Revealing of Non-Cultivable Bacteria Associated with the Mycelium of Fungi in the Kerosene-Degrading Community Isolated from the Contaminated Jet Fuel


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and Cultures Isolation
2.2. Morphological Features of Micromycetes
2.3. Growth Ability on the TS-1 Jet Fuel
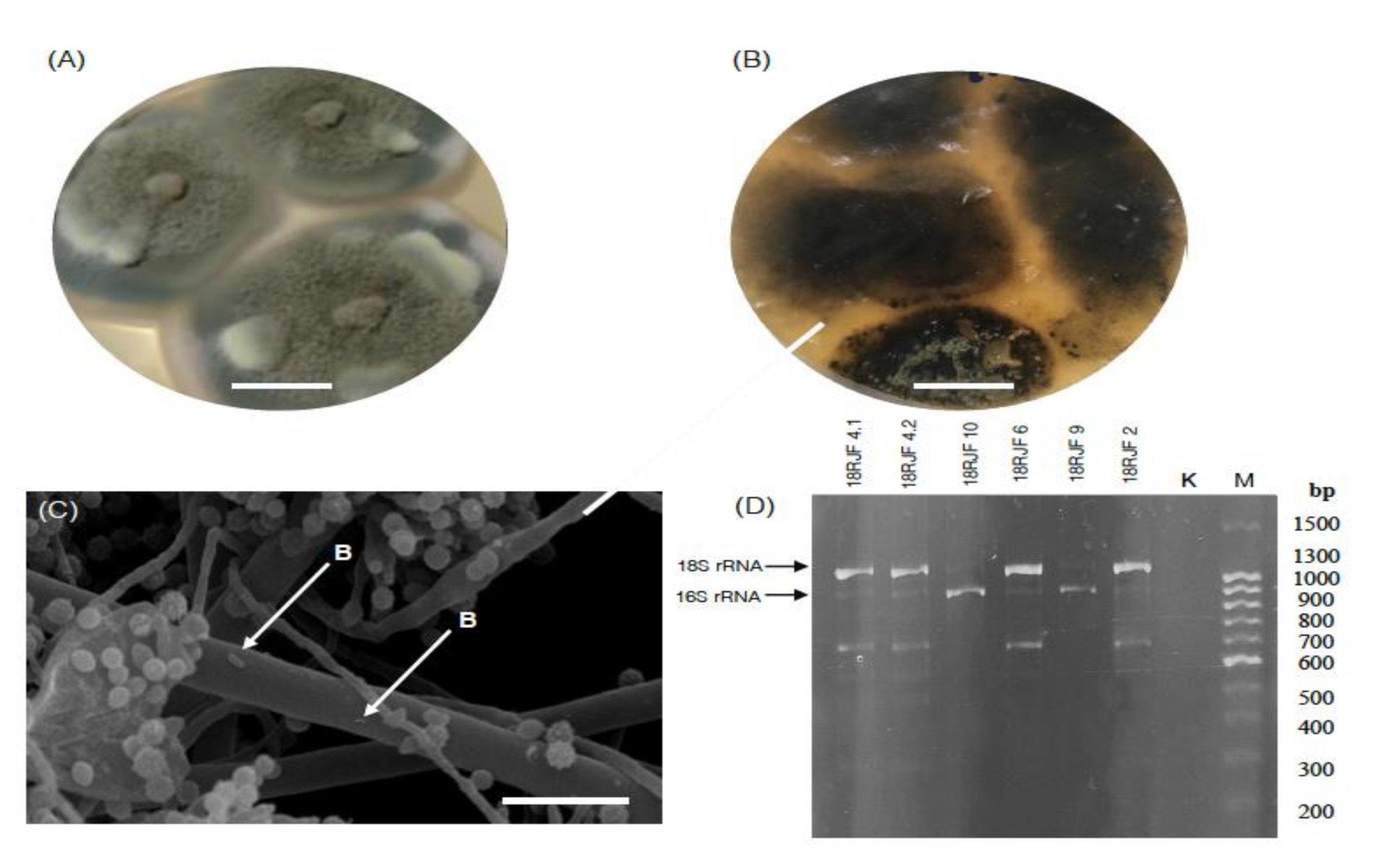
2.4. Express Assay for the Detection of 16S/18S rRNA Genes
2.5. Next-Generation 16S rRNA Amplicon Sequencing (16S rRNA Metabarcoding) and Bioinformatic Analysis
3. Results
3.1. Micromycetes Isolated from the TS-1 Jet Fuel
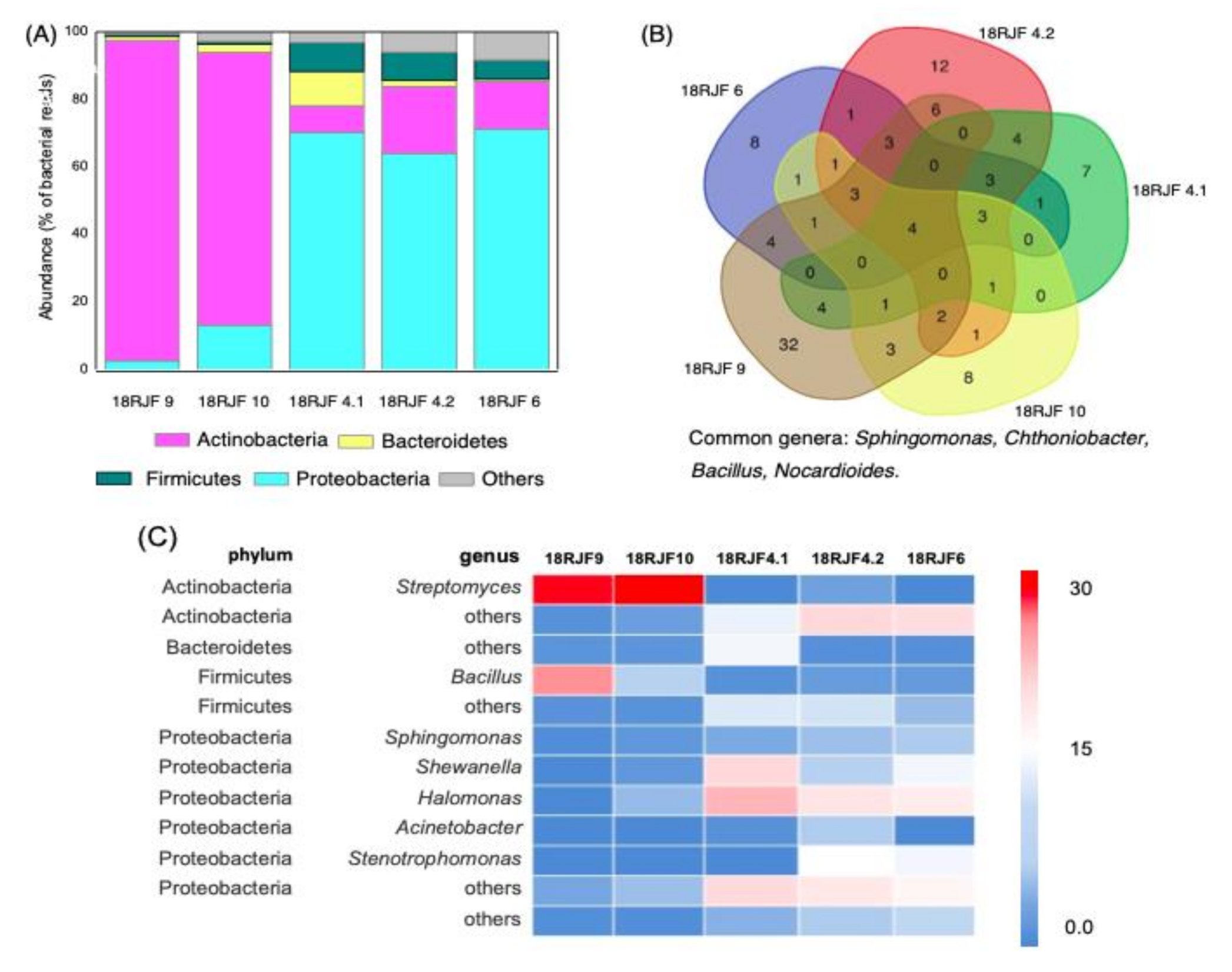
3.2. Bacterial Component of the Micromycetes’ Biomass Isolated from Jet Fuel Communities Revealed by 16S rRNA Data
3.3. The Growth Capacity of Isolated Communities on the TS-1 Jet Fuel
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Iverson, W.P. Microbial corrosion of metals. In Advances in Applied Microbiology; Academic Press: Cambridge, MA, USA, 1987; Volume 32, pp. 1–36. [Google Scholar]
- Gerasimenko, A.A.; Yampol’skaya, T.E. Exfoliation corrosion of aluminum alloys II. Protection methods, their effectiveness and improvement. Prot. Met. 2000, 36, 397–407. [Google Scholar] [CrossRef]
- Itah, A.Y.; Brooks, A.A.; Ogar, B.O.; Okure, A.B. Biodegradation of international jet A-1 aviation fuel by microorganisms isolated from aircraft tank and joint hydrant storage systems. Bull. Environ. Contam. Toxicol. 2009, 83, 318–327. [Google Scholar] [CrossRef]
- Buddie, A.G.; Bridge, P.D.; Kelley, J.; Ryan, M.J. Candida keroseneae sp. nov., a novel contaminant of aviation kerosene. Lett. Appl. Microbiol. 2011, 52, 70–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soriano, A.U.; Martins, L.F.; Ventura, E.S.D.; de Landa, F.H.G.T.; da Auraujo Valoni, E.; Faria, F.R.D.; Ferreira, R.F.; Clara, M.; Fallera, K.; Valério, R.R.; et al. Microbiological aspects of biodiesel and biodiesel/diesel blends biodeterioration. Int. Biodeterior. Biodegrad. 2015, 99, 102–114. [Google Scholar] [CrossRef]
- Passman, F.J. Microbial contamination and its control in fuels and fuel systems since 1980—A review. Int. Biodeterior. Biodegrad. 2013, 81, 88–104. [Google Scholar] [CrossRef]
- Rodríguez-Rodríguez, C.E.; Rodríguez, E.; Blanco, R.; Cordero, I.; Segura, D. Fungal contamination of stored automobile-fuels in a tropical environment. J. Environ. Sci. 2010, 22, 1595–1601. [Google Scholar] [CrossRef]
- Varjani, S.J. Microbial degradation of petroleum hydrocarbons. Bioresour. Technol. 2017, 223, 277–286. [Google Scholar] [CrossRef]
- Koshlaf, E.; Ball, A. Soil bioremediation approaches for petroleum hydrocarbon polluted environments. Aims Microbiol. 2017, 3, 25–49. [Google Scholar] [CrossRef]
- Leahy, J.G.; Colwell, R.R. Microbial degradation of hydrocarbons in the environment. Microbiol. Mol. Biol. Rev. 1990, 54, 305–315. [Google Scholar] [CrossRef]
- Bücker, F.; de Moura, T.M.; da Cunha, M.E.; de Quadros, P.D.; Beker, S.A.; Cazarolli, J.C.; Caramão, E.B.; Frazzon, A.P.G.; Bento, F.M. Evaluation of the deteriogenic microbial community using qPCR, n-alkanes and FAMEs biodegradation in diesel, biodiesel and blends (B5, B10, and B50) during storage. Fuel 2018, 233, 911–917. [Google Scholar] [CrossRef]
- Das, N.; Chandran, P. Microbial degradation of petroleum hydrocarbon contaminants: An overview. Biotechnol. Res. Int. 2011, 2011, 941810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shabir, G.; Afzal, M.; Anwar, F.; Tahseen, R.; Khalid, Z.M. Biodegradation of kerosene in soil by a mixed bacterial culture under different nutrient conditions. Int. Biodeterior. Biodegrad. 2008, 61, 161–166. [Google Scholar] [CrossRef]
- Boychenko, S.; Shkilnuk, I.; Turchak, V. The problems of biopollution with jet fuels and the way of achieving solution. Transport. 2008, 23, 253–257. [Google Scholar] [CrossRef] [Green Version]
- Sørensen, G.; Pedersen, D.V.; Nørgaard, A.K.; Sørensen, K.B.; Nygaard, S.D. Microbial growth studies in biodiesel blends. Bioresour. Technol. 2011, 102, 5259–5264. [Google Scholar] [CrossRef] [PubMed]
- Shkilniuk, I.; Boichenko, S. Methodically organizational principles of biological stability providing of aviation fuels. Pr. Inst. Lotnictwa 2014, 237, 76–83. [Google Scholar] [CrossRef]
- Bücker, F.; Barbosa, C.S.; Quadros, P.D.; Bueno, M.K.; Fiori, P.; te Huang, C.; Frazzon, A.P.G.; Ferrão, M.F.; de Oliveira Camargo, F.A.; Bento, F.M. Fuel biodegradation and molecular characterization of microbial biofilms in stored diesel/biodiesel blend B10 and the effect of biocide. Int. Biodeterior. Biodegrad. 2014, 95, 346–355. [Google Scholar] [CrossRef]
- Chiciudean, I.; Mereuţă, I.; Ionescu, R.; Vassu, T.; Tănase, A.M.; Stoica, I. Jet A-1 Bacterial contamination: A case study of cultivable bacteria diversity, alkane degradation and biofilm formation. Pol. J. Environ. Stud. 2019, 28, 4139–4146. [Google Scholar]
- Atlas, R.M. Microbial degradation of petroleum hydrocarbons: An environmental perspective. Microbiol. Rev. 1981, 45, 180–209. [Google Scholar] [CrossRef]
- Chaillan, F.; Le Flèche, A.; Bury, E.; Phantavong, Y.H.; Grimont, P.; Saliot, A.; Oudot, J. Identification and biodegradation potential of tropical aerobic hydrocarbon-degrading microorganisms. Res. Microbiol. 2004, 155, 587–595. [Google Scholar] [CrossRef]
- De Azambuja, A.O.; Bücker, F.; de Quadros, P.D.; Zhalnina, K.; Dias, R.; Vacaro, B.B.; Bento, F.M. Microbial community composition in Brazilian stored diesel fuel of varying sulfur content, using high-throughput sequencing. Fuel 2017, 189, 340–349. [Google Scholar] [CrossRef]
- Pereira, E.; Napp, A.P.; Allebrandt, S.; Barbosa, R.; Reuwsaat, J.; Lopes, W.; Maria do Carmo, R.P. Biodegradation of aliphatic and polycyclic aromatic hydrocarbons in seawater by autochthonous microorganisms. Int. Biodeterior. Biodegrad. 2019, 145, 104789. [Google Scholar] [CrossRef]
- Hasan, I. Biodegradation of kerosene by Aspergillus niger and Rhizopus stolonifer. Appl. Environ. Microbiol. 2014, 2, 31–36. [Google Scholar]
- Lotfinasabasl, S.; Gunale, V.R.; Rajurkar, N.S. Assessment of petroleum hydrocarbon degradation from soil and tarball by fungi. Biosci. Discov. 2012, 3, 186–192. [Google Scholar]
- Dhar, K.; Dutta, S.; Anwar, M.N. Biodegradation of petroleum hydrocarbon by indigenous fungi isolated from ship breaking yards of Bangladesh. Int. Res. J. Biol. Sci. 2014, 3, 22–30. [Google Scholar]
- Mnif, S.; Chamkha, M.; Sayadi, S. Isolation and characterization of Halomonas sp. strain C2SS100, a hydrocarbon-degrading bacterium under hypersaline conditions. J. Appl. Microbiol. 2009, 107, 785–794. [Google Scholar] [CrossRef]
- Ameen, F.; Moslem, M.; Hadi, S.; Al-Sabri, A.E. Biodegradation of diesel fuelhydrocarbons by mangrove fungi from Red Sea Coast of Saudi Arabia. Saudi J. Biol. Sci. 2016, 23, 211–218. [Google Scholar]
- Boonchan, S.; Britz, M.L.; Stanley, G.A. Degradation and mineralization of high-molecular-weight polycyclic aromatic hydrocarbons by defined fungal-bacterial cocultures. Appl. Environ. Microbiol. 2000, 66, 1007–1019. [Google Scholar] [CrossRef] [Green Version]
- White, J.; Gilbert, J.; Hill, G.; Hill, E.; Huse, S.M.; Weightman, A.J.; Mahenthiralingam, E. Culture-independent analysis of bacterial fuel contamination provides insight into the level of concordance with the standard industry practice of aerobic cultivation. Appl. Environ. Microbiol. 2011, 77, 4527–4538. [Google Scholar] [CrossRef] [Green Version]
- Boer, W.D.; Folman, L.B.; Summerbell, R.C.; Boddy, L. Living in a fungal world: Impact of fungi on soil bacterial niche development. FEMS Microbiol. Rev. 2005, 29, 795–811. [Google Scholar]
- Furuno, S.; Päzolt, K.; Rabe, C.; Neu, T.R.; Harms, H.; Wick, L.Y. Fungal mycelia allow chemotactic dispersal of polycyclic aromatic hydrocarbon-degrading bacteria in water-unsaturated systems. Environ. Microbiol. 2010, 12, 1391–1398. [Google Scholar]
- Wick, L.Y.; Furuno, S.; Harms, H. Fungi as transport vectors for contaminants and contaminant-degrading bacteria. In Handbook of Hydrocarbon and Lipid Microbiology; Springer: Berlin/Heidelberg, Germany, 2010; pp. 1555–1561. [Google Scholar]
- Partida-Martinez, L.P.; Hertweck, C. Pathogenic fungus harbours endosymbiotic bacteria for toxin production. Nature 2005, 437, 884–888. [Google Scholar] [CrossRef] [PubMed]
- Wick, L.Y.; Remer, R.; Würz, B.; Reichenbach, J.; Braun, S.; Schäfer, F.; Harms, H. Effect of fungal hyphae on the access of bacteria to phenanthrene in soil. Environ. Sci. Technol. 2007, 41, 500–505. [Google Scholar] [CrossRef] [PubMed]
- Furuno, S.; Remer, R.; Chatzinotas, A.; Harms, H.; Wick, L.Y. Use of mycelia as paths for the isolation of contaminant-degrading bacteria from soil. Microb. Biotechnol. 2012, 5, 142–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banitz, T.; Johst, K.; Wick, L.Y.; Schamfuß, S.; Harms, H.; Frank, K. Highways versus pipelines: Contributions of two fungal transport mechanisms to efficient bioremediation. Environ. Microbiol. Rep. 2013, 5, 211–218. [Google Scholar] [CrossRef]
- Schamfuß, S.; Neu, T.R.; Harms, H.; van der Meer, J.R.; Tecon, R.; Wick, L.Y. Mycelial networks enhance the bioavailability of PAH in water unsaturated environments. Environ. Sci. Technol. 2013, 47, 6908–6915. [Google Scholar] [CrossRef]
- Wick, L.; De Munain, A.; Springael, D.; Harms, H. Responses of mycobacterium sp. LB501T to the low bioavailability of solid anthracene. Appl. Microbiol. Biotechnol. 2002, 58, 378–385. [Google Scholar]
- Ferradji, F.Z.; Mnif, S.; Badis, A.; Rebbani, S.; Fodil, D.; Eddouaouda, K.; Sayadi, S. Naphthalene and crude oil degradation by biosurfactant producing Streptomyces spp. isolated from Mitidja plain soil (North of Algeria). Int. Biodeterior. Biodegrad. 2014, 86, 300–308. [Google Scholar] [CrossRef]
- Rapp, P.; Backhaus, S. Formation of extracellular lipases by filamentous fungi, yeasts, and bacteria. Enzym. Microb. Technol. 1992, 14, 938–943. [Google Scholar] [CrossRef]
- Thom, C.; Church, M.B. The Aspergilli; Williams and Wilkins Co.: Baltimore, MD, USA, 1926; p. 272. [Google Scholar]
- Matlakowska, R.; Sklodowska, A. The culturable bacteria isolated from organic-rich black shale potentially useful in biometallurgical procedures. J. Aapplied Microbiol. 2009, 107, 858–866. [Google Scholar] [CrossRef]
- Evans, C.G.T.; Herbert, D.; Tempest, D.W. Chapter XIII the continuous cultivation of microorganisms: 2, construction of a chemostat. In Methods in Microbiology; Academic Press: Cambridge, MA, USA, 1970; Volume 2, pp. 277–327. [Google Scholar]
- Lobakova, E.; Vasilieva, S.; Kashcheeva, P.; Ivanova, E.; Dolnikova, G.; Chekanov, K.; Dedov, A. New bio-hybrid materials for bioremoval of crude oil spills from marine waters. Int. Biodeterior. Biodegrad. 2016, 108, 99–107. [Google Scholar] [CrossRef]
- Shapiro, T.; Dolnikova, G.; Nemtseva, N.; Sanjieva, D.; Lobakova, E. Identification and physiological characterisationtics of athe consortium of hydrocarbon-oxidizing bacteria of oil and oilpetroleum products. J. Microbiol. Epidemiol. Immunobiol. 2018, 107–113. [Google Scholar] [CrossRef]
- Ivanova, E.A.; Lobakova, E.S.; Idiatulov, R.K.; Shapiro, T.N.; Sandzhieva, D.A.; Kuznetsova, O.V.; Zaitseva, Y.u.N.; Dzhabrailova, K.h.S.; Dedov, A.G. Biocomposite materials for purification of aqueous media contaminated with hydrocarbons. Pet. Chem. 2019, 59, 420–426. [Google Scholar] [CrossRef]
- Gorelova, O.A.; Kosevich, I.A.; Baulina, O.I.; Fedorenko, T.A.; Torshkhoeva, A.Z.; Lobakova, E.S. Associations between the White Sea invertebrates and oxygen-evolving phototrophic microorganisms. Mosc. Univ. Biol. Sci. Bull. 2009, 64, 16–22. [Google Scholar] [CrossRef]
- Raper, K.B.; Fennel, D.I. The Genus Aspergillus; Williams & Wilkins: Baltimore, MD, USA, 1965; p. 686. [Google Scholar]
- Raper, K.B.; Thom, C.; Fennell, D.I. A Manual of the Penicillia; Hefner Publishing Co.: New York, NY, USA, 1949; p. 875. [Google Scholar]
- Pitt, G.I. A Laboratory Guide to Common Penicillium Species, 2nd ed.; CSIRO Food Research Laboratory: North Ryde, NSW, Australia, 1968; p. 188. [Google Scholar]
- Klich, M. Identification of Common Aspergillus Species; CBS: New York, NY, USA, 2002; p. 116. [Google Scholar]
- Samson, R.A.; Frisvard, J.C. Penicillium subgenus Penicillium: New taxonomic schemes, mycotoxins and other extrolites. Stud. Mycol. 2004, 49, 251. [Google Scholar]
- Yilmaz, N.; Houbraken, J.; Hoekstra, E.S.; Frisvad, J.C.; Visagie, C.M.; Samson, R.A. Delimitation and characterisation of Talaromyces purpurogenus and related species. Pers. Mol. Phylogeny Evol. Fungi 2012, 29, 39–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yilmaz, N.; Visagie, C.M.; Houbraken, J.; Frisvad, J.C.; Samson, R.A. Polyphasic taxonomy of the genus Talaromyces. Stud. Mycol. 2014, 78, 175–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, A.J.; Frisvad, J.C.; Sun, B.D.; Varga, J.; Kocsubé, S.; Dijksterhuis, J.; Kim, D.H.; Hong, S.B.; Houbraken, J.; Samson, R. Aspergillus section Nidulantes (formerly Emericella): Polyphasic taxonomy, chemistry and biology. Stud. Mycol. 2016, 84, 1–118. [Google Scholar] [CrossRef] [Green Version]
- Vasilieva, A.A.; Chekunova, L.N.; Polyakova, A.V. Effect of temperature on growth and viability of Hormoconis resinae and Phialophora sp. developing in the aviation fuel. Mycol. Phytopatologiya 2009, 43, 312–316. [Google Scholar]
- Wang, Y.; Tian, R.M.; Gao, Z.M.; Bougouffa, S.; Qian, P.-Y. Optimal eukaryotic 18S and universal 16S/18S ribosomal RNA primers and their application in a study of symbiosis. PLoS ONE 2014, 9, e90053. [Google Scholar] [CrossRef] [Green Version]
- Chekanov, K.; Kublanovskaya, A.; Lobakova, E. Eukaryotic sequences in the 16Sr RNA metagenomic dataset of algal–bacterial consortia of the White Sea coastal zone. J. Eukaryot. Microbiol. 2019, 66, 853–856. [Google Scholar] [CrossRef]
- Bates, S.T.; Berg-lyons, D.; Caporaso, J.G.; Walters, W.A.; Knight, R.; Fierer, N. Examining the global distribution of dominant archaeal populations in soil. ISME J. 2010, 5, 908–917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kublanovskaya, A.; Chekanov, K.; Solovchenko, A.; Lobakova, E. Cyanobacterial diversity in the algal–bacterial consortia from Subarctic regions: New insights from the rock baths at White Sea Coast. Hydrobiologia 2019, 830, 17–31. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huse, S.M.; Welch, D.B.M.; Voorhis, A.; Shipunova, A.; Morrison, H.G.; Eren, A.M.; Sogin, M.L. VAMPS: A website for visualization and analysis of microbial population structures. BMC Bioinform. 2014, 15, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beccati, A.; Gerken, J.; Quast, C.; Yilmaz, P.; Glöckner, F.O. SILVA tree viewer: Interactive web browsing of the SILVA phylogenetic guide trees. BMC Bioinform. 2017, 18, 433. [Google Scholar] [CrossRef] [Green Version]
- Gaylarde, C.C.; Bento, F.M.; Kelley, J. Microbial contamination of stored hydrocarbon fuels and its control. Rev. Microbiol. 1999, 30, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Rauch, M.E.; Graef, H.W.; Rozenzhak, S.M.; Jones, S.E.; Bleckmann, C.A.; Kruger, R.L.; Naik, R.R.; Stone, M.O. Characterization of microbial contamination in United States Air Force aviation fuel tanks. J. Ind. Microbiol. Biotechnol. 2006, 33, 29–36. [Google Scholar] [CrossRef]
- Yemashova, N.A.; Murygina, V.P.; Zhukov, D.V.; Zakharyantz, A.A.; Gladchenko, M.A.; Appanna, V.; Kalyuzhnyi, S.V. Biodeterioration of crude oil and oil derived products: A review. Rev. Environ. Sci. Bio Technol. 2007, 6, 315–337. [Google Scholar] [CrossRef]
- Martin-Sanchez, P.M.; Becker, R.; Gorbushina, A.A.; Toepel, J. An improved test for the evaluation of hydrocarbon degradation capacities of diesel-contaminating microorganisms. Int. Biodeterior. Biodegrad. 2018, 129, 89–94. [Google Scholar]
- Korshunova, T.; Chetverikov, S.; Bakaeva, M.; Kuzina, E.; Rafikova, G.; Chetverikova, D.; Loginov, O. Microorganisms in the elimination of oil pollution consequences. Appl. Biochem. Microbiol. 2019, 55, 344–354. [Google Scholar] [CrossRef]
- Mitchell, R.; Alexander, M. Lysis of soil fungi by bacteria. Can. J. Microbiol. 1963, 9, 169–177. [Google Scholar] [CrossRef]
- Balachandran, C.; Duraipandiyan, V.; Balakrishna, K.; Ignacimuthu, S. Petroleum and polycyclic aromatic hydrocarbons (PAHs) degradation and naphthalene metabolism in Streptomyces sp. (ERI-CPDA-1) isolated from oil contaminated soil. Bioresour. Technol. 2012, 112, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Zhang, Y.; Ding, R.; Li, D.; Gao, Y.; Yang, M. Comparison of archaeal and bacterial community structures in heavily oil-contaminated and pristine soils. J. Biosci. Bioeng. 2009, 108, 400–407. [Google Scholar] [CrossRef]
- Tapilatu, Y.H.; Grossi, V.; Acquaviva, M.; Militon, C.; Bertrand, J.C.; Cuny, P. Isolation of hydrocarbon-degrading extremely halophilic archaea from an uncontaminated hypersaline pond (Camargue, France). Extremophiles 2010, 14, 225–231. [Google Scholar] [CrossRef]
- Le Borgne, S.; Paniagua, D.; Vazquez-Duhalt, R. Biodegradation of organic pollutants by halophilic bacteria and archaea. J. Mol. Microbiol. Biotechnol. 2008, 15, 74–92. [Google Scholar] [CrossRef]
- Baoune, H.; El Hadj-Khelil, A.O.; Pucci, G.; Sineli, P.; Loucif, L.; Polti, M.A. Petroleum degradation by endophytic Streptomyces spp. isolated from plants grown in contaminated soil of southern Algeria. Ecotoxicol. Environ. Saf. 2018, 147, 602–609. [Google Scholar] [CrossRef]
- Chen, J.; Huang, P.T.; Zhang, K.Y.; Ding, F.R. Isolation of biosurfactant producers, optimization and properties of biosurfactant produced by Acinetobacter sp. from petroleum-contaminated soil. J. Appl. Microbiol. 2012, 112, 660–671. [Google Scholar] [CrossRef]
- Yanan, W.; Yingying, L.; Jianliang, X.; Ke, S.; Yu, G.; Xiaolong, X. Exploring the degradation potential of Halomonas bacteria from oil-contaminated marine environment. China Pet. Process. Petrochem. Technol. 2018, 20, 91–98. [Google Scholar]
- Bianciotto, V.; Bandi, C.; Minerdi, D.; Sironi, M.; Tichy, H.V.; Bonfante, P. An obligately endosymbiotic mycorrhizal fungus itself harbors obligately intracellular bacteria. Appl. Environ. Microbiol. 1996, 62, 3005–3010. [Google Scholar]
- Pion, M.; Spangenberg, J.E.; Simon, A.; Bindschedler, S.; Flury, C.; Chatelain, A.; Bshary, R.; Job, D.; Junier, P. Bacterial farming by the fungus Morchella crassipes. Proc. R. Soc. B Biol. Sci. 2013, 280, 20132242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haq, I.U.; Graupner, K.; Nazir, R.; van Elsas, J.D. The genome of the fungal-interactive soil bacterium Burkholderia terrae BS001—A plethora of outstanding interactive capabilities unveiled. Genome Biol. Evol. 2014, 6, 1652–1668. [Google Scholar] [CrossRef] [PubMed]
- Abbasian, F.; Lockington, R.; Mallavarapu, M.; Naidu, R. A comprehensive review of aliphatic hydrocarbon biodegradation by bacteria. Appl. Biochem. Biotechnol. 2015, 176, 670–699. [Google Scholar] [CrossRef] [PubMed]
- Young, D.; Rice, J.; Martin, R.; Lindquist, E.; Lipzen, A.; Grigoriev, I.; Hibbett, D. Degradation of bunker C fuel oil by white-rot fungi in sawdust cultures suggests potential applications in bioremediation. PLoS ONE 2015, 10, e0130381. [Google Scholar] [CrossRef] [Green Version]
- Johnsen, A.R.; Wick, L.Y.; Harms, H. Principles of microbial PAH-degradation in soil. Environ. Pollut. 2005, 133, 71–84. [Google Scholar] [CrossRef]
- Seneviratne, G.; Zavahir, J.S.; Bandara, W.M.M.S.; Weerasekara, M.L.M.A.W. Fungal-bacterial biofilms: Their development for novel biotechnological applications. World J. Microbiol. Biotechnol. 2008, 24, 739–743. [Google Scholar] [CrossRef]
- Costerton, J.W.; Lewandowski, Z.; DeBeer, D.; Caldwell, D.; Korber, D.; James, G. Biofilms, the customized microniche. J. Bacteriol. 1994, 176, 2137. [Google Scholar] [CrossRef] [Green Version]
- Herath, H.M.L.I.; Rajapaksha, A.U.; Vithanage, M.; Seneviratne, G. Developed fungal–bacterial biofilms as a novel tool for bioremoval of hexavelant chromium from wastewater. Chem. Ecol. 2014, 30, 418–427. [Google Scholar] [CrossRef]
- Pandit, A.; Adholeya, A.; Cahill, D.; Brau, L.; Kochar, M. Microbial biofilms in nature: Unlocking their potential for agricultural applications. J. Appl. Microbiol. 2020, 129, 199–211. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Isolate | Cultural Characteristics | Morphological Characteristics | |
|---|---|---|---|
| Mycelial Morphology | Sporulation Characteristics | ||
| 18RJF6 | Widely grown, velvety surface with individual | 1–2 mm height | Sporulation: grayish-green, in some cases, the edge of the colonies lighter |
| Conidiophores: 100–150 μm × 2.5–3 μm Typically, biverticillate and symmetrical; each conidiophore contained 3–5 metules and lanceolate phialides | |||
| 18RJF9 | felted areas; smooth reverse side; with exudate; reddish brown | ||
| 18RJF10 | Elliptic conidia: 2–3 µm × 1.5–2.5 μm; smooth or slightly rough | ||
| 18RJF2 | Colonies of 10–20 mm in diameter with low growth; velvety surface, rugose | 1–2 mm height | Sporulation: bright green |
| Conidiophores: 40–110 μm × 2–3 μm | |||
| Typically, biverticillate and symmetrical, as a rule, with additional branches (10–25 μm length); each conidiophore contained 5 or 6 metals and were from lanceolate to flask-shaped phialides | |||
| Elliptic to fusiform conidia: 2.5–6 μm × 2.5–4 μm; smooth or slightly rough | |||
| 18RJF4.1 | Colonies of 2.5–3.5 mm in diameter; moderate growth rate; velvety surface; radially folded; colorless or yellow drops of exudate | 1–3 mm height | Sporulation: yellow-green |
| Conidiophores: 250–500 μm × 2.5–3.5 μm | |||
| Typically, terverticillate and asymmetric, with a pressed lateral twig; each conidiophore contained 5 or 6 metules and from 3 to 6 bottle-shaped phialides | |||
| Subspherical to ellipsoidal conidia: 3.0–4.0 μm × 2.8–3.8 μm; smooth | |||
| 18RJF4.2 | Colonies of 10–20 mm in diameter with slow growth rate; strongly folded with a well-defined edge; | 2–4 mm height | Sporulation: blue-green |
| Conidiophores formed on the substrate mycelium only: 500 μm × 5–8 μm | |||
| the reverse side radially folded, initially without a specific coloration and then wine-purple | The spore heads biseriate radial, up to 20 µm in diameter; metules (6–7 μm length), phialides 7–10 µm × 2.0–2.5 µm | ||
| Globular conidia: 2.5 µm × 3.5 µm; prickly, green in mass | |||
| Isolate | Isolation Medium | Micromycete | TS-1 Degradation Score | Mycelium Lysis |
|---|---|---|---|---|
| 18RJF2 | EM | Talaromyces rugulosus | 0 | − |
| 18RJF4.1 | CP | Penicillium chrysogenum | 3 | + |
| 18RJF4.2 | CP | Aspergillus sydowii | 5 | + |
| 18RJF6 | WA | Talaromyces amestolkiae | 5 | + |
| 18RJF9 | WA | Talaromyces amestolkiae | 3 | + |
| 18RJF10 | WA | Talaromyces amestolkiae | 2 | + |
| Isolate | Mitochondrial rRNA | GenBank ID | Bacteria with Putative Petroleum Destruction Activity |
|---|---|---|---|
| 18RJF2 | Talaromyces | MW393516 | None |
| 18RJF4.1 | Penicillium | MW393517 | Sphingomonas, Bacillus, Rhodococcus, Halomonas, Nocardioides |
| 18RJF4.2 | Aspergillus | MW393518 | Sphingomonas, Bacillus, Pseudomonas, Stenotrophomonas, Arthrobacter, Halomonas, Nocardioides |
| 18RJF6 | Talaromyces | MW393519 | Sphingomonas, Bacillus, Pseudomonas, Stenotrophomonas, Arthrobacter, Halomonas, Nocardioides |
| 18RJF9 | Talaromyces | MW393520 | Sphingomonas, Bacillus, Pseudomonas, Stenotrophomonas, Arthrobacter, Streptomyces, Nocardioides |
| 18RJF10 | Talaromyces | MW393521 | Sphingomonas, Bacillus, Arthrobacter, Halomonas, Streptomyces, Nocardioides |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shapiro, T.; Chekanov, K.; Alexandrova, A.; Dolnikova, G.; Ivanova, E.; Lobakova, E. Revealing of Non-Cultivable Bacteria Associated with the Mycelium of Fungi in the Kerosene-Degrading Community Isolated from the Contaminated Jet Fuel. J. Fungi 2021, 7, 43. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7010043
Shapiro T, Chekanov K, Alexandrova A, Dolnikova G, Ivanova E, Lobakova E. Revealing of Non-Cultivable Bacteria Associated with the Mycelium of Fungi in the Kerosene-Degrading Community Isolated from the Contaminated Jet Fuel. Journal of Fungi. 2021; 7(1):43. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7010043
Chicago/Turabian StyleShapiro, Tatiana, Konstantin Chekanov, Alina Alexandrova, Galina Dolnikova, Ekaterina Ivanova, and Elena Lobakova. 2021. "Revealing of Non-Cultivable Bacteria Associated with the Mycelium of Fungi in the Kerosene-Degrading Community Isolated from the Contaminated Jet Fuel" Journal of Fungi 7, no. 1: 43. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7010043