Trichoderma asperellum T76-14 Released Volatile Organic Compounds against Postharvest Fruit Rot in Muskmelons (Cucumis melo) Caused by Fusarium incarnatum



Abstract
:1. Introduction
2. Materials and Methods
2.1. Trichoderma Species and Pathogen Isolates
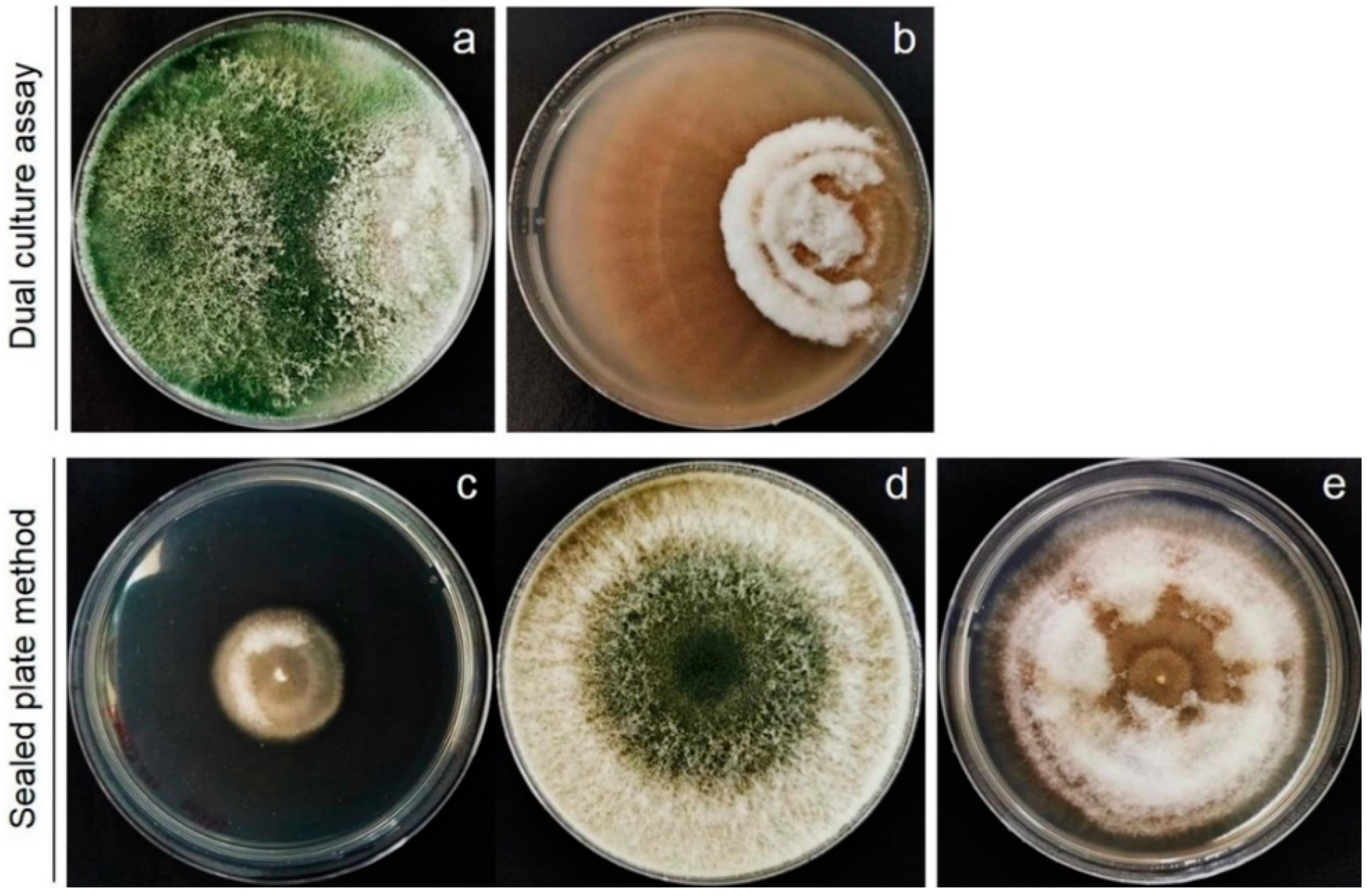
2.2. In Vitro Screening of Trichoderma spp. against F. incarnatum by Dual-Culture Assay
2.3. Volatile Antifungal Bioassay
2.4. GC/MS Analysis
2.5. Sealed Plate Method of Commercial Volatile against Mycelial Growth of F. incarnatum
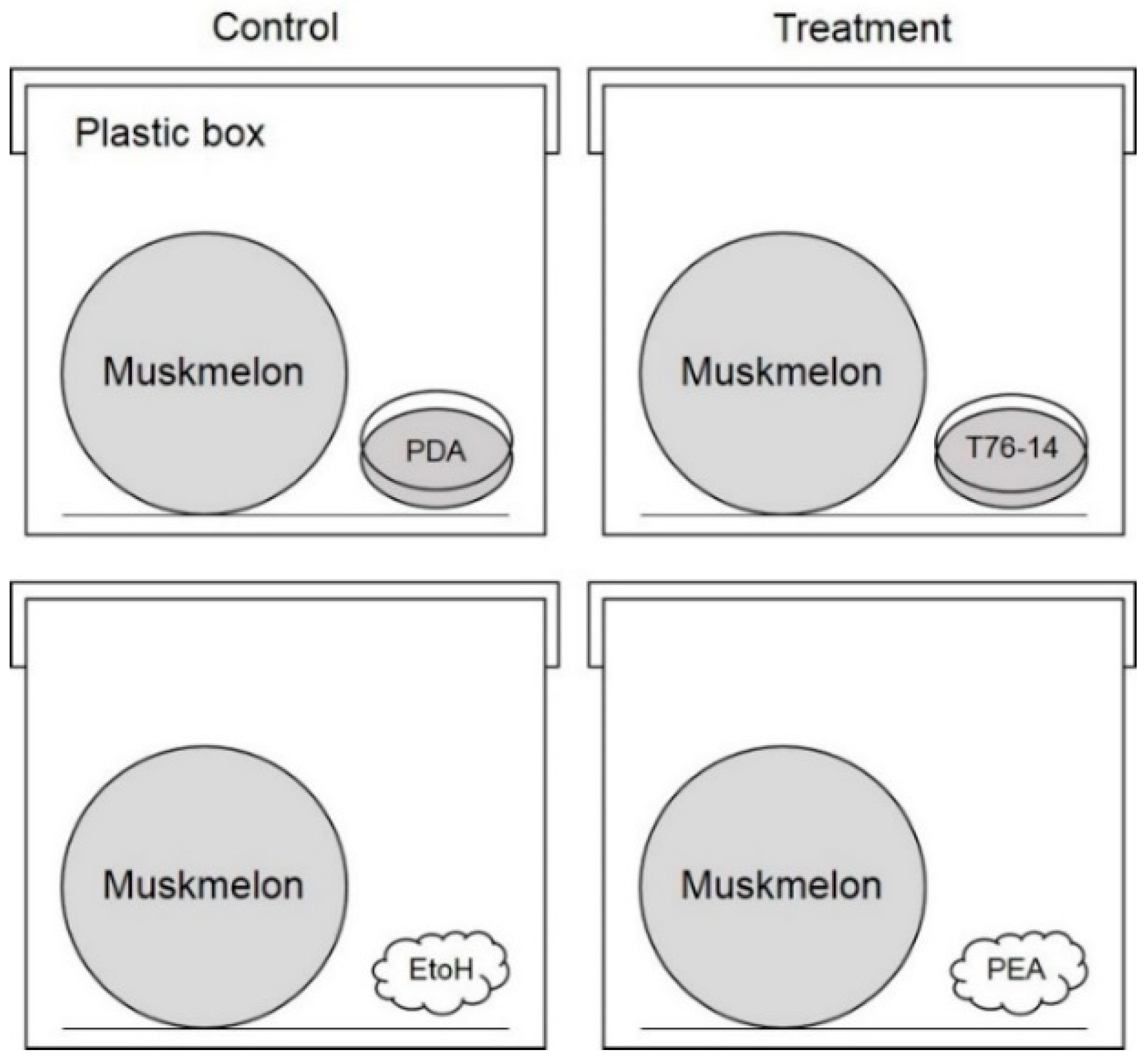
2.6. In Vivo Test of Volatile against Postharvest Fruit Rot
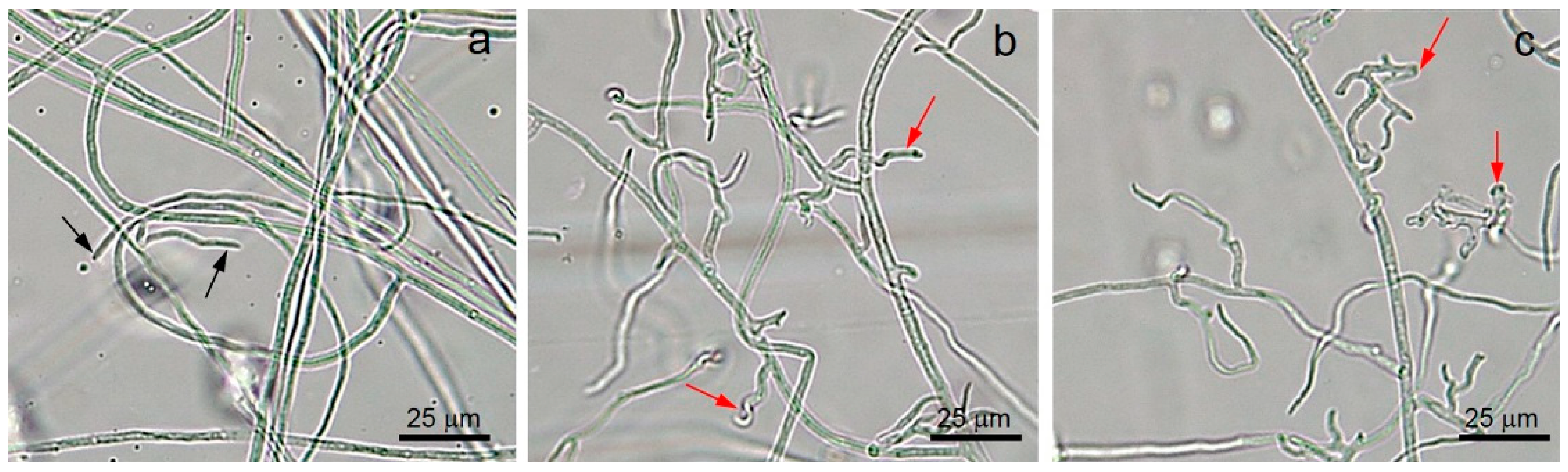
2.7. Morphological Study of F. incarnatum Exposed to Volatiles
2.8. Effect of VOCs Emitted from T. asperellum T76-14 on Fruit Rot
2.9. Statistical Analysis
3. Results
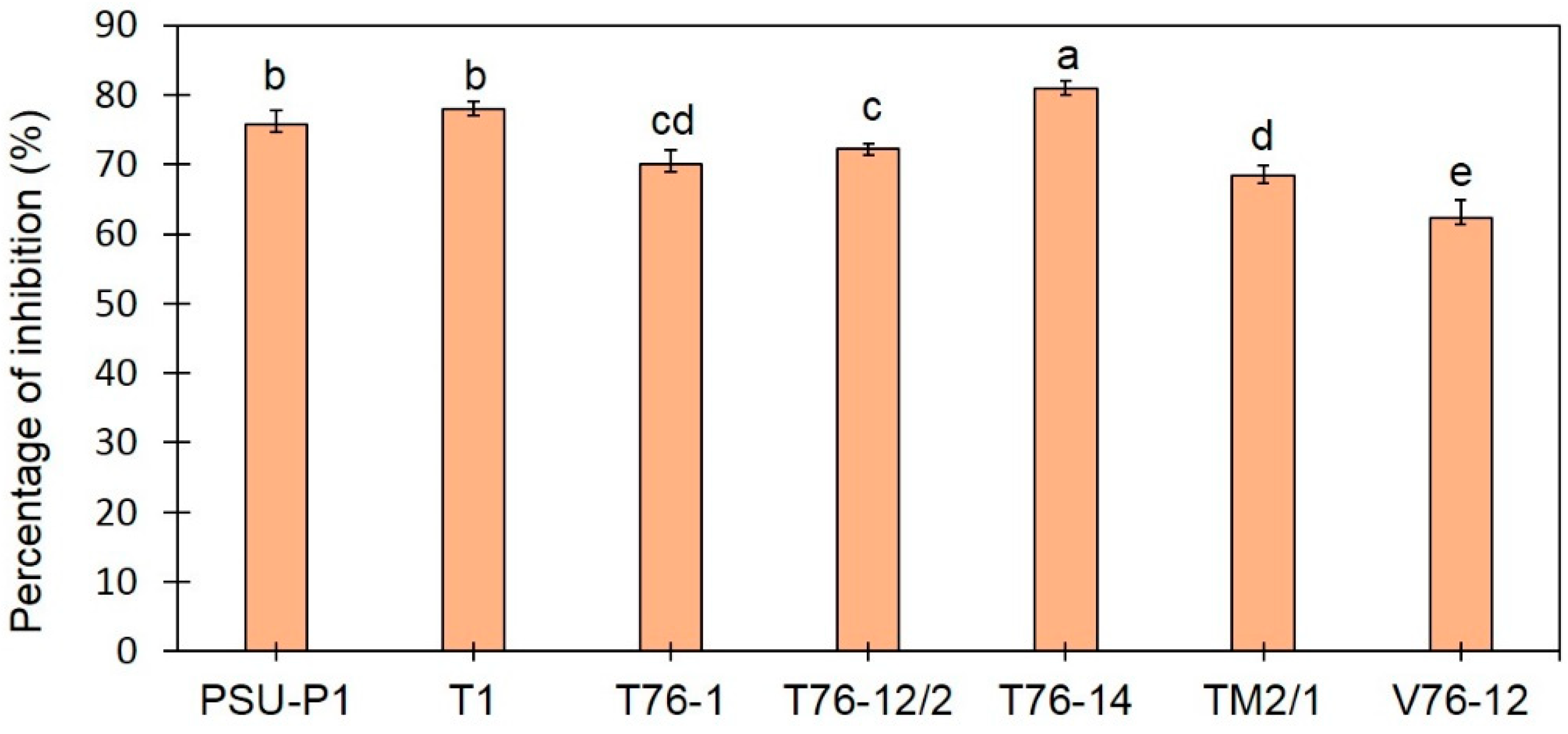
3.1. Trichoderma Species Inhibit Mycelial Growth of F. incarnatum
3.2. Volatiles Emitted by T. asperellum T76-14 Inhibit Mycelial Growth of F. incarnatum
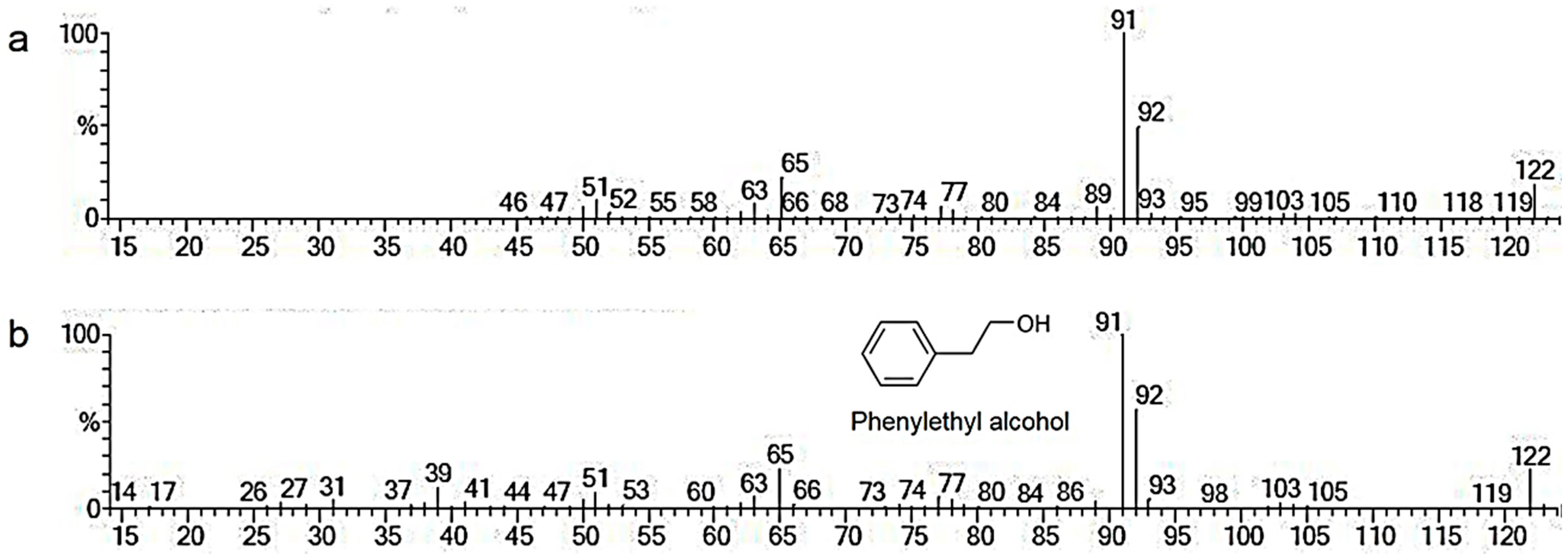
3.3. Identifying Volatile Organic Compounds
3.4. Effect of Commercial Volatile Compounds on Antifungal Activity against F. incarnatum
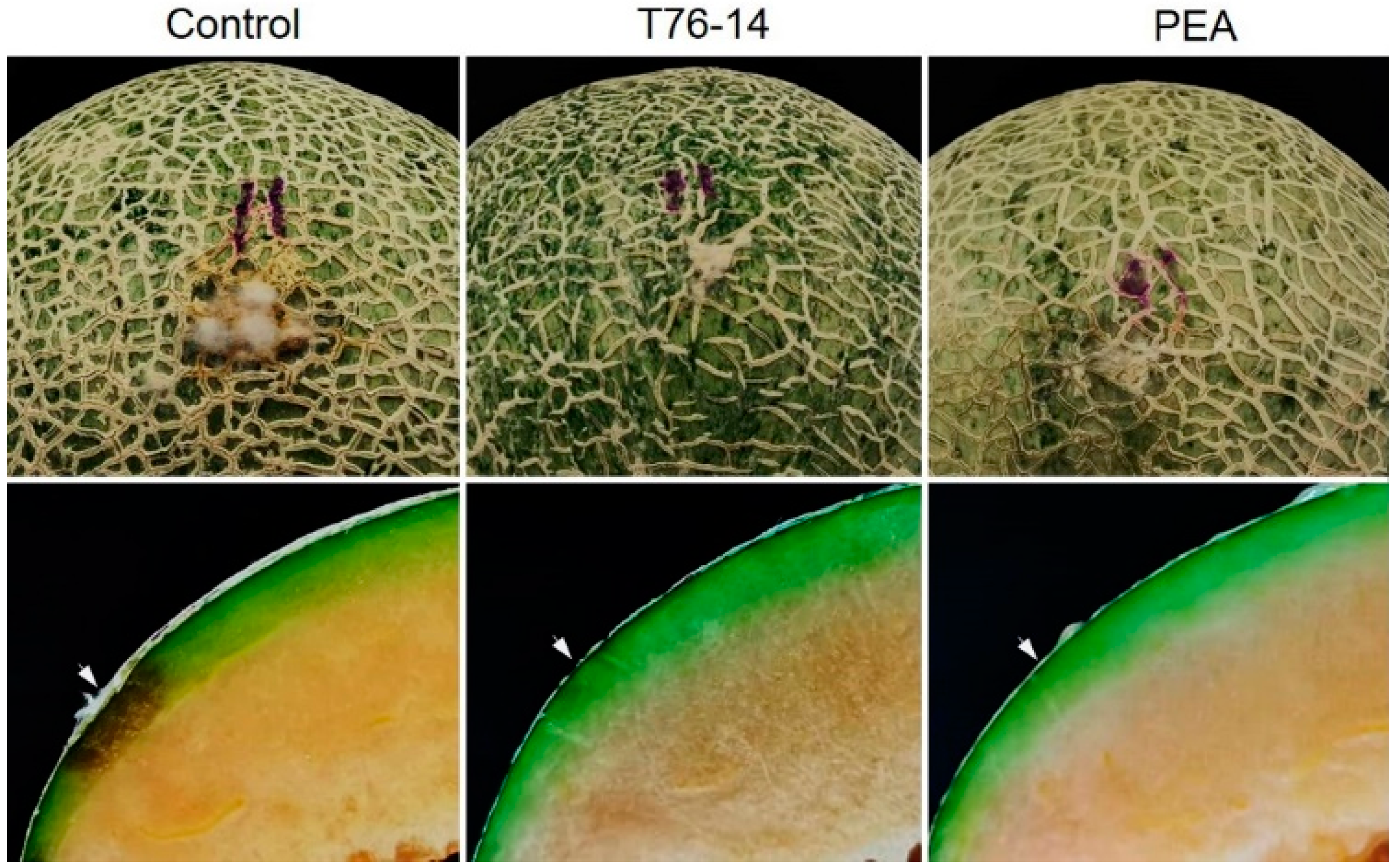
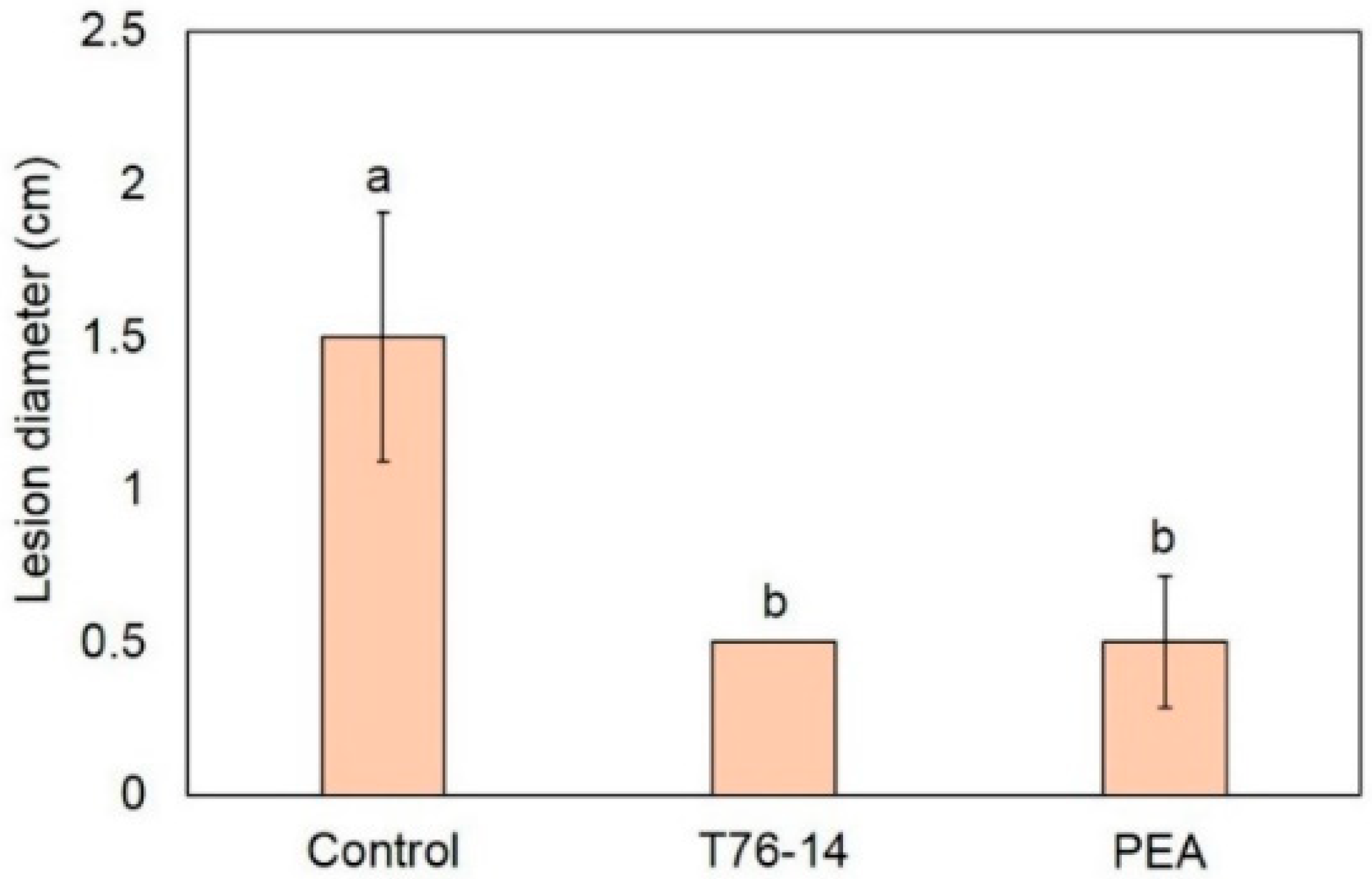
3.5. VOCs of T. asperellum T76-14 Reduced Postharvest Fruit Rot in Muskmelon Fruits
3.6. VOCs of T. asperellum T76-14 Suppressed Fruit Rot in Muskmelons
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wonglom, P.; Sunpapao, A. Fusarium incarnatum is associated with postharvest fruit rot of muskmelon (Cucumis melo). J. Phytopath. 2020, 168, 204–210. [Google Scholar] [CrossRef]
- Yoon, M.Y.; Cha, B.; Kim, J.C. Recent trends in studies on botanical fungicides in agriculture. Plant Pathol. J. 2013, 29, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Fattahi, E.; Mousavi Moghadam, M.; Khanbabaei, R. The effect of fricyclazole on testosterone changes and testicular structure in mice. J. Babol Univ. Med. Sci. 2015, 17, 43–49. [Google Scholar]
- Sunpapao, A.; Chairin, T.; Ito, S. The biocontrol by Streptomyces and Trichoderma of leaf spot disease caused by Curvularia oryzae in oil palm seedlings. Biol. Control. 2018, 123, 36–42. [Google Scholar] [CrossRef]
- Baiyee, B.; Pornsuriya, C.; Ito, S.; Sunapapo, A. Trichoderma spirale T76-1 displays biocontrol activity on lettuce (Lactuca sativa L.) caused by Corynespora cassiicola or Curvularia aeria. Biol. Control. 2019, 129, 195–200. [Google Scholar] [CrossRef]
- Wonglom, P.; Ito, S.; Sunpapao, A. Volatile organic compounds emitted from endophytic fungus Trichoderma asperellum T1 mediate antifungal activity, defense response and promote plant growth in lettuce (Lactuca sativa). Fungal Ecol. 2020, 43, 100867. [Google Scholar] [CrossRef]
- Sunpapao, A. Antagonistic Microorganisms: Current Research and Innovations; Lambert Academic Publishing: Saarbrücken, Germany, 2020; p. 120. [Google Scholar]
- Elad, Y.; David, D.R.; Levi, T.; Kapat, A.; Kirshner, B. Trichoderma harzianum T-39- mechanisms of biocontrol of foliar pathogens. In Modern Fungicides and Antifungal Compounds; Lyr, H., Russell, P.E., Dehne, H.W., Sisler, H.D., Eds.; Intercept: Andover, UK, 1999; pp. 459–467. [Google Scholar]
- Troian, R.F.; Steindorff, A.S.; Ramada, M.H.; Arruda, W.; Ulhoa, C.J. Mycoparasitism studies of Trichoderma harzianum against Sclerotinia sclerotiorum: Evaluation of antagonism and expression of cell wall-degrading enzymes genes. Biotechnol. Lett. 2014, 36, 2095–2101. [Google Scholar] [CrossRef]
- Howell, C.R. Mechanisms employed by Trichoderma species in the biological control of plant diseases: The history and evolution of current concepts. Plant Dis. 2003, 87, 4–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baiyee, B.; Ito, S.; Sunapapo, A. Trichoderma asperellum T1 mediated antifungal activity and induced defense response against leaf spot fungi in lettuce (Lactuca sativa L.). Physiol. Molec. Plant Pathol. 2019, 106, 96–101. [Google Scholar] [CrossRef]
- Phoka, N.; Suwannarach, N.; Lumyong, S.; Ito, S.-I.; Matsui, K.; Arikit, S.; Sunpapao, A. Role of Volatiles from the Endophytic Fungus Trichoderma asperelloides PSU-P1 in Biocontrol Potential and in Promoting the Plant Growth of Arabidopsis thaliana. J. Fungi 2020, 6, 341. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Medina, A.; Alguacil, M.D.M.; Pascual, J.A.; VanWees, S.C. Phytohormone profiles induced by Trichoderma isolates correspond with their biocontrol and plant growth-promoting activity on melon plants. J. Chem. Ecol. 2014, 40, 804–815. [Google Scholar] [CrossRef] [PubMed]
- Brader, G.; Compant, S.; Mitter, B.; Trognitz, F.; Sessitsch, A. Metabolic potential of endophytic bacteria. Curr. Opin. Biotechnol. 2014, 27, 30–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, M.; Chen, L.; Wang, X.W.; Zhang, T.; Zhao, P.B.; Song, X.Y.; Sun, C.Y.; Chen, X.L.; Zhou, B.C.; Zhang, Y.Z. Antimicrobial petaibols from Trichoderma pseudokoningii induce programmed cell death in plant fungal pathogens. Microbiology 2012, 158, 166–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reino, J.L.; Guerrero, R.F.; Hernández-Galán, R.; Collado, I.G. Secondary metabolites from species of the biocontrol agent Trichoderma. Phytochem. Rev. 2007, 7, 89–123. [Google Scholar] [CrossRef]
- Marcías-Rubalcava, M.L.; Hernández-Bautista, B.E.; Oropeza, F.; Duarte, G.; González, M.C.; Glenn, A.E.; Hanlin, R.T.; Anaya, A.L. Allelochemical effects of volatile compounds and organic extracts from Muscodor yucatanensis, a tropical endophytic fungus from Bursera simaruba. J. Chem. Ecol. 2010, 36, 1122–1131. [Google Scholar] [CrossRef]
- Wu, Q.; Sun, R.; Ni, M.; Yu, J.; Li, Y.; Yu, C.; Dou, K.; Ren, J.; Chen, J. Identification of a novel fungus, Trichoderma asperellum GDFS1009, and comprehensive evaluation of its biocontrol efficacy. PLoS ONE 2017, 12, e0179957. [Google Scholar] [CrossRef]
- Loulier, J.; Lefort, F.; Stocki, M.; Asztemborska, M.; Szmigielski, R.; Siwek, K.; Grzywacz, T.; Hsiang, T.; Ślusarski, S.; Oszako, T.; et al. Detection of Fungi and Oomycetes by Volatiles Using E-Nose and SPME-GC/MS Platforms. Molecules 2020, 25, 5749. [Google Scholar] [CrossRef]
- Stracquadanio, C.; Quiles, J.M.; Meca, G.; Cacciola, S.O. Antifungal Activity of Bioactive Metabolites Produced by Trichoderma asperellum and Trichoderma atroviride in Liquid Medium. J. Fungi 2020, 6, 263. [Google Scholar] [CrossRef]
- Tao, L.Y.; Zhang, Y.W.; Li, Y.Q.; Luo, L.P.; Zhang, Z.L.; Chen, J. Antagonistic activity of volatile metabolites from Trichoderma asperellum. Chin. J. Biotechnol. 2020, 36, 1181–1189. [Google Scholar]
- Vinale, F.; Sivasithamparam, K.; Ghisalberti, E.L.; Ruocco, M.; Woo, S.; Lorito, M. Trichoderma secondary metabolites that affect plant metabolism. Nat. Prod. Commun. 2012, 7, 1545–1550. [Google Scholar] [CrossRef] [Green Version]
- Castillo, F.D.H.; Padilla, A.M.B.; Morales, G.G.; Siller, M.C.; Herrera, R.R.; Gonzales, C.N.A.; Reyes, F.C. In vitro antagonistic action of Trichoderma strains against Sclerotinia sclerotiorum and Sclerotium cepivorum. Am. J. Agr. Biol. Sci. 2011, 6, 410–417. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.A.; Begum, M.F.; Alam, M.F. Screening of Trichoderma isolates as a biological control agent against Ceratocystis paradoxa causing pineapple disease of sugarcane. Microbiology 2009, 37, 277–285. [Google Scholar]
- Dennis, C.; Webster, J. Antagonistic properties of species-groups of Trichoderma, II. Production of volatile antibiotic. Trans. Br. Mycol. Soc. 1971, 57, 41–48. [Google Scholar] [CrossRef]
- Arthur, C.L.; Killam, L.M.; Buchholz, K.D.; Pawliszyn, J. Automation and optimization of solid-phase microextraction. Anal. Chem. 1992, 64, 1960–1966. [Google Scholar] [CrossRef]
- Gomez, K.A.; Gomez, A.A. Statistical Procedure for Agricultural Research; John Wiley and Sons: New York, NY, USA, 1984. [Google Scholar]
- Wonglom, P.; Daengsuwan, W.; Ito, S.; Sunpapao, A. Biological control of Sclerotium fruit rot of snake fruit and stem rot of lettuce by Trichoderma sp. T76-12/2 and the mechanisms involved. Physiol. Molec. Plant Pathol. 2019, 107, 1–7. [Google Scholar] [CrossRef]
- Zhu, Y.J.; Zhou, H.T.; Hu, Y.H.; Tang, J.Y.; Su, M.X.; Guo, Y.J.; Chen, Q.X.; Liu, B. Antityrosinase and antimicrobial activities of 2-phenylethanol, 2-phenylacetaldehyde and 2-phenylacetic acid. Food Chem. 2011, 124, 298–302. [Google Scholar] [CrossRef]
- Fraud, S.; Rees, E.L.; Mahenthiralingam, E.; Russell, A.D.; Maillard, J.Y. Aromatic alcohols and their effect on gram-negative bacteria, cocci and mycobacteria. J. Antimicrob. Chemother. 2003, 51, 1435–1436. [Google Scholar] [CrossRef]
- Gao, F.; Daugulis, A.J. Bioproduction of the aroma compound 2-phenylethanol in a solid–liquid two-phase partitioning bioreactor system by Kluyveromyces marxianus. Biotechnol. Bioeng. 2009, 104, 332–339. [Google Scholar] [CrossRef]
- Lester, G. Inhibition of growth, synthesis, and permeability in Neurospora crassa by phenethyl alcohol. J Bacteriol. 1965, 90, 29–37. [Google Scholar] [CrossRef] [Green Version]
- Lucchini, J.J.; Bonnaveiro, N.; Cremieux, A.; Le Goffic, F. Mechanism of bactericidal action of phenethyl alcohol in Escherichia coli. Curr. Microbiol. 1993, 27, 295–300. [Google Scholar] [CrossRef]
- Mo, E.K.; Sung, C.K. Phenylethyl alcohol (PEA) application slows fungal growth and maintains aroma in strawberry. Postharvest Biol. Technol. 2007, 45, 234–239. [Google Scholar] [CrossRef]
- Angel, L.P.L.; Yusof, M.T.; Ismail, I.S.; Ping, B.T.Y.; Azni, I.N.A.M.; Kamarudin, N.H.; Sundram, S. An in vitro study of antifungal activity of Trichoderma virens 7b and profile of its non-polar antifungal components released against Ganoderma boninense. J. Microbiol. 2016, 54, 732–744. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Cheng, Y.; Yang, M.; Liu, Y.; Chen, K.; Long, C.; Deng, X. Mechanisms of action for 2-phenylethylethanol isolated from Kloeckera apiculata in control of Penicillium molds of citrus fruits. BMC Microbiol. 2014, 14, 242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoppacher, N.; Kluger, B.; Zeilinger, S.; Krska, R.; Schuhmacher, R. Identification and profiling of volatile metabolites of the biocontrol fungus Trichoderma atroviride by HS-SPME-GC-MS. J. Microbiol. Methods 2010, 81, 187–193. [Google Scholar] [CrossRef]
- Siddiquee, S.; Cheong, B.E.; Taslima, K.; Kausar, H.; Hasan, M.M. Separation and identification of volatile compounds from liquid culture of Trichoderma hazianum by GC-MS using three different capillary columns. J. Chromatogr. Sci. 2012, 50, 358–367. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Wu, L.; Hao, J.; Luo, L.; Cao, Y.; Li, J. Biofumigation on post-harvest diseases of fruits using a new volatile-producing fungus of Ceratocystis fimbriata. PLoS ONE 2015, 10, e0132009. [Google Scholar] [CrossRef]
- Di Francesco, A.; Di Foggia, M.; Baraldi, E. Aureobasidium pullulans volatile organic compounds as alternative postharvest method to control brown rot of stone fruits. Food Microbiol. 2020, 87, 103395. [Google Scholar] [CrossRef]
- Elsherbiny, E.A.; Amin, B.H.; Aleem, B.; Kingsley, K.L.; Bennett, J.W. Trichoderma volatile organic compounds as a biofumigation tool against late blight pathogen Phytophthora infestans in [ostharvest potato tubers. J. Agric. Food Chem. 2020, 68, 8163–8171. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RT (min) | Volatile Compounds | Formula | % Math | % Area |
|---|---|---|---|---|
| 1.58 | Fluoro(trinitro)methane * | CFN3O6 | 95 | 8.78 |
| 2.29 | Pentan-1-ol | C5H12O | 86.4 | 2.98 |
| 2.31 | 3-Methylpentane | C6H14 | 89.2 | 4.10 |
| 5.66 | 2,4,6-Trimethyloctane | C11H24 | 83.7 | 1.34 |
| 6.28 | 5,7-Dimethylundecane | C13H28 | 84.4 | 1.60 |
| 6.52 | Phenylethyl alcohol | C8H10O | 94.8 | 23.19 |
| 7.45 | Octadec-9-yne | C18H34 | 74.9 | 3.09 |
| 7.65 | 1-Methylideneindene | C10H10 | 92.8 | 1.34 |
| 10.27 | 3-Isopropyl-6,8a-dimethyl-1,2,4,5,8,8a-hexahydroazulene | C15H24 | 85.8 | 1.41 |
| 11.64 | β-Curcumene | C15H24 | 84.4 | 8.49 |
| 11.83 | α-Bisabolene | C15H24 | 83.8 | 1.69 |
| 12.04 | Sesquisabinene B | C15H24 | 83.5 | 1.74 |
| 13.10 | Zingiberenol | C15H26O | 81.8 | 3.11 |
| 13.30 | Cubenol | C15H26O | 83.1 | 6.04 |
| 13.61 | Undeca-3,4-diene-2,10-dione, 5,6,6-trimethyl | C14H22O2 | 75.1 | 1.44 |
| 14.03 | Allyldimethyl(prop-1-ynyl)silane | C8H14Si | 71.4 | 0.77 |
| 15.12 | cis-Z-α-Bisabolene epoxide | C15H24O | 71.3 | 1.73 |
| Commercial Volatiles | Percentage Inhibition (%) a | ||
|---|---|---|---|
| 10−1 | 10−2 | 10−3 | |
| 1-Nonanol | 55.42 ± 13.69 b | 29.71 ± 3.32 b | 19.67 ± 5.69 c |
| 2-Ethylhexanol | 74.29 ± 11.12 a | 29.71 ± 5.01 b | 24.89 ± 9.04 a |
| 2-Methyl-1-butanol | 39.12 ± 8.43 c | 20.88 ± 4.86 d | 16.86 ± 1.20 d |
| 6-Pentyl-2H-pyran-2-one | 21.68 ± 5.25 d | 32.53 ± 14.65 a | 20.08 ± 6.18 b |
| Phenylethyl alcohol | 56.00 ± 5.25 b | 24.00 ± 6.92 c | 12.00 ± 4.00 e |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Intana, W.; Kheawleng, S.; Sunpapao, A. Trichoderma asperellum T76-14 Released Volatile Organic Compounds against Postharvest Fruit Rot in Muskmelons (Cucumis melo) Caused by Fusarium incarnatum. J. Fungi 2021, 7, 46. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7010046
Intana W, Kheawleng S, Sunpapao A. Trichoderma asperellum T76-14 Released Volatile Organic Compounds against Postharvest Fruit Rot in Muskmelons (Cucumis melo) Caused by Fusarium incarnatum. Journal of Fungi. 2021; 7(1):46. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7010046
Chicago/Turabian StyleIntana, Warin, Suchawadee Kheawleng, and Anurag Sunpapao. 2021. "Trichoderma asperellum T76-14 Released Volatile Organic Compounds against Postharvest Fruit Rot in Muskmelons (Cucumis melo) Caused by Fusarium incarnatum" Journal of Fungi 7, no. 1: 46. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7010046