whitefly B biotype, Bemisia tabaci (Hemiptera: Aleyrodidae). Crop Prot. 2009, 28, 333–337. [Google Scholar] [CrossRef]
- Gökce, A.; Er, M.K. Pathogenicity of Paecilomyces spp. to the glasshouse whitefly, Trialeurodes vaporariorum, with some observations on the fungal infection process. Turk. J. Agric. For. 2005, 29, 331–339. [Google Scholar]
- Gerling, D.; Alomar, O.; Arno, J. Biological control of Bemisia tabaci using predators and parasitoids. Crop Prot. 2001, 20, 779–799. [Google Scholar] [CrossRef]
- Joyce, A.L.; Bellows, T.S.; Headrick, D.H. Reproductive biology and search behavior of Amitus bennetti (Hymenoptera: Platygasteridae), a parasitoid of Bemisia argentifolli (Homoptera: Aleyrodidae). Environ. Entomol. 1999, 28, 282–289. [Google Scholar] [CrossRef]
- Drost, Y.C.; Qiu, Y.T.; Posthuma-Doodeman, C.J.A.M.; van Lenteren, J.C. Life-history and oviposition behaviour of Amitus bennetti, a parasitoid of Bemisia argentifolli. Entomol. Exp. Appl. 1999, 90, 183–189. [Google Scholar] [CrossRef]
- Joyce, A.L.; Bellows, T.S. Field evaluation of Amitus bennetti (Hymenoptera: Platygasteridae), a parasitoid of Bemisia argentifolli (Homoptera: Aleyrodidae), in cotton and bean. Biol. Control 2000, 17, 258–266. [Google Scholar] [CrossRef]
- Scorsetti, A.C.; Humber, R.A.; de Gregorio, C.; Lopez-Lastra, C.C. New records of entomopathogenic fungi infecting Bemisia tabaci and Trialeurodes vaporariorum, pests of horticultural crops, in Argentina. BioControl 2008, 53, 787–796. [Google Scholar] [CrossRef]
- Perea, A.E.I.; Rojas, M.E.; Pineda, V.Y.A. Evaluation of different media for isolation of Trialeurodes vaporariorum (Homoptera: Aleyrodidae) molds. Rev. Colomb. Entomol. 2003, 29, 13–19. [Google Scholar]
- Aiswariya, K.K.; Manjunatha, M.; Naik, M.I. Evaluation of fungi Fusarium semitectum Berk, and Ravenel and Verticillium lecanii (Zimm.) Viegas against spiraling whitefly Aleurodicus disperses Russell on guava. Karnataka J. Agric. Sci. 2007, 20, 283–287. [Google Scholar]
- Panyasiri, C.; Attathom, T.; Poehling, H.M. Pathogenicity of entomopathogenic fungi-potential candidates to control insect pests on tomato under protected cultivation in Thailand. J. Plant Dis. Prot. 2007, 114, 278–287. [Google Scholar] [CrossRef]
- Sharma, L.; Marques, G. Fusarium, an entomopathogen-a myth or reality? Pathogens 2018, 7, 93. [Google Scholar] [CrossRef] [PubMed]
- De Faria, M.R.; Wraight, S.P. Mycoinsecticides and mycoacaricides: A comprehensive list with worldwide coverage and international classification of formulation types. Biol. Control 2007, 43, 237–256. [Google Scholar] [CrossRef]
- Shinde, S.V.; Purohit, M.S.; Sabalpara, A.N.; Patel, M.B. First report of entomopathogenic fungus Lecanicillium lecanii (Zimm.) Zare and Games on sugarcane whitefly Aleurolobus barodensis (Maskell) from Gujarat. Trends BioSci. 2010, 3, 76–77. [Google Scholar]
- Fiedler, Ż.; Sosnowska, D. Nematophagous fungus Paecilomyces lilacinus (Thom) Samson is also a biological agent for control of greenhouse insects and mite pests. BioControl 2007, 52, 547–558. [Google Scholar] [CrossRef]
- Vega, F.E.; Dowd, P.F.; McGuire, M.R.; Jackson, M.A.; Nelsen, T.C. In vitro effects of secondary plant compounds on germination of blastospores of the entomopathogenic fungus Paecilomyces fumosoroseus (Deuteromycotina: Hyphomycetes). J. Inverteb. Pathol. 1997, 70, 209–213. [Google Scholar] [CrossRef] [PubMed]
- Lacey, L.A.; Mercadier, G. The effect of selected allelochemicals on germination of conidia and blastospores and mycelial growth of the entomopathogenic fungus, Paecilomyces fumosoroseus (Deuteromycotina: Hyphomycetes). Mycopathologia 1998, 142, 17–25. [Google Scholar] [CrossRef]
- Imran, M.; Rasool, N.; Rizwan, K.; Zubair, M.; Riaz, M.; Zia-Ul-Haq, M.; Rana, U.A.; Nafady, A.; Jaafar, H.Z.E. Chemical composition and biological studies of Ficus benjamina. Chem. Central J. 2014, 8, 12. [Google Scholar] [CrossRef] [PubMed]
- Krauss, U.; Hidalgo, E.; Arroyo, C.; Piper, S.R. Interaction between the entomopathogens Beauveria bassiana, Metarhizium anisopliae and Paecilomyces fumosoroseus and the mycoparasites Clonostachys spp., Trichoderma harzianum and Lecanicillium lecanii. Biocontrol Sci. Technol. 2004, 14, 331–346. [Google Scholar] [CrossRef]
- Hunter, W.B.; Avery, P.B.; Pick, D.; Powell, C.A. Broad spectrum potential of Isaria fumosorosea against insect pests of citrus. Fla. Entmol. 2011, 94, 1051–1054. [Google Scholar] [CrossRef]
- Smits, N.; Fargues, J.; Rougier, M. Modelling the persistence of quiescent conidia of the entomopathogenic hyphomycete Paecilomyces fumosoroseus exposed to solar radiation. Biocontrol Sci. Technol. 1997, 7, 365–375. [Google Scholar] [CrossRef]
- Bouamama, N.; Vidal, C.; Fargues, J. Effects of fluctuating moisture and temperature regimes on the persistence of quiescent conidia of Isaria fumosorosea. J. Invertebr. Pathol. 2010, 105, 139–144. [Google Scholar] [CrossRef]
- Avery, P.B.; Hunter, W.B.; Hall, D.G.; Jackson, M.A.; Powell, C.A.; Rogers, M.E. Diaphorina citri (Hemiptera: Psyllidae) infection and dissemination of the entomopathogenic fungus Isaria fumosorosea (Hypocreales: Cordycipitaceae) under laboratory conditions. Fla. Entomol. 2009, 92, 608–618. [Google Scholar] [CrossRef]
- Skinner, M.; Parker, B.L.; Kim, J.S. Role of entomopathogenic fungus in integrated pest management. In Integrated Pest Management: Current Concepts and Ecological Perspective; Abrol, D.P., Ed.; Academic Press, Elsevier: San Diego, CA, USA, 2014; pp. 169–191. ISBN 978-0-12-398529-3. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Order | Family | Scientific Name | Common Name | Observation |
|---|---|---|---|---|---|
| Insect pests | Hemiptera | Aleyrodidae | Singhiella simplex | ficus whitefly | feeding on leaves |
| Hemiptera | Aleyrodidae | Tetraleurodes fici | whitefly | feeding on leaves | |
| Thysanoptera | Phlaeothripidae | Gynaikothrips uzeli | weeping ficus thrips | in leaf galls | |
| Natural Enemies | Hymenoptera | Aphelinidae | Encarsia protransvena | parasitoid | parasitized nymphs |
| Hymenoptera | Platygasteridae | Amitus bennetti | parasitoid | parasitized nymphs | |
| Coleoptera | Coccinellidae | Harmonia axyridis | Asian lady beetle | roaming on leaves | |
| Coleoptera | Coccinellidae | Curinus coeruleus | metallic blue lady beetle | roaming on leaves | |
| Neuroptera | Chrysopidae | Chrysopid sp. | green lacewing | eggs, larvae on leaves | |
| Hypocreales | Clavicipitaceae | Isaria fumosorosea | fungal species | leaf surface, nymphs | |
| Hypocreales | Ophiocordycipitceae | Purpureocillium lilacinum | fungal species | leaf surface, nymphs | |
| Hypocreales | Plectosphaerellaceae | Lecanicillium sp. | fungal species | leaf surface, nymphs |
| Mean % Occurrence of Each Fungal Species on Ficus Whitefly Nymphs/Treatment/Sampling Day a | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| 0 | 14 | 35 | |||||||
| Fungal Species | PFR-97 | Admire Pro | Control | PFR-97 | Admire Pro | Control | PFR-97 | Admire Pro | Control |
| Aspergillus sp. | 55 | 50 | 40 | 35 | 39 | 85 | 55 | 70 | 65 |
| Lecanicillium sp. | 0 | 5 | 0 | 0 | 0 | 0 | 15 | 0 | 0 |
| Isaria fumosorosea b | 0 | 0 | 0 | 5 | 0 | 0 | 0 | 0 | 0 |
| Purpureocillium lilacinum | 0 | 0 | 0 | 0 | 16 | 15 | 0 | 0 | 0 |
| Fusarium sp. | 45 | 45 | 60 | 60 | 45 | 0 | 30 | 30 | 35 |
| Mean Number of Colony Forming Units (CFUs) ± SEM (×101)/Days Post-Application/Treatment a | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 15 | 28 | |||||||
| PFR-97 | Admire Pro | Control | PFR-97 | Admire Pro | Control | PFR-97 | Admire Pro | Control | |
| Fungal species b | |||||||||
| AS | 13 ± 0.4 | 7 ± 2.5 | 2 ± 1.2 | 0 | 19 ± 4.5 | 1 ± 0.1 | 2 ± 1.1 | 3 ± 1.3 | 9 ± 2.4 |
| LE | 9 ± 6.6 | 0 | 0 | 0 | 0 | 0 | 0 | 1 ± 0.5 | 9 ± 6.1 |
| IFc | 6 ± 2.6 | 0 | 0 | 101 ± 101.0 | 0 | 0 | 2 ± 1.1 | 1 ± 0.5 | 5 ± 5.0 |
| PL | 75 ± 27.9 | 1 ± 0.5 | 34 ± 13.0 | 0 | 8 ± 6.6 | 8 ± 3.5 | 2 ± 2.0 | 3 ± 2.2 | 31 ± 1.1 |
| FU | 32 ± 13.5 | 51 ± 15.2 | 3 ± 1.3 | 0 | 22 ± 10.0 | 0 | 3 ± 2.0 | 7 ± 1.1 | 4 ± 1.7 |
| PN | 0 | 0 | 0 | 0 | 0 | 0 | 8 ± 6.5 | 1 ± 1.0 | 0 |
| TR | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 ± 1.0 |
| Total Mean % of Ficus Whitefly Nymphs Parasitized/Sampling Day a | |||
|---|---|---|---|
| Treatment | 0 | 14 | 35 |
| PFR-97 | 40 ± 21.3 a | 15 ± 11.1 a | 0 ± 0.0 a |
| Admire Pro | 36 ± 21.8 a | 4 ± 3.8 a | 0 ± 0.0 a |
| Control | 14 ± 12.2 a | 39 ± 20.5 a | 13 ± 12.5 a |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Avery, P.B.; Kumar, V.; Skvarch, E.A.; Mannion, C.M.; Powell, C.A.; McKenzie, C.L.; Osborne, L.S. An Ecological Assessment of Isaria fumosorosea Applications Compared to a Neonicotinoid Treatment for Regulating Invasive Ficus Whitefly. J. Fungi 2019, 5, 36. https://0-doi-org.brum.beds.ac.uk/10.3390/jof5020036
Avery PB, Kumar V, Skvarch EA, Mannion CM, Powell CA, McKenzie CL, Osborne LS. An Ecological Assessment of Isaria fumosorosea Applications Compared to a Neonicotinoid Treatment for Regulating Invasive Ficus Whitefly. Journal of Fungi. 2019; 5(2):36. https://0-doi-org.brum.beds.ac.uk/10.3390/jof5020036
Chicago/Turabian StyleAvery, Pasco B., Vivek Kumar, Edward A. Skvarch, Catharine M. Mannion, Charles A. Powell, Cindy L. McKenzie, and Lance S. Osborne. 2019. "An Ecological Assessment of Isaria fumosorosea Applications Compared to a Neonicotinoid Treatment for Regulating Invasive Ficus Whitefly" Journal of Fungi 5, no. 2: 36. https://0-doi-org.brum.beds.ac.uk/10.3390/jof5020036